
Abstract
A systematic review of archaeological and palaeoenvironmental records of cannabis (fibres, pollen, achenes and imprints of achenes) reveals its complex history in Eurasia. A multiregional origin of human use of the plant is proposed, considering the more or less contemporaneous appearance of cannabis records in two distal parts (Europe and East Asia) of the continent. A marked increase in cannabis achene records from East Asia between ca. 5,000 and 4,000 cal bp might be associated with the establishment of a trans-Eurasian exchange/migration network through the steppe zone, influenced by the more intensive exploitation of cannabis achenes popular in Eastern Europe pastoralist communities. The role of the Hexi Corridor region as a hub for an East Asian spread of domesticated plants, animals and cultural elements originally from Southwest Asia and Europe is highlighted. More systematic, interdisciplinary and well-dated data, especially from South Russia and Central Asia, are necessary to address the unresolved issues in understanding the complex history of human cannabis utilisation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Cannabis is a versatile plant, used either as a medicament, hallucinogen, raw fibre material for ropes and textiles, or as a food source (seeds and oil) (Warf 2014). It is regarded as one of the world’s oldest cultivated plants (Russo 2007) and an element of the ethnological myths and social identity of many traditional societies (e.g. Ullah et al. 2013). Despite a large volume of information on the botany, agriculture or medical application of the plant (e.g. Borgelt et al. 2013), the history of cannabis remains poorly understood.
There are different hypotheses on the origin of cannabis domestication, of which the two most frequently cited either refer to China (e.g. Chang 1986; Crawford 2006) or Central Asia (e.g. Vavilov 1992) as the domestication centre. The China hypothesis is largely based on early written records (Li 1974b), whereas the Central Asia hypothesis considers Central Asia as a biodiversity centre of cannabis and thus a possible domestication centre. However, both hypotheses lack a systematic test based on available archaeological evidence. That the domestication centre of a crop does not necessarily match its modern centre of biodiversity has been documented in earlier studies. For example, the Yunnan-Assam region (Fig. 1), the modern biodiversity centre of Asian rice (Oryza sativa) (Zeng et al. 2007), was for a long time believed to be the domestication centre of rice (Chang 1976). However, this hypothesis has been largely refuted by archaeological evidence for domestication activities from the middle and lower reaches of the Yangtze River (Fuller 2011). Thus, a systematic compilation of early records of cannabis becomes an important and reasonable approach in response to a growing interest in understanding cannabis and its early domestication history (e.g. Booth 2003; Clarke and Merlin 2013; Small 2015).

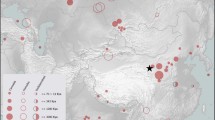
Map of sites/areas or cultures in Eurasia with early (older than ca. 3,000 cal bp) cannabis remains; for numbers see Table 1. I—heartland of ancient East Asian civilisation, II—heartland of ancient South Asian civilisation, III—heartland of ancient Southwest Asian civilisation, IV—heartland of mid-latitude Eurasia. A—possible homeland of the Yamnaya Culture, B—possible homeland of the Scythians, C—possible homeland of the Botai Culture, D—Altai Mountains region, E—homeland of the Xiongnu tribes, F—Hexi Corridor region, G—Yunnan-Assam region, H—Xinjiang. The historic Silk Road and the Bronze Age trans-Eurasian exchange/migration route are shown, respectively, in solid and dashed lines. Equidistant Cylindrical Projection is used for the map
This paper reviews archaeological and palaeoenvironmental case studies related to early cannabis remains in Eurasia, in order to trace the origin of its use by humans and to shed further light on related environmental and cultural contexts. These remains mainly include four different types: fibres, pollen, achenes or imprints of achenes on archaeological artefacts. We follow Small (2015, p. 191) using ‘cannabis’ as a vernacular abstraction when referring to the plant in general and ‘Cannabis’ as the genus name when referring to the binomial nomenclature.
Botany and ecology of cannabis
Cannabis plants are generally considered as one genus, Cannabis, family Cannabaceae, order Urticales (Kuddus et al. 2013). Previously, plants of Cannabaceae were included in the family of Moraceae or Urticaceae, but now are considered as taxa of a separate family based upon accumulating genetic and morphological evidence (Gautam et al. 2013). Cannabis and Humulus are the two acknowledged genera of Cannabaceae, although ongoing studies suggest moving some subfamilies previously under Ulmaceae to Cannabaceae (e.g. Yang et al. 2013).
The taxonomic differentiation within the genus Cannabis is under ongoing debate (Laursen 2015). Based on genetic variations, some authors propose a multitypic genus with at least two putative species, C. sativa and C. indica (possibly supplemented by C. ruderalis and C. chinensis) (e.g. Hillig 2005; Salentijn et al. 2015), while other authors propose a single species C. sativa with the genetic variation explained by differences on subspecies- and variety-levels or on a biotype-level of putative taxa (e.g. de Meijer et al. 2003). Both proposals, however, reveal problems in excluding hybridisation of different taxa and/or determining an adequate size of sample (Small 2015). Notwithstanding the difficulty in nomenclature or in defining species, genetic differentiation is almost certain between the hemp-type fibre cannabis (low in the principal psychoactive constituent delta-9-tetrahydrocannabinol (THC) and mostly used for fibre production) and marijuana-type cannabis (rich in THC and mostly used for drug intoxication) and between European hemp and East Asian hemp (Hillig 2005).
Cannabis is an annual herb growing during the warm season (Raman 1998). The plant is heliotropic and flowering is critically controlled by the length of the photoperiod (Booth 2003). Growing on almost any type of soil, it is most productive on nutrient-rich ground (Angelini et al. 2014). As a nitrophilous plant (Small et al. 2003), cannabis is closely associated with intentionally or unintentionally manured soils near human settlements subjected to organic wastes (Clarke and Merlin 2013). In general, this ruderal plant is considered as one of man’s camp followers in prehistoric time, appearing quickly along roadsides, in dump heaps, and/or on the edges of fields after establishment of settlements (Schultes et al. 1974).
Data sources and criteria of evaluation
A list of early cannabis records was compiled from published sources specifying the location where the remains were recovered, types of remains, and their approximate age. Radiocarbon dates were directly cited with the determined age, while relative ages (e.g. associated with an archaeological culture) referred to the chronology of the given culture. All age information was harmonised with the unit of calendar years before present (cal bp), where ‘present’ is conventionally taken as ad 1950.
Only records having an age older than ca. 3,000 cal bp were selected for our database. The critical value of 3,000 cal bp was chosen because the period between ca. 3,200 and 2,500 cal bp marks the approximate start of the Iron Age and documented contacts across Eurasia (Smith 2014). The pre-Iron Age millennia are of high relevance for understanding the origin of cannabis utilisation. Accordingly, the records were grouped into time intervals: >ca. 8,000 cal bp, ca. 8,000–7,000 cal bp, ca. 7,000–6,000 cal bp, ca. 6,000–5,000 cal bp, ca. 5,000–4,000 cal bp and ca. 4,000–3,000 cal bp. In the compilation process, we followed data interpretation in the original references. For example, pollen from the Banpo Site in China, which was exclusively assigned to Humulus in the original archaeological report (Zhou 1963), but hypothetically interpreted as Cannabis in a widely cited secondary reference (Li 1974a). In this case we did not accept the interpretation of the secondary reference in our database.
