
Abstract
This article presents the archaeobotanical study of Taï, a Neolithic settlement located in Languedoc, southern France. In the western Mediterranean, the Neolithization process occurred during the 6th millennium bc and is supposed to have induced a fundamental change in the subsistence economy, with the development of agriculture and animal husbandry. Unfortunately, the Neolithic farming economy is still poorly known in southern France, mostly from old archaeobotanical studies. At Taï, soil flotation of 127 samples provided the largest dataset available for the Early Neolithic in southern France, which testifies to the changes occurring in cultivated plants and in the subsistence economy during the Neolithic. Sampling also allowed comparisons between the archaeobotanical record inside the cave and from the outside settlement. Naked barley and naked wheat were the most cultivated plants throughout the Neolithic sequence. The contribution of emmer and the probable use of opium poppy during the Early Neolithic should also be highlighted. This encourages us to reconsider the role of glume wheats during the early stages of agriculture in the area. Einkorn was more common in the Late Neolithic, in agreement with results from other sites in the region. Chaff remains were always underrepresented. Remains of weeds and wild fruits were very abundant in the Early Neolithic samples from the cave. Wild plants were probably brought to the site for the tending of domestic animals or by the animals themselves (dung, fodder and/or litter). Livestock was most probably occasionally kept at the site.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
In Mediterranean France, the Neolithic period lasted for more than 3,500 years, from ca. 5800 to ca. 2100 bc. The economy of these societies is generally thought to have been based on growing crops and keeping domesticated stock. However, plant resources and cultivation practices underlying the Neolithic subsistence economy are still poorly known in the area. The first Early Neolithic colonizers of the Impressa culture in ca. 5800–5600 cal bc arrived from Italy, but settled only at a few scattered sites not far from the sea (Guilaine and Manen 2007). The first archaeobotanical results confirmed that these people were already living, at least partly, on cultivation products, especially on hulled wheats (Marinval 2007). Actually, the spread of the Neolithic economy further inland was ensured by the Cardial/Epicardial cultural complex, ca. 5400–4500 cal bc (Perrin et al. 2017). The available archaeobotanical data suggest that the agriculture of that period was mostly based on naked barley and naked wheat (Marinval 1993). However most of this information is available from sites, often in caves, which were excavated during the 1970s and 1980s. These sites were studied according to the methodological standards of the time and rarely involved extensive sampling and fine sieving. Such studies do provide information about the major economic plants present, but make it impossible to investigate cultivation practices and the specific activities related to in situ plant processing. The same is also true for the rest of the Neolithic period. Throughout this period, southern France underwent important cultural, social and techno-economic changes (Guilaine et al. 2007; Cauliez 2011; Binder 2016). It has for example been argued that during the Middle Neolithic (ca. 4500–3500 cal bc) the economy was often focused on itinerant pastoral activities, while cultivation was of secondary importance (Beeching et al. 2000). At present, the available archaeobotanical information is not representative enough to fully reflect the changes and adaptations that may have occurred in the farming economy through the whole of the Neolithic.
The lack of representative archaeobotanical data is also, and above all, attributable to the lack of integrated multidisciplinary research teams and to the relatively small number of extensive excavations with adequate sampling during the last 30 years; this is particularly noticeable when compared to investigations carried out on more recent archaeological periods. Furthermore, whenever extensive excavations and archaeobotanical sampling were carried out (mostly for rescue archaeology), the density of carbonized botanical remains in the sediment was low (Bouby and Figueiral 2014), and this probably did not encourage the development of archaeobotanical investigations.
Cave and rock shelter sites are therefore of primary interest for archaeobotanical research in the area, despite the fact that a restricted number of such sites have been excavated in the last years. Archaeological investigations at Taï (Remoulins, Gard) gave us the opportunity both to thoroughly sample the sediments of the cave for macrobotanical remains and to have a glimpse at the adjoining open air settlement. This paper presents the archaeobotanical data recorded from Taï and attempts to understand how plants were used in a Neolithic site in Mediterranean France. Of particular interest was the identification of the plant resources and farming activities during the Early Neolithic period, which are most difficult to investigate in the region, due to the limited number of sites. After a taphonomic assessment of the assemblages, we also aim to investigate any changes in economic plants and activities that may have occurred from the Early to the Late Neolithic.
The site under study
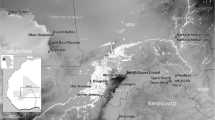
The Taï settlement (Remoulins) is located in the Gard département, about 40 km from the sea and between Avignon and Nîmes (Fig. 1). The site is positioned in one of the numerous valleys deeply carved into the limestone plateau of Garrigues, opening to the plain of Remoulins. The site is at the interface of several ecosystems favourable to human life. It has been excavated for 10 years and thanks to the multidisciplinary research team, it has been possible to understand the sediment dynamics and general topography, to reconstruct the production system of the Neolithic communities and to discuss the functional status of each occupation period.

Location of the site of Taï in southern France
The excavation took place in different sectors of the site: in the cave, in its extension the entrance and in the open-air area in front of the cave (Fig. 2). The different sectors do not offer the same degree of preservation and the different periods are not equally represented there. The history of the site with its different phases has been investigated by field observations, the study of artefacts and 42 radiocarbon dates (Table 1). Five of the radiocarbon dates were directly performed on individual remains of cereal caryopses (Table 2). The earliest human occupations belong to the Early Neolithic (EN) and are dated to 5270–4990 cal bc. These layers are well preserved in the cave (Fig. 3) and in the entrance (about 40 m2) but have been totally eroded in the open-air sector. However, the dwellings probably extended to the open-air area and the cave could have been used for domestic activities and as a refuse area. The Early Neolithic layers (belonging to the Epicardial culture) have been divided into two groups (EN1 and EN2) according to the stratigraphy. The presence of remains linked with all the technological steps indicates a long-lasting stay of the communities at the site. After a period of abandonment, the site was re-occupied during the Middle Neolithic, between 4010 and 3500 cal bc. The Middle Neolithic layers (MN) are badly preserved in the cave, well developed in the entrance and totally eroded in the open-air sector. The available surface (only about 15 m2) is therefore insufficient to understand the status and rhythms of occupation of the site during this period. The stone-working and pottery remains belong to the Chasséen culture. The domestic structures are rare but a high density of burning residues characterizes the deposits. In continuity with the previous occupations, communities of the Late Neolithic (LN—Ferrières culture) settled there between 3575 and 3050 cal bc. The layers mainly extend in the open-air area but are also preserved in the cave and in the entrance. During this period, it is possible to observe the complementary use of the open-air space and the sheltered space. All the sectors include domestic structures (clay and stone architecture, pits, hearths), large amounts of material production and food waste. The habitat seems to have been abandoned around 3000 cal bc, but the site continued to be used for funerary purposes during the Chalcolithic Fontbouisse culture. Indeed, the cave was arranged and used to receive collective burials for more than 300 years. At the beginning of the early Bronze Age, the site was finally abandoned. From a general point of view, one can say that the sediment dynamics are characterized by an alternation of human-derived and natural deposits, and by erosional phases. In the cave, the earliest deposits (EN) are best preserved thanks to the protection by the entrance. The entrance was eroded little by little and the detritus deposits increased in the cave. From the Middle Neolithic onwards, there are fewer signs of human activities in the cave. The unprotected layers outside the cave were eroded. But the Late Neolithic occupations were preserved locally thanks to the severe erosion of the slopes, probably under pressure from human activities, which rapidly covered and preserved the archaeological layers.

