
Abstract
This paper describes the patterns and processes of vegetation change and fire history in the Late Holocene (c. 3,140 cal bp) palaeoecological sequence of El Tiemblo, in a mountainous area in central Spain (Gredos range, Spanish Central System), and provides the first Iberian pollen sequence undertaken within a Castanea sativa-dominated woodland. These new data reassess not only the autochthonous nature of the species in the region and in the Iberian Peninsula, but also the naturalness of well-developed sweet chestnut forests. The study focuses on anthropogenic dynamics linked both to the effects of livestock husbandry and the use of fire for forest clearance. With this aim, non-pollen palynomorphs (coprophilous fungi ascospores) and charcoal accumulation rate are useful indicators for assessing the increasing role of human influence on vegetation.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The history of forests in the Mediterranean basin is the history of forest fragmentation, degradation and eventually deforestation (Scarascia-Mugnozza et al. 2000), a process which began in most of Western Europe during the Neolithic. However, the Holocene history of Mediterranean forests is also a history of temporal natural expansion and sometimes the maintenance of forest formations that have remained intact for millennia, so they have been resilient to multiple climate or anthropogenic impacts that they have been suffered (Chauchard et al. 2007; Carrión et al. 2010). The sweet chestnut (Castanea sativa Mill.) is a tree species that has attracted particular human attention for millennia, as it has been intensively cultivated for centuries as a monoculture (coppices and orchards), even at the limits of its potential range (Pitte 1986). As with many other tree species, these facts have severely limited differentiation among natural or introduced populations as well as the tracing of its original range in the Mediterranean basin and southern Central European regions (Mattioni et al. 2008), where currently chestnut forest ecosystems still represent an important landscape component in the mountainous regions, covering more than 2.2 million ha (Conedera et al. 2004b; Bounous and Marinoni 2005). In fact, many authors only consider the presence of this species in these territories as a result of its intensive cultivation since the Greco-Roman Age (Scarascia-Mugnozza et al. 2000).
The history of chestnut after the last ice period is not yet completely clear, although it is likely to have survived in glacial refugia located in the Mediterranean Peninsulas (Fineschi et al. 2000; Mattioni et al. 2010). Chestnut woods occur naturally in the northern part of the Mediterranean region from the Caspian Sea to the Atlantic Ocean, with some minor occurrences in Central Europe, North Africa and the Caucasus. In Spain, there are a total of c. 84,000 ha occupied by chestnut forests with four principal areas (Ruiz de la Torre 2006), north-western Iberia and the Cantabrian coast, north-eastern (Catalonia), southern Iberia (in some mountain ranges of Andalusia) and, finally, the central-western part of the Peninsula (Gredos and Francia ranges, Tiétar valley). Based on radiocarbon-dated pollen and macrofossil records, six or seven potential Quaternary refugia or macroregions of C. sativa have been delimited (Krebs et al. 2004). From these regions chestnut trees could have expanded elsewhere and there is no doubt that human cultures were also supportive (Conedera et al. 2004a, b; Lauteri et al. 2004; López-Merino et al. 2009a; Mercuri et al. 2013). Indeed, humans have radically modified and controlled the distribution of chestnut over thousands of years in southern Europe, and have long known how to manage the chestnut in extremely profitable and diversified ways (Conedera et al. 2004a). The current distribution of the species in the European continent can only be understood by considering together the complexity of the refugia during the last glaciation, the subsequent natural processes and changing environmental conditions (climate change, natural dispersal, interspecific competition) as well as human influence (Krebs et al. 2004). In any case, during some historical periods, the cultivation of the chestnut became in various regions in Europe so widespread and indispensable for the survival of mountain populations that some authors do not hesitate to identify these cultures as ‘chestnut civilizations’ (Krebs et al. 2012). Despite its extensive distribution and socio-economic role, the history of the chestnut landscape is little known, both with regard to its origins and to when and how chestnut cultivation spread (Conedera et al. 2004a; Di Pasquale et al. 2010).
The native character of the sweet chestnut in the Iberian Peninsula has long been proposed (García-Antón et al. 1990; Carrión 1992; Morla 1996), and palaeobotanical evidence supporting this consideration has been found in recent years (Carrión et al. 2003; Krebs et al. 2004; Muñoz-Sobrino et al. 2004; García-Amorena et al. 2007; Gómez-Orellana et al. 2007; Postigo-Mijarra et al. 2008), demonstrating the survival of this species throughout the Pleistocene and the Holocene mainly in the north, northwest and southeast of the Iberian Peninsula (Postigo-Mijarra et al. 2010). Meanwhile, genetic studies performed on chestnut populations from the Mediterranean basin have shown (i) the low genotypic variability of this species probably related to its spreading by humans and (ii) the existence of refuge areas in the eastern Mediterranean and the north and northwest of the Iberian Peninsula (Villani et al. 1999; Fineschi et al. 2000; Mattioni et al. 2008, 2010). However, except for some perfunctory reference (López-Sáez et al. 1996; Fernández-López and Monteagudo 2010), no reference is known to the naturalness of C. sativa in central Spain or the probability of the existence of a refugial zone in this region, although macroremains of this species have been documented in some pre-Roman archaeological sites as both fuelwood and construction elements (López-Sáez et al. 1991, 2008; Rubiales et al. 2011).
Within the framework of a regional research project on resilience and vulnerability of mountain ecosystems from the Spanish Central System, an attempt was made to examine the natural and anthropogenic causes driving the vegetation dynamics and exploitation of a small mountain valley (Yedra gorge). In this article, we provide palynological data for the last c. 3,140-year vegetation and fire history of a C. sativa-dominated forest of the Gredos range in central Spain, inferred from the palynological study of the peat bog of El Tiemblo. There are several reasons why this investigation has been considered pertinent: (i) because the available palaeobotanical data are insufficient to assess the natural character of the chestnut tree in this part of the Iberian Peninsula; (ii) to our knowledge, this is the first pollen record known in the Iberian Peninsula from a small bog located within a forest dominated by sweet chestnut; (iii) the presented pollen record spans the last three millennia, for which gaps in the palaeoecological knowledge of the region are particularly significant.
Study area
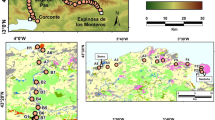
The study area is located in the mountain range of the Sierra de Gredos, in the central-west subdivision of the Spanish Central System, in particular in the Iruelas Valley Natural Reserve, in a territory of 8,828 ha roughly corresponding to the headwaters of the River Alberche catchment (Fig. 1). The climate is of a Mediterranean type, warm-temperate and rainy, with a mean annual precipitation of 1,400 mm and a mean annual temperature of about 13 °C (Molina-Moreno 1992; Ninyerola et al. 2005). Such a climate combined with a predominance of siliceous soils (granites) is favourable for the presence of the sweet chestnut, which is now dominant in south-eastern exposures, forming a continuous chestnut belt between 1,000 and 1,300 m a.s.l. A limited number of ancient giant chestnut trees—aged over 500 years—have been reported (Génova et al. 2009).

The study area in the Gredos range (Spanish Central System, central Spain) and location of the fossil record (El Tiemblo peat bog). Red represents the modern distribution of the forest formations dominated by Castanea sativa in the Iruelas valley and surrounding areas
The slopes of the Iruelas valley are characterized by a mosaic of Quercus pyrenaica stands, dense Pinus pinaster woodlands and isolated P. nigra trees. Those altitudes over 1,600 m a.s.l. are widely occupied by shrublands, where brooms such as Cytisus oromediterraneus and Echinospartum barnadesii constitute the main features of the landscape (Molina-Moreno 1992). Isolated Pinus sylvestris stands are frequent at altitude, shaping the timberline. Riparian forests are characterized by Alnus glutinosa, Ulmus glabra and Fraxinus angustifolia.
El Tiemblo peat bog (30T 4468565N, 370365E), covering a surface area of 0.02 ha, is located in the Yedra gorge (Iruelas Valley Natural Reserve, Gredos range, province of Ávila, Spanish Central System, central Spain), within the supramediterranean belt (1,250 m a.s.l.). The modern vegetation of the peat bog includes Caricetum carpetanae oligotrophic communities dominated by Carex nigra, C. echinata, Parnassia palustris and Calluna vulgaris, and the catchment is mainly covered by several centuries-old sweet chestnut orchard trees.
Materials and methods
Coring and palynological analysis
A 120-cm core was extracted from El Tiemblo peat bog with a Russian corer. 60 samples of 1 cm3 were analysed at intervals of 2 cm. Later, the stratification of the peat bog sediments was recorded (Table 1) in order to clarify certain important events in their formation and evolution. All samples were treated according to the Fægri and Iversen (1989) method, using Thoulet solution for densimetric extraction of pollen and non-pollen microfossils (Goeury and de Beaulieu 1979), although acetolysis was not carried out to allow the identification of any contamination by modern pollen. No macrofossils were discerned throughout the core. Small aliquots of the residues were mounted in glycerine, sealed with Histolaque and all recognizable pollen, spores and non-pollen microfossils were counted under a light microscope using a magnification of ×400, until a pollen count of at least 400 was reached. Ferns, aquatics and NPPs were excluded from the main pollen sum in order to avoid biases from the local pollen representation. Pollen grains were identified with the help of different keys and pollen atlases, such as Fægri and Iversen (1989), Moore et al. (1991) and Reille (1992), and the reference collection of the Archaeobiology Laboratory of CSIC (Madrid). Castanea sativa was palynologically discriminated according to van Benthem et al. (1984) and Pinus pollen differentiation follows Carrión et al. (2000). Non-pollen palynomorphs (NPPs) were identified using the nomenclature proposed by van Geel (2001).