We separated four geographical regions in Eurasia to trace the ancient spatial distribution of the plant. These are East Asia (including China, North Korea, South Korea and Japan), South Asia (including India, Pakistan, Nepal, Bangladesh, Bhutan and Sri Lanka), Southwest Asia (including Iraq, Syria, Lebanon, Turkey, Jordan, Georgia, Azerbaijan, Iran, Armenia and Azerbaijan) and mid-latitude Eurasia (including Europe, Russia, Mongolia, Kazakhstan, Kyrgyzstan, Tajikistan, Uzbekistan and Afghanistan).
Four major types of cannabis remains were distinguished in the database: cannabis achenes, hemp fibres (e.g. for ropes or textiles), pollen assigned to Cannabis-type, and imprints of achenes on archaeological artefacts. The differentiation allows consideration of the relative merits and weaknesses of these different remains in indicating human use (see Fleming and Clarke 1998). In general, cannabis achenes can be accurately identified to the genus level, so achenes recovered from an archaeological context were tentatively considered as potential evidence for human exploitation of cannabis achenes at the site (Bouby 2002). Hemp fibres directly indicate use of cannabis (Ryder 1999), but ambiguity exists in accurately distinguishing these remains from other fibre remains (especially after several millennia of degradation) (Haugan and Holst 2014). Imprints of cannabis achenes on ceramics are evidence for a more intensive human use of cannabis achenes than a few sieved cannabis grains obtained by floatation. However, cord imprints on ceramics with linear decorative patterns that are often assigned to hemp were not accepted in the database, because distinguishing different fibre traces based only on morphological features is particularly unreliable (Bergfjord and Holst 2010).
Lastly, appearance of pollen assigned to Cannabis is seen as an indicator of local growth of cannabis plants near the study site (Godwin 1967), though pollen grains can be transported over long distances in certain circumstances (Moore et al. 1991). The fact that Humulus pollen morphologically resembles that of Cannabis may lead to erroneous identification, although some authors (Dörfler 1990; Fleming and Clarke 1998; Mercuri et al. 2002; Punt and Malotaux 1984) propose criteria for distinguishing between the two. Furthermore, pollen records do not explicitly allow the identification of past human use of cannabis, since plants might have grown spontaneously. However, if a sudden and strong increase in percentages of cannabis pollen is observed in a pollen diagram, the increase is more likely to be due to human activities, such as retting hemp for fibre production (Demske et al. 2016; Lavrieux et al. 2013; Mercuri et al. 2002). A unique advantage of palynological evidence of anemophilous cannabis is the often excellent preservation of its pollen grains in sediment records. Preservation of macrofossils in sediments, in general, is less often recorded (Birks 2001).
Results
Table 1 lists published cannabis records (older than ca. 3,000 cal bp) across Eurasia while Fig. 1 illustrates their location, with records of different ages indicated with different symbols. Records associated with an estimated age range were indicated according to the upper (older) boundary of that age range.
Eastern Europe, notably the region north and west of the Black Sea, has a high frequency of records of fossil Cannabis-type pollen (site numbers 40, 41, 42, 43, 44, 45, 46 and 48) older than ca. 8,000 cal bp, and at least one achene record (site number 51) of the same age. Western Europe and Central Europe has five pollen and achene records (site numbers 47, 49, 50, 52 and 53) older than ca. 8,000 cal bp. A record of achenes (site number 1) also appears in East Asia from ca. 10,000 cal bp.
A continuous series of achene or ceramic imprint records is found in Europe from ca. 8,000–7,000 cal bp (site numbers 56 and 58) through ca. 7,000–6,000 cal bp (site number 64) to ca. 6,000–5,000 cal bp (site numbers 66, 70 and 71). In East Asia, there are also some records of cannabis achenes (site numbers 4, 5 and 8) but also hemp fibre (site numbers 2 and 6) throughout the period between ca. 7,000 and 5,000 cal bp. A marked increase in achene records, however, only takes place in the millennium ca. 5,000–4,000 cal bp. From a total of eight sites or site clusters in East Asia (site numbers 9, 10, 11, 13, 15, 16, 17 and 18) cannabis achenes dating to this millennium were reported.
Outside of Europe and East Asia, records are scattered and appear relatively late in time. Southwest Asia has two geographically isolated records of hemp fibre (site numbers 36 and 38), of which one dates to ca. 6,000–5,000 cal bp and the other to ca. 5,000–4,000 cal bp, and only one record of cannabis achenes (site number 39) dating to ca. 4,000–3,000 cal bp. Cannabis records did not appear in records from South Asia until ca. 4,000–3,000 cal bp.
Analysis and discussion
Scythians in the mid-fifth century bc (ca. 2,500 cal bp) were described by the contemporaneous Greek historian Herodotus as consuming cannabis by inhaling its smoke in rituals (Warf 2014). Scythians was a general term at that time referring to nomads who inhabited the area stretching from present-day Ukraine to Central Asia (González-Ruiz et al. 2012). Herodotus’ record seems to be amongst the earliest texts from Europe describing the use of cannabis (Wills 1998). Excavations carried out in the eastern part of Scythia (i.e. Central Asia) provide evidence supporting his description. Archaeological sites such as Pazyryk Frozen Tombs in the Altai Mountains (Fig. 1; van Noten and Polosmak 1995) and Iron Age burial sites at Yanghai in the Turpan Depression (Fig. 2; Jiang et al. 2006) recovered fur bags or bowls containing cannabis remains dated to ca. 2,500 cal bp. Being of similar age, the earliest written Chinese record concerning cannabis growth and use in the eastern part of Eurasia can be found in the Shi Jing, a collection of poems finalised in the sixth century bc (ca. 2,600 cal bp) (Li 1974a). Texts from Southwest Asia, South Asia and Egypt, quoted as 3,500–4,000 years old (Russo 2007), were suggested as even earlier records of medical uses of cannabis, but the reliability of these records is questionable (Wills 1998).

