
Abstract
Pink salmon, Oncorhynchus gorbuscha (Walbaum, 1792), a native of the North Pacific Ocean, is an invasive fish species rapidly expanding in the North Atlantic, the adjacent waters of the Arctic Ocean, and rivers in their watersheds. We investigated the species composition of parasites and the structure of parasite communities of the introduced pink salmon in Kandalaksha Bay of the White Sea in 2021. The parasites were identified using morphological and genetic methods. A total of 14 parasite species were recorded. The core species of the parasitic fauna of the introduced pink salmon in the Kandalaksha Bay were Clistobothrium sp., ʽDiphyllobothriumʼ spp., Scolex polymorphus Rudolphi, 1819 (plerocercoids), Brachyphallus crenatus (Rudolphi, 1802), Derogenes varicus (Müller, 1784), Lecithaster salmonis Yamaguti, 1934 (adults), Anisakis simplex (Rudolphi, 1809) sensu stricto (juvenile III), and Hysterothylacium aduncum (Rudolphi, 1802) (juveniles III and IV). We provided the first evidence that the Pacific genetic lineage of A. simplex sensu stricto occurs in European marine waters. In 2021, there was a noticeable increase in the mean abundance of the third-stage juveniles of Anisakis in the White Sea pink salmon compared to the period of 1990s-early 2000s. We conclude that the pink salmon occupies the same trophic niche in the new region of its introduction as in the native range.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The pink salmon, Oncorhynchus gorbuscha (Walbaum, 1792), is a monocyclic species with the shortest freshwater stage of the life cycle among all members of the Salmonidae (Zueva 1963; Heard 1991). Due to a rigid 2-year life cycle, generations of this fish species spawning in odd and in even years represent two reproductively isolated lineages. The native range of the pink salmon encompasses most of the North Pacific with the adjacent Arctic Ocean waters and rivers in their watersheds (Heard 1991; Babaluk et al. 2000; Chereshnev et al. 2002).
In the second half of the twentieth century, several attempts have been made to introduce the pink salmon into the watersheds of the White and the Barents Seas (Azbelev and Yakovenko 1963; Karpevich et al. 1991; Alekseev and Kulachkova 2000; Kudersky 2005). For this purpose, fertilized eggs were repeatedly imported and incubated in fish hatcheries in the Murmansk and the Arkhangelsk Regions of Russia (Karpevich et al. 1991; Kudersky 2005). However, steady populations spawning naturally in the newly acquired range did not form until the late 1980s. At present, only odd-year generations of the introduced pink salmon reach commercial abundance (Kudersky 2005).
The non-native range of the pink salmon is currently in the North Atlantic, western areas of the Arctic Ocean, and rivers in their watersheds (Bogdanov and Kizhevatov 2015; Armstrong et al. 2018; Alekseev et al. 2019; Sandlund et al. 2019; Nielsen et al. 2020; Paulsen et al. 2022). Pink salmon has reportedly increased in abundance and expanded its range lately, due to the warming of the North Atlantic and the Arctic (Armstrong et al. 2018). This process has stimulated the research of the ecological implications of the pink salmon introductions (International seminar… 2021).
Parasitological studies are an important area of invasion ecology (Dunn 2009). The introduction of a new animal or plant in an area may enhance (spillback effect and spillover effect) or weaken (dilution effect) the parasite load on the native hosts (Tompkins and Poulin 2006; Kelly et al. 2009; Lymbery et al. 2014). The pink salmon in Northern Europe provides a vivid example that the colonization of new habitats by an introduced species can be accompanied by the formation of the parasitic community consisting only of native parasites. The first data on the parasite fauna of the pink salmon introduced into Northern Europe were published by Ninburg (1963), who found 14 parasite species in the fish ascending the Kola River (Barents Sea watersheds). Later on, repeated surveys of the pink salmon in the Barents Sea basin were performed (Mitenev and Karasev 1995; Mitenev and Shulman 1999). Data on pink salmon parasites are also available for the watersheds of the White Sea (Mitenev and Karasev 1995; Mitenev and Shulman 1999; Malakhova 1972; Grozdilova 1974; Kulachkova 2001; Barskaya et al. 2005; Ieshko et al. 2016; Krupenko et al. 2022), the Norwegian Sea (Fjær 2019; Rullestad 2021), and the Kara Sea (Gavrilov 2016).
How does the increase in abundance of introduced pink salmon observed in recent years affect the diversity of the parasites colonizing them? The data obtained during the long-term parasitological monitoring of the White Sea pink salmon provide valuable information for assessing the trends in the dynamics of parasite diversity of this introduced species. In this study, we examined the composition of metazoan parasites of the pink salmon from the Kandalaksha Bay (White Sea) with the aim of assessing the current parasitic load on this species.
Materials and methods
Samples
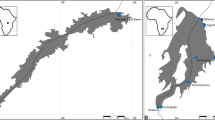
Twenty-five specimens of the pink salmon were caught in the Kandalaksha Bay (Fig. 1) of the White Sea (66° 55′ N; 32° 45′ E), July 2021. They were frozen and dissected some time later. The presence of ecto- and endoparasites was ascertained using a standard method of parasitological examination (Klimpel et al. 2019).

