
Abstract
Today, the East European forest-steppe is an agricultural landscape with very few remains of its former natural vegetation. The history of the transformation from natural vegetation to a human-made landscape in the area of Sudzha (Kursk region, Russia) is studied here. We compare the off-site pollen record Sudzha with three on-site pollen records obtained from the archaeological site Kurilovka-2. The sediment core Sudzha covering the last 2,500 years was taken from an oxbow lake in an area with archaeological sites of the early Slavonic period (3rd–8th centuries ce). The Sudzha pollen record indicates dominance of broadleaf forests and meadow steppes in the area from 2,500 to 200 cal year bp with two major settlement phases one between ~ 2,000 and 1,600 cal year bp (~ 50 bce to 350 ce) and the other between 1,100 and 600 cal year bp (850 and 1350 ce) followed by a total deforestation and transformation to an agricultural landscape over the last 200–300 years. Similar changes in the last 300–400 years are indicated by the three on-site pollen records. It is noteworthy, however, that the record Sudzha does not provide an intensive signal of human impact during the 5th–8th centuries ce. This points to a quite restricted spatial influence of the Early Slavonic settlements on the vegetation, leading to a relatively low contribution of palynological anthropogenic indicators to the regional pollen rain signal.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
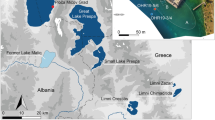
The forest-steppe zone of the East-European plain stretches several hundred kilometres from NW to SE (Fig. 1a). Based on climatic conditions, its natural vegetation should be meadow steppe with patches of broadleaf trees (Bohn et al. 2003). However, this huge territory covered largely by chernozem soils was transformed to an agricultural landscape over previous centuries. Today, it is difficult to find any areas covered with natural vegetation between the crop fields. The exact timing of this transformation is unclear due to difficulties finding appropriate pollen archives, so that information on the vegetation history of this large zone is very fragmentary (e.g. Shumilovskikh et al. 2017).

Map of the study area: a vegetation map of Europe; b vegetation map of the study region—vegetation units: F71—North Ukrainian-south Sarmatian lime-pedunculate oak forests, L3—subcontinental meadow steppes alternating with Tatarian maple-pedunculate oak forests, M1—west and central Pontic herb-rich grass steppes, D57—Southeast European xerophytic herb- and grass-rich pine and oak-pine forests, U12—East Sarmatian hardwood alluvial forests (details in Bohn et al. 2003); c location of pollen records in relation to the archaeological site Kurilovka-2
In this paper, we present vegetation and environmental reconstructions from the sediment core Sudzha, taken from the oxbow lake in the forest-steppe zone (Fig. 1b). The studied oxbow lake is located near the archaeological site Kurilovka-2, representing a promising archive to study human environmental interactions. The settlement Kurilovka-2 is a part of an archaeological complex at the village of Kurilovka (Fig. 1c). Archaeological data reveal traces of human presence on site in the Neolithic and in the following Bronze Age (Rodinkova et al. 2017). However, the first period of intensive settlement activities falls in the 3rd–8th centuries ce. This activity was by Proto- and Early Slavic archaeological cultures such as Kiev (3rd–the first half of the 5th century ce), Kolochin (the second half of the 5th–the third quarter of the 7th century ce) and the Sakhnovka stage of Volyntsevo (the end of the 7th–8th centuries ce). From the end of the first millennium ce, the settlement Kurilovka-2 was abandoned. The new period of its settlement began in the 17th century ce and continued to the second half of the 20th century ce.
The aim of this study is to reconstruct the vegetation history and environmental changes in the Sudzha region during the late Holocene in order to decipher the anthropogenic influence on the forest-steppe ecotone. Within the scope of the study, we ask following questions: (1) What was the course of the general vegetation development in the region? (2) How did the local environment and vegetation at the study site change? (3) How strong was the anthropogenic impact of different cultures on the vegetation? (4) When did the transformation to an agricultural landscape take place? In order to answer these questions, we provide the details of our analysis of pollen and non-pollen palynomorphs in the Sudzha sediment core. Furthermore, we compare this to the existing but still unpublished on-site pollen profiles from Kurilovka-2 and archaeological information in order to identify the intensity of the human impact on forest-steppe over the studied time period.
Study area
The study area is situated on the Russian-Ukrainian border in the south-west of the Kursk Oblast (Russia). The archaeological site Kurilovka-2 is located on the remnant of a low terrace at the confluence of the Sudzha and Psiol Rivers, belonging to the Dnieper River basin of the Black Sea catchment area. The site is surrounded by oxbow lakes of former meanders of the Sudzha and the Psiol Rivers, in one of which the off-site core Sudzha was obtained (Fig. 1c).
The climate of the region is cold temperate and classified as Dfb according to Köppen and Geiger. The annual average temperature in Sudzha town is 6.5 °C, reaching 19.5 °C in July and − 7.6 °C in January. The annual precipitation is 598 mm with highest precipitation in July (76 mm) and lowest in February (32 mm). The investigated area is a part of the forest-steppe zone with zonal soils as typical chernozems with different degrees of erosion. Inter-stream areas are composed mainly of chalky rocks, clays and loess-like loams, low terraces and floodplains consisting of sands and sandy loams.
The study site is situated to the middle of the forest-steppe zone with a potential vegetation of alternating meadow steppes and dry grasslands with temperate deciduous forests. The forests are composed of Quercus robur, Tilia cordata, Ulmus glabra, U. laevis, Fraxinus excelsior, Acer platanoides, A. campestre and A. tataricum, growing together with shrubs of Corylus avellana, Rhamnus cathartica, Prunus spinosa and Cerasus fruticosa (Bohn et al. 2003). However, the majority of the area is today covered by an agricultural landscape. The potential natural vegetation at the site Kurilovka-2 is estimated to consist of alluvial hardwood forests (Bohn et al. 2003). Until the 1990s, the site was intensively ploughed. Today it is covered by ruderal vegetation and used for haymaking.
Numerous archaeological investigations in the Sudzha region provide a list of 148 archaeological sites (Fig. 2, ESM). Among them there are 118 settlements (including a large number of multilayer ones) commonly in valleys of the Sudzha and Psiol rivers and their tributaries, and 41 burial grounds mostly located on watershed (Fig. 2, ESM; Kashkin 2000 updated by V. Rodinkova). Most of the burial grounds (38 sites) are kurgans with largely unclear chronological attribution. The settlement materials show presence of humans in the Sudzha region since the early or mid-Palaeolithic (one site) and sporadically during the late Palaeolithic (two sites) and the Neolithic (seven sites). The settlement activity was the highest in the Bronze Age (mid third–beginning of first millennium bce) with 64 known sites. In the early Iron Age (7th–1st century bce), the density of the archaeological sites decreases (25 sites). Materials from the Roman period (1st–5th century ce) are registered on 21 sites. The objects of the late Roman period (3rd–the first half of the 5th century ce) belong to the Cherniakhov and Kiev cultures. In the mid-5th century ce, the Cherniakhov culture disappeared, while the Kiev culture transformed to the Kolochin culture (14 sites), which was the only archaeological culture of the Migration Period (the second half of the 5th–the 7th century ce) in the Sudzha region, and later was replaced by Volyntsevo (Sakhnovka stage) (the end of the 7th–8th century ce), represented by two settlements. In the late Early Middle Ages (9th–10th century ce), the number of discovered archaeological sites increases to 29, belonging to the Romny-Borshevo culture, while the Kievan Rus period (11th–13th century ce) decreased to 15 settlements. After a period of abandonment of the territory due to the danger from nomad tribes, the settlement process started again in the 17th century. At this point, the Sudzha region was included into the Moscow State and natives of the Polish-Lithuanian Commonwealth and other parts of Russia settled here (Babin 2015).