Temporal and spatial distribution of cannabis achenes (ca. 5,000–2,000 cal bp) and wheat (ca. 5,000–3,000 cal bp) records in North China and adjacent regions. Sites with cannabis remains: 1 Haminmangha, 2 Yanggua, 3 Buziping, 4 Linjia, 5 Erdaojingzi, 6 Shangtaizi, 7 Jinchankou, 8 Lajia, 9 Ejia, Xinjia, Qingquanhantai, 10 Daxinzhuang, 11 Erfang, Lamafeng, Baojia, Wenjiagou, Zhongchuannongchang, 12 Yanghai. Information about sites 1-11 can be found in Table 1 and Fig. 1, and information about site 12 can be found in Jiang et al. (2006). Sites with early wheat remains: 13 Donghuishan, 14 Xishanping, 15 Zhouyuan, 16 Zhaojialai, 17 Jiaochangpu, 18 Liangchengzhen, 19 Zhaojiazhuang, 20 Huoshiliang, 21 Ganggangwa, 22 Xiaohe, 23 Gumugou, 24 Luanzagangzi, 25 Sidaogou, 26 Xintala, 27 Wupaer. The sites with wheat remains are based on Betts et al. (2014). Possible Bronze Age trans-Eurasian exchange/migration routes through the Eurasian steppe and possible dispersal directions of western (Southwest Asia and Europe) cultural elements within China are shown in dashed lines and arrowed lines, respectively. The topographic base map was generated from ETOPO1 Global Relief Model data (Amante and Eakins 2009). National boundaries were drawn from Natural Earth vector data v2.0 (http://www.naturalearthdata.com). Equidistant Cylindrical Projection is used for the map, in which the scale is true along the meridians
According to the collected data, all these written records are late reflections of a much earlier tradition of cannabis use across Eurasia, dating back to the early Holocene or even earlier periods. On one hand, cannabis seems to have grown as a component of natural vegetation across the entire continent at least since the early Holocene, represented by pollen and achene records appearing older than ca. 8,000 cal bp from both Europe and East Asia, a plant resource ready to be used by humans. The scarcity of cannabis pollen records predating ca. 8,000 cal bp from East Asia and other regions outside Europe might be due to the fact that Europe has been more thoroughly studied through pollen analysis (Grimm et al. 2007) and that the genus Cannabis was placed in different taxonomic groups depending on the flora to which the authors referred. For example, in previous Chinese standard floras (Editorial Board of ‘Flora of China’ of Chinese Academy of Sciences 1959–2004), Cannabis was listed as a genus of the Moraceae family (sensu Engler). Accordingly, some palynologists in China enumerated Cannabis pollen as unspecified Moraceae-type pollen (Wang et al. 1995). On the other hand, the three archaeological sites (site numbers 1, 52 and 53) with achene remains provide more direct evidence of human use of cannabis in both Europe and East Asia during the early Holocene. In light of the availability of the cannabis plant resource and contemporaneous records of cannabis achenes in the two distal parts of the continent, a multiregional origin of cannabis exploitation in Eurasia is, at least on a Holocene scale, more likely than a single-region origin (e.g. China) proposed in some conventional hypotheses. At least two centres, Europe and East Asia, appear as origins based on our compiled dataset. Prior to the Holocene, however, a more constrained centre of natural distribution of Cannabis remains possible.
In the cannabis exploitation history of Europe, there seems to be a long tradition of using cannabis achenes, which is reflected by the continuous series of achene or ceramic imprint records in combination with a few hemp fibre records. How achenes were exploited by prehistoric people cannot be clarified based on the present dataset. Archaeologists suggest that the practice of inhaling cannabis smoke might have appeared in Eastern Europe during the Yamnaya Culture (ca. 5,500/5,300–4,500/4,300 cal bp; Milisauskas and Kruk 2011; Anthony 2007) or even earlier (Balter and Gibbons 2015). The Yamnaya Culture originally occupied the region north of the Black Sea and is thought to be one of the most likely candidates for suggested Proto-Indo-European ancestors (Haak et al. 2015). Their westward migration is believed to have had influences on large parts of Europe linguistically and genetically (Haak et al. 2015). It is suggested that at this time cannabis was one of the key trade goods in the East European steppe and that smoking for psychoactive purposes became a ritual practice (Sherratt 2003). While achenes are not highly psychoactive, bracts attached to them contain the highest content of THC (Jiang et al. 2006). Thus, achene finds can be associated with the consumption of cannabis as a hallucinogen, as e.g. shown in a study of an Iron Age tomb in Central Asia (Russo et al. 2008). The westward migration of the Yamnaya people further spread the practice of cannabis smoking in Europe. One example for this is the record of a clay vessel with carbonised achenes and signs of cannabis burning from a tomb in Romania (site number 71).
Although more or less continuous cannabis records are reported from East Asia prior to ca. 5,000 cal bp, they are scattered in space and time. The marked increase in cannabis records in East Asia, notably achene records, between ca. 5,000 and 4,000 cal bp, might indicate a change in utilisation at that time. At least in one (Linjia, site number 13) of the eight sites or site clusters from which cannabis records date to this age, achenes appear to have been of high value as they are found to have been collectively stored in a clay pot (Institute of Botany in Northwest Normal University and Gansu Provincial Museum 1984). All the eight sites or site clusters are located in western or northern borderlands of China that are at similar distances from the eastern part of the Eurasian steppe and from the ancient (agri-)cultural centres in East Asia (the middle and lower reaches of the Yangtze and Yellow Rivers) (e.g. Zhao 2011), in particular the Hexi Corridor region and Northeast China. In the Hexi Corridor region, the oldest remains of wheat (Triticum spp.) in China were also found. This crop is considered to be of Southwest Asian origin and was first introduced to East Asia around ca. 5,000 cal bp or immediately after. Betts et al. (2014) cite an age of directly-dated carbonised wheat grains at the Donghuishan Site (Hexi Corridor region) as old as ca. 5,300–3,900 cal bp. Fig. 2 shows records of earliest wheat from northern China as compiled in Betts et al. (2014) and cannabis achene remains dating to ca. 5,000–2,000 cal bp. The radial distance from the Hexi Corridor region of early cannabis and wheat records is increasing over time.
A possible explanation for such a distribution pattern is that the Hexi Corridor region might have acted as a gateway through which the ancient exchange/migration network in Eurasia stretched to the heartland of the Chinese civilisation located further southeast. Although the Hexi Corridor was an easy passage later followed by the historical Silk Road, which accounted for much of the trade in Eurasian written history, the Silk Road itself was not firmly established until the second century bc (ca. 2,200 cal bp), when the Chinese Emperor Han Wu attempted to defeat the nomadic Xiongnu (living in the eastern part of the Eurasian steppe) through allying with political powers and seeking to obtain horses in Central Asia (Xie et al. 2007). However, available archaeological evidence (Warburton 2011) supports the hypothesis that connections between East Asia and Southwest Asia/Europe were already established since the late Neolithic or the early Bronze Age (e.g. Anthony 2007). The third millennium bc (ca. 5,000–4,000 cal bp) seems to have been the key period for firm establishment of this connection. Pastoralists, particularly the Yamnaya and their eastern neighbours (e.g. the Botai), living on the Eurasian steppe at that time, played an important role in this exchange/migration network. It is believed that these people domesticated wild horses (Equus ferus) (Levine 1999), probably from ca. 6,000 and 5,000 cal bp (Bendrey 2014), and were able to travel long distances across the relatively flat steppe zone (Frachetti 2011). It might have been this domesticated horse-based mobility—much enhanced compared to other agricultural societies located in the southern part of the continent—that enabled these populations to become a key promoter in the early exchange/migration network connecting far distant parts of the continent (Boivin et al. 2012), comparable to the merchants of the Silk Road several millennia later. Recent genetic evidence suggests that the Yamnaya population not only spread westwards to the heartland of Europe, but also migrated to Central Asia (e.g. Altai region) and later to Mongolia and northern China, suggesting a dynamic mobility throughout the entire Eurasian steppe zone during the Bronze Age (Allentoft et al. 2015). This mobility might have also facilitated the spread of bronze-making technology throughout Eurasia, contributing to the formation of the Bronze Age World System across the continent and to fundamental societal changes towards the end of the third millennium bc (ca. 5,000–4,000 cal bp) (Warburton 2011). In China, the utilisation of bronze, horses and new cereal crops, probably introduced via the network, changed the mode of warfare, which resulted in the establishment of the first dynastic states in the middle Yellow River region (Yi 2014). Interestingly, the first appearance of bronze artefacts in China was also in the Hexi Corridor region: the Qijia Culture, which was a representative archaeological culture in the region at the time, is famous for its early bronze mirrors, knives, rings and spears (Yi 2014). Also, the first bone remains of the domesticated form of horse (Equus ferus ssp. caballus) in China are associated with Qijia Culture sites (Yuan 2010).