Map of the Kandalaksha Bay, where pink salmon was collected for parasitological studies
All parasites were fixed in 70% ethanol and 3 days later were transferred to 96% ethanol for long-term storage. For morphological study, digeneans were stained with acetic carmine, dehydrated, contrasted (cleared) with dimethyl phthalate, and finally mounted in the Canada balsam; nematode specimens were mounted in glycerol.
Molecular data
Molecular data were obtained for anisakid juveniles III, as well as diphyllobothriid, bothriocephalidean, and phyllobothriidean plerocercoids, whose species identification by morphological characters is unreliable or impossible. Whole DNA was extracted from single ethanol-fixed specimen of the nematodes and the cestodes using DNeasy Blood & Tissue kits (Qiagen, Hilden, Germany) following the manufacturer’s protocol with overnight lysis and double elution for increased DNA yield. A total of 12 anisakid juveniles III and 10 metacestode specimens were studied molecularly. For anisakid juveniles III, the mitochondrial cytochrome c oxidase subunit II (cox2) gene was amplified using the primers 211F (5′-TTTTCTAGTTATATAGA TTGRTTTYAT-3′) and 210R (5′-CACCAACTCTTAAAATT ATC-3′) (Mattiucci et al. 2014). Polymerase chain reaction (PCR) was carried out according to Mattiucci et al. (2014). For diphyllobothriid, bothriocephalidean and phyllobothriidean plerocercoids, two molecular markers were used: the large subunit nuclear ribosomal RNA gene (28S rRNA) and the cytochrome oxidase subunit I (cox1) mitochondrial gene. Partial 28S rRNA gene sequences (ca. 1000 bp) were amplified using the primer set: ZX-1 (5ʹ-ACCCGCTGAATTTAAGCATAT-3ʹ), 1500R (5ʹ-GCTATCCTGAGGGAAACTTCG-3ʹ), LSU_300F (5ʹ-CAAGTACCGTGAGGGAAAGTTG3ʹ), 1090F (5ʹ-TGAAACACGGACCAAGG-3ʹ), LSU_1200F (5ʹ-CCCGAAAGATGGTGAACTATGC-3ʹ), and ECD2 (5ʹ-CTTGGTCCGTGTTTCAAGACGGG3ʹ) (Waeschenbach and Littlewood 2017). Partial cox1 (ca. 550 bp) was amplified using the primers PBIcox1F_PCR (5′-CATTTTGCTGCCGGTCARCAYATGTTYTGRTTTTTTGG-3′), PBIcox1R_PCR (5′-CCTTTGTCGATACTGCCAAARTAATGCATDGGRAA-3′) (Waeschenbach and Littlewood 2017). PCR were performed as described by Waeschenbach and Littlewood (2017). Sequences of the cox2 and cox1 genes have a high resolving power to identify species in anisakids and metacestodes, respectively, while the 28S rRNA gene sequences can be used as a marker for analysis of evolutionary events at the genus level and above (e.g., Mattiucci and Nascetti 2008; Hernández-Orts et al. 2021).
PCR products were purified and sequenced bidirectionally using an Applied Biosystems BigDye Terminator v3.1 Cycle Sequencing Kit (Thermo Fisher Scientific Inc., Waltham, MA) with the PCR primers for cox2 and 28S rRNA, and with sequencing primers PBI-cox1F_seq (5′-CATTTTGCTGCCGGTCA-3′), PBI–cox1R_seq (5′-TAATGCATDGGRAAAAAAC-3′) for cox1.
The sequences were edited and aligned using Geneious, vers. 9.1.8 (Kearse et al. 2012) to obtain consensus sequences and to check for deletions, insertions, and stop codons. All the obtained sequences were deposited in GenBank (OP946146–OP946157, OP933782–OP933788, OP935162–OP935169). Haplotypes of cox2 were defined by using FaBox (Villesen 2007).
To identify the place of our specimens of Clistobothrium Dailey & Vogelbein, 1990 and Diphyllobothrium Cobbold, 1858 among other phyllobothriid and diphyllobothriid taxa available in GenBank (Bouzid et al. 2008; Arizono et al. 2009; Waeschenbach et al. 2012, 2017; Yamasaki et al. 2012a, b; Kuchta et al. 2013; Ebmer et al. 2020; Fraija-Fernández et al. 2021; Hernández-Orts et al. 2021; Lozano-Cobo et al. 2021), Bayesian inference (BI) analysis was performed for the cox1 mtDNA dataset. The TIM2 + I + G (Phyllobothriidae) and TIM1 + I + G (Diphyllobothriidae) models were chosen as the best nucleotide substitution model under the Akaike information criterion using jModelTest 2.1.10 software (Darriba et al. 2012).
Bayesian inference analyses were performed in BEAST 3.2.6 (Drummond and Rambaut 2007) with standard settings for 10,000,000 generations with a sampling frequency of 1000 generations. Burn-in periods were set to 10% of generations. The resulting tree was visualized in FigTree 1.4.4 (Rambaut 2018). Nomenclature of diphyllobothriid taxa followed the latest study by Hernández-Orts et al. (2021).
Terminology and statistical analyses
In accordance with the established approaches to the description of parasite communities, we used the terms infracommunity (all parasites of a particular host specimen) and component community (all parasites in the host population) (Bush and Holmes 1986; Holmes and Price 1986). We also used basic parameters describing the parasitic load on the host: the prevalence of parasites (P), the intensity of infection and the mean abundance (M) (Bush et al. 1997). The parasites species were classified as the core species and the satellite species in accordance with Hanski (1982). Prevalence was used as a key parameter for this classification.