Archaeological sites of the Sudzha region: a type of the sites: 1—kurgan, kurgan cemetery; 2—burial ground; 3—hillfort, settlement, temporary site; b chronological position of the sites: 1—kurgan, kurgan cemetery without a clear date; 2—Stone Age site; 3—Bronze Age site; 4—Early Iron Age site; 5—Late Roman Period site; 6—Migration Period site; 7—Early Middle Ages, Middle Ages (Kievan Rus Period), Late Middle Ages site. The numbers of sites on the map correspond to the numbers in the ESM
Materials and methods
In order to provide vegetation and environmental reconstructions, in 2010 we obtained a sediment core from the oxbow lake located at a distance of about 120 m from the archaeological site Kurilovka-2. For comparison with the off-site record Sudzha, pollen data from three on-site soil profiles from Kurilovka-2 are presented here. Since these profiles were studied by different laboratories and at different times and have not been published until now, we present here the laboratory treatment for every archive in detail.
Sediment core Sudzha
The sediment core Sudzha (51°08′15″N, 35°17′17″E, 134 m a.s.l.) is composed of organic-rich material from 20 to 200 cm covered by 20 cm of water, changing to a transition zone between 200 and 215 and finally to bluish green clays below 215 cm. The bottom (253–267 cm) is rich in sand. At 30 cm and 50 cm Phragmites rhizomes occur, while from 137 to 152 cm a few narrow clay layers appear. For the chronology, one AMS radiocarbon date on bulk peat and two on plant remains (Table 1) were carried out in the Radiocarbon Laboratory of Poznan (Poland) and Erlangen (Germany). An establishment of the age-depth model (Fig. 3) was carried out using the Clam 2.2 package (Blaauw 2010) and the Intcal13 calibration curve (Reimer et al. 2013).

Age-depth model of the sediment core Sudzha
A total of 29 subsamples were collected at 6–10 cm intervals from the sediment core Sudzha. The laboratory treatment included demineralization with cold hydrochloric acid (10%), followed by cold hydrofluoric acid (70%) overnight, acetolysis (Erdtmann 1960) and sieving through 200 µm metallic mesh and 6 µm nylon mesh using an ultrasound bath (less than 1 min). One tablet of Lycopodium spores (Batch number 177,745 or 1,031) was added at the beginning of the preparation in order to calculate the concentration of microfossils (Stockmarr 1971). Prepared subsamples were stored in glycerin and counted under 400 × to 1,000 × magnification. Counts of 300 pollen grains of terrestrial plants per sample were made. In cases of low pollen concentration, about 100 pollen grains were counted if possible. Pollen identification and taxonomy follows Beug (2004) and Moore et al. (1999). Beside pollen and plant spores, non-pollen palynomorphs (NPP) and charcoal particles were identified and counted. For NPP identification we mostly used Pals et al. (1980), van der Wiel (1982) as well as the NPP database http://nonpollenpalynomorphs.tsu.ru/. Pollen and NPP are expressed as percentages of the total sum of pollen excluding water and wetland plants. All diagrams (Figs. 4, 5, 6, 7) were constructed using C2 version 1.5.6 (Juggins 2007).

Palynological %-diagram of the sediment core Sudzha: a arboreal pollen (AP) and non-arboreal pollen (NAP), b spores, pollen of water and wetland plants, algae, animal remains, fungal spores and other non-pollen palynomorphs (NPP). Circles indicate presence of pollen clumps. Grey bands indicate anthropogenic activity phases

Macroremains diagram of the sediment core Sudzha. Stratigraphical description: 1—Phragmites peat, 2—herbaceous peat, 3—Nuphar peat, 4—tree leaf peat

Pollen diagram of the soil profiles 2/15. Legend to the stratigraphy: 1—brown sand with mollusc shells; 2—yellow sand; 3—grey sand with thin layers; 4—brown silty loam; 5—light brown sand, laminated, with mollusc shells; 6—dark-brown siltstone; 7—greyish-green sand

Pollen diagram of the soil profile 3/15. Legend to the stratigraphy: 1—litter and turf; 2—brown silty sand with mollusc shells and roots; 3—dark-grey highly compressed sand; 4—light-grey sand; 5—whitish sand; 6—greyish-brown sand
The Holocene changes in forest coverage in a 20 km radius the Sudzha site were reconstructed using the Best Modern Analogue (BMA) technique (Overpeck et al. 1985; Nakagawa et al. 2002). The details of this approach are described in our previous publications (Novenko et al. 2014; Shumilovskikh et al. 2017). In this study, we used squared-chord distances (SCD) as the index of dissimilarity between modern and fossil pollen spectra with a threshold T = 0.4 and use eight best analogues to calculate an average value of reconstructed forest coverage. The dataset of modern analogues consist of 720 surface pollen assemblages from a wide variety of landscapes in Europe and West Siberia (Novenko et al. 2014). Forest coverages around the surface pollen spectra were derived from MODIS satellite images (Hansen et al. 2003).
Plant remains larger than 200 µm from pollen samples were studied for the botanical composition of the organic-rich parts of the sediment. Samples were analyzed with a microscope with a magnification of 100 ×. Plant macrofossils were identified following Katz et al. (1977) and Dombrovskaya et al. (1959). The plant macrofossil content is presented by volume percentages for each taxon, estimated in steps of 5% and subsequently calculated for the total volume.
On-site records Kurilovka-2
Three soil profiles were studied from the archaeological site Kurilovka-2 (Fig. 1c). The soil profiles 2/15, 3/15 and 10/16 were dated by humic acids in the Laboratory of Radiocarbon Dating and Electronic Microscopy, Institute of Geography RAS (Russia), using the LSC method.
Profiles 2/15 and 3/15 were located on the NE slope of the outcrop facing the River Sudzha, on the border of the archaeological site. Profile 2/15 is situated closer to the water, while profile 3/15 is about 5 m further up the slope. Both profiles represent alternating layers of organic-rich sandy loam and sand (Figs. 6, 7). The upper parts of both profiles contain mollusc shells. In profile 3/15, pottery fragments of the Neolithic, the Early Slavonic and Modern periods were found. Subsampling of the soil profiles 2/15 and 3/15 was carried out in the field in July 2015 with intervals of 5 cm in profile 2/15 (11 samples) and in profile 3/15 (12 samples). Chemical preparation of the samples was carried out using 10% HCl, 10% KOH and heavy liquid (KI, СdI2) with a density of 2.35 (Grichuk 1938, 1940; Chernova 2004). One tablet of Lycopodium spores (Batch number 177,745) was added at the beginning of the preparation in order to calculate the concentration of microfossils (Stockmarr 1971). Prepared subsamples were stored in glycerin and counted under 400 × magnification up to at least 300 pollen grains.
Soil profile 10/16 is located on the lower northern part of the outcrop, inside the archaeological site. It is composed of sandy loam in the upper part changing to the heavy loam in the middle and to the silty loess-like loam in the lower part of the profile (Fig. 8). In total, 12 samples were collected in the field from the soil profile 10/16 in November 2016. The samples (10 g of soil) were processed as recommended for mineral soils, using acidification with 10% HCl, with one tablet of Lycopodium spores (Batch number 3862), boiling in 10% KOH, and centrifuging with sodium polytungstate (Torresan 1987). Counting was carried out at a magnification of 400 × up to 100 pollen grains per sample due to the low pollen concentration. Pollen of all three soil profiles is expressed as percentages of the total sum of pollen of terrestrial plants.