a Plan; b photo of the site of Taï indicating the three sectors of the site and the areas excavated in each case

Main stratigraphic profile of the Taï cave with indication of the main phases
Materials and methods
During all the excavation campaigns sediments were sampled to undergo water flotation only when the risk of disturbance was considered low, according to field observations. In total, 127 samples were analysed, representing 1,752 l of sediment.
Samples cover all the occupation phases of the site and come from the interior of the cave, from the entrance and from the settlement outside (Table 1). However, the majority of the samples are related to the two Early Neolithic phases and to the interior of the cave.
Sediments were processed using mesh sizes of 2 and 0.4 mm. The mineral residues, which do not float, were sieved with a single mesh of 0.8 mm. All the fractions were slowly dried and entirely sorted using a stereo microscope. All plant remains were then identified according to their anatomical and taxonomical origins, using scientific reference literature and the comparison collections of modern seeds at the archaeobotany laboratories of ISEM (Montpellier) and TRACES (Toulouse). Plant remains were systematically counted (NPR) and added up by sample regardless of the mesh and residue, distinguishing entire seeds (NER) and fragments (NF). Minimum number of individuals (MNI) was calculated using the formula: MN = NER + ½ NF. Botanical nomenclature follows TAXREF (Gargominy et al. 2016). Ecological characteristics of wild plants were described according to Braun-Blanquet et al. (1952), Ellenberg (1988) and Julve (2015) as regards phytosociology, and according to Pignatti (2005) and Julve (2015) concerning autecological values.
In order to explore the variation in sample composition, a correspondence factor analysis (CFA) was carried out with XLStat software. Raw counts of palaeoecological remains are highly determined by sampling and laboratory methods, especially by sample size (Van der Veen 1992; Birks 2012). Therefore, CFA was performed on percentage data per sample rather than raw counts. Due to their high fragmentation, Cerealia remains were excluded from the reference sums before calculating percentages. In order to have assemblages of at least 30 MNI, samples were grouped according to archaeological structures (hearths, pits), sub-squares and adjoining sub-layers. In order to keep as much diversity as possible, grouping of samples was only undertaken when strictly necessary. If small samples could not be grouped with any other neighbouring small samples in order to reach 30 MNI, they were simply left out of the CFA or processed as passive observations (13 samples). Only taxa recorded in at least 5% of the samples were taken into consideration, rare taxa being regarded mostly as providers of detrimental noise (Van der Veen 1992). Only occasionally, and when individual taxa could not be included due to their low seed number, plants were grouped according to large phytosociological units, such as segetal weeds. Three taxa (Papaver, Linum and Galium aparine) were considered as supplementary (passive) variables because their seeds were strongly concentrated in one sample. Supplementary variables are positioned in the plot but do not enter the computations.
Among our primary goals we wanted to describe the variations in the preservation of plant remains and in taphonomic characteristics of the samples within the site, especially between the different sectors and occupation phases. The condition of the plant remains was evaluated by recording their state of preservation and their fragmentation at the sample level. For each sample we calculated the taxa diversity (minimum number of identified taxa/litre), the global density of plant remains (NPR/litre and MNI/l) and the individual density of cultivated taxa. Regarding glume wheats, the contribution of grain (primary product) and crop processing by-products to the assemblages was evaluated by the calculation of ratios between the number of glume bases and the number of glume wheat grains (Van der Veen 1992). Before calculating the ratios, we converted the number of glume bases into the equivalent number of grains according to the anatomy of each species (one glume per grain for emmer; two glumes per grain for einkorn). For the grains we used the MNI. The number of Middle Neolithic remains was too restricted to allow the calculation of such ratios.
Results
Preservation and distribution of plant remains
A total of 7,590 fruit and seed remains were recovered. Most of the material was preserved by carbonization. Only 11 mineralized seeds were recovered from the cave sector and they were generally poorly preserved, making their identification difficult. Here we will only consider the carbonized plant remains (Table 3).
The calculated MNI was 4,737 and the minimal number of taxa 67. The density of plant remains in the sediments was variable, regardless of the chronology and the spatial origin of the samples (4.73 ± 4.11 NPR/l; min = 0, max = 21.5) (Fig. 4). The diversity of identified taxa per litre of sediment decreased from the cave to the entrance and then to the exterior sector. The lower number of identified taxa/l in the Middle and Late Neolithic samples probably only reflected this spatial trend, as Early Neolithic material was only sampled within the cave and Ferrières mostly outside.

Box-plot diagrams showing the distribution of the archaeobotanical material in the site. Density of plant remains (number of plant remains/l) according to; a chronological phases (EN1 Early Neolithic 1, EN2 Early Neolithic 2, MN Middle Neolithic, LN Fer Late Neolithic Ferrières, Fer/Font Late Neolithic Ferrières/Fontbouisse, LN Font Late Neolithic Fontbouisse); b sectors of the site. Number of taxa/l of sieved sediment according to, c chronological phases; d sectors of the site. The number of samples per group is indicated at the top of each diagram. Statistical differences between groups of samples were determined using non parametric Kruskal–Wallis tests followed by comparisons with the Steel–Dwass–Critchlow–Fligner method (probability values indicated on the diagrams when significant differences were detected)
The quality of preservation of carbonized plant remains was also variable. Seeds of cultivated plants were generally poorly preserved, especially caryopses of cereals, which were highly fragmented and corroded. Corrosion of chaff remains was also noticeable even if not as strongly as in the cereal grains. On the other hand, the preservation of fruits and seeds from wild plants was much better.
Domesticated plants
Cereal grains were to some extent over-represented due to the large quantity of unidentifiable fragments of Cerealia (Fig. 5). Hordeum vulgare var. nudum (naked barley), Triticum aestivum/turgidum (naked wheat), T. turgidum ssp. dicoccon (emmer) and T. monococcum (einkorn) were identified. Barley seems to be the most abundant species but many wheat grains were poorly identified. Most of the grains identified as barley belonged to the naked variety. Less well preserved specimens could not be identified more precisely than as H. vulgare s.l. Among wheat species, naked wheat was the most common. Cereal chaff was much less abundant than grain (3.2% of MNI) and mostly belonged to glume wheats, especially einkorn.

Representation of the main categories of plant remains in the Taï samples
Pulses were weakly represented. Many of the large pulse seeds believed to belong to cultivated plants were fragmented. Lens culinaris (lentil) was identified by at least three seeds and the identification of cf. Pisum sativum (pea) remained uncertain.
It was not possible to assign the seeds of Papaver somniferum (poppy) either to the domesticated form (P. somniferum ssp. somniferum) or its wild ancestor (P. somniferum ssp. setigerum), based on morphological criteria. The wild poppy grows wild in the fields of the western Mediterranean, including the area around Taï (Julve 2016). However, as several seeds have been found concentrated around an Early Neolithic pit (EN1) alone, we consider that they are likely to belong to the cultivated poppy.