For a better understanding of the structuring of the parasitic communities, we assessed variation in fish specimens’ infection depending on the size of pink salmons. The frequencies of size–weight indicators of the captured pink salmon individuals were checked for compliance with the normal distribution according to the Kolmogorov–Smirnov test with Lilliefors modification (KSL).
Species richness was described by cumulative curves s = anb, where a predicts the number of species in the parasite infracommunity of the first fish under study, b characterizes the growth rate of the number of parasite species s with the increase in the number of the studied fish n. A change in the order in which fish were included in the initial sample results in a different series of accumulation of species richness of the parasites. We calculated 100 variants of shuffled rows taking the mean values of a and b for the final equations (Ieshko et al. 2019). The frequency distributions of the parameters were compared using the Kolmogorov–Smirnov test (KS). Variables of variability ranges were measured by the simple non-parametric bootstrap with the number of tests B = 1000. Confidence ranges were set using the percentile method. Indicators were compared with the use of the randomization test (Monte Carlo Randomization, MCR) with the same number of iterations B. The significance of differences p in this approach is an adjusted share of null model combinations (empirical difference no greater than randomized, |dobs| ≤|dran|) of the total number of tests B. The standard value α = 0.05 was taken as the critical value for p (Shitikov and Rosenberg 2013). The use of iterative procedures (randomization, bootstrap), free from the requirement of distribution normality, allowed us to analyze samples of different size, including small samples. The data were processed in MS Excel and R 4.0.1 (R Core Team 2022) with the use of basic functions.
Results
A total of 14 parasites species were identified in the pink salmon spawning in the rivers flowing into Kandalaksha Bay (Table 1). Eight of them were identified based on the morphological data only, while six parasites species were identified based on a combination of morphological and molecular data.
The sequence of cox2 in the third-stage juvenile morphologically assigned to Phocanema Myers, 1959 (575 bp, GenBank accession no. OP946157) matched the sequence of Phocanema bulbosum (Cobb, 1888) (GenBank accession no. KU558720) from Newfoundland, the similarity being 99.6%. All 11 cox2 sequences obtained from the third-stage juveniles morphologically assigned to Anisakis Dujardin, 1845 (574 bp, GenBank accession nos. OP946146–OP946156) matched the sequences of Anisakis simplex (Rudolphi, 1809) sensu stricto. Ten of them matched the sequences from the Norwegian Sea, the Baltic Sea, and the Cantabrian Sea, the similarity being 99.5–100%. However, one sequence (GenBank accession no. OP946147) matched (similarity 99.6%) two sequences obtained from specimens from the North Pacific (GenBank accession no. KC633441, LC543811).
Sequences of the 28S rRNA (GenBank accession nos. OP935162, OP935167, OP935168) gene obtained from three phyllobothriidean plerocercoids genotyped in our study, which had an apical sucker and four long foliose bothridia with an anterior round muscular accessory sucker and posterior folding loculus, were very similar to those of Clistobothrium sp. of Guardone et al. (2020) (= Clistobothrium sp. 1 of Caira et al. (2020)) (BLAST identity indices were 100%). One of these three phyllobothriidean plerocercoids was genotyped on the cox1 gene. In the phylogenetic tree based on this marker, the mentioned plerocercoid is in the same clade as Clistobothrium sp. of Guardone et al. (2020) (Fig. 2). Thus, the plerocercoid specimens found in the current study with the morphological features having been described above were identified as Clistobothrium sp. of Guardone et al. (2020). BLAST analysis of 28S rRNA (GenBank accession nos. OP935165, OP935169) and cox1 (GenBank accession nos. OP933786, OP933788) gene sequences of two Eubothrium plerocercoids genotyped showed a match with Eubothrium crassum (Bloch, 1779) (100% and 99.3–99.4%, respectively).

Phylogenetic relationships of phyllobothriid plerocercoids from the pink salmon caught in the Kandalaksha Bay based on Bayesian analysis of cox1 gene sequences data. Newly obtained sequences are in bold. Numbers represent posterior probabilities from Bayesian inference analysis (> 0.95 shown only)
One specimen of the five diphyllobothriid plerocercoids genotyped in our study was identified as ʽDiphyllobothriumʼ schistochilus (Germanos, 1895) based on both 28S rDNA (GenBank accession no. OP935166) and cox1 (GenBank accession no. OP933787) sequences, with the matching of 100% and 99.6%, respectively. Four diphyllobothriid specimens were classified as ʽDiphyllobothriumʼ sp. (GenBank accession nos. for 28S rDNA sequences: OP935163, OP935164, for cox 1 sequences: OP933783–OP933785). Their taxonomic status needs to be clarified in future phylogenetic studies. On the phylogenetic tree based on BI analyses of cox1 sequences, ʽDiphyllobothriumʼ sp. formed a well-supported sister taxon to ʽDiphyllobothriumʼ sprakeri Hernández-Orts, Kuzmina, Gomez-Puerta & Kuchta, 2021 (Fig. 3).