Pollen diagram of the soil profile 10/16. Stratigraphical description: Ao—sod, dark grey sandy loam, crumb to granular structure, intertwined plant roots; Ap—dark grey sandy loam, firm, crumb to granular structure, chalk crumbles and ceramic fragments, intertwined plant roots, lower boundary straight, distinct transition in density; AE—grey sandy loam, loose, with off-white skeletans, lower boundary straight, distinct transition in colour and density; 2Ab—dark gray medium textured loam, humified, powdery, granular structure, loose; AEBt—brownish-gray silt loam, homogeneous, skeletans and thin humus-clay cutans
Results
Off-site diagram Sudzha
Pollen
In total, 96 pollen taxa were documented in the Sudzha core. Based on changes of pollen taxa percentages, the Sudzha pollen diagram was divided in three local pollen zones (Fig. 4).
The lowest part of the sediment (260–240 cm) is characterized by very low pollen concentrations of 160–700 pollen grains/ml. Due to low counts the data are not presented in the pollen diagram. The pollen spectra are represented by Pinus diploxylon-type and Chenopodiaceae, accompanied by the Pinus haploxylon-type, Ephedra distachya-type and E. fragilis. Algae are represented by the green algae Pediastrum and Botryococcus and fungi by spores of the mycorrhizal fungus Glomus.
The pollen spectra of zone Sud-1 (240–220 cm, ~ 2.8–2.5 cal ka bp) are dominated by broadleaf tree taxa like Quercus robur-type (18–31%), Ulmus (5%), and Tilia (7%) as well as maxima in the pioneer Betula (10%) and the wetland tree Alnus (16%). Non-arboreal pollen (NAP) spectra are dominated by Artemisia (4–8%) and Poaceae (6–10%). The algal assemblage changes from Pediastrum and Botryococcus at 234 cm to HdV 128, accompanied by remains of Cyanobacteria (sheaths of Gleotrichia-type, heterocysts of Rivularia-type), Zygnemataceae and dinoflagellate cysts. For the first time, testate amoebae, oocytes of Rhabdocoela and saprotrophic fungi appear.
The zone Sud-2 (220–48 cm, 2.5–0.2 cal ka bp) is characterized by a dominance of Quercus robur-type (40–74%) and the presence of anthropogenic indicators. The zone is divided into two subzones. In the subzone Sud-2a (220–130 cm, 2.5–1.3 cal ka bp), Pinus diploxylon-type (1–16%), Ulmus (3–10%) and Betula (2–7%) are dominant alongside Quercus robur-type (42–74%) with an increase of Corylus up to 9% towards the end of the subzone. NAP is characterized by a dominance of Poaceae (14%) and Artemisia (1–3%) and the regular presence of Plantago major-media-type, Cichorioideae, Rumex acetosella-type, Ranunculus acris-type, Cannabaceae and Chenopodiaceae with a maximum between 190–150 cm corresponding to a period between 2,000 and 1,600 cal year bp (lower grey band in Fig. 4a). In the subzone 2b (130–48 cm, 1.3–0.2 cal ka bp), Quercus robur-type reduces to 40–53% while Tilia (3–6%), Alnus (6–13%) and Betula (5–8%) increase. NAP is similar to the previous subzone, but exhibit the first appearance of Secale and regular presence of Cerealia-type, Ranunculus acris-type, Cannabaceae, Chenopodiaceae and Plantago lanceolata-type. Maximum of values occur between 120–80 cm corresponding to 1,100 and 600 cal year bp (upper grey band in Fig. 4a).
Pollen assemblages of wetland and water plants are similar for the entire zone Sud-2. They are represented by Cyperaceae, Lythrum, Filipendula, Sparganium-type, Lemna, Nuphar, Nymphaea, Myriophyllum spicatum, Sagittaria sagittifolia and Potamogeton. From 182 cm upwards, mucilaginous hairs and trichosclereids of Nymphaceae are present in the sediment. Algal assemblages are dominated by HdV 128 and several maxima of Gleotrichia-type and Zygnemataceae. Pediastrum is more common in the subzone Sud-2b.
Animal remains are more frequent in the subzone Sud-2a with testate amoebae, Rhabdocoela, eggs of Rotatoria and Tardigrada, several NPP as well as helminth eggs of Dicrocoelium and Diphyllobothrium. Lignicolous fungi (Diporotheca, Helicoon, Savoriella) are more frequent in Sud-2a, while spores of coprophilous fungi occur sporadically during Sud-2. There is a characteristic charcoal maximum at 182 cm.
The zone Sud-3 (48–20 cm, 0.2 cal ka bp–present) shows a dominance of NAP with Poaceae (24–36%), Artemisia (3–9%), Chenopodiaceae (3–10%), Cerealia-type (1–6%), Secale (2–7%), Fagopyrum (1%) and Cannabaceae (3–18%). Several other herb types have increased occurrence such as Ranunculus acris-type, Rumex acetosella-type, R. scutatus-type, Cichorioideae, Plantago major-media-type, P. lanceolata-type, Xanthium strumarium, Centaurea cyanus and Polygonum aviculare. Pollen assemblages of wetland and water plants are quite diverse including Rumex aquaticus, Hottonia palustris, Persicaria maculosa-type and Typha latifolia-type. While algae, animal remains and saprotrophic fungi do not change significantly, the coprophilous fungal spores Podospora and Sordaria occur more frequently. The charcoal maximum of the diagram is noted at 28 cm depth.
Plant macroremains
The organic-rich part of the sediment core Sudzha is represented by eutrophic peat (Fig. 5). The diagram of plant macrofossils can be divided into three main zones (Fig. 5). MRZ 1 (215–178 cm, 4 samples) is dominated by fragments of bark and wood of Alnus and the remains of Humulus lupulus, while Calla palustris remains are sporadic. In MRZ 2 (depth 178–55 cm, 12 samples), the plant assemblages become more diverse. Nuphar lutea 15–30%, Phragmites sp. 10–15%, Calamagrostis lanceolata 5–15%, Comarum palustre and Carex cespitosa are important components together with Alnus remains (55–90%). In the upper zone MRZ 3 (55–22 cm, 5 samples), Phragmites dominates (40–95%) together with C. palustris (10–15%), Typha angustifolia (5–20%) and C. cespitosa (5–15%).
On-site diagrams of Kurilovka-2
Soil profile 2/15
In total 67 pollen taxa were documented. The pollen diagram was divided in three local pollen zones KII (Fig. 6). The zone KII-1 (92–112 сm) is characterized by the dominance of NAP (up to 80%), composed of Poaceae (7–10%), Asteraceae (5–10%), Cichorioideae (12–21%), Rosaceae (4–14%), Fabaceae, Ranunculaceae and others. Tree taxa are presented by Pinus (6–12%), Alnus (6–11%) and broadleaf trees such as Tilia (6%), Quercus robur-type (2%), Ulmus and Acer. During the zone KII-2 (92–63 сm), the amount of AP decreases to 22%, while Poaceae (13%) increase together with Cerealia-type (4%) and Fagopyrum (2%) as well as ruderal taxa such as Chenopodiaceae, Onagraceae, Urtica, Plantago and Artemisia. The zone KII-2 is followed by a pollen sterile sand layer (63–42 cm). In the zone KII-3 (32–42 сm), Pinus (17–24%) and Salix (3–10%) increase while most broadleaf taxa decrease. The NAP is dominated by Chenopodiaceae (17–28%). Characteristic is an increase in Potamogeton (up to 30%) and in spores of Bryales (up to 75%).