The above-mentioned ancient exchange network is termed the ‘Bronze Road’ by some authors (e.g. Liu and Li 2014 p. 79), as bronze might have been a key object of trade. Nonetheless, the transfer of plants and plant utilisation techniques may also have played an important role in this trade network (Lightfoot et al. 2013). The multiple usability of cannabis makes it an ideal candidate for being a ‘cash crop before cash’, a plant that is cultivated primarily for exchange purposes (Sherratt 1999, p. 16), even if the ritual obsession of cannabis smoking in East Europe and the potential role of hemp-made rope in the first horse reins (Merlin 2003) are not emphasized. The marked increase in cannabis achene remains at ca. 5,000–4,000 cal bp (site numbers 9, 10, 11, 13, 15, 16, 17 and 18) in distal parts of East Asia, in particular the Hexi Corridor and Northeast China, may be interpreted as evidence for the increased importance of the plant in the exchange network established during the early Bronze Age. Firm archaeobotanical evidence of cannabis appearing at archaeological sites in areas such as South Asia (e.g. site number 34) during ca. 4,000–3,000 cal bp (e.g. Stevens et al. 2016) could be associated with a further expansion of the network.
This ancient exchange/migration network might have passed largely through the Eurasian steppe in the earliest stage, differing from the historical Silk Road to the south (Christian 2000) (Fig. 1). The difference could partly be due to the lower degree of landscape complexity in the steppe zone. Extreme environments (i.e. arid deserts and precipitous mountainous terrain) along the Silk Road (Fig. 2) could have hindered early exchange activities of people who were probably less well equipped compared with their later counterparts of the historical period (Liu and Li 2014). The steppe route of the Bronze Road (Christian 2000) might also explain the relatively late appearance of cannabis remains (from ca. 3,000–2,000 cal bp) (e.g. Jiang et al. 2006) and Southwest Asian crops such as wheat (from ca. 4,000–3,000 cal bp) (e.g. Dodson et al. 2013) in Xinjiang (Fig. 1). This indicates that the Hexi Corridor region was probably the central hub in China from which introduced plants and cultural elements from Southwest Asia/Europe were further diffused south-eastwards and westwards.
The fact that traits like smoking cannabis or using its achenes as funerary objects are closely related to Indo-European cultures may also facilitate the reconstruction of migrations of Proto-Indo-European populations. The Scythians were likely an Indo-European population (Ning et al. 2015). There is strong evidence of ritual/medicinal use of cannabis dating to ca. 2,500 cal bp from the burial site at Yanghai, Tarim Basin, Northwest China (Jiang et al. 2006). Several anatomical features of the skulls recovered from the site show more similarities to those of European populations than to East Asian features (Yang et al. 2010). Even clearer European anatomical and genetic features are observed in populations in the Tarim Basin dating to ca. 4,000 cal bp (Li et al. 2010), for example at the Xiaohe and Gumugou burial sites (Fig. 2). These people possibly spoke Tocharian (extinct today), which was a western branch of the Indo-European language family (Jia and Betts 2010; Li and Li 2011). Hypothesizing that ritual burning of cannabis was an important Indo-European tradition, cannabis remains may be regarded as a valuable palaeobotanical proxy for the reconstruction of migration activities of Proto-Indo-European people across Eurasia, which are still poorly understood (Bouckaert et al. 2012).
We found only three spatially scattered cannabis records (site numbers 62, 67 and 70) from the large area of South Russia and Central Asia published in international literature. The scarcity is partly due to the current focus in archaeobotany on regions such as Europe and East Asia, leaving large parts of the Eurasian steppe and adjacent regions to its south understudied (Outram et al. 2012). This outlines the need for more high-resolution, interdisciplinary and well-dated data from the Eurasian steppe to strengthen or to falsify our hypothesis that cannabis was closely associated with the Bronze Age trans-continental exchange. One promising method in studying ancient cannabis remains appears to be the analysis of phytochemical and genetic features, as it may allow for identifying the genetic type of the remains and their possible utilisation (Russo et al. 2008). The results can be correlated with modern phylogenetic-biogeographical relations of different putative taxa of cannabis.
Conclusion
In light of archaeobotanical and palynological records of cannabis available to date, the history of human use of this plant remains a complex and unresolved issue. More well-dated data, especially from the Eurasian steppe zone, are called for to address open questions. Existing evidence, however, suggests that a multiregional origin of cannabis utilisation in different parts of Eurasia is more likely than a single-region origin as argued for in some conventional hypotheses. It appears that at least Europe and East Asia were two independent centres of early cannabis use. Additional domestication centres cannot be ruled out, since other regions in Eurasia still lack systematic archaeobotanical investigations. There seems to be a continuous emphasis on exploiting cannabis achenes in Europe from the early Holocene. This practice might have also influenced how cannabis was used in East Asia since the onset of the Bronze Age exchange/migration network across Eurasia (ca. 5,000–4,000 cal bp).