Phylogenetic relationships of diphyllobothriid plerocercoids from the pink salmon caught in the Kandalaksha Bay based on Bayesian analysis of cox1 gene sequences data. Newly obtained sequences are in bold. Numbers represent posterior probabilities from Bayesian inference analysis (> 0.95 shown only)
Based on the prevalence values, the parasites found in our study could be divided into two groups: the core species of the component community and the satellite species. The actual values of prevalence were higher than the mean value (60.6) in the core species and lower than that in the satellite species (Table 1).
The distribution of the fish by length and body weight did not correspond to the normal law (KSL = 0.248, p < 0.001; KSL = 0.187, p = 0.042), demonstrating a possible heterogeneity of the sample. After decomposition, the optimal result was the division of the initial frequencies of the two characters into two empirical distributions that did not differ significantly from the normal one (KSL = 0.217, p = 0.130; KSL = 0.194, p = 0.075 for body total length and KSL = 0.139, p = 0.977; KSL = 0.154, p = 0.269 for body weight). The first empirical distribution included relatively small fish individuals (n = 7) with a length of up to 45 cm (39–45, mean 42.7) and a body weight of up to 1000 g (755–1000, mean 875.7), while the second one included larger individuals (n = 18) with body total length 47–53 cm, mean 49.5 cm, and body weight 1100–1750 g, mean 1337.4 g. The small fishes differed from the large ones in the length and the body weight (MCR test: p < 0.001), while the differences in the sex were insignificant (MCR test: p > 0.150).
The samples of the small and the large individuals of the pink salmon had an identical composition of the core group of the parasites. The abundance of parasite individuals in each studied host fish was counted only for the six core species (Table 2).
The prevalence of these six core species in the small and the large fish groups did not differ significantly (MCR test: p > 0.071) (Table 2). The abundance distribution of these six species across the host individuals was very uneven: the value of aggregation index (ratio of variance to arithmetic mean, DM−1) varied in the range of 10.1–188.2 in the small fish group and in the range of 24.0–374.6 in the large fish group. The mean abundance of most of the core species of parasites in large pink salmon individuals was 2–5 times higher than in small ones, but due to the strong variability in the abundance values the statistical differences were also not significant (MCR test: p > 0.081).
Differences between abundance distributions in small and large individuals of the host were significant only for Derogenes varicus (Müller, 1784) (Kolmogorov–Smirnov test: KS = 0.635, p = 0.021).
The species richness accumulation models plotted for the two size groups of the pink salmon differed from each other. The most likely number of the parasite species forming the infracommunity in the first captured individual (coefficient a of the regression model s = anb) in the large fish group was significantly greater than in the small fish group (MCR test: p < 0.001) (Table 3, Fig. 4A). Mean rates of species accumulation associated with the increasing number of examined host individuals (coefficient b of the regression model s = anb) were also higher in the large fish group (MCR test: p < 0.001). At the same time, the results of studying the series of model parameters obtained in the process of resampling indicate the differentiation of the values of coefficient b in the small fish group: while the prevailing values of this parameter were low, in quite a few sample combinations the growth rates of the number of parasite species with the increasing number of the examined host individuals predicted by the model were similar to those in the large fish group (Fig. 4B).

Smoothed distributions of the regression model coefficients s = anb calculated for 100 variants for accumulation of parasites in two size groups of pink salmon from the Kandalaksha Bay: a distribution of coefficient a; b distribution of coefficient b. Differences of distributions are significant (Kolmogorov–Smirnov MCR test: KS > 0.43; p < 0.001). 1 = host size group > 45 cm; 2 = host size group ≤ 45 cm; KS = Kolmogorov–Smirnov; MCR = Monte Carlo Randomization
Discussion
The parasite fauna of the pink salmon in our survey was represented only by species belonging to the marine ecological group. This is unusual because both marine and freshwater species are mentioned in most of the previous parasitic studies of the introduced pink salmon (Ninburg 1963; Malakhova 1972; Grozdilova 1974; Mitenev and Karasev 1995; Mitenev and Shulman 1999; Barskaya et al. 2005; Gavrilov 2016; Ieshko et al. 2016; Fjær 2019). These results can be explained by the features of the pink salmon life cycle, namely, the downstream migration of the early larval stages, which prevents the fish from getting infected by freshwater species (except for parasitic protists, which are not retained during the marine period), and the anadromous migration of mature fish. So, the freshwater parasites recorded in the mature pink salmon in the literature mostly infect it during the spawning run into rivers. However, it cannot be ruled out that the pink salmon is re-infected with some endohelminths of freshwater group such as Dibothriocephalus ditremus (Creplin, 1825) and Dibothriocephalus dendriticus (Nitzsch, 1824) in the sea, when feeding on the young of other fish that has run there (Ninburg 1963; Mitenev and Karasev 1995; Mitenev and Shulman 1999). We believe that the difference between our results and data from the literature regarding the freshwater component of the pink salmon parasite fauna arises from nuances of the sampling. All researchers who reported freshwater parasites in mature pink salmon took samples in the lower course of the spawning rivers (Ninburg 1963; Malakhova 1972; Grozdilova 1974; Mitenev and Karasev 1995; Mitenev and Shulman 1999; Barskaya et al. 2005; Gavrilov 2016; Ieshko et al. 2016; Fjær 2019), while we sampled the fish in the Kandalaksha Bay, before it entered the rivers.