Soil profile 3/15
In total, 47 pollen taxa were identified. The pollen diagram is divided in five zones (Fig. 7). The first zone KIII-1 is represented by one sample at 66 cm depth. Its spectrum is dominated by AP (62%) with Alnus (38%), Tilia (11%) and Betula (9%). Among the NAP, Cichorioideae (12%) and Poaceae (6%) have the highest percentages, while Cerealia-type and Fagopyrum are present. A rapid decline in AP (27–34%) characterizes the zone KIII-2 (63–42 сm) due to an increase in Cerealia-type together with Cichorioideae (up to 23%) and Chenopodiaceae (up to 23%). Zone KIII-3 (42–28 cm) is characterized by a strong increase in Pinus (19–26%) and a maximum in Chenopodiaceae (up to 29%). Moreover, there is an increase in Potamogeton (6%) and a maximum in spores of Bryales (up to 52%). In the zone KIII-4 (28–17 сm), the role of Pinus (10%) decreases with an increase in Alnus (22%) and Tilia (6%). Poaceae increase up to 16% together with the Cerealia-type (4%), Artemisia, Ranunculaceae, Campanula and Urtica. Wetland and water plants show maxima of 13% and 6%, respectively. In the upper zone KIII-5 (17–3 сm), there is an increase in AP up to 68% with Alnus (31%), Pinus (21%), Quercus robur-type (7%) and Tilia (5%). NAP is dominated by Poaceae (10%) and Chenopodiaceae (5–13%).
Soil profile 10/16
The samples are characterized by very low pollen concentrations and poor pollen preservation, therefore only 5 from 12 samples could be analysed (Fig. 8) and just 20 pollen taxa were verified. The two lower-most samples (70–75 cm and 65–70 cm) show about 50% of AP with a dominance of Betula (28%) together with Tilia (9%), Quercus robur-type (3–5%) and Alnus (6%). NAP is represented mainly by Artemisia (23%) and other Asteraceae (19%). In addition, Apiaceae, Onagraceae, Fabaceae and Chenopodiaceae were found. As primary anthropogenic indicators, pollen of Cerealia-type and Fagopyrum as well as panicoid forms of phytoliths are present. The samples are rich in charcoal particles (not shown on diagram). The sample from 40 to 50 cm shows an increase in Pinus (13%), disappearance of broadleaf tree taxa (Quercus robur-type, Tilia) and a high Cerealia-type value up to 20%. The upper two samples (10–15 cm and 0–1.5 cm) are characterized by the dominance of Pinus (38–44%), Poaceae (10–24%), Cichorioideae (4–13%) and Cerealia-type (12–18%). Other NAP taxa present are Chenopodiaceae and Fabaceae, while Polygonum aviculare appears for the first time.
Discussion
In the discussion, we present the regional vegetation change reconstructed from the Sudzha sediment core as well as the local development of the site, followed by the reconstructions from on-site records in Kurilovka-2. We then compare our palynological reconstructions with archaeological and historical data from the Sudzha region.
Regional vegetation change reconstructed from the pollen diagram Sudzha
The pollen spectra from the core bottom indicate the presence of conifers such as Pinus sibirica, Picea and dry steppe indicators such as Ephedra. In the absence of radiocarbon dates and taking the low pollen concentration into account, it is highly speculative to make a correlation with other records. However, we compare this phase palynologically with the Late Glacial or beginning of the Holocene from the core SV-8 in the floodplain of the Svapa River, a tributary of the Seim River about 60 km north of Sudzha (Borisova et al. 2006), and the Avdeevo profile from the Seim River near Kursk (Panin et al. 2017). Alternatively, these pollen assemblages may also result from erosion as a local event in the river channel or as a fire event sometime during the Holocene. A large amount of Glomus-type at the base of the core clearly indicates erosion.
The first zone (pollen zone Sud-1) reflects the dominance of broadleaf trees, suggesting the spread of mixed oak forests and alder-willow carr by strong reduction of pine in the surroundings. Considering the low pollen concentration and minerogenic composition of the sediment, this zone most probably represents mixed assemblages on the basis of the oxbow lake.
The pollen zone Sud-2 reflects a very stable environment with filling of the oxbow lake between 2,500 and 200 cal year bp (Fig. 4). The sediment accumulation rate for this period is 12–14 year/cm. The pollen reflects the presence of mixed oak forests and forest patches with elm, lime, hazel and ash. The forest cover varied between 30 and 36% suggesting open landscapes covered by herb-rich meadows or meadow steppe with broadleaf and birch-poplar patches.
Compilation of the records from the forest-steppe region shows that Sudzha was located in the forest-steppe ecotone, close to the southern border of closed deciduous forests (Shumilovskikh et al. 2017). Located in the south, Sudzha has the highest values of broadleaf tree taxa in comparison to the northern records but a comparable forest cover between 50% in Klukva (Novenko et al. 2015) and 20% in Podkosmovo (Novenko et al. 2014). The more eastern record Krasivo in the western Belgorod region (Ershova 2017) has lower values of broadleaf forests and higher values of pioneers such as Betula due to the more continental climate. Similarly, pollen data from the basin of the Don River demonstrate the presence of forest-steppe vegetation during the late Holocene (Spiridonova 1991).
Later on, human-forced changes can be traced in the Sudzha area. Between ~ 2,000 and 1,600 cal year bp (~ 50 bce to 350 ce), there is a pronounced phase of agricultural (Cerealia-type, secondary anthropogenic indicators) and pastoral (spores of coprophilous fungi) activities, including deforestation (decrease in AP, spread of Corylus, Poaceae maximum) and burning (maximum in charcoals). This signal corresponds well to the general pattern of increased anthropogenic activities in the Mid-Russian Plain since the Iron Age (Khotinsky 1993; Shumilovskikh et al. 2017). It is interesting that the charcoal maximum in Sudzha coincides with maximum of macrocharcoals in the Selikhovo record (Novenko et al. 2016) as well as in the Podkosmovo record (Novenko et al. 2014), indicating increased fire activities over the whole region that could have had anthropogenic or climatic causes.
Starting from about 1,500 cal year bp (450 ce), an increase of Alnus indicates a general paludification of the region possibly due to the ongoing filling of the oxbow lakes. The second pronounced occupation phase is indicated between 1,100 and 600 cal year bp (850–1350 ce). The further decrease in oaks suggests deforestation, maxima in Cerealia-type and Secale indicate agricultural activities, a general increase in anthropogenic indicators with the first occurrences of Plantago lanceolata-type, Cirsium and Xanthium strumarium points to higher anthropogenic pressure than before. However, low charcoal values might indicate that settlements were located further away from the studied oxbow lake.