Pollen concentration (grains cm−3) was estimated by adding a Lycopodium tablet to each sample (Stockmarr 1971), and these values were divided by deposition time (year cm−1) to calculate pollen accumulation rate (PAR; grains cm−2 year−1). Data processing and graphic representation of pollen data was performed with the help of the TILIA and TGView software (Grimm 1992, 2004). The program IBM SPSS Statistics 21 was used to conduct the statistical analysis. To establish the zonation of the pollen sequence, we tested several divisive and agglomerative methods. Based on the ecological meaning of the obtained zones, local pollen assemblage zones (LPAZs) were constructed on the basis of agglomerative constrained cluster analysis of incremental sum of squares (CONISS) with square root transformed percentage data (Grimm 1987). The number of statistically significant zones was determined using the broken-stick model (Bennett 1996).
Radiocarbon dating
Nine bulk organic sediment samples were 14C dated using AMS technique. The AMS dating was conducted at the Centro Nacional de Aceleradores (CNA) Laboratory of the University of Sevilla, Junta de Andalucía and CSIC. The dates were calibrated using CALIB 7.1 with the IntCal13 curve (Reimer et al. 2013) (Table 2). An age-depth model (Fig. 2) was produced using Clam 2.2 software (Blaauw 2010), fitting a smoothing spline to the available radiocarbon dates. Confidence intervals of the calibrations and the age-depth model were calculated at 95 % (2σ) with 1,000 iterations.

Age-depth model for El Tiemblo peat bog. 95.4 % CI are represented in grey
Charcoal analysis
In order to reconstruct local fire history at El Tiemblo peat bog, macroscopic charcoal (>125 µm) was identified and counted from subsamples of 1 cm3 at every 2 cm depth by sediment sieving. The samples were soaked in a 3 % sodium metaphosphate solution (72 h) to deflocculate any particles and then washed through a 125 µm mesh sieve. Particles >125 µm diameter are not transported far from their source and thus provide information on local fire history (Whitlock and Larsen 2001). Charcoal accumulation rate (CHAR) was calculated by sedimentation rate (cm year−1) and is expressed in particles cm−2 year−1.
Results
Sedimentology
The core taken from El Tiemblo peat bog has a thickness of 120 cm and consists mainly of peat (Table 1). The lowest 60 cm (60–120 cm) of the core are a moderately humified dark peat with a high concentration of pollen (>400 × 103 grains/cm3) in the lower samples (77–120 cm) compared to the overlaying sediments (60–77 cm, 115–375 × 103 grains/cm3). The next layer (37–60 cm) consists of 23 cm of dark peat with silts, some gravel, fine and coarse sands and also low concentration of pollen (101–259 × 103 grains/cm3). This thin layer gradually transforms into a 37 cm-thick sedge peat that constitutes the uppermost layers (0–37 cm) with increasing pollen concentration (214–462 × 103 grains/cm3). The degree of humic decomposition of the peat is low to medium and fluctuates between 2 and 3 through the core.
Dating
The results of the AMS-radiocarbon dating are shown in Table 2 and Fig. 2. The 120 cm basal radiocarbon date is 3,140 cal bp for the onset of peat deposition. The age-depth model (Fig. 2) is based on nine radiocarbon data points and shows the variability in accumulation rates of El Tiemblo peat bog. The peat formation began at c. 3,140 cal bp (120 cm) with an accumulation rate of about 0.043 cm year−1. It remained fairly constant (0.042 cm year−1) c. 3,140–2,950 cal bp (120–112 cm), although it declined progressively throughout the Early Iron Age (c. 3,100–2,350 cal bp) to 0.038 cm year−1 (110–89 cm). Subsequently during the Late Iron Age (c. 2,350–2,000 cal bp) the accumulation rate declined about 0.033 cm year−1 (89–75 cm), then gradually increased to 0.034 cm year−1 during Roman times (c. 2,000–1,500 cal bp; 75–61 cm). During the Early Middle Ages (c. 1,500–850 cal bp), peat accumulation grew to 0.042 cm year−1 until c. 1,090 cal bp (44 cm) and declined again to 0.036 cm year−1 at the end of this period. Throughout the Feudal/Christian period (c. 850–500 cal bp) the accumulation rate decreased to near 0.030 cm year−1 and during the Modern period (500 cal bp to present) the peat bog grows faster at a rate of 0.065 cm year−1.
Vegetation history
The results of identification and counting of pollen grains, spores and NPPs are presented as a percentage diagram (Fig. 3). The sequence was divided into six zones. Figure 4 shows CHAR and PAR related to selected pollen and NPP percentage curves and climatic events.

Percentage pollen diagram for trees, shrubs, herbs, hydro-hygrophytic taxa, pteridophytes and NPPs of El Tiemblo peat bog (exaggeration curves are ×10)

Climatic events, Charcoal accumulation rate (CHAR), pollen accumulation rate (PAR) and selected pollen and NPP percentage diagram from the palynological record from El Tiemblo plotted against age (cal bp)
LPAZ 1 (120–76 cm; c. 3,140–1,915 cal bp)
Castanea sativa percentages are high (31–42 %) indicating a sweet chestnut forest in the vicinity. Deciduous Quercus (15–22 %) and evergreen Quercus (2–6 %) show continuous occurrences, while Betula, Corylus, Fraxinus, Ilex, Pinus pinaster, P. pinea, Salix, Taxus and Ulmus types are usually low (<5 %). In this zone, oscillations of Alnus (3–12 %) and Pinus sylvestris/nigra (4–11 %) types are noticed. The amounts of shrub pollen are between 2–6 % including Cistus ladanifer, Erica arborea, Genista, Prunus spinosa, Lonicera and Lamiaceae types. Herbaceous pollen is between 6 and 9.5 % and PAR very high (418–911 × 103 grains cm−2 year−1). Coprophilous fungi—Sordaria, Sporormiella—and both anthropozoogenic—Plantago lanceolata—and anthropogenic pollen taxa—Aster, Cardueae, Cichorioideae—show discontinuous and sporadic presence (<1 %). Macrocharcoal particles are not present. Alnus (11–12 %), Betula (3 %), Fraxinus (3 %), Ilex (2.7 %), Salix (3 %) and Cyperaceae (17–19 %) show a greater presence at 106–104 cm (c. 2,780–2,730 cal bp), but Castanea (33 %) and Pinus sylvestris/nigra (4–5 %) sharply decline.
LPAZ 2 (76–61 cm; c. 1,915–1,465 cal bp)
In this zone, C. sativa values are always high (34–41 %). The remaining arboreal pollen taxa keep similar percentages to those of the previous LPAZ 1 zone, except Pinus pinaster (<2 %) and P. pinea (<1 %) which decrease and Betula (2.5 %) and Corylus (6 %) which slightly increase. Olea europaea (0–1 %) is documented for the first time. Most shrubs and herbs maintain a continuous and significant presence throughout, reflecting the increased importance of the understorey. Anthropogenic (Aster 1–1.5 %, Cardueae 1–2 %, Cichorioideae 1–3 %) and anthropozoogenic taxa (Plantago lanceolata 0–1 %, Urtica dioica 0–1 %) increase, as do Pteridium aquilinum (1.5–2.5 %) and coprophilous fungi (Sordaria 0.5–1.2 %, Sporormiella 1.4–2.3 %). Macrocharcoal particles are absent except for the upper sample (62 cm), where CHAR reaches 2 particles cm−2 year−1, while PAR sharply decreases (225–375 × 103 grains cm−2 year−1).
LPAZ 3 (61–38 cm; c. 1,465–925 cal bp)
This zone is characterized by oscillations in the C. sativa pollen curve (between 30.5 and 38 %). Also noteworthy are the continuous presence of Olea europaea (2–4 %), Alnus, Corylus, Ilex, Fraxinus and Taxus, and the increases of Betula (5–8 %), Pinus pinaster, P. pinea and evergreen Quercus (5–6.5 %). Deciduous Quercus (12–13 %) and Pinus sylvestris/nigra (3–6 %) pollen types decrease. Anthropogenic (Aster 2–4 %, Cardueae 2.5–4.2 %, Cichorioideae 2.5–4.5 %) and anthropozoogenic taxa (Plantago lanceolata 1–1.5 %, Urtica dioica 1.2–2.5 %), together with coprophilous fungi (Sordaria 1.5–3.5 %, Sporormiella 1.4–3.5 %) and Pteridium aquilinum (maxima 6.6 %) show a greater presence in LPAZ 3, while Cyperaceae sharply decline. PAR decreases ranging between 101 and 158 × 103 grains cm−2 year−1. This zone is also characterized by rapid accumulation of charcoal between 60 and 52 cm (CHAR 23–43 particles cm−2 year−1) and 46 and 38 cm (CHAR maxima 22 particles cm−2 year−1).