References
Allentoft ME, Sikora M, Sjögren K-G et al (2015) Population genomics of Bronze Age Eurasia. Nature 522:167–172
Amante C, Eakins BW (2009) ETOPO1 1 Arc-minute Global Relief Model: Procedures, Data Sources and Analysis: NOAA Technical Memorandum NESDIS NGDC-24. National Geophysical Data Center, National Oceanic and Atmospheric Administration, Boulder, USA
Andrič M (2007) Holocene vegetation development in Bela krajina (Slovenia) and the impact of first farmers on the landscape. Holocene 17:763–776
Angelini LG, Tavarini S, Cestone B, Beni C (2014) Variation in mineral composition in three different plant organs of five fibre hemp (Cannabis sativa L.) cultivars. Agrochimica 58:1–18
Anthony DW (2007) The Horse, the Wheel, and Language: How Bronze-Age Riders from the Eurasian Steppes Shaped the Modern World. Princeton University Press, Princeton, USA
Balter M, Gibbons A (2015) Indo-European languages tied to herders. Science 347(6224):814–815
Barber EJW (1991) Prehistoric textiles: The development of cloth in the Neolithic and Bronze Ages with special reference to the Aegean. Princeton University Press, Princeton
Bendrey R (2014) Population genetics, biogeography, and domestic horse origins and diffusions. J Biogeog 41:1,441–1,442
Bergfjord C, Holst B (2010) A procedure for identifying textile bast fibres using microscopy: flax, nettle/ramie, hemp and jute. Ultramicroscopy 110:1,192–1,197
Berthel N, Schwörer C, Tinner W (2012) Impact of Holocene climate changes on alpine and treeline vegetation at Sanetsch Pass, Bernese Alps, Switzerland. Rev Palaeobot Palynol 174:91–100
Betts A, Jia PW, Dodson J (2014) The origins of wheat in China and potential pathways for its introduction: a review. Quat Int 348:158–168
Birks HH (2001) Plant Macrofossils. In: Smol JP, Birks HJB, Last WM (eds) Tracking Environmental Change Using Lake Sediments, vol 3, Terrestrial, algal, and siliceous indicators. Kluwer, Dordrecht, pp 49–74
Blyakharchuk TA, Chernova NA (2013) Vegetation and climate in the Western Sayan Mts according to pollen data from Lugovoe Mire as a background for prehistoric cultural change in southern Middle Siberia. Quat Sci Rev 75:22–42
Bodnariuc A, Bouchette A, Dedoubat JJ, Otto T, Fontugne M, Jalut G (2002) Holocene vegetational history of the Apuseni mountains, central Romania. Quat Sci Rev 21:1,465–1,488
Boivin N, Fuller DQ, Crowther A (2012) Old World globalization and the Columbian exchange: comparison and contrast. World Archaeol 44:452–469
Booth M (2003) Cannabis: A history. St. Martin’s Press, New York, USA
Borgelt LM, Franson KL, Nussbaum AM, Wang GS (2013) The pharmacologic and clinical effects of medical cannabis. Pharmacotherapy 33:195–209
Bouby L (2002) Le chanvre (Cannabis sativa L.): une plante cultivée à la fin de l’âge du Fer en France du Sud-Ouest ? C R Palevol 1:89–95
Bouckaert R, Lemey P, Dunn M et al (2012) Mapping the origins and expansion of the Indo-European language family. Science 337:957–960
Chang K-c (1986) The Archaeology of Ancient China, 4th edn. Yale University Press, New Haven, USA
Chang T-T (1976) The origin, evolution, cultivation, dissemination, and diversification of Asian and African rices. Euphytica 25:425–441
Chaussé C, Leroyer C, Girardclos O, Allenet G, Pion P, Raymond P (2008) Holocene history of the River Seine, Paris, France: bio-chronostratigraphic and geomorphological evidence from the Quai-Branly. Holocene 18:967–980
Chen X (2007) Paleoethnobotany and agriculture across the transition from the Late Neolithic to the Bronze Age in Northeastern China: A case study. Ph.D. Thesis, Shandong University
Christian D (2000) Silk Roads or Steppe Roads? The Silk Roads in world history. J World Hist 11:1–26
Clarke RC (2006) Hemp (Cannabis) cultivation and use in the Republic of Korea. J Ind Hemp 11:51–86
Clarke RC, Merlin MD (2013) Cannabis: Evolution and ethnobotany. University of California Press, Berkeley, USA
Connor SE, Thomas I, Kvavadze EV (2007) A 5600-yr history of changing vegetation, sea levels and human impacts from the Black Sea coast of Georgia. Holocene 17:25–36
Crawford GW (2006) East Asian plant domestication. In: Stark MT (ed) Archaeology of Asia. Blackwell, Oxford, pp 77–95
Crawford GW (2011) Advances in understanding early agriculture in Japan. Curr Anthropol 52(S4):S331–S345
Crawford GW, Takamiya H (1990) The origins and implications of late prehistoric plant husbandry in northern Japan. Antiquity 64(245):889–911
Cyprien A-L, Visset L, Carcaud N (2004) Evolution of vegetation landscapes during the Holocene in the central and downstream Loire basin (Western France). Veget Hist Archaeobot 13:181–196
D’Andrea AC (1995) Later Jomon subsistence in Northeastern Japan: new evidence from palaeoethnobotanical studies. Asian Perspect 34:195–227
de Meijer EPM, Bagatta M, Carboni A, Crucitti P, Moliterni VMC, Ranalli P, Mandolino G (2003) The inheritance of chemical phenotype in Cannabis sativa L. Genetics 163:335–346
Demske D, Tarasov PE, Leipe C, Kotlia BS, Joshi LM, Long T (2016) Record of vegetation, climate change, human impact, and retting of hemp in Garhwal Himalaya (India) during the past 4600 years. Holocene. doi:10.1177/0959683616650267
Dodson JR, Li X, Zhou X, Zhao K, Sun N, Atahan P (2013) Origin and spread of wheat in China. Quat Sci Rev 72:108–111
Dörfler W (1990) Die Geschichte des Hanfanbaus in Mitteleuropa aufgrund palynologischer Untersuchungen und von Großrestnachweisen. Prähist Z 65:218–244
Ecsedy I (1979) The people of the pit-grave Kurgans in Eastern Hungary. Adademia Kiado, Budapest, Hungary
Editorial Board of ‘Flora of China’ of Chinese Academy of Sciences (1959–2004) Flora of China. Science Press, Beijing, China
Edwards KJ, Whittington G (2000) Multiple charcoal profiles in a Scottish lake: taphonomy, fire ecology, human impact and inference. Palaeogeog Palaeoclimatol Palaeoecol 164:67–86
Fărcaş S, Tanţău I, Mîndrescu M, Hurdu B (2013) Holocene vegetation history in the Maramureş Mountains (Northern Romanian Carpathians). Quat Int 293:92–104
Fernane A, Penaud A, Gandouin E, Goslin J, Van Vliet-Lanoë B, Vidal M (2015) Climate variability and storm impacts as major drivers for human coastal marsh withdrawal over the Neolithic period (Southern Brittany, NW France). Palaeogeog Palaeoclimatol Palaeoecol 435:136–144
Feurdean A, Astalos C (2005) The impact of human activities in the Gutaiului Mountains, Romania. Stud Univ Babeş-Bolyai Geol 50:63–72
Feurdean A, Parr CL, Tanţău I, Fărcaş S, Marinova E, Perşoiu I (2013) Biodiversity variability across elevations in the Carpathians: parallel change with landscape openness and land use. Holocene 23:869–881