The last 200 years (18th century, Sud-3) reflect strong and rapid changes in the area. Total deforestation took place with a reduction of forest cover below 20% mainly due to felling of broadleaf forests but also birch. Ploughing, cereal, rye and buckwheat cultivation are visible from the pollen data. These activities led to a strong spread of ruderal plants and weeds, also including a general increase of open areas and spread of steppes. There is an interesting maximum of Cannabaceae, which we interpret as hemp rotting in the oxbow lake for hemp fibre production. A charcoal peak suggests the presence of local fires or possibly a settlement in the close vicinity of the site. In addition, pasture is suggested close to the site. Forest cover decreases from 35% to 12–18%, which is comparable with the modern forest cover estimates of 14.2% based on MODIS. With visible and very strong human impact in 17th–18th century, the Sudzha record is in line with other palynological reconstructions from the Mid-Russian Plain (e.g. Khotinsky 1993; Shumilovskikh et al. 2017).
Local development of the oxbow lake
The local development of the coring site is connected to a variety of processes. The bottom of the core represents the lacustrine conditions, indicated by the dominance of Botryococcus and Pediastrum. After a hiatus during the mid-Holocene, formation of the oxbow lake initiated the sedimentation of organic-rich material (pollen zones Sud-1 to Sud-3). Several species identified in the peat layers through pollen and botanical macroremains indicate the ecological conditions on the site.
In general, the peat composition indicates presence of alder carr on the shore of the oxbow lake, which was slowly filled over 2,500 years. In the lower part (212–60 cm), the sediment was formed under shallow lake conditions from wood and leaf remains of Alnus and later by Nuphar lutea and peatland plants (Fig. 5). Humulus lupulus was covering the trees along the river valleys. Calamagrostis grew together with sedge peat hillocks on nutrient-rich ground. Comarum palustre is one of the typical peatland species (Lapshina 2003). Calla grew along the banks of standing and flowing waters. Beside Nuphar lutea, several other water and wetland plants like Nymphaea, Lemna, Myriophyllum spicatum, Sagittaria sagittifolia, Potamogeton, Typha angustifolia and/or Sparganium, Lythrum and Filipendula were growing in the oxbow lake and on its shore.
At the top (60–20 cm), the continuing filling of the lake led to the development of Phragmites reed and several Carex spp. Carex cespitosa is the indicator of tussock microlandscape, characterized by two microhabitats: the tussock tops with an abundance of light and normal moisture level, and pools which are strongly shaded and waterlogged. Pools were filled with water or inhabited by hygrophilous species such as Equisetum fluviatile, Phragmites australis and Comarum palustre. Carex lasiocarpa and C. globularis are typical peatland species, growing in eutrophic and meso-eutrophic conditions. Newly invading wetland species are documented by pollen such as Rumex aquaticus-type, Hottonia palustris, Persicaria maculosa-type, Typha latifolia-type and Caltha. In general, the botanical composition indicates that the peat was formed under eutrophic conditions.
After the formation of the oxbow lake, algal assemblages are dominated by HdV 128, cyanobacteria and Zygnemataceae (Fig. 4b). HdV 128 is still not affiliated to any known species but by its palaeoecological records it is related to shallow eu- to mesotrophic open water (Pals et al. 1980; van Geel et al. 1982). Zygnemataceae inhabit shallow, stagnant, oxygen-rich water (van Geel 2001). The presence of Gleotrichia-type and Rivularia-type indicates an alkaline environment (pH 7.5–8), rich in oxidizable organic compounds (van Geel et al. 1982).
Animal remains are represented by a wide variety of testate amoeba, rotifer eggs, tardigrade eggs, oocytes of Rhabdocoela and further taxonomically unknown resting eggs, indicating open water conditions (Fig. 4b). Types HdV 179 and HdV 187D indicate stagnant shallow open water with eutrophic conditions (van Geel et al. 1982, 1989). The finds of helminth eggs in the sediment are interesting. The lancet liver fluke Dicrocoelium sp. parasitizes the bile ducts of a wide range of wild and domestic animals such as Bovidae, rabbits and rodents (Le Bailly and Bouchet 2010). Its eggs found in natural lake sediments indicate presence of the disease in the area and animals close to the site. Diphyllobothrium infects the small intestine of mammals, including humans, through the consumption of raw fish infected by the plerocercoid larvae of the tapeworm (Lardín and Pacheco 2015). An adult tapeworm produces eggs, which are passed in the feces, which need to be deposited in freshwater for the further development of the worm. The presence of the eggs indicates infected mammals or humans at the shore of the oxbow lake in the first millennium ce.
Fungal assemblages are represented mainly by lignophilous species, which are more common in the zone Sud-2a (Fig. 4b). Their maxima correlates very well with the peat composition of Alnus wood and leaf remains (Fig. 5). Coprophilous fungi are mainly represented by generalists such as Cercophora and sporadically by more strictly coprophilous species, suggesting rather low grazing pressure in the surroundings of the lake.
From NPP with unknown taxonomical origins, the occurrence of the type TEP-2 in the upper part of the record is interesting. This microfossil was first described from soil sediments taken in the savannas of Roraima State in Amazonia (Rodríguez-Zorro et al. 2017). It occurred during the dry periods with grassland dominance and correlates with microcharcoal peaks and Gelasinospora (Rodríguez-Zorro et al. 2017). In our record, TEP-2 only corresponds to a charcoal maximum once and is present under forested and open landscape conditions. Another NPP of unknown origin UAB-27 was described from an Early Neolithic settlement of La Draga (Revelles et al. 2016) and from lacustrine sediments in Lake Banyoles (Revelles and van Geel 2016) in Spain. It occurs during times of soil erosion, in waterlogged layers and in charred storage structures (Revelles et al. 2016; Revelles and van Geel 2016). In Sudzha it correlates with increased soil erosion and increased human activities, implied by a peak of microcharcoals and a maximum in Glomus-type (Fig. 4b). In our opinion, UAB-27 might be an egg or resting stage of a water or wetland invertebrate, as it has also been found in the wetland core Kongor from NE Iran (unpublished).
Vegetation change derived from the on-site soil profiles at Kurilovka-2
Soil profiles 2/15 and 3/15 located close to each other at the rim of the archaeological occupation site show similar features in the stratigraphy. Both profiles are composed of sandy loams intercalated with sandy layers, indicating formation of alluvial soils with possible flooding. For chronology, we use radiocarbon dates by humic acids (HA) from soil organic matter. HA are the most biologically stable fractions in the study soils and we can use them as a “dating fraction” both for natural soils (Chichagova and Cherkinsky 1993) and for soils of archaeological sites (Wild et al. 2013; Zazovskaya et al. 2017). Dates by HA reflect the mean residence time of carbon in these soils. These dates can be interpreted as the minimum age of the studied soils. Based on radiocarbon dates (Table 1), both soil profiles are quite young and do not exceed 500 years. In both profiles, AP curves vary mostly between 20 and 40% and never exceed 60%, indicating rather open local conditions. Most likely the vegetation was patches of broadleaf forests and alder-willow carr alternating with herb-rich meadows and the ruderal vegetation of the settlement.
Changes in trees and ruderal plants in the soil profile 3/15 indicate at least two occupation phases. In the zone KIII-2, AP decreases to 30% together with an increase in Cerealia-type, Chenopodiaceae, Cichorioideae and Rumex. Furthermore, pollen of buckwheat (Fagopyrum), present throughout the profile 3/15, and low AP values correlate with the zone Sud-3 of the Sudzha record, supporting a chronological position in the last 250–300 years. The second occupation period can be interpreted in the zone KIII-4 by maxima of Cerealia-type, Poaceae, Artemisia, Ranunculaceae and Urtica. This phase is modern (Table 1) and was followed by a period of abandonment of the territory and recovery of the broadleaf forests in the surroundings. An increase in pollen of water and wetland vegetation towards the top of the record suggests an increase of wetness in the area and possible flooding events in spring or summer.