LPAZ 4 (38–15 cm; c. 925–430 cal bp)
This zone is characterized by synchronous increases of C. sativa (34–44.5 %), Alnus (6–9.5 %) and deciduous Quercus (17–21 %). Betula, Pinus pinaster, P. pinea and Olea europaea decrease, while evergreen Quercus, Pinus sylvestris/nigra and other tree taxa remain more or less constant. Shrub taxa percentages (Erica arborea, Genista) experience a decreasing tendency, while Cistus ladanifer continue to be relatively abundant. The trend in CHAR (17–23 particles cm−2 year−1) corresponds with shifts in those pollen assemblages (38–34 cm) that have higher values of anthropogenic taxa (Aster, Cardueae, Cichorioideae) and Pteridium aquilinum. Anthropozoogenic types such as Plantago lanceolata and Urtica dioica continue to be present. Hydro-hygrophytic elements such as Cyperaceae or Ranunculaceae increase. Coprophilous fungi increase (>5 %) c. 780 cal bp after CHAR maxima in the bottom samples. PAR oscillates between 232 and 430 × 103 grains cm−2 year−1.
LPAZ 5 (15–7 cm; c. 430–195 cal bp)
In this zone increasing levels of C. sativa (37–46 %) are noticed. Betula, Corylus, Olea europaea, deciduous Quercus and Pinus sylvestris/nigra decrease, while P. pinea, Taxus and Cistus ladanifer disappear, and P. pinaster increases from 1 to 3 %. Genista, Prunus spinosa and Lonicera also increase. Cyperaceae attains higher values (5–8.5 %), and Sordaria (7.5 %), Plantago lanceolata (4.3 %) and Urtica dioica (2.7 %) reach a maximum. Aster, Cardueae and Cichorioideae remained low (<1 %). PAR remains relatively stable (246–214 × 103 grains cm−2 year−1) and CHAR values are null.
LPAZ 6 (0–7 cm; c. 195 cal bp–present)
Castanea sativa is at its maximum in this pollen sequence (48.6 %) but with a decreasing trend. Other components increase such as Pinus pinaster (7 %), P. sylvestris/nigra (6.5 %), Taxus and Ilex, while other arboreal taxa (Alnus, Betula) decrease. Among shrubs, only Erica arborea, Genista, Prunus spinosa and Lonicera show significant values, as well as Cyperaceae (6–10 %) among the hygrophilic taxa. Coprophilous fungi and anthropozoogenic taxa are absent. Anthropogenic taxa continue with low values. PAR increases (296–462 × 103 grains cm−2 year−1) and macrocharcoal particles are present in low concentrations (CHAR 1–2 particles cm−2 year−1).
Discussion
Pollen representation and representativeness of the coring site
So far little has been done in modern pollen rain studies to estimate the percentage representation of chestnut pollen inside and outside of its forests and in relation to the topographical variability (e.g. differences between valleys and mountains). Detailed studies on current pollen rain in northwestern Iberia (Aira and Barthelemy 1990) show that (i) Castanea pollen is present basically in valley areas and it is not documented in elevated ones, and, (ii) inside chestnut forests, Castanea pollen percentage is high (>20 %), but on the outside is greatly reduced (<5 %). The data provided by the pollen record from El Tiemblo (Figs. 3, 4), where Castanea pollen values range from 30.4 to 48.6 %, reflect what is stated in the above mentioned research, i.e. the situation of a valley within a chestnut forest. In fact, chestnut pollen is absent or appears only sporadically (<5 %) in modern pollen rain samples from topographical and altitudinal situations other than the pollen record from El Tiemblo in the Spanish Central System (López-Sáez et al. 2010a, 2013, 2015). This forces us to point out some issues relating to the representativeness of the site in terms of pollen source area: (i) the coring site consists of a small bog of only 0.02 ha; (ii) it is located in a relatively enclosed river valley; (iii) the pollen record from El Tiemblo shows a relatively high sweet chestnut cover for more than three millennia. These data lead us to consider the coring site as a ‘microsite’ which can only be useful for local vegetation reconstruction. Therefore, distinguishing between the local and regional part of a single pollen assemblage at a sampling site can only be achieved with modelling approaches, which require that estimates of pollen productivity are available for the taxa occurring in the pollen record. Unfortunately, no research on the pollen source area has so far been conducted in central Spain.
Usually, the scarce and usually sporadic presence of chestnut in Quaternary pollen records of the Iberian Peninsula has been interpreted as a result of its regional distribution, thanks to the medium and long distance transportation capacity of C. sativa pollen highlighted by several authors (Peeters and Zoller 1988; Frei 1997; García-Mozo et al. 2007). Huntley and Birks (1983) observed that chestnuts are not great pollen producers and that the little pollen which chestnuts do produce is poorly dispersed, so a value >5 % almost certainly reflects the widespread occurrence of Castanea woodlands near the site, while lower values can be regarded as a fairly reliable indication of the local occurrence of mixed woodland with some chestnut trees. Consequently, Morales-Molino et al. (2015) considered a ‘rational limit’ when chestnut pollen represents >5 % of the pollen spectra, and an ‘empirical limit’ when a continuous pollen record of C. sativa is detected even with values below 5 %. Undoubtedly the pollen record from El Tiemblo reflects the in situ persistence of a dense natural chestnut forest during the last three millennia. In short, a difficult issue to resolve is the determination of the origin of chestnut pollen and therefore of the effective spatial representation of the pollen curves (Krebs et al. 2004; Conedera et al. 2006).
Chestnut forests linked to anthropic and climate dynamics and fire occurrence
In the Gredos range the onset of the Iron Age has been linked to the so-called 2.8 ka cal bp event (López-Sáez and Blanco-González 2005; López-Sáez et al. 2009a), an abrupt and short climatic change on a global scale which occurred c. 2,800–2,710 cal bp, and which represents the evolution from warm and dry conditions to wetter and colder ones (van Geel et al. 1998; van Geel and Berglund 2000). Pollen sequences from both high and low-altitude peat bogs in the Gredos range show how the increasing rainfall caused a higher environmental humidity, triggering the decline of high-mountain pine forests and increases in riparian trees and wetlands taxa (López-Sáez et al. 2014 and references therein). These facts are well documented in the pollen record from El Tiemblo c. 2,780–2,730 cal bp (LPAZ 1) by increased values of Alnus, Betula, Fraxinus, Ilex, Salix and Cyperaceae, and by the drop in the percentages of Castanea and Pinus sylvestris/nigra (Figs. 3, 4). From c. 2,900 cal bp onwards a gradual process of human presence in mountain environments of the Gredos range has been recognized, with the emergence of the first nucleated and long-lasting permanent villages during the Iron Age along with the persistence of small isolated farmsteads in the more fertile lowlands (Álvarez-Sanchís 2000, 2005). However, no evidence of these facts is documented in the pollen record from El Tiemblo (Figs. 3, 4), while in other pollen sequences from this mountain range percentage increases of anthropogenic pollen taxa and grazing indicators including coprophilous fungi are often recorded c. 2,850–2,650 cal bp, and even cereal cultivation, a decrease in oak forests and higher macrocharcoal concentration directly related to fire activity (Franco-Múgica et al. 1997; Dorado et al. 2001; Abel-Schaad and López-Sáez 2013). The explanation of the data provided by the pollen record from El Tiemblo is that the Iruelas Valley Natural Reserve was not populated throughout the Iron Age as evidenced by the total lack of archaeological sites of this period c. 3,100–2,000 cal bp (Mariné 1995).
LPAZ 2 coincides roughly with the Roman period (c. 2,000–1,500 cal bp) in the study area (Figs. 3, 4). Although the landscape is still dominated by a dense chestnut forest with patches of deciduous Quercus, Betula and Corylus, the first clear evidence of human impact is already apparent in the pollen record from El Tiemblo through slight increases in anthropogenic and anthropozoogenic taxa, coprophilous fungi and Pteridium aquilinum, and the first appearance of the olive tree. In any case, it would be a residual anthropic pressure as it probably did not involve the use of fire as an element in deforestation (minimum values of CHAR). Pollen diagrams from the whole of the Spanish Central System show a higher intensity of human pressure during the Roman period in low-altitude deposits (<1,400 m a.s.l.) (Abel-Schaad et al. 2009b; López-Sáez et al. 2010b; Abel-Schaad and López-Sáez 2013; Morales-Molino et al. 2013), usually related to the existence of small villages and farming communities (López-Merino et al. 2009b; López-Sáez et al. 2009b). In contrast, the landscape appears scarcely altered by human activities in high-altitude deposits c. 1,400–1,700 m a.s.l. (Atienza et al. 1990; López-Sáez et al. 2009b). During this time, the Gredos range was just an access route, and the area was a marginal territory sparsely populated due to its inhospitable character for the development of urban centres (López-Merino et al. 2009b). In general, in the Gredos range Roman presence is not definitively apparent until the Late Roman period, from the first century ad (c. 1,950–1,850 cal bp) when a significant population increase is documented (Mariné 1995). During the Roman period the settlement was concentrated on plains and easily accessible fertile lands such as river valleys, as their main interests were agriculture and livestock. For these reasons most of the Roman settlements (especially villae) were located north of these mountains (Adaja and Tormes valleys), far from the study area (Hernando 2002). The scattered evidence of human impact recorded in the pollen record from El Tiemblo can be assigned to the aforementioned settlement model of small villages mainly devoted to livestock and agriculture. The dense chestnut forest present in the study area remained unchanged during the Roman period.