Wild plants
A large number of wild plant fruits and seeds were recovered at Taï (Fig. 5). Their MNI represents 34% of the total assemblage and at least 60 wild taxa were recorded. Many wild plant remains are from fruit trees and shrubs typical of open Mediterranean evergreen oak woods (Arbutus unedo, Juniperus communis/oxycedrus, J. phoenicea, Pistacia terebinthus, Quercus sp.). A group of taxa with more mesophilous preferences (Cornus sanguinea, Corylus avellana, Prunus spinosa, Sambucus nigra, S. ebulus, Vitis vinifera) should probably be assigned to the alluvial woods by the river Gardon, which runs less than 1 km from the site. Under a Mediterranean climate, mesophilous trees and shrubs are largely restricted to the heavy soils, with good water retention, of the alluvial plains.
At least 44 wild herbaceous taxa were identified. Most of these plants grow typically in habitats disturbed by human activity (synanthropic) (Fig. 6). Seeds of segetal weeds were identified only sporadically, but some are obviously related to the weed community typical of cereals grown on calcareous soils (Ajuga chamaepitys, Asperula arvensis, Galium aparine ssp. spurium, Sherardia arvensis, Thymelaea passerina). Such soils are predominant on the plateau around the site. Scleranthus annuus and Vicia angustifolia-type, or even Raphanus raphanistrum, are more typical components of cereal weed communities growing on lime-deficient soils. Around the site such conditions can be found on the sandy alluvial soils along the river Gardon.

Detail of the main ecological groups represented by wild herbaceous taxa in the Taï samples
The most abundant wild plant remains were those from ruderals, most particularly from one single species, Chenopodium album. Ruderal weeds are today encountered in gardens, rich arable soils, vineyards and, for most taxa, also on wasteland. Other typical ruderal plants were classified into the group of fallow fields and wasteland. Annual nitrophilous plants (Chenopodium hybridum, Hyoscyamus albus/niger) are common on trampled ground close to human habitat areas. Other taxa (Carthamus lanatus, Saponaria officinalis, Verbena officinalis) are usually to be found in persistent fallow areas such as abandoned arable land, on path margins and in livestock enclosures. Another group of taxa is associated with grasslands, mostly the communities that are common on the thin and rocky soils of the Mediterranean landscapes. However, this group is heterogeneous according to the ecological requirements of plants (some are more mesophilous) and include several taxa that are not identified to species level. Lastly, several species belong to the wet soil communities that are often encountered in the region on occasionally flooded river banks (Cladium mariscus, Eleocharis palustris, Persicaria lapathifolia, Schoenoplectus lacustris s.l.)
Data analysis and variation in sample composition
The correspondence factor analysis (CFA) was performed on a matrix composed of 45 active samples, 13 supplementary samples, 44 active taxa and 3 supplementary taxa (ESM 1). It allowed us to identify variations in sample composition associated with chronological and spatial patterns (Chi2 = 11497.67, df = 1892, p < 0.0001; total inertia = 1.562). The first two axes accounted for 40.03% of the total variance (Fig. 7). Axis 1 (23.8% of total variance) strongly separated the Early Neolithic samples from those of later periods, especially Late Neolithic (Ferrières and Fontbouisse). Most of the Early Neolithic samples clustered together in the negative part of axis 1, associated with wild plants, especially trees and shrubs and some cultivated plant remains (grain and chaff of T. dicoccon, pulses and poppy). All the Middle and Late Neolithic samples were situated in the positive part of axis 1, closely associated with cereal remains. Axis 1 therefore separates the samples dominated by wild plant remains from the samples mostly composed of cereal grain and chaff (ESM 2). Axis 2 (13.23% of total variance) introduced a separation within this last group, between samples from the cave, in the positive part, and those from the entrance and the exterior sector located in the negative part. Chaff remains, chiefly T. monococcum, contributed more importantly to the samples from the cave while those from the entrance and exterior were associated with cereal grain (unidentified cereals, barley, naked wheat and einkorn).

First plot of the correspondance factor analysis on taxa abundance (percentage) in the samples. Three taxa (Papaver, Linum and Galium aparine) are considered as supplementary data. a Plot of the samples according to main archaeological periods; b plot of samples according to the sectors of the site; c plot of the taxa
Even when considering additional axes it was not possible to recognize any structure beyond these three groups: (1) Early Neolithic samples from the cave, (2) Middle and Late Neolithic samples from the entrance and the exterior, (3) Late Neolithic samples from the cave.
Discussion
Taphonomy and composition of the assemblages
Physical and chemical characteristics of plant remains cannot account for the differences of preservation reported in our plant spectra, between taxa, plant organs and sectors of the site. Experiments have shown that cereal grains resist charring better than chaff (Boardman and Jones 1990) and that larger elements, such as cereal grains and pulse seeds, resist better than small seeds, such as those from weeds (Märkle and Rösch 2008). Therefore, the better preservation of wild plant remains than cereal grains at Taï is likely to result from differences in the conditions of their carbonization and/or in the depositional processes.
Preservation conditions seemed better in the cave than in the exterior sector. For example, carbonized remains of fleshy fruits, fruit flesh and occasional fruit flesh still adhering to fruit stones and pips (Juniperus sp., Rubus sp.) were reported only in the samples from the cave. The existence of a gradient in preservation quality from the cave to the exterior sector could account for the parallel progressive decrease in taxa diversity.
Most of the chaff remains at the site come from hulled wheats. We saw here that chaff remains had a different distribution in the samples than cereal grain. The contribution of primary product (grain) and crop processing by-products to the constitution of the assemblages can be studied by the calculation of ratios between the number of glume bases and the number of glume wheat grains (Table 4). In Taï these ratios were clearly less than 1 for the Early and Late Neolithic, suggesting the global predominance of clean grain. Dehusking by-products were present but were of restricted importance in all the archaeobotanical assemblages. There were more however from the Late Neolithic, especially in the cave/entrance area and from einkorn.
The density of cereal grains in the sediment increased through time. This is especially striking as regards einkorn (Fig. 8). The calculation of the Mann–Whitney test indicates a significantly higher density in Late Neolithic than in Early Neolithic samples (U = 648.5, p < 0.0001). A similar pattern was observed for barley (U = 510.5, p < 0.0001) and even naked wheat (U = 870, p = 0.014). The situation is similar when we consider chaff remains instead of grain (Fig. 8).

Density of cereal remains, expressed in MNI, in the sediments of Taï according to chronological phases and sectors of the site as in Table 1. The number of samples for each phase/sector is indicated in brackets. a Overall picture based on cereal grains. Unidentified cereals, Hordeum sp. and Triticum spp.; b detail of the wheat species based on wheat grains; c overall picture based on chaff remains
The diversity of cultivated plants seems less during the Middle and Late Neolithic than in the earlier period, as poppy is no longer present and remains of pulses are very scarce. It is however not clear how significant this observation should be considered, as more Early Neolithic samples were studied and preservation conditions were better in the cave.