Three parasite species, Clistobothrium sp. of Guardone et al. (2020), ʽDiphyllobothriumʼ sp., and P. bulbosum, were found in the introduced pink salmon for the first time. Their detection became possible owing to the use of advanced identification techniques. Juveniles of P. bulbosum have previously been reported from non-salmonid fishes of the North Atlantic and adjacent areas of the Arctic Ocean (Paggi et al. 1991; Najda et al. 2018). Plerocercoids of Clistobothrium sp. of Guardone et al. (2020) have previously been detected in cephalopods, sharks and marine actinopterygians from various areas of the Atlantic and waters of Santa Catalina Island (Pacific Ocean) and South Africa (probably Indian Ocean) (Brickle et al. 2001; Kuris et al. 2015; Caira et al. 2020; Guardone et al. 2020) and adults in sharks Lamna nasus (Bonnaterre, 1788) and Isurus oxyrinchus (Rafinesque, 1810) from the Atlantic (Olson et al. 2001; Randhawa and Brickle 2011; Caira et al. 2020). We obtained the first evidence of the presence of the Pacific genetic lineage of A. simplex sensu stricto in European marine waters (one “Pacific-like” haplotype out of the 11 haplotypes obtained). This finding contradicts the conclusions about a clear genetic segregation between the Pacific and the Atlantic isolates of this species (Kijewska et al. 2009).
Overall, the parasitic fauna of the pink salmon in non-native range had a poorer species composition than that of the pink salmon in its native range in the North Pacific: 50 vs. 83 (Mamaev et al. 1959; Strelkov 1960; Ninburg 1963; Malakhova 1972; Grozdilova 1974; Love and Moser 1983; Nagasawa et al. 1987; Mitenev and Karasev 1995; Mitenev and Shulman 1999; Ermolenko et al. 1998; Kulachkova 2001; Barskaya et al. 2005; Ieshko et al. 2016; Gavrilov 2016; Fjær 2019; Rullestad 2021; Krupenko et al. 2022). This observation agrees with the literature data that parasite species richness or prevalence is lower in host populations in invasive ranges as compared to those in the native range (Torchin and Lafferty 2009; Sarabeev et al. 2017).
Analysis of the initial pink salmon sample collected by us revealed eight core parasite species. Three of these species, taking into account the taxonomic re-evaluation of specimens of Lecithaster Lühe, 1901 (see below), are the core species (actual p value > mean p value) in the parasite communities of the White Sea pink salmon recorded in all the previous study periods of this introduced species: Brachyphallus crenatus (Rudolphi, 1802), Lecithaster salmonis Yamaguti, 1934, and plerocercoids of the collective name of Scolex spp. (Malakhova 1972; Grozdilova 1974; Barskaya et al. 2005; Ieshko et al. 2016). Molecular phylogenetic data of Krupenko et al. (2022) on the species affiliation of Lecithaster Lühe, 1901 from the White Sea suggest that specimens of this genus previously recorded in introduced pink salmon (Malakhova 1972; Grozdilova 1974; Barskaya et al. 2005; Ieshko et al. 2016) are probably conspecific with L. salmonis. It is also likely that at least some of the S. pleuronectis specimens detected in the White Sea pink salmon prior to the “molecular taxonomy” era (Malakhova 1972; Grozdilova 1974; Barskaya et al. 2005; Ieshko et al. 2016) are conspecific with Clistobothrium sp. of Guardone et al. (2020) plerocercoids in our study. Diphyllobothriid plerocercoids and D. varicus have been mentioned as the core parasites of the White Sea pink salmon by Grozdilova (1974), Barskaya et al. (2005), and Ieshko et al. (2016); Hysterothylacium aduncum (Rudolphi, 1802), by Malakhova (1972), Barskaya et al. (2005), and Ieshko et al. (2016); juveniles of Anisakis, by Grozdilova (1974). Thus, during the time that has elapsed since the first study of the White Sea pink salmon (1961), that is, slightly more than 60 years, the composition of its core parasite species has included both stable and fluctuating components.
Among the eight core parasite species of the White Sea pink salmon revealed in our study, a consistent analysis of the long-term dynamics of the mean abundance is possible only for third-stage juveniles of Anisakis. The abundance of other histozoic parasites from the cores species group (diphyllobothriid plerocercoids) was not estimated in our study. The abundance of intestinal endohelminths depends on the duration spent by anadromous pink salmon in fresh water. Therefore, the previous data collected during the study of fish in fresh water do not reflect the actual abundance of these parasites before the hosts enter the river. Given certain requirements for fish samples (n ≥ 10 specimens, odd years), the dynamics of anisakid abundance in the White Sea pink salmon can be traced using only data from 1965, 1991, 1993, 1995, 1997, 1999, 2003, 2021 (Grozdilova 1974; Kulachkova 2001; Barskaya et al. 2005; Ieshko et al. 2016; present data). Previously, Kulachkova (2001) revealed a downward trend in the mean abundance of Anisakis spp. in these fish in the period from the 1960s to the 1990s (from 6 to 0.3–1.2 ind). The reasons for the decline of the White Sea pink salmon infection with Anisakis spp. have not been elucidated. Kulachkova (2001) attributed the trend she revealed to the decline in the numbers of the harp seal, Pagophilus groenlandicus (Erxleben, 1777), in the White Sea in the last quarter of the twentieth century. This explanation, however, is incorrect, since the obligate definitive hosts for Anisakis spp. are cetaceans (Mattiucci et al. 2017). Expansion of the dataset (Barskaya et al. 2005; Ieshko et al. 2016; present data) showed that the mean abundance of third-stage juveniles of Anisakis in the White Sea pink salmon was relatively low during the 1990s and in the early 2000s (0.3–1.4 ind.). In 2021, however, this infection parameter increased notably (2.4 ind.).