Raw CHAR values usually range between 0 and 2 particles cm−2 year−1 between c. 3,140 and 1,495 cal bp, followed by increasing values from c. 1,440 cal bp (Fig. 4). Maximum values over 20 particles cm−2 year−1 are present around c. 1,385, 970 and 830 cal bp. This suggests that fire events were of local origin (Whitlock and Larsen 2001). Finally, CHAR values drop again during the last millennium (0–2 particles cm−2 year−1). The above mentioned largely forested landscape dominated for a relatively long period of low or no fire activity and human impact between the Iron Age and the Roman period. Notwithstanding this general trend, a certain forest opening is apparent at c. 1,465–830 cal bp (LPAZ 3 and bottom of LPAZ 4) linked to local fire activity during the Early Middle Ages (Fig. 4). This episode is followed by CHAR increases, slight decreases of chestnut values (to 30 % c. 1,240 and 1,105 cal bp) and by a rapid and remarkable spread of the olive tree and indicators of human impact (anthropogenic and anthropozoogenic taxa and coprophilous fungi). At the same time, a large amount of Pteridium aquilinum spores occurred suggesting fernland spread, which could be related to increasing burning (López-Sáez et al. 2010b).
During the first part of this cultural period, the Visigothic period (c. 1,500–1,240 cal bp), a phase of widespread deforestation processes occurred in the Gredos range, especially in montane areas, within a clear livestock-oriented pattern (López-Sáez et al. 2009b, 2014). Forests were cleared to obtain new pastures in a period of rural habitat proliferation and a land exploitation led by peasantry (Blanco-González et al. 2009). These forest clearances were mainly driven by means of fire, as documented in low-altitude deposits (Lanzahíta, Peña Negra) following the trend set by the Romans, but more intensely in higher ones like Serranillos, Narrillos del Rebollar, Hoyos del Espino and Navarredonda (Franco-Múgica et al. 1997; Dorado et al. 2001; López-Merino et al. 2009b; López-Sáez et al. 2009b, 2010b; Abel-Schaad and López-Sáez 2013). As in all such pollen sequences, in the pollen record from El Tiemblo these facts are probably related to grazing activities (high values of anthropozoogenic taxa and coprophilous fungi) and fire occurrences (CHAR maximum c. 1,385 cal bp) (Figs. 3, 4), in order to preserve upland pastures and extent the olive tree.
Climatic conditions also affected the evolution of vegetation, particularly with the onset of the Early Medieval Cold Episode (c. 1,500–1,000 cal bp), marked by lower temperatures and greater aridity (Martín-Puertas et al. 2008; Blanco-González et al. 2015). This climate change clearly influenced the vegetation of the Iruelas Valley Natural Reserve in the Early Middle Ages (Fig. 4), as evidenced in the pollen record from El Tiemblo, which reports an arid phase extending from c. 1,440 to 970 cal bp. As in many pollen records from the Spanish Central System (López-Sáez et al. 2014), this cold episode is documented in the pollen record from El Tiemblo by a percentage decrease in Cyperaceae and an increase in birch, possibly indicating a lowering of the treeline at high altitude, also demonstrated by the growing at lower altitudes of high-mountain pines (Pinus sylvestris/nigra). Later, during the Late Medieval Warm Episode (c. 1,000–600 cal bp) temperature increases with moderate rainfall and severe drought episodes (Desprat et al. 2003; Martín-Puertas et al. 2008; Blanco-González et al. 2015). This mild episode is reflected in the pollen record from El Tiemblo by the rise of Alnus, deciduous Quercus, Ranunculaceae and Cyperaceae, and the decline of Betula and P. sylvestris/nigra, better adapted to cold and continental conditions.
During the post-Visigothic or Muslim period (c. 1,240–850 cal bp), corresponding to the top of LPAZ 3 and the bottom of LPAZ 4, values of C. sativa and Alnus increase, while those of Olea europaea and Erica arborea decrease indicating a significant reduction of human pressure on the environment. The historical data about this period fully corroborate the presented pollen record. They describe an economic model based on livestock grazing and small subsistence crops managed by relatively self-sufficient peasants, in a sparsely populated land (García-Garcimartín 2002; Blanco-González et al. 2015). Until the effective political possession (the so-called Reconquest) and colonization of the Gredos range by the northern Christian kingdom c. 950 cal bp, this massif acted as a borderline between Muslim and Christian powers for several centuries (c. 1,240–950 cal bp) (Manzano 1991). From the political Reconquest and demographic repopulation of the Gredos range by c. 950 cal bp, some of the members of the urban councils directed a process of intensification of the rural economy (Monsalvo 2003). This trend was aimed at supplying the urban markets and was based on the intensification and specialization of livestock husbandry during the following Feudal/Christian period (c. 850–500 cal bp). At this time the climate was still under the influence of the Late Medieval Warm Episode. In the pollen record from El Tiemblo (top of LPAZ 4) it is characterized by the intensification of livestock activities (increased coprophilous fungi) and oscillating values of C. sativa. CHAR and grazing indicators also provide a stronger signal, especially c. 970 and 830 cal bp (Fig. 4), pointing to a slightly more intense human pressure, probably related to the presence of northern Christian settlers.
Climatic conditions between the end of the Feudal/Christian period and the beginning of the Late Modern period correspond to Little Ice Age (LIA), which would have started around c. 600 cal bp and lasted until c. 100 cal bp (Desprat et al. 2003). In the Iberian Peninsula, the LIA was a cold interval with an initial dry phase c. 600–400 cal bp and a later and more humid one until c. 100 cal bp (Manrique and Fernández-Cancio 2000). The first dry phase of the LIA is documented in the pollen record from El Tiemblo by the rise of Betula and decreasing values of both hydro-hygrophytic taxa and Castanea, while the humid second phase is demonstrated by the opposite trend (Fig. 4).
The Early Modern period (c. 500–150 cal bp) was, in the Spanish Central System, a time of overall agricultural expansion and increasing livestock, due to the continuous population growth (Blanco-González et al. 2015). The pollen record from El Tiemblo concurrently displays an increasing human pressure, primarily with a reduction of chestnut (minimum 37 %, Fig. 3) and high values of both anthropozoogenic taxa and coprophilous fungi (Fig. 4). Birch forests recorded in El Tiemblo start a sharp decline c. 400 cal bp parallel to the spread of shrublands (mainly Genista type; Fig. 3), pointing to an overall increasing grazing pressure. From a palaeoenvironmental point of view, the most striking feature of this period in central Spain was the development of the olive groves (López-Sáez et al. 2014). These facts are evident at the beginning of this period (c. 500–400 cal bp) in the pollen record from El Tiemblo, although such olive cultivation is substantially reduced during the second humid phase of the LIA (Fig. 4). In any case, the presence of Olea europaea pollen is continuous throughout the Early Modern period (top of LPAZ 4, LPAZ 5 and bottom of LPAZ 6) (Fig. 3). In the late 18th century (c. 200–150 cal bp) new regulations were established with the growing concern about the scarcity of forest resources in the Spanish Central System (Pardo and Gil 2005), which would have favoured the development of chestnut stands (maximum 48.6 % c. 170 cal bp).
Finally, during the Late Modern period (c. 150 cal bp to present) several afforestation plans were developed in the Iberian Peninsula with the aim of increasing tree cover, mainly with massive pine plantations in the case of the Iruelas valley (Molina-Moreno 1992). These could be the reasons for the percentage increase in both Pinus sylvestris/nigra (6.5 %) and P. pinaster (7 %) in the pollen record from El Tiemblo (Figs. 3, 4; top of LPAZ 6), although both taxa have been present in the study area during the last three millennia. But such afforestation with pines could also explain the downward trend experienced by C. sativa values since c. 150 cal bp. Unlike other pollen records from the Spanish Central System displaying a higher prevalence of livestock with maxima of coprophilous fungi and anthropozoogenic taxa during the Late Modern period (López-Sáez et al. 2014 and references therein), the palynological sequence of El Tiemblo shows no evidence of such activities since both coprophilous fungi and anthropozoogenic taxa disappear from the record at this time (Fig. 4). Instead, it shares a new recurrence of fire incidence (low CHAR values), probably related with monospecific pine plantations that favour large-scale fires.
Three millennia of naturalness and resilience of chestnut forests in the Gredos range
The pollen record from El Tiemblo is the irrefutable proof not only of the naturalness of sweet chestnut tree in central Spain, but also of the existence of well characterized forests of this species (Castanea pollen >30 %) during at least the last c. 3,140 years, supporting a pre-Roman origin for the Iruelas Valley Natural Reserve chestnut forest. Our data demonstrate the millenary resilience of chestnut forests in the Gredos range, probably related to some kind of human management since the Middle Ages. Even in those moments when the human impact was little or absent, this chestnut forest was not invaded by other species such as the Pyrenean oak (Quercus pyrenaica) but remained constant, resilient to climate changes and to anthropic dynamics of the last three millennia. More than 90 % of all chestnut stands in the Gredos range occur in the Iruelas valley and surrounding areas (Fig. 1), where these forests have been managed since at least the 13th century following traditional ecosystem management practices (Seijo et al. 2015). However, a few potential natural stands of chestnut have persisted away elsewhere in this massif (Jerte valley, Béjar range, Barranco de las Cinco Villas) (Pulido et al. 2007; Génova et al. 2009). In the absence of management chestnut coppices started to be invaded by other species (e.g. Betula, deciduous Quercus etc.). In fact, these forests were impoverished, floristically speaking, during the Visigothic and Christian periods, in a process related to general increase in anthropization, grazing and regional olive cultivation.