Filipova-Marinova M, Pavlov D, Coolen M, Giosan L (2013) First high-resolution marinopalynological stratigraphy of Late Quaternary sediments from the central part of the Bulgarian Black Sea area. Quat Int 293:170–183
Filipova-Marinova M, Pavlov D, Giosan L (2015) Multi-proxy records of Holocene palaeoenvironmental changes in the Varna Lake area, western Black Sea coast. Quat Int 401:99–108
Fleming MP, Clarke RC (1998) Physical evidence for the antiquity of Cannabis sativa L. J Int Hemp Assoc 5:80–92
Frachetti MD (2011) Migration concepts in Central Eurasian archaeology. Annu Rev Anthropol 40:195–212
Fuller DQ (2008) The spread of textile production and textile crops in India beyond the Harappan zone: an aspect of the emergence of craft specialization and systematic trade. In: Osada T, Uesugi A (eds) Occasional paper 3: Linguistics, archaeology and the human past. Indus Project, Research Institute for Humanity and Nature, Kyoto, Japan, pp 1–26
Fuller DQ (2011) Pathways to Asian civilizations: tracing the origins and spread of rice and rice cultures. Rice 4:78–92
Fuller DQ, Boivin N (2009) Crops, cattle and commensals across the Indian Ocean: current and potential archaeobiological evidence. Études Océan Indien 42–43:13–46
Gao H, Wang R, Chen Y (1979) Textiles excavated from Taixicun Site (Shang Dynasty). Cult Relics 6:44–48
Gautam AK, Kant M, Thakur Y (2013) Isolation of endophytic fungi from Cannabis sativa and study their antifungal potential. Arch Phytopathol Plant Prot 46:627–635
Geantă A, Gałka M, Tanţău I, Hutchinson SM, Mîndrescu M, Feurdean A (2014) High mountain region of the Northern Romanian Carpathians responded sensitively to Holocene climate and land use changes: a multi-proxy analysis. Holocene 24:944–956
Godwin H (1967) Pollen-analytic evidence for the cultivation of Cannabis in England. Rev Palaeobot Palynol 4:71–80
González-Ruiz M, Santos C, Jordana X et al (2012) Tracing the origin of the East-West population admixture in the Altai Region (Central Asia). PLOS One 7(11):1–11
Grimm EC, Keltner J, Cheddadi R, Hicks S, Lézine A-M, Berrio JC, Williams JW (2007) Pollen methods and studies / Databases and their application. In: Elias SA (ed) Encyclopedia of Quaternary Science. Elsevier, Amsterdam, pp 2521–2528
Gronenborn D (2003) Migration, acculturation and culture change in western temperate Eurasia, 6500–5000 cal bc. Doc Praehist 30:79–91
Haak W, Lazaridis I, Patterson N et al (2015) Massive migration from the steppe was a source for Indo-European languages in Europe. Nature 522:207–211
Haugan E, Holst B (2014) Flax look-alikes: pitfalls of ancient plant fibre identification. Archaeometry 56:951–960
Hillig KW (2005) Genetic evidence for speciation in Cannabis (Cannabaceae). Genet Resour Crop Evol 52:161–180
Hörnberg G, Bohlin E, Hellberg E et al (2005) Effects of Mesolithic hunter-gatherers on local vegetation in a non-uniform glacio-isostatic land uplift area, northern Sweden. Veget Hist Archaeobot 15:13–26
Hörnberg G, Josefsson T, Bergman I, Liedgren L, Östlund L (2015) Indications of shifting cultivation west of the Lapland border: multifaceted land use in northernmost Sweden since AD 800. Holocene 25:989–1001
Hudson M, Barnes GL (1991) Yoshinogari. a Yayoi settlement in northern Kyushu. Monum Nippon 46:211–235
Innes JB, Zong Y, Wang Z, Chen Z (2014) Climatic and palaeoecological changes during the mid- to Late Holocene transition in eastern China: high-resolution pollen and non-pollen palynomorph analysis at Pingwang, Yangtze coastal lowlands. Quat Sci Rev 99:164–175
Institute of Botany in Northwest Normal University, Gansu Provincial Museum (1984) Millets and cannabis recovered from Linjia Site (Majiayao Culture), Gansu. Archaeology 7:654–655
Jia PWM, Betts AVG (2010) A re-analysis of the Qiemu’erqieke (Shamirshak) cemeteries, Xinjiang, China. J Indoeur Stud 38:275–317
Jia X, Dong G, Li H et al (2012) The development of agriculture and its impact on cultural expansion during the late Neolithic in the Western Loess Plateau, China. Holocene 23:85–92
Jiang H-E, Li X, Zhao Y-X et al (2006) A new insight into Cannabis sativa (Cannabaceae) utilization from 2500-year-old Yanghai Tombs, Xinjiang, China. J Ethnopharmacol 108:414–422
Kühn D (1987) Textile technology: spinning and reeling. In: Needham J (ed) Science and civilization in China, vol 5. Cambridge University Press, Cambridge, pp 865–867
Kabelik J, Krejci Z, Santavy F (1960) Cannabis as a medicament. Bull Narc 12:5–23
Kaltenrieder P, Procacci G, Vannière B, Tinner W (2010) Vegetation and fire history of the Euganean Hills (Colli Euganei) as recorded by Lateglacial and Holocene sedimentary series from Lago della Costa (northeastern Italy). Holocene 20:679–695
Krenke N, Erschov I, Erschova E, Lazukin A (2013) Corded ware, Fatjanovo and Abashevo culture sites on the flood-plain of the Moskva River. Spraw Archaeol 65:413–424
Kuddus M, Ginawi IAM, Al-Hazimi A (2013) Cannabis sativa: an ancient wild edible plant of India. Emir J Food Agric 25:736–745
Kvavadze E, Sagona A, Martkoplishvili I, Chichinadze M, Jalabadze M, Koridze I (2015) The hidden side of ritual: new palynological data from Early Bronze Age Georgia, the Southern Caucasus. J Archaeol Sci Rep 2:235–245
Latalowa M (1999) Palaeoecological reconstruction of the environmental conditions and economy in early medieval Wolin: against a background of the Holocene history of the landscape. Acta Palaeobot 39:183–271
Laursen L (2015) The cultivation of weed. Nature 525:S4–S5
Lavrieux M, Jacob J, Disnar J-R, Bréheret J-G, Milbeau CL, Miras Y, Andrieu-Ponel V (2013) Sedimentary cannabinol tracks the history of hemp retting. Geology 41:751–754
Lawler A (2009) Going the distance to uncover the roots of trade in the Near East. Science 324:717
Lawson IT, Al-Omari S, Tzedakis PC, Bryant CL, Christanis K (2005) Lateglacial and Holocene vegetation history at Nisi Fen and the Boras mountains, northern Greece. Holocene 15:873–887
Leipe C, Demske D, Tarasov PE, HIMPAC Project Members (2014) A Holocene pollen record from the northwestern Himalayan lake Tso Moriri: implications for palaeoclimatic and archaeological research. Quat Int 348:93–112
Levine MA (1999) Botai and the origins of horse domestication. J Anthropol Archaeol 18:29–78
Li C, Li H, Cui Y et al (2010) Evidence that a West-East admixed population lived in the Tarim Basin as early as the early Bronze Age. BMC Biol 8:1–12
Li H-L (1974a) An archaeological and historical account of cannabis in China. Econ Bot 28:437–448
Li H-L (1974b) The origin and use of cannabis in Eastern Asia linguistic-cultural implications. Econ Bot 28:293–301
Li S (1984) The ‘Boat Coffins’ of the Wuyi Mountains. In: Stockwell F, Bowen T (eds) Recent Discoveries in Chinese Archeology. Foreign Language Press, Beijing, China, pp 79–80