Farming activities
Considering the small number of sites studied so far and the lack of regionally representative archaeobotanical data, it becomes evident that the very significant sampling carried out at Taï provides a decisive contribution to the knowledge of agricultural resources during the early Neolithic in southern France. Cultivated plant remains were recorded at all periods and in all sectors. The density of cereal grain remains in the sediment seems to increase with time (Fig. 8). Comparison between Early Neolithic and Late Neolithic samples using a non-parametric Mann–Whitney test shows that the density of caryopses was significantly higher in the Late Neolithic sediments (U = 765, p = 0.002). This could be evidence of either an increasing use of cereals in the excavated areas of the site, or of changes in processing activities implying increased contact with heat and/or fire.
On-site farming activities are nevertheless poorly documented, given that mostly cereal grain was recovered in comparison to chaff. Does this mean that the grain was grown elsewhere and brought to the cave in the form of a cleaned product? The under-representation of chaff is a rather common feature in Neolithic sites and several explanations have been put forward (Jones 2000; Jones and Rowley-Conwy 2007; Bishop et al. 2009; McClatchie et al. 2014): (1) chaff resists high temperatures during carbonization less than grain, (2) cereal processing can be done in bulk after the harvest, restricting the opportunities for chaff to be carbonized compared to household day-to-day processing (this is especially true for free-threshing cereals), (3) cereal processing can be done outside of the dwelling area and (4) chaff can be used for animal fodder or in building material, restricting its chances of being carbonized. Another valuable hypothesis to explain the scarcity of carbonized chaff remains considers that crop processing could have been done on a small scale in Neolithic settlements, without prior parching of spikelets, which would also have reduced the possibilities of carbonization (Antolín 2016).
On the one hand, chaff remains found at Taï belong above all to glume wheats. Chaff remains of free threshing cereals are limited to two rachis segments of barley. Naked wheat is totally absent, although it is the most abundant wheat species when it comes to grain. Therefore, our remains represent mostly rubbish from cleaned products, typical of the last steps of crop processing sequences. This does not mean that arable fields were not close to the site and that the crop processing was not done at the site. We will later on discuss further evidence for local cultivation in the light of the presence of potential arable weeds at the site. There are several reasons why chaff of free threshing cereals may be even more heavily underrepresented than glume wheat chaff. Of course processing of free-threshing cereals may have been done in other parts of the site that have not yet been excavated. Processing of naked cereals is usually done in bulk after threshing and therefore outside of the dwelling places. This reduces considerably the chances of chaff from naked cereals to become carbonized. Furthermore it is even less resistant to carbonization than glume bases of hulled wheats (Boardman and Jones 1990). As a consequence, the rarity of free threshing cereal chaff is a common feature in pre- and protohistoric dry settlements of southern France. In 104 sites ranging from the Late Bronze Age to the end of the Roman times, free threshing cereal chaff was better represented than naked cereal grain in only 17 samples from a total of 199. Most strikingly, 14 of these samples came from waterlogged assemblages, where carbonized naked cereal chaff preserves better than in more aggressive dry sediments (Bouby 2014). Consequently free threshing cereals could easily have been processed close to Taï without leaving any trace in the archaeobotanical record.
On the other hand, dehusking by-products of glume wheats were identified in all periods, but these were always under-represented compared to cereal grain; this is also true for the Late Neolithic when an increase of glume bases is detected. This is maybe more surprising because in non-mechanized agrarian societies still cultivating glume wheats today, dehusking is usually done day-by-day, according to needs related to food preparation. This would increase the chances of finding glume wheat chaff in archaeological sites, as is common in contemporary LBK sites (Kreuz et al. 2005; Kreuz and Marinova 2017; Salavert 2011). But processing in bulk during summer can also be done in areas with dry summers (Hillman 1984), such as Languedoc, and this would reduce the chances of glume wheat chaff to reach the site. In our opinion, the scarcity of chaff cannot be used in this case to argue that crops only arrived as clean products at the site. Unfortunately, no in situ burnt stored material was recovered.
Spatial variability in crop processing activities should especially be considered in the case of Taï, as only a section of the site was investigated. For that matter, Early Neolithic occupations could only be excavated in a small area of the cave when we know that the site extended outside. However, Late Neolithic chaff was also under-represented in the settlement outside the cave.
There is a strong discrepancy between the scarcity of chaff remains and the abundance and diversity of potential weed seeds. It is therefore uncertain that on-site crop processing was in that case a major route of entry of the potential weed seeds in the deposits. In any case, the strong representation of wild taxa from synanthropic communities may be considered as evidence of the existence of cultivation activities around Taï. Most of the seeds were from ruderal plants, but segetal weeds were also recorded for each main period of occupation.
Origin and significance of wild plant remains
The diversity and abundance of wild plant remains raises questions about their origins. As in all Neolithic sites, several pathways and activities could explain the presence of charred seeds of wild plants.
Of course, wild plant fruits and seeds could have been collected for human food. Most, if not all the fruits identified at Taï are known to be edible (Couplan 1983). Several herbaceous seeds could also possibly have been consumed, especially the seeds of Chenopodium album, which make up 61% of the total number of wild herbaceous taxa. Strong hints exist for the consumption of C. album seeds in several European Neolithic sites (Bogaard 2004). One species is of special interest, Carthamus lanatus, which is a ruderal plant rather common on Mediterranean, dry, calcareous soils. While it is quite rare in archaeobotanical assemblages it has been recorded in two other Early Neolithic western Mediterranean sites, la Marmotta, in central Italy (Rottoli and Pessina 2007), and La Draga, in Catalonia, Spain (Antolín 2016). The abundance of the seeds at la Marmotta and the record of wild Carthamus seeds at Pre Pottery and Neolithic sites in eastern Europe and in the Near East led to the hypothesis that wild Carthamus could have been used in the same way as the later domesticated species C. tinctorius, especially as a dye and as an oil plant (Rottoli 2002; Marinova and Riehl 2009).
However, many of the wild plant seeds recorded at Taï are not recognized as edible. Wild plant remains were generally better preserved than cereals and pulses, which shows that they followed different pathways during carbonization and formation processes. Their collection as human food does not seem to be a better explanation for their presence than crop processing activities, to justify the importance and diversity of wild plant remains.
Of course, many other activities could explain the presence of one or other taxa. But they can only partially explain the wild plant record. The fact that wild fruits and herbaceous plants were both strongly recorded following a similar pattern, with a strong representation in the Early Neolithic layers, is consistent with the hypothesis that their representation was the result of one main single process. Only a connection to animal husbandry activities seems likely to justify the carbonization of such abundance of wild plant fruits and seeds. This explanation is supported by data from Neolithic circum-alpine lakeshore settlements where the excellent waterlogged preservation conditions have provided evidence of large quantities of wild plant remains entering the sites via animal dung, fodder or litter (Rasmussen 1993; Kühn et al. 2013). Leaf-foddering has been recognized in several Neolithic caves (the so-called “grottes-bergeries”) in the southern margins of the Alps and Massif Central (Thiébault 2005; Delhon et al. 2008; Martin 2014). Many of the most common wild fruits and seeds in the dung layers of la Grande Rivoire, a rock shelter in Sassenages, Isère, are also abundant at Taï (Chenopodium sp., Cornus sanguinea, Corylus avellana, Galium sp., Rubus sp., Sambucus sp.). Fruits of Mediterranean species such as Arbutus unedo and Pistacia lentiscus were also recorded at Can Sadurni (Begues, Catalonia), in the Middle Neolithic layers when the cave was used for animal stabling, and interpreted as remains of leaf fodder (Antolín and Jacomet 2015). In the Languedoc region, the analysis of goat /sheep dung from Late Bronze Age layers of la Fangade (Sète, Hérault) showed that the animals had eaten leaves of Juniperus phoenicia and seeds of several other plants, for example Chenopodium sp., Pistacia sp. and Rubus sp. This suggests that people brought leafy branches of evergreen Quercus, Juniperus and Pistacia lentiscus to the settlement to be used as fodder and/or litter (Chabal et al. 2010; Bouby 2014).