Although the chestnut forests of the Gredos range have numerous floristic affinities with Pyrenean oak forests, they have certain characteristics species of flora such as Paradisea lusitanica, Neottia nidus-avis, Delphinium fissum ssp. sordidum and Pulmonaria longifolia (Luceño and Vargas 1991; Pulido et al. 2007), which makes us rethink the nature of such native chestnut forests. Furthermore, within these chestnut forests many extremely rare plants find shelter in the centre of the Iberian Peninsula such as Allium scorzonerifolium, Aquilegia vulgaris, Arctium minus, Cephalanthera rubra, Circaea lutetiana, Doronicum plantagineum, Erysimum lagascae, Hesperis laciniata, Lilium martagon, Narcissus pseudonarcissus, Paeonia officinalis or Pentaglottis sempervirens, which probably grow in its last southern Iberian refuge in the forest of El Tiemblo (Génova et al. 2009). Some of these species are present in the pollen record from El Tiemblo (Fig. 3), more or less continuously, represented by different pollen types (Amaryllidaceae, Circaea, Lilium martagon, Paeonia), except during the Middle Ages, when the highest degree of human impact was achieved. In short, our pollen data allow the application of the concept of the “eco-cultural niche” proposed by Krebs et al. (2012) to the chestnut forests of the Iruelas Valley Natural Reserve (Gredos range), as this population includes some giant chestnut trees and has been exposed to both climatic and anthropogenic/cultural factors during at least the last two millennia, when from the Roman period the study area began to suffer the first clear influences of human impact (increase of anthropogenic and anthropozoogenic taxa, coprophilous fungi and Olea) and chestnut forest began to be handled by the man in a consistent way. Although chestnut forests from the Iruelas Valley Natural Reserve have not been considered in the phytosociological literature, the conjunction of palynological data and the unique flora of these forests—Populus tremula, Prunus avium, P. spinosa, Ulmus glabra, Ilex aquifolium, Corylus avellana, Cytisus multiflorus, C. scoparius, Erica arborea, Genista falcata, G. florida, Lonicera periclymenum, Allium scorzonerifolium, Aquilegia vulgaris, Arctium minus, Cephalanthera rubra, Circaea lutetiana, Doronicum plantagineum, Erysimum lagascae, Lilium martagon, Paeonia officinalis, Pulmonaria longifolia (Génova et al. 2009)—suggests that they must be studied in detail with the aim of defining such communities testified by the pollen record.
Since the second half of the last century, chestnut forest maintenance and cultivation substantially decreased in the Iberian Peninsula because of the loss of importance of this species as staple food or for timber, progressive depopulation of the countryside—especially in mountain areas—and the onset of pathogens causing ink disease (Phytophthora cinnamomi and P. cambivora) and chestnut blight (Cryptonectria parasitica) (Taveira 1979). As a consequence, chestnut stands, orchards and forests across large areas were partially or entirely abandoned (Manetti et al. 2001). In the absence of cultural inputs of management open-structure chestnut forests tend to be invaded by other trees, giving way to evolutionary dynamics leading toward potential climax communities associated usually with Fagus sylvatica or Quercus species, as has been observed in many European regions in recent decades (e.g. Conedera et al. 2001; Gallardo-Lancho 2001; Pezzi et al. 2011). Despite its relatively limited distribution in the Gredos range, chestnut had been of great importance for the population in the region. In central Spain these forests have been managed both for timber production as coppice stands and for edible nut production as high forest stands, although intermediate structures can also be found (Rubio et al. 1999; Gallardo et al. 2000). Coppicing under traditional practices in central Spain has led to the conservation of a high level of diversity in chestnut forests, it being possible to find endangered flora elements such as Ilex aquifolium, Rhaponticum exaltatum, Linaria triornithophora, Melittis melissophyllum or Paeonia broteroi (Rubio and Sánchez-Mata 1995). The key factor of coppicing must be related to the fact that it mimics the natural gap-phase dynamics, which should allow both forest exploitation and maintenance of the natural high herb-layer diversity (Rubio et al. 1997).
Although Europe-wide diffusion of chestnut cultivation is very evident from the Roman and Christian periods in the Gredos range (e.g. López-Sáez et al. 2010b; Abel-Schaad and López-Sáez 2013), including the so-called ‘golden age’ in western Europe (Conedera et al. 2004a, b; Conedera and Krebs 2008; Mercuri et al. 2013), in the case of the pollen record from El Tiemblo we must admit the existence of a dense and native C. sativa forest in this area of the Gredos range during the last three millennia whose origin is unclear. As pointed out, palaeoecological records indicate that the chestnut tree was present in the Iberian Peninsula before the last glaciations and then became largely extinct except for some northern populations that survived during the Last Glacial Maximum (Krebs et al. 2004). Then it appeared again in many parts of Iberia in Prehistoric times, presumably re-introduced by man in the last two millennia (Conedera et al. 2004a). The presence of chestnut in the Spanish Central System is mainly supported by pollen finds (~5 %) in the Middle Pleistocene palaeontological site of Villacastín (Carrión et al. 2007), in the Chalcolithic archaeological sites of Aldeagordillo and Valdeprados c. 4,800–3,600 cal bp (~2 %) (López-Sáez et al. 2003; Fabián et al. 2006), as well as with sporadic appearances (1–2 %) in the Garganta del Trampal, Puerto de Santa Clara-El Payo, Peña Negra, Lanzahíta, La Meseguera and La Panera peat bog pollen records dating from c. 6,000, 3,800, 3,200, 2,700, 1,200 and 200 cal bp respectively (Atienza et al. 1990; Abel-Schaad et al. 2009a, b, 2014a; López-Sáez et al. 2010b; Abel-Schaad and López-Sáez 2013), and by the timber macroremains found in the Vettonian (pre-Roman) settlement of El Raso de Candeleda c. 2,350–1,900 cal bp (López-Sáez et al. 1991, 2008). These findings are more or less concentrated in southern exposures in the central-western part of this mountainous massif (Gredos range) at altitudes between 1,000 and 1,400 m a.s.l. (López-Sáez et al. 2014). Anyway, they are already scattered, sporadic (usually the values of chestnut pollen do not exceed 5 %) and far enough apart to assume the possibility of a large refuge area in the whole of the Gredos range. These data are likely to represent the presence of isolated chestnut trees within other deciduous formations (e.g. deciduous oak woodlands). However, the data provided by El Tiemblo are conclusive (Castanea 30.4–48.6 %) and lead us to seek a plausible explanation beyond its pre-Roman origin.
The sweet chestnut tree (Castanea sativa) is a mesophilic or moderately thermophilic species, well adapted to ecosystems with an annual mean temperature ranging between 8 and 15 °C and monthly mean temperatures for 6 months over 10 °C (Bounous and Beccaro 2002), these being the most adverse cold weather and drought growth conditions in the Iberian Peninsula (Gandullo et al. 2004). According to García-López and Allué-Camacho (2008), the Gredos range has one of the highest phytoclimatic suitability rates for the presence of chestnut trees in the Iberian Peninsula, which means that these mountains can be considered as a habitat of high durability of its forests (capacity for self-regeneration, ability to compete with other species, resistance to diseases). In the Iruelas Valley Natural Reserve this tree occupies well-drained fertile soils, rich in potassium and with low pH and with an optimal precipitation range of 1,400 mm per year (Fig. 1), representing very suitable soil and climate conditions for the survival of chestnut trees (Berrocal et al. 1998). According to the detailed genetic studies of Fernández-López and Monteagudo (2010), the chestnut subpopulation from El Tiemblo is the purest within the Iberian Mediterranean group of populations, based on the longest estimated ancestry. These data allow us to suggest that the chestnut forest of the Iruelas Valley Natural Reserve could be probably older than 3,140 years.
Benito-Garzón et al. (2007) modelled potential distribution during the Last Glacial Maximum (LGM) and the mid-Holocene for C. sativa. Their reconstruction suggests that climatic conditions in the north-western and northern Iberian Peninsula and the foothills of the Pyrenees at the LGM may have enabled the persistence of chestnut trees. Their model (ECHAM3 scenario) also proposed the potential existence of scattered populations of this species along the Spanish Central System including the Gredos range, as well as a great expansion of chestnut trees during the mid-Holocene in these mountains. These data agree with palaeoenvironmental data that have already demonstrated the importance of the Spanish Central System as an area of persistence of tree relicts from the Tertiary, such as Taxus baccata, Ulmus glabra, Quercus robur, Prunus lusitanica, Fagus sylvatica, Carpinus betulus, Tilia sp. and Betula sp. (López-Sáez and López-García 1994; López-Sáez 1995; Pulido et al. 2008; Abel-Schaad et al. 2014b). Unfortunately, within the Spanish Central System we do not have detailed pollen records for the LGM and the Lateglacial period (López-Sáez et al. 2014). However, the discontinuous distribution of chestnut in the Gredos range today could be attributed to the great antiquity of the species in the region (López-Sáez et al. 1996). We can explain the origin of these scattered populations in two ways: (i) as local penetration of forests where chestnut is not present in a natural way, but rather through management by man which has allowed this species to thrive at the expense of deciduous oaks; (ii) as being the preservation of suitable refugia for survival of the diverse flora of mesic chestnut forests. It is important to note that the Iruelas Valley Natural Reserve meets one of the main requirements given by Krebs et al. (2004), since this potential chestnut refuge (microrefugia sensu Rull 2009) includes an area connected to orographic systems (Gredos range) and is located in a habitat with a favourable microclimate (Fig. 1). Therefore we can argue that the chestnut forest of El Tiemblo could have found a refuge area where the effects of cold and drought would be attenuated (Blanco et al. 1997) and have persisted unharmed for at least the last three millennia, thus representing a new “surprise” in the Late Holocene vegetation history of the Iberian Peninsula (Carrión et al. 2010). In short, considering the limitation that the pollen record from El Tiemblo covers only 3,140 years, this location of chestnut trees in the Iruelas Valley Natural Reserve could be considered as a microrefuge. At the moment, this statement is just a preliminary hypothesis which requires to be validated with new data.