Li Y, Li B (2011) The role of Tocharian in establishing the relationship between Chinese and Indo-European languages. Linguist Sci 10:586–592
Lightfoot E, Liu X, Jones MK (2013) Why move starchy cereals? A review of the isotopic evidence for prehistoric millet consumption across Eurasia. World Archaeol 45:574–623
Liu X, Li W (2014) Pre-history “Bronze Road” and civilization of Central Plains. J Xinjiang Norm Univ (Philos Soc Sci) 35:79–88
Long T, Qin J, Atahan P, Mooney S, Taylor D (2014) Rising waters: new geoarchaeological evidence of inundation and early agriculture from former settlement sites on the southern Yangtze Delta, China. Holocene 24:546–558
Marinova E (2003) The new pollen core Lake Durankulak-3: a contribution to the vegetation history and human impact in Northeastern Bulgaria. In: Tonkov S (ed) Aspects of palynology and palaeoecology. Pensoft Publishers, Moscow, pp 279–288
Matsui A, Kanehara M (2006) The question of prehistoric plant husbandry during the Jomon Period in Japan. World Archaeol 38:259–273
Mercuri AM, Accorsi CA, Mazzanti MB (2002) The long history of Cannabis and its cultivation by the Romans in central Italy, shown by pollen records from Lago Albano and Lago di Nemi. Veget Hist Archaeobot 11:263–276
Mercuri AM, Accorsi CA, Mazzanti MB et al (2006) Economy and environment of Bronze Age settlements - Terramaras - on the Po Plain (Northern Italy): first results from the archaeobotanical research at the Terramara di Montale. Veget Hist Archaeobot 16:43–60
Mercuri AM, Mazzanti MB, Torri P et al (2012) A marine/terrestrial integration for mid-late Holocene vegetation history and the development of the cultural landscape in the Po Valley as a result of human impact and climate change. Veget Hist Archaeobot 21:353–372
Mercuri AM, Montecchi MC, Pellacani G, Florenzano A, Rattighieri E, Cardarelli A (2015) Environment, human impact and the role of trees on the Po Plain during the Middle and Recent Bronze Age: pollen evidence from the local influence of the terramare of Baggiovara and Casinalbo. Rev Palaeobot Palynol 218:231–249
Merlin MD (2003) Archaeological evidence for the tradition of psychoactive plant use in the Old World. Econ Bot 57:295–323
Milisauskas S, Kruk J (2011) Late Neolithic/Late Copper Age 3500–2200 bc. In: Milisauskas S (ed) European prehistory: A survey, 2nd edn. Springer, New York, pp 293–325
Miotk-Szpiganowicz G (1992) The history of the vegetation of Bory Tucholskie and the role of man in the light of palynological investigations. Acta Palaeobot 32:39–122
Moore PD, Webb JA, Collinson ME (1991) Pollen analysis, 2nd edn. Blackwell, Oxford
Murphy TM, Ben-Yehuda N, Taylor RE, Southon JR (2011) Hemp in ancient rope and fabric from the Christmas Cave in Israel: talmudic background and DNA sequence identification. J Archaeol Sci 38:2,579–2,588
Niinemets E, Saarse L (2009) Holocene vegetation and land-use dynamics of south-eastern Estonia. Quat Int 207:104–116
Ning C, Gao S, Deng B et al (2015) Ancient mitochondrial genome reveals trace of prehistoric migration in the east Pamir by pastoralists. J Hum Genet 61:103–108
Nunome J (1992) The archeology of fiber. Senshoku to Seikatsusha, Kyoto, Japan
Okazaki H, Kobayashi M, Momohara A et al (2011) Early Holocene coastal environment change inferred from deposits at Okinoshima archeological site, Boso Peninsula, central Japan. Quat Int:87–94
Outram AK, Kasparov A, Stear NA, Varfolomeev V, Usmanova E, Evershed RP (2012) Patterns of pastoralism in later Bronze Age Kazakhstan: new evidence from faunal and lipid residue analyses. J Archaeol Sci 39:2,424–2,435
Park J, Kim M, Lim HS, Choi J (2013) Pollen and sediment evidence for late-Holocene human impact at the Seonam-dong archeological site, Gwangju, Korea. Rev Palaeobot Palynol 193:110–118
Pashkevich G (2003) Palaeoethnobotanical evidence of agriculture in the steppe and the forest-steppe of East Europe in the Late Neolithic and Bronze Age. In: Levine M, Renfrew C, Boyle K (eds) Prehistoric steppe adaptation and the horse. McDonald Institute for Archaeological Research, Cambridge, pp 287–297
Pashkevych G (2012) Environment and economic activities of Neolithic and Bronze age populations of the Northern Pontic area. Quat Int 261:176–182
Poska A, Saarse L (2006) New evidence of possible crop introduction to north-eastern Europe during the Stone Age - Cerealia pollen finds in connection with the Akali Neolithic settlement, East Estonia. Veget Hist Archaeobot 15:169–179
Punt W, Malotaux M (1984) Cannabaceae, Moraceae and Urticaceae. Rev Palaeobot Palynol 42:23–44
Raman A (1998) The cannabis plant: botany, cultivation and processing for use. In: Brown DT (ed) Cannabis: The genus Cannabis. Harwood, Amsterdam, pp 29–54
Rasmussen P, Anderson NJ (2005) Natural and anthropogenic forcing of aquatic macrophyte development in a shallow Danish lake during the last 7000 years. J Biogeog 32:1,993–2,005
Russo EB (2007) History of cannabis and its preparations in saga, science, and sobriquet. Chem Biodivers 4:1,614–1,648
Russo EB, Jiang H-E, Li X et al (2008) Phytochemical and genetic analyses of ancient cannabis from Central Asia. J Exp Bot 59:4,171–4,182
Ryder ML (1999) Probable Fibres from Hemp (Cannabis sativa L.) in Bronze Age Scotland. Environ Archaeol 4:93–95
Salentijn EMJ, Zhang Q, Amaducci S, Yang M, Trindade LM (2015) New developments in fiber hemp (Cannabis sativa L.) breeding. Ind Crops Prod 68:32–41
Sarianidi V (1994) Temples of Bronze Age Margiana: traditions of ritual architecture. Antiquity 68(259):388–397
Schultes RE, Klein WM, Plowman T, Lockwood TE (1974) Cannabis: an example of taxonomic neglect. In: Rubin V (ed) Cannabis and culture. De Gruyter Mouton, den Hague, pp 21–38
Sherratt A (1999) Cash-crops before cash: organic consumables and trade. In: Gosden C, Hather J (eds) The prehistory of food: Appetites for change. Routledge, London, pp 12–32
Sherratt A (2003) The horse and the wheel: the dialectics of change in the circum-Pontic region and adjacent areas, 4500–1500 bc. In: Levine M, Renfrew C, Boyle K (eds) Prehistoric steppe adaptation and the Horse. McDonald Institute for Archaeological Research, Cambridge, pp 233–252
Sherratt AG (1991) Sacred and profane substances: the ritual use of narcotics in later Neolithic Europe. In: Sherratt A (ed) Economy and society in prehistoric Europe: Changing perspectives. Princeton University Press, Princeton, pp 403–430
Shin S-C, Rhee S-N, Aikens CM (2012) Chulmun Neolithic intensification, complexity, and emerging agriculture in Korea. Asian Perspect 51:68–109
Small E (2015) Evolution and classification of Cannabis sativa (marijuana, hemp) in relation to human utilization. Bot Rev 81:189–294