During the marine life stage, the pink salmon individuals are notably segregated by body size both within one generation and between adjacent (even–odd) generations (Azbelev and Yakovenko 1963; Utne 2021). As stated above, we examined two size groups of the pink salmon, differing significantly in length and weight (Table 2). The core of the component parasite community in both size groups of the pink salmon was made up of species whose life cycles involve pelagic animals such as planktonic crustaceans and young marine fish as intermediate or paratenic hosts (Køie 1979; Delyamure et al. 1985; Klimpel et al. 2004; Klimpel and Rückert 2005; Sokolov et al. 2016; Klotz et al. 2018; Krupenko et al. 2022). Based on the data on the core parasite species obtained in our study and the information on the diet of pink salmon in the Norwegian Sea (Utne 2021), we can describe the introduced pink salmon as a pelagic euryphagous fish showing a tendency to consume zooplankton and micronekton. This means that, when introduced to a new region, pink salmon occupies the same trophic niche as its marine life cycle stage in the native range (compare with Karpenko et al. 2007; Graham et al. 2021). According to Barskaya et al. (2005), the species composition of marine parasites of introduced pink salmon has a high degree of similarity with that of anadromous Atlantic salmon, Salmo salar Linnaeus, 1758, which indicates the similarity of the food base of these fish during the marine feeding period. Recently, there has been a sharp decrease in the abundance of anadromous S. salar throughout the range (ICES 2018). One of the key hypotheses explaining the decline of this fish species is an increased interspecific competition with marine pelagic fish (Potter and Crozier 2000). Utne et al. (2021) studied interactions between salmon postsmolts and some other pelagic fish (Scomber scombrus Linnaeus, 1758 and Clupea harengus Linnaeus, 1758) in the Northeast Atlantic and found no evidence to support the hypothesis. The feeding interactions between the pink salmon and Atlantic salmon require a close examination in the light of the hypothesis under consideration.
The parameters a and b of an exponential equation describing the parasite species accumulation, s = anb, are generalized metrics of the species richness and β-diversity of infracommunities (Dove and Cribb 2006; Ieshko et al. 2019). We have identified a trend towards an increase in species richness and β-diversity of parasite infracommunities in the pink salmons with a body length > 45 cm as compared to specimens with a body length ≤ 45 cm (Table 3). In addition, we detected a lower level of infection in the host specimens with a body length ≤ 45 cm by D. varicus compared to the larger pink salmons. These differences are probably associated with the difference in feeding intensity and species composition of food objects. The heterogeneity of the resampled values of the average rate of parasite species accumulation, the model parameter b, for small fish group noted in our study (Fig. 4B) may be either an effect of randomness or that of an actual differentiation of parasite species richness. This issue requires verification in future studies based on a larger sample. Reports of a dependence of the infection rate of fish individuals on their size are frequent in the literature (e.g., Zdzitowiecki 1988; Oliva et al. 1990; Muzzall and Bowen 2002; Brickle et al. 2006). Our data demonstrate that the size structure of host samples should be given closer attention in further parasitological studies of this fish species.
Some authors have pointed out potential parasitological risks associated with increasing numbers of pink salmon in recipient freshwater and marine environments (Sandlund et al. 2019; Hindar et al. 2020). These concerns were probably evoked by cases of parasite spillback and spillover upon animal introductions (Kelly et al. 2009; Paterson et al. 2013; Lymbery et al. 2014). The data on parasites of introduced pink salmon available so far are insufficient for discussing these risks.
Data availability
All data used in this study are included within this paper and they can be available upon request.