Unfortunately, it is not possible to say on the basis of the available data whether the above mentioned pollen records were connected to a huge refuge covering the Gredos range. Fernández-López and Monteagudo (2010) have confirmed, according to genetic studies in Iberian chestnut populations, that the introduction of chestnuts into the Iberian Peninsula from other Mediterranean countries, entering through the South as has been usually hypothesized (Aravanopoulos et al. 2005), is not supported by their data as the highest allelic richness is in northern populations. In addition, these authors indicated a pattern of structured genetic variability caused by natural selection. Thus, the natural selection could be considered another evolutionary force acting over the different identified gene pools (Fernández-López et al. 2005), increasing the adaptation of populations to local conditions (e.g. El Tiemblo).
Conclusions
El Tiemblo peat bog was formed c. 3,140 cal bp in a landscape dominated by C. sativa. The data presented here provide the first evidence in the Iberian Peninsula of continued persistence of a dense and natural chestnut forest for at least three millennia.
Comparison with pollen data from the Spanish Central System suggests that chestnut forests had reduced their extension because C. sativa pollen percentages are generally low. Only from the Roman period and even more intensely during the Middle Ages, chestnut stands developed in this massif, probably through cultivation. However, the pollen record presented here is an exception to these facts. El Tiemblo is unique as a case of Late Holocene dominance of chestnut woodlands.
Palynological and charcoal data records show that vegetation of the Iruelas Valley Natural Reserve was highly resilient to fire disturbances and anthropic dynamics during the Late Holocene. Currently available data support that livestock and olive culture were practiced in the study area from the Roman period to the Early Modern period. However, its impact on vegetation and landscape seems to have been extremely low.
The pollen record from El Tiemblo peat bog strengthens the hypothesis of the existence of a refuge area localized in the foothills of the Gredos range in the Iruelas Valley Natural Reserve. Relationships between refuge areas and first evidence of chestnut usage in the whole of the Spanish Central System should be more deeply investigated. Similarly, it is necessary to find longer records and replicate these results with new pollen sequences in the area.
References
Aaby B, Berglund BE (1986) Characterization of the peat and lake deposits. In: Berglund BE (ed) Handbook of Holocene palaeoecology and palaeohydrology. Wiley, Chichester, pp 231–246
Abel-Schaad D, López-Sáez JA (2013) Vegetation changes in relation to fire history and human activities at the Peña Negra mire (Bejar Range, Iberian Central Mountain System, Spain) during the past 4000 years. Veget Hist Archaeobot 22:199–214
Abel-Schaad D, Hernández AM, López-Merino L, Pulido FJ, López-Sáez JA (2009a) Cabras y quemorros: tres siglos de cambios en el paisaje de la vertiente extremeña de la Sierra de Gredos. Rev Est Extremeños 65:449–478
Abel-Schaad D, Hernández AM, López-Sáez JA, Pulido FJ, López-Merino L, Martínez-Cortizas A (2009b) Evolución de la vegetación en la Sierra de Gata (Cáceres-Salamanca, España) durante el Holoceno reciente. Implicaciones biogeográficas. Rev Esp Micropal 41:91–105
Abel-Schaad D, López-Sáez JA, Pulido FJ (2014a) Heathlands, fire and grazing. A paleoenvironmental view of Las Hurdes (Cáceres, Spain) history during the last 1200 years. For Syst 23:247–258
Abel-Schaad D, Pulido FJ, López-Sáez JA et al (2014b) Persistence of tree relicts through the Holocene in the Spanish Central System. Lazaroa 35:107–131
Aira MJ, Barthelemy L (1990) Étude de la répartition de la pluie pollinique dans la Serra do Bocelo (Galice, Espagne). CR Soc Biogéogr 63:129–146
Álvarez-Sanchís JR (2000) The Iron Age in Western Spain (800 bc–ad 50): an overview. Oxford J Archaeol 19:65–89
Álvarez-Sanchís JR (2005) Oppida and Celtic society in western Spain. J Interdiscip Celtic Stud 6:255–285
Aravanopoulos FA, Bucci G, Akkak A et al (2005) Molecular population genetics and dynamics of chestnut (Castanea sativa) in Europe: inferences for gene conservation and tree improvement. Acta Hortic 693:403–412
Atienza M, Gómez-Lobo A, Ruiz-Zapata MB (1990) Estudio polínico de un depósito localizado en la Garganta del Trampal (Sierra de Béjar, Ávila). Actas de Gredos 10:19–23
Benito-Garzón M, Sánchez de Dios R, Sainz-Ollero H (2007) Predictive modelling of tree species distributions on the Iberian Peninsula during the Last Glacial Maximum and Mid-Holocene. Ecography 30:120–134
Bennett KD (1996) Determination of the number of zones in a biostratigraphical sequence. New Phytol 132:155–170
Berrocal M, Gallardo JF, Cardeñoso JM (1998) El castaño. Mundi-Prensa, Madrid
Blaauw M (2010) Methods and code for “classical” age-modelling of radiocarbon sequences. Quat Geochronol 5:512–518
Blanco E, Casado M, Costa M et al (1997) Los bosques ibéricos: una interpretación geobotánica. Planeta, Barcelona
Blanco-González A, López-Sáez JA, López-Merino L (2009) Ocupación y uso del territorio en el sector centromeridional de la cuenca del Duero entre la Antigüedad y la Alta Edad Media (siglos I-XI D.C.). Arch Esp Arqueol 82:275–300
Blanco-González A, López-Sáez JA, Alba F, Abel D, Pérez S (2015) Medieval landscapes in the Spanish Central System (450–1350): a palaeoenvironmental and historical perspective. J Mediev Iber Stud 7:1–17
Bounous G, Beccaro G (2002) Chestnut culture: directions for establishing new orchard. Nucis 11:30–34
Bounous G, Marinoni DT (2005) Chestnut: botany, horticulture and utilization. Hortic Rev 31:291–347
Carrión JS (1992) Late Quaternary pollen sequence from Carihuela Cave, southeastern Spain. Rev Palaeobot Palynol 71:37–77
Carrión JS, Navarro C, Navarro J, Munuera M (2000) The distribution of cluster pine (Pinus pinaster) in Spain as derived from palaeoecological data, relationships with physociological classification. Holocene 10:243–252
Carrión JS, Yll EI, Walker MJ, Legaz AJ, Chain C (2003) Glacial refugia of temperate, Mediterranean and Ibero-North African flora in south-eastern Spain: new evidence from cave pollen at two Neanderthal man sites. Glob Ecol Biogeogr 12:119–129
Carrión JS, Scott L, Arribas A, Fuentes N, Gil-Romera G, Montoya E (2007) Pleistocene landscapes in central Iberia inferred from pollen analysis of hyena coprolites. J Quat Sci 22:191–202
Carrión JS, Fernández S, González-Sampériz P et al (2010) Expected trends and surprises in the Lateglacial and Holocene vegetation history of the Iberian Peninsula and Balearic Islands. Rev Palaeobot Palynol 162:458–475
Chauchard S, Carcaillet C, Guibal F (2007) Patterns of land-use abandonment control tree-recruitment and forest dynamics in Mediterranean mountains. Ecosystems 10:936–948
Conedera M, Krebs P (2008) History, present situation and perspective of the chestnut cultivation in Europe. Acta Hortic 784:2–8
Conedera M, Stanga P, Oester B, Bachmann P (2001) Different post-culture dynamics in abandoned chestnut orchards. For Snow Landsc Res 76:487–492
Conedera M, Krebs P, Tinner W, Pradella M, Torriani D (2004a) The cultivation of Castanea sativa (Mill.) in Europe, from its origin to its diffusion on a continental scale. Veget Hist Archaeobot 13:161–179
Conedera M, Manetti MC, Giudici F, Amorini E (2004b) Distribution and economic potential of the sweet chestnut (Castanea sativa Mill.) in Europe. Ecol Mediterr 30:179–193
Conedera M, Tinner W, Crameri S, Torriani D, Herold A (2006) Taxon-related pollen source areas for lake basins in the southern Alps: an empirical approach. Veget Hist Archaeobot 15:263–272
Desprat S, Sánchez-Goñi MF, Loutre MF (2003) Revealing climatic variability of the last three millennia in northwestern Iberia using pollen influx data. Earth Planet Sci Lett 213:63–78
Di Pasquale G, Allevato E, Ermolli ER et al (2010) Reworking the idea of chestnut (Castanea sativa Mill.) cultivation in Roman times: new data from ancient Campania. Plant Biosyst 144:865–873