Soil profile 2/15 was radiocarbon dated, but the ages are inverted. Even so, they do not pre-date the 16th century (Table 1). Similar to the soil profile 3/15, the vegetation had a rather open character with broadleaf forests and meadows. Zone KII-2 shows a settlement phase, indicated by maxima of Cerealia-type, Fagopyrum, Artemisia, Poaceae and a general decrease of broadleaf trees especially Tilia. The lack of Fagopyrum in KII-1 is remarkable, possibly indicating that the base of soil profile 2/15 is older than the beginning of soil profile 3/15. Palynologically, the occupation in KII-2 correlates with Sud-3 by low AP and the presence of Fagopyrum, and with KIII-2 by low Pinus and Tilia, a maximum in Cerealia-type and the presence of Fagopyrum. The youngest zone KII-3 shows high Pinus and Chenopodiaceae values, indicating abandonment of the settlement and therefore correlating to KIII-3. In addition, maxima in water plants and spores of Bryales correlate well with KIII-3.
Soil profile 10/16 was taken inside the archaeological site, which is characterized by dry conditions. Therefore, pollen spectra of this profile are very limited due to the poor pollen preservation and should be interpreted with caution (Dimbleby 1985). The general vegetation development is reflected by this record. The bottom of the profile (65–75 cm) indicates the presence of forest-steppe with broadleaf and birch patches in a meadow steppe. Local anthropogenic activities are indicated by a few finds of Cerealia-type, Fagopyrum and panicoid phytoliths, suggesting slash and burn agriculture. Rare archaeological material of the Early Slavonic tradition included in this layer suggests that its accumulation started in the 3rd–8th century ce. Analysis of the cultural layer (sample 40–55 cm) demonstrates that local agricultural activities increased (20% of Cerealia-type) and a deforestation of broadleaf trees took place. A single radiocarbon date at 50–55 cm (Table 1) indicates that this period was around 13th–14th century ce. However, this is in contrast to the archaeological finds, represented by Early Slavonic pottery fragments. The two samples from the modern soil (0–15 cm) show a strong increase in pine values, also reflected in soil records 2/15 and 3/15. In general, the vegetation was rather open and actively used for agriculture as suggested by high Cerealia-type values. The change from birch-oak-lime assemblages to cereals and herb meadow assemblages correlates with the transition Sud-2/Sud-3 of the Sudzha record.
Pollen records in context of archaeological and historical data
The record Sudzha reveals the presence of humans in the area during the last 2,500 years with three phases of increased anthropogenic activities.
The first phase from the 1st century bce to the 4th century ce correlates to the so-called Roman period covering the rather warm and/or dry climate conditions of the Roman Warm Period (Bianchi and McCave 1999). In the Sudzha region, mainly antiquities of the late stage of the Roman Period (3rd–the first half of the 5th century ce) are present. These are the Kiev and the Cherniakhov cultures, known for their well-developed agriculture (Gorbanenko and Pashkevich 2010; Terpylovsky and Gorbanenko 2010; Radiush 2015). After the collapse of the Cherniakhov culture and during the spread of the Early Slavonic Kolochin culture in the second half of the 5th–7th century ce, the mixed oak forests recovered (Fig. 4a). This recovery can be due to climatic reasons or to a decrease in human impact or both. On the one hand, climate reconstructions show the cooling and glacier advances during the “Dark Ages Cold Period” of 400–600 ce (Patterson et al. 2010). On the other hand, poor evidence of the Early Slavonic cultures of Kolochin and Volyntsevo (Sakhnovka) is the most striking feature of the Sudzha record. We know that the settlements were close to the lake (Fig. 1c), but apparently their way of life must have had a little impact on the natural environment in comparison to the other cultures. Even a very local oak deforestation and slash and burn agriculture at the site Kurilovka-2 close by, suggested by soil profile 10/16 (Rodinkova et al. 2017), is not strongly reflected in the pollen diagram of Sudzha, which might have a much bigger pollen source area and reflect therefore regional processes rather than the local site.
The second visible human impact phase occurs in the 9th–14th century ce, corresponding to the Medieval Warm Period (Bianchi and McCave 1999; Mann et al. 2009), which possibly favoured an increase of agricultural activity in the area. During this period, the Romny-Borshevo culture and later the Kievan Rus established and occupied the high terraces and steep banks of the rivers.
We suggest that density of settlements as well as their topographical position may have a crucial influence on the composition of pollen rain. On the one hand, the density of settlements changes considerably through the time with 21 sites during the Roman time (1st–5th century ce), 14 sites during the Migration Period (the second half of the 5th–the 7th century ce) and just 2 sites of Volyntsevo (the end of the 7th–8th century ce) and increasing again to 29 sites in the late Early Middle Ages (9th–10th century ce) and 15 in the Kievan Rus time (11th–13th century ce) (see Study area section). The periods with the higher numbers of sites correspond to the periods of higher anthropogenic activities in the pollen diagram Sudzha. On the other hand, the members of different cultures occupied different landscape niches. For example, members of the Cherniakhov culture are known for their agricultural activities and preferred the higher parts of terraces with chernozem soils outside the floodplain. Kiev, Kolochin and Volyntsevo settlements tended to be located on the remnants of low terraces in the floodplain area (Gorbanenko and Pashkevich 2010). Only in the late Early Middle Ages (9th–10th century ce), members of the Romny-Borshevo culture moved out of the floodplains to the high terraces and steep banks of the rivers, and the number of recovered archaeological sites increases to 29. The 15 settlements of the Kievan Rus time (11th–13th century ce) are located in similar high topographical locations. Well-developed agriculture and higher topographic location of the settlements of the Romny-Borshevo culture, the Kievan Rus and possibly of the Cherniakhov culture led to larger areas covered by ruderal plants and easier pollen transport by the wind, so that anthropogenic indicators could be transported over longer distances, contributing to the regional pollen rain. In contrast, small settlements of Early Slavs during the 5th–8th century ce inside the alluvial forests are protected by trees and anthropogenic indicators contribute to a very local signal, remaining invisible for regional records. Similar results are obtained from a comparison of the on-site and off-site pollen profiles from the Early Iron Age settlements in the hemiboreal forest zone near Moscow (Ershova and Krenke 2014). This study demonstrates that the early signs of developed agriculture can be seen only in the areas of intensive economic development immediately around the settlements, and only when agriculture covers large areas does the pollen signal become visible in the regional records.
After a period of abandonment of the territory due to the danger from nomad tribes, the settlement process started again in the 17th century, when the Sudzha region was included into the Moscow State and natives of the Polish-Lithuanian Commonwealth and other parts of Russia settled here (Babin 2015). At that time, the active agricultural development of watersheds began, which was connected with the use of improved agricultural tools. This phase of the final forest devastation and spread of agriculture is clearly reflected in the Sudzha record and coincides well with historical data documenting large migration waves from the Polish-Lithuanian Commonwealth to the expanded Moscow State in the 17th century (Babin 2015) and the formation of new cities to defend the borders. Furthermore, Sudzha city was formed in 1661, becoming a local centre of trade and crafts and developing into an administrative centre in 1779 (Chistiakov 2015). The first increase of Secale and Cerealia-type occur in the mid-18th century, showing that transformation from natural to agricultural landscapes lasted at least 100 years. Soil profiles 2/15 and 3/15 provide deeper insights in the local occupation history at Kurilovka-2. They indicate at least two phases of occupation and abandonment of the territory during the last 400 years.