References
Alekseev AP, Kulachkova VG (2000) Far-Eastern pink salmon in the basins of the White and Barents seas (The second, “Magadan” stage of acclimatization). In: Matishov GG (ed) Vidy-vselentsy v evropeiskikh moryakh Rossii. Kol’skiy nauchnyy tsentr RAN, Apatity, pp 249–257 (In Russian)
Alekseev MYu, Tkachenko AV, Zubchenko AV, Shkatelov AP, Nikolaev AM (2019) Distribution, spawning and the possibility of fishery of introduced pink salmon (Oncorhynchus gorbusha Walbaum) in rivers of Murmansk region. Russ J Biol Invasions 10:109–117. https://doi.org/10.1134/S2075111719020024
Arizono N, Shedko M, Yamada M, Uchikawa R, Tegoshi T, Takeda K, Hashimoto K (2009) Mitochondrial DNA divergence in populations of the tapeworm Diphyllobothrium nihonkaiense and its phylogenetic relationship with Diphyllobothrium klebanovskii. Parasitol Int 58:22–28. https://doi.org/10.1016/j.parint.2008.09.001
Armstrong JD, Bean CW, Wells A (2018) The Scottish invasion of pink salmon in 2017. J Fish Biol 93:8–11. https://doi.org/10.1111/jfb.13680
Azbelev VV, Yakovenko AA (1963) Materials on acclimatization of pink salmon in the basins of the Barents and White Seas. Trudy Polyarnogo Nauchno-Issledovatel’skogo Instituta Morskogo Rybnogo Khozyaystva i Okeanografii 15:7–26 (In Russian)
Babaluk JA, Reist JD, Johnson JD, Johnson L (2000) First records of sockeye (Oncorhynchus nerka) and pink salmon (O. gorbuscha) from Banks Island and other records of pacific salmon in Northwest Territories, Canada. Arctic 53:161–164. https://doi.org/10.14430/arctic846
Barskaya YuYu, Ieshko EP, Novokhatskaya OV (2005) Formation of parasite fauna of pink salmon Oncorhynchus gorbuscha (Walbaum, 1792) under acclimatization. In: Alimov AF, Berger VYa, Ieshko EP, Nemova NN (eds) Problemy izucheniya, ratsional’nogo ispol’zovaniya i okhrany resursov Belogo morya. Materialy IX mezhdunarodnoy konferentsii. Publishing Company PIN, Petrozavodsk, pp 39–43 (In Russian)
Bogdanov VD, Kizhevatov YaA (2015) Pink salmon (Oncorhynchus gorbuscha, Walbaum, 1792) a new species of aquatic biological resources in the Yamalo-Nenets Autonomous Okrug. Vestnik Astrakhanskogo gosudarstvennogo tekhnicheskogo universiteta. Seriya: Rybnoye Khozyaystvo 3:7–14 (In Russian)
Bouzid W, Stefka J, Hypsa V, Lek S, Scholz T, Legal L, Ben Hassine OK, Loot G (2008) Geography and host specificity: two forces behind the genetic structure of the freshwater fish parasite Ligula intestinalis (Cestoda: Diphyllobothriidae). Int J Parasitol 38:1465–1479. https://doi.org/10.1016/j.ijpara.2008.03.008
Brickle P, Olson PD, Littlewood DTJ, Bishop A, Arkhipkin AI (2001) Parasites of Loligo gahi from waters off the Falkland Islands, with a phylogenetically based identification of their cestode larvae. Can J Zool 79:2289–2296. https://doi.org/10.1139/cjz-79-12-2289
Brickle P, MacKenzie K, Pike A (2006) Variations in the parasite fauna of the Patagonian Toothfish, Dissostichus eleginoides with Length, Season and Depth of habitat around the Falkland Islands. J Parasitol 92:282–291. https://doi.org/10.1645/GE-539R.1
Bush AO, Holmes JC (1986) Intestinal helminths of lesser scaup ducks: patterns of association. Can J Zool 64:132–141
Bush AO, Lafferty KD, Lotz JM, Shostak AW (1997) Parasitology meets ecology on its own terms: Margolis et al. Revisited. J Parasitol 83:575–583. https://doi.org/10.2307/3284227
Caira JN, Jensen K, Pickering M, Ruhnke TR, Gallagher KA (2020) Intrigue surrounding the life-cycles of species of Clistobothrium (Cestoda: Phyllobothriidea) parasitising large pelagic sharks. Int J Parasitol 50:1043–1055. https://doi.org/10.1016/j.ijpara.2020.08.002
Chereshnev IA, Volobuev VV, Shestakov AV, Frolov SV (2002) Salmonids of the North-East of Russia. Dal’nauka, Vladivostok (In Russian)
Darriba D, Taboada GL, Doallo R, Posada D (2012) jModelTest 2: more models, new heuristics and parallel computing. Nat Methods 9:772. https://doi.org/10.1038/nmeth.2109
Delyamure SL, Skrjabin AS, Serdukov AM (1985) Diphyllobothriids – tapeworms of man, mammals and birds. In: Sudarikov VE (ed) Osnovoy Tsestodologii, vol 11. Nauka, Moscow (In Russian)
Dove AD, Cribb TH (2006) Species accumulation curves and their applications in parasite ecology. Trends Parasitol 22:568–574
Drummond AJ, Rambaut A (2007) BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol Biol 7:214. https://doi.org/10.1186/1471-2148-7-214
Dunn AM (2009) Parasites and biological invasions. Adv Parasitol 68:161–184. https://doi.org/10.1016/S0065-308X(08)00607-6
Ebmer D, Navarrete MJ, Muñoz P, Flores LM, Gärtner U, Brabec J, Poppert S, Taubert A, Hermosilla C (2020) Anthropozoonotic parasites circulating in synanthropic and pacific colonies of South American Sea Lions (Otaria flavescens): Non-invasive techniques data and a review of the literature. Front Mar Sci 7:543829. https://doi.org/10.3389/fmars.2020.543829