At Taï the carbonization of seeds and fruits from animal food or litter could have been either intentional, if for example dung was used as fuel or dung accumulations were destroyed by fire, or unintentional, if for example hearths were situated in areas containing plentiful dung or fodder/litter remains. The use of dung as fuel can lead to the charring of many wild seeds (Anderson and Ertug-Yaras 1998). At Taï numerous charcoal fragments were found, testifying to the use of wood as fuel. Ethnographical and archaeological studies show that burning of animal dung was regularly practised at sites where animals were intensely penned (Brochier et al. 1992).
No evidence of an in situ burnt layer of dung has been found in the cave of Taï. In any case, due to the small size of the cave and the grouping of several structures (storage pits, large hearths) in this small area, penning of the animals directly inside the cave is unlikely. They were more probably kept outside, close to the cave. However, sedimentological and micromorphological studies give evidence of the presence of burnt animal dung and coprolites in the sediment matrix of several hearths and ashy layers in the cave (Sordoillet unpublished).
Whether of intentional origin or not, charring of wild seeds and fruits when embedded in a dung and/or sedimentary matrix could explain the good preservation of these remains compared to cereals and pulses. This has also been noticed in Neolithic sites in the Iberian Pyrenees (Antolín et al. 2017). The collection of leaf-fodder or litter could also explain the presence of entire fruits in the sediments. According to the taxon and the cutting season, branches can carry leaves and fruits at the same time. For several of the taxa identified at Taï the fruits stay for a long time on the trees, which would favour their collection when leafy branches were cut. This is especially true for Juniperus cones, Arbutus berries or even Pistacia fruits. Furthermore, some deciduous taxa, such as Cornus sanguinea, Prunus spinosa, Rubus spp. and Sambucus spp., can keep their fruits, and even their leaves, for some time in autumn and early winter.
During historical times, evergreen trees were very often cut to provide litter for the cattle in the Mediterranean area in general (Moriceau 1999). Even juniper, which is well represented at Taï, can be collected and used as leaf fodder. Examples from North Africa show that leaf fodder can be collected from Juniperus phoenicia and J. oxycedrus to be given to goats and occasionally to sheep. J. communis is less palatable but can nevertheless also be used for goats (Le Houérou 1980). In this area, J. phoenicia, J. oxycedrus and oaks are conspicuously used as animal fodder (Bourbouze and Donadieu 1987).
Although fruit remains most probably represent remains of leaf fodder collected by people to feed their animals at the site or for their litter, they could also have been directly consumed by the animals around the settlement. Grazing and/or browsing seem, on the other hand, the most likely explanation for the presence of wild herbaceous plant seeds in the site, as the droppings of domestic animals would sooner or later be in contact with fire. According to the taxa recognized in our study, animals would have grazed or browsed in different areas: typical Mediterranean dried grasslands, wet land along the river Gardon and above all in ruderal places around the site, in fallows and harvested fields. However, these same areas could also have been used for the collection of fodder. Before the mechanization of agriculture, fodder was still regularly collected in the Pyrenees and in Languedoc along paths and from animal enclosures (Chevalier 1956; Durand-Tullou 1972).
Seasonality of agro-pastoral activities and Early Neolithic human occupation of the site
If we accept the hypothesis that the Early Neolithic inhabitants of Taï cultivated their own crops, this implies that they occupied the site at various seasons of the year. People were there during summer for the harvest, and probably also during the autumn for the sowing of winter cereals, which fits with the identification of autumn-sown segetal weeds. The numerous ruderal spring-sown weeds indicate tilling activities in spring. Some plants could have been sown in spring, especially opium poppy or pulses. Growing of ruderal spring weeds could also have been favoured by the hoeing and weeding at that season of cereal fields sown during the previous autumn.
The presence of several fruits that stay for some time on the trees after summer suggests that pastoral activities took place around the site in autumn or even early winter. Leaf fodder was probably cut and/or eaten by animals during the cold season. Recent traditional pastoral practices from Greece and Morocco included the collection of leaf fodder principally in late summer or autumn to feed the animals in winter, because needs for fodder are less pressing during spring, cutting of branches is less detrimental to trees in autumn, already fairly dried leaves store better, and young leaves can cause digestive problems to animals (Bourbouze and Donadieu 1987; Forbes 1998; Halstead 1998).
If we consider that many of the ruderal weeds probably come from fallows and harvested fields, this would point to browsing by the animals around the site at the end of summer or during the autumn.
In the end, the archaeobotanical results let us suppose that the Early Neolithic farmers were probably occupying the site all through the year. This is in agreement with many other observations made at the site (Manen et al. 2004, forthcoming). The multi-disciplinary studies do not show any crafting or economical specialization, but on the contrary, a wide range of activities comparable to those observed in permanent open air settlements. Storage pits give evidence of long-term storage in the site. The faunal remains show that the subsistence economy was strongly based on domestic animals. The entire sequence of activities, from slaughtering of the animals to the consumption of meat, took place at the site (Vigne et al., unpublished).
Chronological changes in cultivated plants
The range of cultivated plants identified at Taï is largely similar throughout the full occupation of the site, at least as regards cereal species. The CFA however allows detection of some changes in the proportions of the main species which, for a large part, mirror wider regional economic patterns.
In the Early Neolithic samples, the significant role of naked wheat and naked barley is in total agreement with what is already known of the Cardial/Epicardial agriculture in southern France (Erroux and Courtin 1974; Marinval 1993). On the other hand, the substantial contribution of emmer and the record of einkorn at Taï are more unexpected, as hulled wheats were traditionally regarded as having been of minor importance and restricted to Epicardial sites (Marinval 1993). We should however bear in mind that plant macrofossil data on the Early Neolithic are still scarce in southern France and largely provided by old studies. As a result, the traditional picture of early agriculture is probably not very accurate and the role of hulled wheats may have been underrated. It has recently been shown that hulled wheats were of primary importance at Impressa culture sites, in the very early Neolithic of Mediterranean France, in agreement with the Italian tradition, where Impressa colonists came from (Marinval 2007). For the moment, the agriculture of the Cardial is better known in Spain than in France. From a general point of view, the main cereals in Spain were naked wheat and naked barley, while hulled wheats and hulled barley were locally significant, as for example at the earliest Neolithic sites of the Valencia area, in the Soria area, high Ebro basin, or on the central and southern Catalan coast (Zapata et al. 2004; Antolín et al. 2015; Antolín 2016; Pérez-Jordà et al. 2017). Most likely the spatial variability was comparable in southern France. Hulled wheats probably did not disappear after the Impressa phase, but their role became secondary and more variable, according to patterns that cannot be identified today.