Dorado M, Valdeolmillos A, Ruiz-Zapata MB (2001) Actividad humana y dinámica de la vegetación en la Sierra de Ávila (Sistema Central Español) desde el Bronce Medio. Polen 11:39–49
Fabián J, Blanco-González A, López-Sáez JA (2006) La transición Calcolítico-Bronce Antiguo desde una perspectiva arqueológica y ambiental: el Valle Amblés (Ávila) como referencia. Arqueología Espacial 26:37–56
Fægri K, Iversen J (1989) In: Fægri K, Kaland PE, Krzywinski K (eds) Textbook of pollen analysis, 4th edn. Wiley, Chichester
Fernández-López J, Monteagudo AB (2010) Genetic structure of wild Spanish populations of Castanea sativa as revealed by isozyme analysis. For Syst 19:156–169
Fernández-López J, Zas R, Blanco-Silva R, Díaz R (2005) Geographic differentiation in adaptative traits of wild chestnut Spanish populations (Castanea sativa Miller). Invest Agrar: Sist Recur For 14:13–26
Fineschi S, Taurchini D, Villani F, Vendramin GG (2000) Chloroplast DNA polymorphism reveals little geographical structure in Castanea sativa Mill. (Fagaceae) throughout southern European countries. Mol Ecol 9:1,495–1,503
Franco-Múgica F, García-Antón M, Sainz-Ollero H (1997) Impacto antrópico y dinámica de la vegetación durante los últimos 2000 años BP en la vertiente septentrional de la Sierra de Gredos: Navarredonda (Ávila, España). Rev Paléobiol 16:29–45
Frei T (1997) Pollen distribution at high elevation in Switzerland: evidence for medium range transport. Grana 30:201–209
Gallardo J, Rico M, González MI (2000) Some ecological aspects of a chestnut coppice located at the Sierra de Gata mountains (Western Spain) and its relationship with a sustainable management. Ecol Mediterr 26:53–69
Gallardo-Lancho J (2001) Distribution of chestnut (Castanea sativa Mill.) forests in Spain: possible ecological criteria for quality and management (focusing on timber coppices). For Snow Landsc Res 76:477–481
Gandullo JM, Blanco A, Sánchez O, Rubio A, Elena R, Gómez V (2004) Las estaciones ecológicas de los castañares españoles., Monografías INIA Serie Forestal 7INIA, Madrid
García-Amorena I, Gómez-Manzaneque F, Rubiales JM, Granja HM, Soares de Carvalho G, Morla C (2007) The Late Quaternary coastal forests of western Iberia: a study of their macroremains. Palaeogeogr Palaeoclimatol Palaeoecol 254:448–461
García-Antón M, Morla C, Sainz H (1990) Consideraciones sobre la presencia de algunos vegetales relictos terciarios durante el Cuaternario en la Península Ibérica. Bol R Soc Esp Hist Nat Sec Biol 86:95–105
García-Garcimartín HJ (2002) Articulación jurisdiccional y dinámica socioeconomic de un espacio natural: la Cuenca del Alberche (siglos XII-XV). Ph.D. Dissertation, Universidad Complutense, Madrid
García-López JM, Allué-Camacho C (2008) Factorial ambits with high phytoclimatic viability for chestnut (Castanea sativa) in Spain. Acta Hortic 784:113–118
García-Mozo H, Domínguez-Vilches E, Galán C (2007) Airborne allergenic pollen in natural areas: Hornachuelos Natural Park, Córdoba, Southern Spain. Ann Agric Environ Med 14:63–69
Génova M, Gómez-Manzaneque F, Morla C (2009) Los bosques de Gredos a través del tiempo. Junta de Castilla y León, Valladolid
Goeury C, De Beaulieu JL (1979) À propos de la concentration du pollen à l’aide de la liqueur de Thoulet dans les sédiments minéraux. Pollen Spores 21:239–251
Gómez-Orellana L, Ramil-Rego P, Muñoz-Sobrino C (2007) The Würm in NW Iberia, a pollen record from Area Longa (Galicia). Quat Res 67:438–452
Grimm EC (1987) Coniss: a Fortran 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Comput Geosci 13:13–35
Grimm EC (1992) Tilia, version 2. Illinois State Museum, Research and Collection Center, Springfield
Grimm EC (2004) TGView. Illinois State Museum, Research and Collection Center, Springfield
Hernando MR (2002) Indigenismo y romanización del territorio abulense (s. V a.C.-s. III d.C.). Ph.D. Dissertation, Universidad Complutense, Madrid
Huntley B, Birks HJB (1983) An atlas of past and present pollen maps for Europe: 0–13000 years ago. Cambridge University Press, Cambridge
Krebs P, Conedera M, Pradella M, Torriani D, Felber M, Tinner W (2004) Quaternary refugia of the sweet chestnut (Castanea sativa Mill.): an extended palynological approach. Veget Hist Archaeobot 13:145–160
Krebs P, Koutsias N, Conedera M (2012) Modelling the eco-cultural niche of giant chestnut trees: new insights into land use history in southern Switzerland through distribution analysis of a living heritage. J Hist Geogr 38:372–386
Lauteri M, Pliura A, Monteverdi MC, Brugnoli E, Villani F, Eriksson G (2004) Genetic variation in carbon isotope discrimination in six European populations of Castanea sativa Mill. originating from contrasting localities. J Evol Biol 17:1,286–1,296
López-Merino L, López-Sáez JA, Sánchez-Palencia FJ, Reher GS, Pérez-Díaz S (2009a) Castaños, nogales y cereales: la antropización de los paisajes de Asturias y León en época romana. Cuad Soc Esp Cienc For 30:93–99
López-Merino L, López-Sáez JA, Alba-Sánchez F, Pérez-Díaz S, Carrión JS (2009b) 2000 years of pastoralism and fire shaping high-altitude vegetation of Sierra de Gredos in central Spain. Rev Palaeobot Palynol 158:42–51
López-Sáez JA (1995) Las comunidades de Prunus lusitanica L. del Valle del Tiétar (Sierra de Gredos, Ávila). Anales de Biología 20 (Biología Vegetal 9), pp 111–113
López-Sáez JA, Blanco-González A (2005) La mutación Bronce Final/Primer Hierro en el suroeste de la Cuenca del Duero (provincia de Ávila): ¿cambio ecológico y social? In: Blanco A, Cancelo C, Esparza A (eds) Bronce Final y Edad del Hierro en la Península Ibérica. Universidad de Salamanca, Salamanca, pp 229–250
López-Sáez JA, López-García P (1994) Contribution of the palaeoecological knowledge of Quaternary in the Tiétar Valley (Sierra de Gredos, Ávila, Spain). Rev Esp Micropal 26:61–66
López-Sáez JA, López-García P, Macías R (1991) Análisis polínico del yacimiento arqueológico de El Raso de Candeleda (Ávila). Actas de Gredos 11:39–44
López-Sáez JA, López-García P, Gómez C, Gil P (1996) Acerca del origen del castaño (Castanea sativa) en el valle del Tiétar (Sierra de Gredos, Ávila). In: Ruiz-Zapata MB (ed) Estudios Palinológicos. Universidad de Alcalá, Alcalá de Henares, pp 79–82
López-Sáez JA, Dorado M, Burjachs F, Ruiz-Zapata MB, López-García P, Fabián JF (2003) Paleoambiente y paleoeconomía durante la Prehistoria en el Valle Amblés (Ávila). Polen 13:129–141
López-Sáez JA, López-Merino L, Pérez-Díaz S (2008) Los vettones y sus paisajes: paleoambiente y paleoeconomía de los castros de Ávila. Zona Arqueológica 12:140–152
López-Sáez JA, Blanco-González A, López-Merino L et al (2009a) Landscape and climatic changes during the end of the Late Prehistory in the Amblés Valley (Ávila, central Spain), from 1200 to 400 cal bc. Quat Int 200:90–101
López-Sáez JA, López-Merino L, Alba-Sánchez F, Pérez-Díaz S (2009b) Contribución paleoambiental al estudio de la trashumancia en el sector abulense de la Sierra de Gredos. Hispania 231:9–38
López-Sáez JA, Alba-Sánchez F, López-Merino L, Pérez-Díaz S (2010a) Modern pollen analysis: a reliable tool for discriminating Quercus rotundifolia communities in Central Spain. Phytocoenologia 40:57–72
López-Sáez JA, López-Merino L, Alba-Sánchez F, Pérez-Díaz S, Abel-Schaad D, Carrión JS (2010b) Late Holocene ecological history of Pinus pinaster forests in the Sierra de Gredos of central Spain. Plant Ecol 206:195–209
López-Sáez JA, Sánchez-Mata D, Alba-Sánchez F, Abel-Schaad D, Gavilán RG, Pérez-Díaz S (2013) Discrimination of Scots pine forests in the Iberian Central System (Pinus sylvestris var. iberica) by means of pollen analysis. Phytosociological considerations. Lazaroa 34:191–208
López-Sáez JA, Abel-Schaad D, Pérez-Díaz S et al (2014) Vegetation history, climate and human impact in the Spanish Central System over the last 9,000 years. Quat Int 353:98–122
López-Sáez JA, Alba-Sánchez F, Sánchez-Mata D, Abel-Schaad D, Gavilán RG, Pérez-Díaz S (2015) A palynological approach to the study of Quercus pyrenaica forest communities in the Spanish Central System. Phytocoenologia 45:107–124