Small E, Pocock T, Cavers PB (2003) The biology of Canadian weeds. 119. Cannabis sativa L. Can J Plant Sci 83:217–237
Smith C (2014) Encyclopedia of global archaeology. Springer, New York
Stähli M, Finsinger W, Tinner W, Allgöwer B (2006) Wildfire history and fire ecology of the Swiss National Park (Central Alps): new evidence from charcoal, pollen and plant macrofossils. Holocene 16:805–817
Stančikaite M, Kabailiene M, Ostrauskas T, Guobyte R (2002) Environment and man around Lakes Duba and Pelesa, SE Lithuania, during the Late Glacial and Holocene. Geol Q 46:391–409
Stančikaitė M, Daugnora L, Hjelle K, Hufthammer AK (2009) The environment of the Neolithic archaeological sites in Šventoji, Western Lithuania. Quat Int 207:117–129
Stefanova I, Ammann B (2003) Lateglacial and Holocene vegetation belts in the Pirin Mountains (southwestern Bulgaria). Holocene 13:97–107
Stevens CJ, Murphy C, Roberts R, Lucas L, Silva F, Fuller DQ (2016) Between China and South Asia: a Middle Asian corridor of crop dispersal and agricultural innovation in the Bronze Age. Holocene. doi:10.1177/0959683616650268
Sun Y (2014) Research on plant remains from Neolithic to Early Bronze Age in Upper West Liao River region. Ph.D. Thesis, Inner Mongolia Normal University
Tarasov P, Jin G, Wagner M (2006) Mid-Holocene environmental and human dynamics in northeastern China reconstructed from pollen and archaeological data. Palaeogeog Palaeoclimatol Palaeoecol 241:284–300
Tonkov S, Panovska H, Possnert G, Bozilova E (2002) The Holocene vegetation history of Northern Pirin Mountain, southwestern Bulgaria: pollen analysis and radiocarbon dating of a core from Lake Ribno Banderishko. Holocene 12:201–210
Tonkov S, Bozilova E, Marinova E, Jüngner H (2008a) History of vegetation and landscape during the last 4000 years in the area of Straldzha mire (southeastern Bulgaria). Phytol Balc 14:185–191
Tonkov S, Bozilova E, Possnert G, Velčev A (2008b) A contribution to the postglacial vegetation history of the Rila Mountains, Bulgaria: the pollen record of Lake Trilistnika. Quat Int 190:58–70
Ullah M, Khan MU, Mahmood A et al (2013) An ethnobotanical survey of indigenous medicinal plants in Wana district south Waziristan agency, Pakistan. J Ethnopharmacol 150:918–924
Van Noten F, Polosmak N (1995) The frozen tombs of the Scythians. Endeavour 19:76–82
Vavilov NI (1992) Origin and geography of cultivated plants. Cambridge University Press, Cambridge
Wang F, Qian N, Zhang Y, Yang H (1995) Pollen flora of China, 2nd edn. Science Press, Beijing
Warburton DA (2011) What might the Bronze Age world-system look like? In: Wilkinson TC, Sherratt S, Bennet J (eds) Interweaving worlds: Systemic interactions in Eurasia, 7th to 1st Millennia bc. Oxbow Books, Oxford, pp 120–134
Warf B (2014) High points: an historical geography of cannabis. Geogr Rev 4:414–438
Wehrli M, Mitchell EAD, van der Knaap WO, Ammann B, Tinner W (2010) Effects of climatic change and bog development on Holocene tufa formation in the Lorze Valley (central Switzerland). Holocene 20:325–336
Wehrli M, Tinner W, Ammann B (2007) 16,000 years of vegetation and settlement history from Egelsee (Menzingen, central Switzerland). Holocene 17:741–761
Wieckowska M, Dörfler W, Kirleis W (2012) Vegetation and settlement history of the past 9000 years as recorded by lake deposits from Großer Eutiner See (Northern Germany). Rev Palaeobot Palynol 174:79–90
Willerding U (1970) Vor- und frühgeschichtliche Kulturpflanzenfunde in Mitteleuropa. Lax, Hildesheim
Wills S (1998) Cannabis use and abuse by man: an historical perspective. In: Brown DT (ed) Cannabis: The genus Cannabis. Harwood Academic Publishers, Amsterdam, pp 1–27
Xie Y, Ward R, Fang C, Qiao B (2007) The urban system in West China: a case study along the mid-section of the ancient Silk Road - He-Xi Corridor. Cities 24:60–73
Yang C-p, Zhao Y-f, Wu X-j, (2010) Exploring the craniofacial morphology of Bronze Age-Iron Age of Shanshanyanghai populations in Xinjiang. In: Dong W (ed) Proceedings of the 12th annual meeting of the Chinese society of vertebrate paleontology. China Ocean Press, Beijing, pp 121–132
Yang M-Q, van Velzen R, Bakker FT, Sattarian A, Li D-Z, Yi T-S (2013) Molecular phylogenetics and character evolution of Cannabaceae. Taxon 62:473–485
Yang Y (2014) The analysis of charred plant seeds at Jinchankou site and Lijiaping site during Qijia Culture period in the Hehuang region, China. Master thesis, Lanzhou University
Yanushevich ZV (1989) Agricultural evolution north of the Black Sea from the Neolithic to the Iron Age. In: Harris DR, Hillman GC (eds) Foraging and farming - the evolution of plant exploitation. Unwin Hyman, London, pp 607–619
Yi H (2014) From Qijia to Erlitou: in search of Xia Culture. Acad Mon 46:134–144
Yin J (2003) A regional investigation into prehistoric cultures on the Liyang Plain, Hunan Province. Archaeology 3:56–68
Yuan J (2010) Zooarchaeological study on the domestic animals in ancient China. Quat Sci 30:298–306
Zeng Y, Zhang H, Li Z et al (2007) Evaluation of genetic diversity of rice landraces (Oryza sativa L.) in Yunnan. China. Breed Sci 57:91–99
Zhang C (2013) Archaeobotanical remains from Lajia Site, Minhe, Qinghai. Master thesis, Northwest University
Zhang S, Gao H (1999) Observations on silk and hemp textiles excavated from Qingtai Site, Xingyang. Relics Cent Plain, 10–16
Zhang X (2012) Archaeobotanical investigation into the Guanting Basin, Qinhai. Archaeol Cult Relics:26-33
Zhao Z (2011) New archaeobotanic data for the study of the origins of agriculture in China. Curr Anthropol 52(S4):295–306
Zhou K-s (1963) Appendix 3: Analysis of pollen from the Neolithic site at Banpo. In: Institute of Archaeology in Academia Sinica, Pan P’o Museum (eds) The Neolithic Village at Pan P’o, SiAn. Wenwu Press, Beijing, China, pp 270–272
Zhou X, Li X, Zhao K, Dodson J, Sun N, Yang Q (2011) Early agricultural development and environmental effects in the Neolithic Longdong basin (eastern Gansu). Chin Sci Bull 56:762–771
Acknowledgements
The research was financially supported via a postdoctoral fellowship from the Fritz Thyssen Foundation (Germany) to T. Long. We thank Ernest Small and an anonymous reviewer for their constructive comments on an earlier version of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by F. Bittmann.
Rights and permissions
About this article

Cite this article
Long, T., Wagner, M., Demske, D. et al. Cannabis in Eurasia: origin of human use and Bronze Age trans-continental connections. Veget Hist Archaeobot 26, 245–258 (2017). https://doi.org/10.1007/s00334-016-0579-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-016-0579-6