At the end of the 19th century, 90.7% of the population in the Sudzha region settled in rural areas, and agriculture accompanied by animal husbandry was the basis of life here (Berezhnaya 2015). The crops were mainly rye, oats and wheat, with much less millet, barley and peas; buckwheat, sun flower, sugar beet, wild tobacco and hemp were planted occasionally (Berezhnaya 2015; Chistiakov 2015). Traditionally, horses and oxen were involved in ploughing, sheep were kept for wool and cows for milk products (Berezhnaya 2015). The pollen record from Sudzha reflects well the regional development of agriculture with Secale, Cerealia-type, Fagopyrum and Cannabaceae. Interestingly, Cannabaceae pollen is absent in all on-site profiles. We assume that the studied oxbow lake was used for hemp retting, but the hemp fields were located further from the archaeological site Kurilovka-2.
Conclusions
Palynological data derived from the Sudzha oxbow lake sediments provide deeper insights into the vegetation history of the area. They demonstrate the dominance of mixed oak forests between 2,500 and 200 cal year bp with two major settlement phases between ~ 2,000 and 1,600 cal year bp (~ 50 bce to 350 ce) and between 1,100 and 600 cal year bp (850 and 1350 ce) followed by a total deforestation and transformation to an agricultural landscape in the last 200–300 years. The on-site soil profiles from the archaeological excavation Kurilovka-2 have very young ages, providing details of the vegetation change around the site probably over the last 300–400 (maximum 800) years. The data suggest that the Early Slavs in the 5th–8th centuries ce had a very restricted spatial impact on the vegetation, much lower than that of the previous Cherniakhov culture and of the following late Early Middle Ages and Kievan Rus. We suggest that this is connected with different land use strategies, population sizes and topographic situations of the sites, explaining different contributions of the palynological anthropogenic indicators to the regional pollen rain signal.
References
Babin IP (2015) Istoriya osnovaniya goroda Sudzhi [History of Sudzha city foundation]. In: Razdorskiy AI (ed) Sudzha i sudzhane v otechastvennoy i zarubezhnoy istorii i kul’ture [Sudzha and its population in Russian and foreign history and culture]. Kursk University, Kursk, pp 90–107 (in Russian)
Berezhnaya SV (2015) Byt russkogo i ukrainskogo naseleniya Sudzhanskogo uezda v kontse XVIII—nachale XX v. [Daily routine of Russian and Ukrainian populations of Sudzha uezd at the end of XVIII—beginning of XX cent.]. In: Razdorskiy AI (ed) Sudzha i sudzhane v otechastvennoy i zarubezhnoy istorii i kul’ture [Sudzha and its population in Russian and foreign history and culture]. Kursk University, Kursk, pp 172–181
Beug HJ (2004) Leitfaden der Pollenbestimmung. Pfeil, München
Bianchi GG, McCave N (1999) Holocene periodicity in North Atlantic climate and deep-ocean flow south of Iceland. Nature 397:515–517
Blaauw M (2010) Methods and code for ‘classical’ age-modelling of radiocarbon sequences. Quat Geochronol 5:512–518
Bohn U, Neuhäusl R, Gollub G, Hettwer C, Neuhäuslová Z, Schlüter H, Weber H (2003) Map of the Natural Vegetation of Europe. Landwirtschaftsverlag, Münster
Borisova O, Sidorchuk A, Panin A (2006) Palaeogeography of the Seim River basin, Mid-Russian Upland, based on palaeochannel morphology and palynological data. Catena 66:53–73
Chernova GM (2004) Sporovo-pyl’tsevoy analiz otlotzheniy pleistotsena-golotsena. St. Petersburg University, St. Petersburg (in Russian)
Chichagova OA, Cherkinsky AE (1993) Problems in radiocarbon dating of soils. Radiocarbon 35:351–362
Chistiakov AYu (2015) Traditsionnye promysly i remesla Sudzhi i Sudzhanskogo uezda v kontse XVIII–XIX v. [Traditional crafts of Sudzha and Sudzha uezd at the end of XVIII–XIX cent.]. In: Razdorskiy AI (ed) Sudzha i sudzhane v otechastvennoy i zarubezhnoy istorii i kul’ture [Sudzha and its population in Russian and foreign history and culture]. Kursk University, Kursk, pp 182–187 (in Russian)
Dimbleby GW (1985) The palynology of archaeological sites. Academic Press, London
Dombrovskaya A, Korenev MM, Turemnov SN (1959) Atlas of Plant Remains in Peat. Nauka, Moscow-Leningrad (in Russian)
Erdtmann G (1960) The acetolysis method. Svensk Botanisk Tidskrift 54:561–564
Ershova EG (2017) Rekonstruktsiya prirodnoy sredy na osnove analiza bolotnykh otlozheniy v basseine verkhnego techeniya Vorskly (Belgorodskaya oblast). Bulleten Moskovskogo obshchestva ispytateley prirody. Otdel Biol 122:71–79 (in Russian)
Ershova EG, Krenke NA (2014) Izuchenie prirodnykh i kulturnykh landshaftov zheleznogo veka v doline Moskvy-reki metodami palinologii i arkheologii. Vestnik arkheologii. Antropologii i Entografii 3:159–172 (in Russian)
Gorbanenko SA, Pashkevich GO (2010) Zemlerobstvo davnikh slavian. Akademperiodika, Kiev (in Ukrainian)
Grichuk VP (1938) Novyi metod obrabotki osadochnyh porod dlia tseley pyl’tsevogo analiza. Trudy Soveshchaniya Sektsii INQUA 3:159–165 (in Russian)
Grichuk VP (1940) Metodika obrabotki osadochnykh porod bednykh organicheskimi ostatkami dlia tseley pyl’tsevogo analiza. Problemy Fizichskoy Geografii 8:53–58 (in Russian)
Hansen M, de Fries RS, Townshend JRG, Carroll M, Dimiceli C, Sohlberg RA (2003) Global percent tree cover at a spatial resolution of 500 meters: first results of the MODIS vegetation continuous fields algorithm. Earth Interact 7:1–15
Juggins S (2007) C2 Version 1.5 User guide. Software for ecological and palaeoecological data analysis and visualisation. Newcastle University, Newcastle upon Tyne
Kashkin AV (2000) Arkheologicheskaya karta Rossii. Kurskaya oblast. Chast 2. Institute of Archaeology of the Russian Academy of Sciences Press, Moscow (in Russian)
Katz NJ, Katz SW, Skobejewa E (1977) Atlas rastitel’nyh ostatkov v torfah (Atlas of Plant Remnants in Peat). Nedra, Leningradskoje otdelenie, Moscow (in Russian)
Khotinsky NA (1993) Anthropogenic changes in the landscapes of the Russian Plain during the Holocene. Grana 2:70–74
Lapshina ED (2003) Flora bolot yuga-vostoka Zapadnoy Sibiri. Publishing house of Tomsk University, Tomsk (in Russian)
Lardín C, Pacheco S (2015) Helminths: handbook for identification and counting of parasitic helminths eggs in urban wastewater. IWA Publishing, London
Le Bailly M, Bouchet F (2010) Ancient dicrocoeliosis: occurrence, distribution and migration. Acta Trop 115:175–180