Luceño M, Vargas P (1991) Guía Botánica del Sistema Central Español. Anaya, Madrid
Manetti M, Amorini E, Becagli C, Conedera M, Giudici F (2001) Productive potential of chestnut (Castanea sativa Mill.) stands in Europe. For Snow Landsc Res 76:471–476
Manrique E, Fernández-Cancio A (2000) Extreme climatic events in dendroclimatic reconstructions from Spain. Clim Chang 44:123–138
Manzano E (1991) La frontera de al-Andalus en la época de los Omeyas. CSIC, Madrid
Mariné M (1995) El patrimonio arqueológico de la Sierra de Gredos. In: Troitiño MA (ed) Gredos: territorio, sociedad y cultura. Diputación de Ávila, Ávila, pp 19–48
Martín-Puertas C, Valero-Garcés BL, Mata MP, González-Sampériz P, Bao R, Moreno A, Stefanova V (2008) Arid and humid phases in southern Spain during the last 4000 years: the Zoñar Lake record, Córdoba. Holocene 18:907–921
Mattioni C, Cherubini M, Michelli E, Villani F, Bucci G (2008) Role of domestication in shaping Castanea sativa genetic variation in Europe. Tree Genet Genomes 4:563–574
Mattioni C, Martin MA, Cherubini M, Taurchini D, Villani F (2010) Genetic diversity in European chestnut populations. Acta Hortic 866:163–167
Mercuri AM, Bandini-Mazzanti M, Florenzano A, Montecchi MC, Rattighieri E (2013) Olea, Juglans and Castanea: the OJC group as pollen evidence of the development of human-induced environments in the Italian península. Quat Int 303:24–42
Molina-Moreno JR (1992) Flora y vegetación del valle de Iruelas (Ávila). Cuadernos Abulenses 18:11–150
Monsalvo JM (2003) Frontera pionera, monarquía en expansión y formación de los concejos de villa y tierra. Relaciones de poder en el realengo concejil entre el Duero y el Tajo (c. 1072-c. 1222). Arqueol Territ Med 10:45–126
Moore PD, Webb JA, Collinson ME (1991) Pollen analysis. Blackwell, London
Morales-Molino C, García-Antón M, Postigo-Mijarra JM, Morla C (2013) Holocene vegetation, fire and climate interactions on the westernmost fringe of the Mediterranean Basin. Quat Sci Rev 59:5–17
Morales-Molino C, Vescovi E, Krebs P et al (2015) The role of human-induced fire and sweet chestnut (Castanea sativa Mill.) cultivation on the long-term landscape dynamics of the southern Swiss Alps. Holocene 25:482–494
Morla C (1996) Especies forestales autóctonas y alóctonas en la restauración del tapiz vegetal de la península Ibérica. In: García-Fernández J (ed) Medio Ambiente y crisis rural. Universidad de Valladolid, Valladolid, pp 33–61
Muñoz-Sobrino C, Ramil-Rego P, Gómez-Orellana L (2004) Vegetation of the Lago de Sanabria area (NW Iberia) since the end of the Pleistocene: a palaeoecological reconstruction on the basis of two new pollen sequences. Veget Hist Archaeobot 13:1–22
Ninyerola M, Pons X, Roure JM (2005) Atlas Climático Digital de la Península Ibérica. Metodología y aplicaciones en bioclimatología y geobotánica. Universidad Autónoma de Barcelona, Bellaterra
Pardo F, Gil L (2005) The impact of traditional land use on woodlands: a case study in the Spanish Central System. J Hist Geogr 31:390–408
Peeters AG, Zoller H (1988) Long range of transport of Castanea sativa pollen. Grana 27:203–207
Pezzi G, Maresi G, Conedera M, Ferrari C (2011) Woody species composition of chestnut stands in the Northern Apennines: the result of 200 years of changes in land use. Landsc Ecol 26:1,463–1,476
Pitte JR (1986) Terres de castanides: Hommes et paysages du châtaignier de l’Antiquité à nos jours. Librairie Arthème Fayard, Evreux
Postigo-Mijarra JM, Gómez F, Morla C (2008) Survival and long-term maintenance of tertiary trees in the Iberian Peninsula during the Pleistocene: first record of Aesculus L. (Hippocastanaceae) in Spain. Veget Hist Archaeobot 17:351–364
Postigo-Mijarra JM, Morla C, Barrón E, Morales-Molino C, García S (2010) Patterns of extinction and persistence of Arctotertiary flora in Iberia during the Quaternary. Rev Palaeobot Palynol 162:416–426
Pulido F, Sanz R, Abel-Schaad D et al (2007) Los bosques de Extremadura, evolución, ecología y conservación. Junta de Extremadura, Mérida
Pulido F, Valladares F, Calleja JA, Moreno G, González-Bornay G (2008) Tertiary relict trees in a Mediterranean climate: abiotic constraints on the persistence of Prunus lusitanica at the eroding edge of its range. J Biogeogr 35:1,425–1,435
Reille M (1992) Pollen et Spores d’Europe et d’Afrique du Nord. Laboratoire de Botanique Historique et Palynologie, Marseille
Reimer PJ, Bard E, Bayliss A et al (2013) Intcal13 and marine13 radiocarbon age calibration curves 0–50,000 years cal bp. Radiocarbon 55:1,869–1,887
Rubiales JM, Hernández L, Romero F, Sanz C (2011) The use of forest resources in central Iberia during the Late Iron Age. Insights from the wood charcoal of Pintia, a Vaccaean oppidum. J Archaeol Sci 38:1–10
Rubio A, Sánchez-Mata D (1995) Consideraciones edafogeobotánicas sobre los castañares occidentales del Sistema Central Ibérico. In: Actas XX Reunión Nacional sobre Suelos: Degradación y conservación de suelos. Madrid, pp 265–273
Rubio A, Escudero A, Gandullo JM (1997) Sweet chestnut silviculture in an ecological extreme of its range in the West of Spain (Extremadura). Ann Sci For 54:667–680
Rubio A, Gavilán R, Escudero A (1999) Are soil characteristics and understorey composition controlled by forest management? For Ecol Manag 113:191–200
Ruiz de la Torre J (2006) Flora Mayor. MIMAM, Madrid
Rull V (2009) Microrefugia. J Biogeogr 36:481–484
Scarascia-Mugnozza G, Oswald H, Piussi P, Radoglou K (2000) Forests of the Mediterranean region: gaps in knowledge and research needs. For Ecol Manag 132:97–109
Seijo F, Millington JDA, Gray R et al (2015) Forgetting fire: traditional fire knowledge in two chestnut forest ecosystems of the Iberian Peninsula and its implications for European fire management policy. Land Use Policy 47:130–144
Stockmarr J (1971) Tablets with spores used in absolute pollen analysis. Pollen Spores 13:614–621
Taveira C (1979) Enfermedad de la tinta del castaño. Bol Serv Plagas 5:59–66
Van Benthem F, Clarke GCS, Punt W (1984) The Northwest European Flora, 33. Fagaceae. Rev Palaeobot Palynol 42:87–110
Van Geel B (2001) Non-pollen palynomorphs. In: Smol JP, Birks HJB, Last WM (eds) Tracking environmental change using lake sediments. Terrestrial, algal, and siliceous indicators, vol 3. Kluwer, Dordrecht, pp 99–119
Van Geel B, Berglund BE (2000) A causal link between a climate deterioration around 850 cal bc as a subsequent rise in human population density in NW-Europe? Terra Nostra 7:126–130
Van Geel B, van der Plicht J, Kilian MR et al (1998) The sharp rise of Δ14C ca. 800 cal bc: possible causes, related climatic teleconnections and the impact on human environments. Radiocarbon 40:535–550
Villani F, Sansotta A, Cherubini M, Cesaroni D, Sbordoni V (1999) Genetic structure of Castanea sativa in Turkey: evidence of a hybrid zone. J Evol Biol 12:233–244
Whitlock C, Larsen C (2001) Charcoal as a fire proxy. In: Smol JP, Birks HJB, Last WM (eds) Tracking environmental change using lake sediments. Terrestrial, algal, and siliceous indicators, vol 3. Kluwer, Dordrecht, pp 75–97
Acknowledgments
We sincerely thank three anonymous reviewers and the editor for their constructive suggestions and comments. This work was funded by the project HAR2013-43701-P (Spanish Economy and Competitiveness Ministry) “Dinámicas socio-ecológicas, resiliencia y vulnerabilidad en un paisaje de montaña: el Sistema Central (9,000 cal bc-1850 cal ad) (DESIRÈ)”. J.A. López-Sáez is currently supported by a research Grant (Spanish Ministry of Education, Culture and Sport) at Caen Basse-Normandie University. We are grateful to A. López Andreu (RIP) and P. Sáez Navas for their collaboration and assistance during the field work.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by W. Tinner.
Rights and permissions
About this article

Cite this article
López-Sáez, J.A., Glais, A., Robles-López, S. et al. Unraveling the naturalness of sweet chestnut forests (Castanea sativa Mill.) in central Spain. Veget Hist Archaeobot 26, 167–182 (2017). https://doi.org/10.1007/s00334-016-0575-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-016-0575-x