In the Early Neolithic samples from Taï, together with cereal remains, were also found seeds of poppy and pulses. The probable presence of cultivated poppy in our site is of special interest because it had never been identified in southern France as early as the Early Neolithic. This should not however be regarded as a surprise. In fact, the western Mediterranean area is home to Papaver somniferum ssp. setigerum, the wild progenitor of opium poppy, and is the probable zone of its domestication (Bakels 1982). In actual fact, P. somniferum ssp. setigerum/somniferum is well known from several Early Neolithic sites in the area. In Spain, it is reported at least as early as 5300 cal bc, especially at the Cueva de los Murciélagos de Zuheros (Córdoba Province, Andalucia), at Meseta sites in central Spain and at la Draga, in Catalonia (Peña-Chocarro 1999; Antolín 2016; Pérez-Jordà et al. 2017). The presence of poppy in central Italy, at la Marmotta, could even be more ancient (Rottoli and Pessina 2007). From cal 5200 bc onwards, poppy became common at many sites in the Linearbandkeramik area (Salavert 2011), which probably indicates very early contacts and exchanges between the Cardial/Epicardial and Linearbandkeramik areas. These exchanges could have occurred through the Rhône valley, major North–South circulation axis of western Europe, running less than 15 km away from Taï.
The scarcity of seeds of pulses at Taï and the species identified (Lens culinaris and cf. Pisum sativum) are in good agreement with the general situation in western Europe (Antolín 2016). In the western Mediterranean, remains of pulses are only numerous and diversified at some specific sites, such as Sammardenchia-Cueis in northern Italy (Rottoli and Pessina 2007), or maybe in southern Spain (Zapata et al. 2004).
As expected, the limited area excavated from the Middle Neolithic at Tai provided only restricted information on cultivated plants. It is therefore unrealistic to expect to identify any differences between the two Middle Neolithic phases. Cultivated plant remains are still dominated by naked barley and naked wheat grains, together with a lower proportion of einkorn, represented by caryopses and glume bases. This is representative of the wider situation in southern France, where naked barley is of special importance (Marinval 1991; Martin et al. 2016). In France more generally, naked cereals are especially favoured during the first half of the Chasséen culture (4400–4000 cal bc), while the role of glume wheats increased during the second part of the period (Martin et al. 2016). At Taï this increase can be perceived only in the Late Neolithic samples. The growing importance of einkorn is actually the most prominent feature for this period. Again for the Late Neolithic, the results from Taï largely mirror the more global situation in southern France, with a clear prevalence of cereal remains over other cultivated plants. Naked barley, naked wheat, emmer and einkorn are often associated in the sites, in variable proportions. Naked barley, however, is probably the most common cereal in the region. The cultivation of this cereal was a lasting and strong tradition in the Neolithic of southern France. For the moment, there is no evidence in the area of a change towards hulled barley, which took place later, during the Bronze Age, in contrast to what seems to happen in north-eastern Spain at the same period (Antolín et al. 2015). The prevalence of einkorn over emmer has already been reported in certain Middle Neolithic sites in south-eastern France, but it seems more widespread during the Late Neolithic. It has been mentioned at various Late Neolithic sites, but without a clear spatial pattern and the reasons for the choice of einkorn over emmer remain unclear (Bouby et al. 2017).
Conclusions
The site of Taï offers one of the most important Neolithic archaeobotanical datasets in Mediterranean France, especially when we consider the Early Neolithic period. Even if a large part of the information deals with this period, the site also provided the opportunity to investigate the changes in economic plants occurring until the Late Neolithic. For this last period, it was also possible to compare the results obtained from inside the cave and from the area outside it, which helped us to better understand the role of taphonomy.
The by-products of crop processing were underrepresented in comparison to grain in all periods. Nevertheless, the evidence from weeds indicates that agriculture was probably practised close to the site. In the Early Neolithic samples a strong taxonomic diversity and a large number of remains of wild herbaceous plants and wild fruits was recorded. We believe that most of these wild plants were brought to the settlement by people for the tending of livestock (leaf fodder, litter) or by the animals themselves (dung). Therefore, animals were most probably penned at the site, maybe near the cave entrance. Considering the list of identified taxa we can argue that the animals were fed at the site during the summer and autumn/winter. All things considered, the site was certainly occupied permanently throughout the year during the Early Neolithic.
The range of cultivated plants stayed largely the same from the Early to the Late Neolithic. The substantial contributions of emmer as well as the presence of einkorn and probably of opium poppy during the Early Neolithic are especially noteworthy. This raises new questions about the role of glume wheats in the region. Their contribution was probably variable in time and space while plant cultivation was dominated by naked cereals. Data is limited for the Middle Neolithic, but cultivated plants were still dominated by naked wheat and naked barley. In the Late Neolithic, the density of cereal remains in the sediments was higher, whereas the proportion of einkorn increased sharply, in agreement with what has been observed in other Late Neolithic sites in the region. The example of Taï shows the good potential of regional cave sites for archaeobotanical investigations, provided an extensive and adapted sampling procedure is applied. It shows the better preservation of carbonized plant remains in cave sediments than in the exterior adjoining settlement.
The Early Neolithic assemblage from the cave was especially rich, most probably due to the bringing in of plants in connection with livestock, even if other activities were also concerned. However, due to the restricted area available for excavation in the cave and to the impossibility of investigating the Early Neolithic levels in the outside settlement, it is difficult to understand in detail the activities carried out at the site.