Mann ME, Zhang Z, Rutherford S, Bradley RS, Hughes MK, Shindell D, Ammann C, Faluvegi G, Ni F (2009) Global signatures and dynamical origins of the “Little Ice Age” and “Medieval Climate Anomaly”, Science 362:1,256–1,260
Moore PD, Webb JA, Collinson ME (1999) Pollen Analysis. Blackwell Science, Oxford
Nakagawa T, Tarasov P, Kotoba N, Gotanda K, Yasuda Y (2002) Quantitative pollen-based climate reconstruction in Japan: application to surface and late Quaternary spectra. Quat Sci Rev 21:2,099–2,113
Novenko EY, Eremeeva AP, Chepurnaya AA (2014) Reconstruction of Holocene vegetation, tree cover dynamics and human disturbances in central European Russia, using pollen and satellite data sets. Veget Hist Archaeobot 23:109–119
Novenko EY, Tsyganov AN, Volkova EM, Babeshko KV, Lavrentiev NV, Payne RJ, Mazei YA (2015) The Holocene paleoenvironmental history of central European Russia reconstructed from pollen, plant macrofossil, and testate amoeba analyses of the Klukva peatland, Tula region. Quat Res 83:459–468
Novenko EY, Tsyganov AN, Rudenko OV et al (2016) Mid- and late- Holocene vegetation history, climate and human impact in the forest-steppe ecotone of European Russia: new data and a regional synthesis. Biodivers Conserv 25:2,453–2,472
Overpeck JT, Webb T III, Prentice ICA (1985) Quantitative interpretation of fossil pollen spectra: dissimilarity coefficients and the method of modern analogs. Quat Res 23:87–108
Pals JP, van Geel B, Delfos A (1980) Palaeoecological studies in the Klokkeweel bog near Hoogkarspel (prov of Noord-Holland). Rev Palaeobot Palynol 30:371–418
Panin A, Adamiec G, Buylaert J-P, Matlakhova E, Moska P, Novenko E (2017) Two Late Pleistocene climate-driven incision/aggradation rhythms in the middle Dnieper River basin, west-central Russian Plain. Quat Sci Rev 166:266–288
Patterson WP, Dietrich KA, Holmden C, Andrews JT (2010) Two millennia of North Atlantic seasonality and implications for Norse colonies. PNAS 107:5,306–5,310
Radiush OA (2015) Issledovaniya pamiatnikov rubezha pozdney antichnosti i rannego srednevekov’ya (III-V vv. n.e.) v verkhov’yakh reki Sudzha [Study of late antique to early Middle Ages sites (III-V cent. AD) in the upper Sudzha River]. In: Razdorskiy AI (ed) Sudzha i sudzhane v otechastvennoy i zarubezhnoy istorii i kul’ture [Sudzha and its population in Russian and foreign history and culture]. Kursk University, Kursk, pp 18–39 (in Russian)
Reimer PJ, Bard E, Bayliss A et al (2013) IntCal13 andMarine13 radiocarbon age calibration curves, 0–50,000 years cal bp. Radiocarbon 55:1,869–1,887
Revelles J, van Geel B (2016) Human impact and ecological changes in lakeshore environments. The contribution of non-pollen palynomorphs in Lake Banyoles (NE Iberia). Rev Palaeobot Palynol 232:81–97
Revelles J, Burjachs F, van Geel B (2016) Pollen and non-pollen palynomorphs from the Early Neolithic settlement of La Draga (Girona, Spain). Rev Palaeobot Palynol 225:1–20
Rodinkova VYe, Sycheva SA, Shumilovskikh LS, Ershova EG, Ponomarenko EV, Batrachenko EA (2017) Pochvy i landshafty tsentra Russkoy ravniny v ranneslavianskoe vremia (po materialam poseleniya Kurilovka-2 v Sudzhanskom rayone Kurskoy oblasti). Materialy Vserossiyskoy mezhdistsiplinarnoy nauchnoy konferentsii s mezhdunarodnym uchastiem “Paleopochvy, paleoecologiya, paleoeconomika”, Pushchino, 24–24.05.2017. Pushchino, pp 162–167 (in Russian)
Rodríguez-Zorro PA, da Costa ML, Behling H (2017) Mid-Holocene vegetation dynamics with an early expansion of Mauritia flexuosa palm trees inferred from the Serra do Tepequém in the savannas of Roraima State in Amazonia, northwestern Brazil. Veget Hist Archaeobot 26:455–468
Shumilovskikh LS, Novenko EY, Giesecke T (2017) Long-term dynamics of the East European forest-steppe ecotone. J Veget Sci 29:416–426
Spiridonova EA (1991) Evolutsiya rastitelnogo pokrova basseina Dona v verkhnem Pleistotsene—Golotsene (Vegetation changes of the Don River basin in the upper Pleistocene and Holocene). Nauka, Moscow (in Russian)
Stockmarr J (1971) Tablets with spores used in absolute pollen analysis. Pollen Spores 13:615–621
Terpylovsky RB, Gorbanenko SA (2010) Agriculture of habitants of settlement Oleksandrivka I. Arkheologiya i davnia istoriya Ukrainy 4:137–145 (in Ukrainian)
Torresan M (1987) The use of sodium polytungstate in heavy mineral separations. U.S. Geological Survey Open-File Report 87–590
Van Geel B (2001) Non-pollen palynomorphs. In: Smol JP, Birks HJB, Last WM (eds) Tracking environmental change using lake sediments, vol 3. Terrestrial, algal, and siliceous indicators. Kluwer, Dordrecht, pp 1–17
Van Geel B, Hallewas DP, Pals JP (1982) A late Holocene deposit under the Westfriese Zeedijk near Enkhuizen (Prov. of Noord-Holland, The Netherlands): palaeoecological and archaeological aspects. Rev Palaeobot Palynol 38:269–335
Van Geel B, Coope GR, van der Hammen T (1989) Palaeoecology and stratigraphy of the lateglacial type section at Usselo (the Netherlands). Rev Palaeobot Palynol 60:25–129
Van der Wiel AM (1982) A palaecological study of a section from the foot of the Hazendonk (Zuid-Holland), based on the analysis of pollen, spores and macroscopic remains. Rev Palaeobot Palynol 38:35–90
Wild EM, Steier P, Fischer P, Hoflmayer F (2013) 14С dating of humus acids from Bronze and Iron Age plant remains from the Eastern Mediterranean. Radiocarbon 55:599–607
Zazovskaya E, Shishkov V, Dolgikh A, Alexndrovskiy A, Skripkin V, Chichagova O (2017) Organic matter of cultural layers as a material for radiocarbon dating. Radiocarbon 59:1,931–1,944
Acknowledgements
The authors are grateful to Maria Belen Tomaselli for sampling and lab preparation of pollen from the Sudzha sediment core, the editor and two anonymous reviewers for constructive comments and Laura Sutcliffe for polishing the English. This study was partly supported by the Russian Foundation for Basic Research, research projects 16-35-60083 and 16-36-00293, and by the Georg-August-University of Göttingen (Germany). Reconstruction of forest coverage performed by E. Novenko was supported by Russian Science Foundation (Grant 16-17-10045).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by W. Tinner.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Shumilovskikh, L.S., Rodinkova, V.Y., Rodionova, A. et al. Insights into the late Holocene vegetation history of the East European forest-steppe: case study Sudzha (Kursk region, Russia). Veget Hist Archaeobot 28, 513–528 (2019). https://doi.org/10.1007/s00334-018-00711-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-018-00711-4