References
Anderson S, Ertug-Yaras F (1998) Fuel fodder and faeces: an ethnographic and botanical study of dung fuel use in central Anatolia. Environ Archaeol 1:99–109
Antolín F (2016) Local, intensive and diverse? Early farmers and plant economy in the North-East of the Iberian Peninsula (5500–2300 cal bc). (Advances in Archaeobotany 2). Barkhuis Publishing, Groningen
Antolín F, Jacomet S (2015) Wild fruit use among early farmers in the Neolithic (5400–2300 cal bc) in the north-east of the Iberian Peninsula: an intensive practice? Veget Hist Archaeobot 24:19–33
Antolín F, Jacomet S, Buxó R (2015) The hard knock life. Archaeobotanical data on farming practices during the Neolithic (5400–2300 cal bc) in the NE of the Iberian Peninsula. J Archaeol Sci 61:90–104
Antolín F, Navarrete V, Saña M, Viñerta Á, Gassiot E (2017) Herders in the mountains and farmers in the plains? A comparative evaluation of the archaeobiological record from Neolithic sites in the eastern Iberian Pyrenees and the southern lower lands. Quat Int. https://doi.org/10.1016/j.quaint.2017.05.056
Bakels C (1982) Der Mohn, die Linearbandkeramik und das westliche Mittelmeergebiet. Archäologisches Korrespondenzblatt 12:11–13
Beeching A, Berger J-F, Brochier J-L, Ferber F, Helmer D, Sidi Maamar H (2000) Chasséens: agriculteurs ou éleveurs, sédentaires ou nomades ? Quels types de milieux, d’économies et de sociétés ? In: Leduc M, Valdeyron N, Vaquer J (eds) Sociétés et espaces, Rencontres méridionales de Préhistoire récente, Troisième session, Toulouse, 6–7 novembre 1998. Archives d’Ecologie Préhistorique, Toulouse, pp 59–79
Binder D (2016) Le Chasséen et ses « cultures sœurs »: apports du colloque de 2014. In: Perrin T, Chambon P, Gibaja Bao JF, Goude G (eds) Le Chasséen, des Chasséens… Retour sur une culture nationale et ses parallèles, Sepulcres de fossa, Cortaillod, Lagozza. Actes du colloque international tenu à Paris (France) du 18 au 20 novembre 2014. Archives d’Ecologie Préhistorique, Toulouse, pp 537–44
Birks HJB (2012) Overview of numerical methods in palaeolimnology. In: Birks HJB, Lotter AF, Juggins S, Smol JP (eds) Tracking environmental change using lake sediments. Springer, Dordrecht, pp 19–92
Bishop RR, Church MJ, Rowley-Conwy PA (2009) Cereals, fruits and nuts in the Scottish Neolithic. Proc Soc Antiq Scotl 139:47–103
Boardman S, Jones G (1990) Experiments on the effects of charring on cereal plant components. J Archaeol Sci 17:1–11
Bogaard A (2004) Neolithic farming in central Europe. Routledge, London
Bouby L (2014) L’agriculture dans le bassin du Rhône du Bronze final à l’Antiquité. Agrobiodiversité, économie, cultures. Archives d’Ecologie Préhistorique, Toulouse
Bouby L, Figueiral I (2014) Les ressources végétales du Néolithique ancien nîmois: Mas de Vignoles X et Mas Neuf. In: Perrin T, Manen C, Séjalon P (eds) Le Néolithique ancien de la plaine de Nîmes (Gard, France). Archives d’Ecologie Préhistorique, Toulouse, pp 339–343
Bouby L, Rovira N, Rousselet O (2017) Ressources agricoles dans les sites de plein air du Néolithique final des Grands causses et de la vallée de l’Hérault: données carpologiques. In: Rovira N, Bouby L, Bouchette A, Ruas MP (eds) Plantes, produits et pratiques: diffusion et adoption de la nouveauté dans les sociétés préindustrielles. ADALR, Lattes, pp 15–30 (Monographies d’Archéologie Méditerranéenne HS 8)
Bourbouze A, Donadieu R (1987) L’élevage sur parcours en régions méditerranéennes. Options Méditerranéennes. CIHEAM/IAM, Montpellier
Braun-Blanquet J, Roussine N, Negre R (1952) Les groupements végétaux de la France Méditerranéenne. CNRS, Paris
Brochier JE, Villa P, Giacomarra M (1992) Shepherds and sediments. Geo-ethnoarchaeology of pastoral sites. J Anthropol Archaeol 11:47–102
Cauliez J (2011) Restitution des aires culturelles au Néolithique final dans le sud-est de la France. Dynamiques de formation et d’évolution des styles céramiques. Gallia Préhistoire 53:85–202
Chabal L, Bouby L, Figueiral I, Catanzano J, Leroy F, Guibal F, Greck S (2010) Vivre sur un littoral lagunaire au Bronze final: milieu et activités humaines d’après les macrorestes végétaux du site palafittique de La Fangade, étang de Thau (Sète, Hérault). In: Delhon C, Théry-Parisot I, Thiébault S (eds) Des hommes et des plantes: exploitation du milieu et gestion des ressources végétales de la Préhistoire à nos jours. APDCA, Antibes, pp 235–252
Chevalier M (1956) La vie humaine dans les Pyrénées ariégeoises. Génin, Paris
Couplan F (1983) Le régal végétal. Plantes sauvages comestibles, vol 1. Equilibres Aujourd’hui, Flers
Delhon C, Martin L, Argant J, Thiébault S (2008) Shepherds and plants in the Alps: multi-proxy archaeobotanical analysis of Neolithic dung from ‘‘La Grande Rivoire’’ (Isère, France). J Archaeol Sci 35:2,937–2,952
Durand-Tullou A (1972) Rôle des végétaux dans la vie de l’homme au temps de la civilisation traditionnelle (Étude ethnobotanique sur le Causse de Blandas, Gard). Journal d’Agriculture Tropicale et de Botanique Appliquée 19:222–248
Ellenberg H (1988) Vegetation ecology of central Europe. Cambridge University Press, Cambridge
Erroux J, Courtin J (1974) Aperçu sur l’agriculture préhistorique dans le sud-est de la France. Bull Soc Languedoc Geogr 8:325–336
Forbes H (1998) European agriculture viewed bottom-side upwards: fodder and forage provision in a traditional Greek community. Environ Archaeol 1:19–34
Gargominy O, Tercerie S, Régnier C et al (2016) TAXREF v10.0, référentiel taxonomique pour la France: méthodologie, mise en oeuvre et diffusion. Rapport SPN 2016-101. Muséum national d’Histoire naturelle, Paris
Guilaine J, Manen C (2007) From Mesolithic to Early Neolithic in the western Mediterranean. In: Whittle A, Cummings V (eds) Going over: the Mesolithic-Neolithic transition in north-west Europe. British Academy, London, pp 21–51
Guilaine J, Manen C, Vigne J-D (2007) Pont de Roque-Haute (Portiragnes, Hérault). Nouveaux regards sur la néolithisation de la France méditerranéenne. Archives d’Écologie préhistorique, Toulouse
Halstead P (1998) Ask the fellow who lop the hay: Leaf-fodder in the mountains of northwest Greece. Rural History 9:211–234
Hillman G (1984) Interpretation of archaeological plant remains: the application of ethnographic models from Turkey. In: van Zeist W, Casparie WA (eds) Plants and ancient man, studies in palaeoethnobotany. Balkema, Rotterdam, pp 1–41
Jones G (2000) Evaluating the importance of cultivation and collecting in Neolithic Britain. In: Fairbairn AS (ed) Plants in Neolithic Britain and beyond. Oxbow, Oxford, pp 79–80
Jones G, Rowley-Conwy PA (2007) On the importance of cereal cultivation in the British Neolithic. In: Colledge S, Conolly J (eds) The origins and spread of domestic plants in southwest Asia and Europe. Left Coast Press, Walnut Creek, pp 391–419
Julve P (2015) Baseveg. Répertoire synonymique des groupements végétaux de France. Version: 16 février 2015. http://perso.wanadoo.fr/philippe.julve/catminat.htm. Accessed Feb 2017
Julve P (ed) (2016) chorodep. Listes départementales des plantes de France. http://www.tela-botanica.org/page:liste_projets?id_projet=9. Accessed Feb 2017
Kreuz A, Marinova E (2017) Archaeobotanical evidence of crop growing and diet within the areas of the Karanovo and the Linear Pottery Cultures: a quantitative and qualitative approach. Veget Hist Archaeobot 26:639–657
Kreuz A, Marinova E, Schäfer E, Wiethold J (2005) A comparison of early Neolithic crop and weed assemblages from the Linearbandkeramik and the Bulgarian Neolithic cultures: differences and similarities. Veget Hist Archaeobot 14:237–258
Kühn M, Maier U, Herbig C, Ismail-Meyer C, Le Bailly M, Wick L (2013) Methods for the examination of cattle, sheep and goat dung in prehistoric wetland settlements with examples of the sites Alleshausen-Täschenwiesen and Alleshausen-Grundwiesen (around cal 2900 bc) at Lake Federsee, south-west Germany. J Environ Archaeol 18:43–57
Le Houérou HN (1980) Fourrages ligneux en Afrique du Nord. In: Le Houérou HN (ed) Les fourrages ligneux en Afrique. Etat actuel des connaissances. Centre International pour l’Elevage en Afrique, Addis Abeba, pp 57–84
Manen C, Bouby L, Carrere I et al (2004) Nouvelles données sur le Néolithique ancien gardois: résultats des campagnes de fouilles 2001–2002 de la grotte du Taï (Remoulins). In: Dartevelle H (ed) 5èmes rencontres méridionales de préhistoire récente. Auvergne et Midi, actualité de la recherche. Actes de la cinquième session, Clermont-Ferrand (Puy-de-Dôme), 8 et 9 novembre 2002. Préhistoire du Sud-Ouest, Cressensac, pp 321–336 (Préhistoire du Sud-Ouest, Supplément 9)
Marinova E, Riehl S (2009) Carthamus species in the ancient Near East and south-eastern Europe: archaeobotanical evidence for their distribution and use as a source of oil. Veget Hist Archaeobot 18:341–349
Marinval P (1991) Le chasséen: problèmes d’identité, l’approche carpologique. In: Beeching A, Binder D, Blanchet JC et al (eds) Identité du chasséen, actes du colloque international de Nemours (17–19 mai 1989). (Mémoires du musée de Préhistoire d’Île de France 4). APRAIF, Nemours, pp 379–390
Marinval P (1993) Approche carpologique de la néolithisation du sud de la France. In: Anderson PC (ed) Préhistoire de l’agriculture: nouvelles approches expérimentales et ethnographiques. Monographies du CRA 6. CNRS, Paris, pp 256–263
Marinval P (2007) Premières données sur l’économie végétale. In: Guilaine J, Manen C, Vigne JD (eds) Pont de Roque Haute. Nouveaux regards sur la néolithisation de la France méditerranéenne. Archives d’Ecologie Préhistorique, Toulouse, pp 215–217
Märkle T, Rösch M (2008) Experiments on the effects of carbonization on some cultivated plant seeds. Veget Hist Archaeobot 17:257–263
Martin L (2014) Premiers paysans des Alpes. Alimentation végétale et agriculture au Néolithique. Presses Universitaires de Rennes, Rennes
Martin L, Bouby L, Marinval P et al (2016) L’exploitation des ressources végétales durant le Chasséen: un bilan des données carpologiques en France entre 4400 et 3500 avant notre ère. In: Perrin T, Chambon P, Gibaja Bao JF, Goude G (eds) Le Chasséen, des Chasséens… Retour sur une culture nationale et ses parallèles: Sepulcres de fossa, Cortaillod, Lagozza. Actes du colloque international tenu à Paris (France) du 18 au 20 novembre 2014. Archives d’Ecologie Préhistorique, Toulouse, pp 259–272
McClatchie M, Bogaard A, Colledge S, Whitehouse NJ, Schulting RJ, Barratt P, Mc Laughlin TR (2014) Neolithic farming in north-western Europe: Archaeobotanical evidence from Ireland. J Archaeol Sci 51:206–215
Moriceau JM (1999) L’élevage sous l’Ancien Régime (XVIe-XVIIIe siècles). Sedes, Paris
Peña-Chocarro L (1999) Prehistoric agriculture in Spain: the application of ethnographic models. BAR International Series 818. Archaeopress, Oxford
Pérez-Jordá G, Peña-Chocarro L, Morales Mateos J, Zapata L (2017) Chap. 7: Evidence for early crop management practices in the Western Mediterranean: latest data, new developments and future perspectives. In: García Puchol O, Salazar García DC (eds) Times of Neolithic transition along the Western Mediterranean. Springer, New York, pp 171–197
Perrin T, Manen C, Valdeyron N, Guilaine J (2017) Beyond the sea… The Neolithic transition in the southwest of France. Quat Int. https://doi.org/10.1016/j.quaint.2017.05.027
Pignatti S (2005) Valori di bioindicazione delle piante vascolari della flora d’Italia. Braun Blanquetia 39:3–97
Rasmussen P (1993) Analysis of sheep/goat faeces from Egolzwil 3, Switzerland: evidence for branch and twig foddering of livestock in the Neolithic. J Archaeol Sci 20:479–502
Rottoli M (2002) Zafferanone selvatico (Carthamus lanatus) e cardo della Madonna (Silybum marianum), piante raccolte o coltivate nel Neolitico antico a “La Marmotta”? Bullettino di Paletnologia Italiana (Roma) 2000–2001 91–92:47–61
Rottoli M, Pessina A (2007) Neolithic agriculture in Italy: an update of archaeobotanical data. In: Colledge S, Conolly J (eds) The origins and spread of domestic plants in Southwest Asia and Europe. Left Coast Press, Walnut Creek, pp 141–153
Salavert A (2011) Plant economy of the first farmers of central Belgium (Linearbandkeramik, 5200–5000 bc). Veget Hist Archaeobot 20:321–332
Thiébault S (2005) L’apport du fourrage d’arbre dans l’élevage depuis le Néolithique. Anthropozoologica 40:95–108
Van der Veen M (1992) Crop husbandry regimes. An archaeobotanical study of farming in northern England 1000 bc–ad 500. (Sheffield Archaeological Monographs 3). University of Sheffield, Sheffield
Zapata L, Peña-Chocarro L, Pérez-Jordá G, Stika HP (2004) Early Neolithic agriculture in the Iberian Peninsula. J World Prehist 18:283–325
Acknowledgements
This study was supported by Agence Nationale de la Recherche, project PROCOME, CULT program (ANR-13-CULT-0001-01). We would like to thank Thierry Pastor (ISEM, Université Montpellier), who carried out a large part of flotation and sieving of the Taï sediments, and Isabel Figueiral (ISEM, Inrap) for her correction of the English text. We thank Vincent Bonhomme (ISEM, CNRS) for his help with the preparation of the figures and Dominique Sordoillet (Chrono-Environnemet, Inrap) for sharing unpublished sedimentological and micromorphological data. Many thanks are also due to two anonymous reviewers and to the associate and copy editors for their useful comments and help in improving the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by F. Antolín.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Bouby, L., Durand, F., Rousselet, O. et al. Early farming economy in Mediterranean France: fruit and seed remains from the Early to Late Neolithic levels of the site of Taï (ca 5300–3500 cal bc). Veget Hist Archaeobot 28, 17–34 (2019). https://doi.org/10.1007/s00334-018-0683-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-018-0683-x