Ermolenko AV, Besprozvannykh VV, Shed'ko SV (1998) Fauna of parasites of salmonid fish (Salmonidae, Salmoniformes) of Primorsky Krai. Dal'nauka, Vladivostok (In Russian)
Fjær MA (2019) Pukkellaks (Oncorhynchus gorbuscha) tatt på Vestlandet. – Hvilke parasitter og infeksjoner bærer de på? MS thesis. The University of Bergen (In Norwegian)
Fraija-Fernández N, Waeschenbach A, Briscoe AG, Hocking S, Kuchta R, Nyman T, Littlewood DTJ (2021) Evolutionary transitions in broad tapeworms (Cestoda: Diphyllobothriidea) revealed by mitogenome and nuclear ribosomal operon phylogenetics. Mol Phylogenet Evol 163:107262. https://doi.org/10.1016/j.ympev.2021.107262
Gavrilov AL (2016) Parasites of whitefish and pink salmon in the Yuribey River (Baidaratskaya Bay basin). Vestnik Syktyvkarskogo universiteta. Seriya 2. Vypusk 6:29–35
Graham C, Pakhomov EA, Hunt BP (2021) Meta-analysis of salmon trophic ecology reveals spatial and interspecies dynamics across the North Pacific Ocean. Front Mar Sci 8:618884. https://doi.org/10.3389/fmars.2021.618884
Grozdilova TA (1974) Parasites of pink salmon (O. gorbusha) of the White Sea. Parazitologiya 8:293–297 (In Russian)
Guardone L, Giusti A, Bilska-Zajac E, Malandra R, Różycki M, Armani A (2020) Molecular characterization of Clistobothrium sp viable plerocercoids in fresh longfin inshore squid (Doryteuthis pealeii) and implications for cephalopod inspection. Pathogens 9:596. https://doi.org/10.3390/pathogens9070596
Hanski I (1982) Dynamics of regional distribution: the core and satellite species hypothesis. Oikos 38:210–221. https://doi.org/10.2307/3544021
Heard WR (1991) Life history of pink salmon (Oncorhynchus gorbuscha). In: Groot C, Margolis L (eds) Pacific salmon life histories. UBC Press, Vancouver, pp 121–123
Hernández-Orts JS, Kuzmina TA, Gomez-Puerta LA, Kuchta R (2021) Diphyllobothrium sprakeri n. sp. (Cestoda: Diphyllobothriidae): a hidden broad tapeworm from sea lions off North and South America. Parasit Vectors 14:219. https://doi.org/10.1186/s13071-021-04661-1
Hindar K, Hole LR, Kausrud KL, Malmstrøm M, Rimstad E, Robertson L, Sandlund OT, Thorstad EB, Vollset K, de Boer H, Eldegard K, Järnegren J, Kirkendall L, Måren I, Nielsen A, Nilsen EB, Rueness E, Velle G (2020) Assessment of the risk to Norwegian biodiversity and aquaculture from pink salmon (Oncorhynchus gorbuscha). In: Scientific opinion of the panel on alien organisms and trade in endangered species (CITES). Norwegian Scientific Committee for Food and Environment (VKM). https://doi.org/10.1111/faf.12587
Holmes JC, Price PW (1986) Communities of parasites. In: Anderson DJ, Kikkawa J (eds) Community ecology: patterns and processes. Blackwell Scientific Publications, Oxford, pp 187–213
ICES (2018) Report of the Working Group on North Atlantic Salmon (WGNAS). 4–13 April 2018, Woods Hole, Mass., USA. ICES CM 2018/ACOM:21
Ieshko EP, Shulman BS, Barskaya Y, Novokhatskaya OV (2016) Parasite fauna of pink salmon in the Keret River, White Sea. In: Niemelä E, Johansen N, Zubchenko AV, Dempson JB, Veselov A, Ieshko EP (eds) Pink salmon in the Barents region with special attention to the status in the transboundary rivers Tana and Neiden, rivers in North West Russia and in East Canada. Vadsø, Norway, pp 126–127
Ieshko EP, Korosov AV, Sokolov SG (2019) Species richness of parasite assemblages in the chinese slipper Perccottus glenii Dybowski, 1877 (Actinopterygii) in the host’s native and non–native ranges. Parazitologiya 53:145–158. https://doi.org/10.1134/S0031184719020066. (In Russian)
International seminar on pink salmon in The Barents Region and in Northern Europe (2021) County Governor of Troms and Finnmark, NIBIO, Svanhovd, Norway, pp 1–25
Karpenko VI, Volkov F, Koval MV (2007) Diets of pacific salmon in the Sea of Okhotsk, Bering Sea, and Northwest Pacific Ocean. NPAFC Bull 4:105–116
Karpevich AF, Agapov VS, Magomedov GM (1991) Acclimatization and farming of introduced salmonid fishes. Vserossiyskiy nauchno-issledovatel'skiy institut rybnogo khozyaystva i okeanografii, Moscow (In Russian)
Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, Buxton S, Cooper A, Markowitz S, Duran C, Thierer T, Ashton B, Meintjes P, Drummond A (2012) Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28:1647–1649. https://doi.org/10.1093/bioinformatics/bts199
Kelly DW, Paterson RA, Townsend CR, Poulin R, Tompkins DM (2009) Parasite spillback: a neglected concept in invasion ecology? Ecology 90:2047–2056. https://doi.org/10.1890/08-1085.1
Kijewska A, Dzido J, Rokicki J (2009) Mitochondrial DNA of Anisakis simplex s.s. as a potential tool for differentiating populations. J Parasitol 95:1364–1370. https://doi.org/10.1645/GE-1950.1
Klimpel S, Rückert S (2005) Life cycle strategy of Hysterothylacium aduncum to become the most abundant anisakid fish nematode in the North Sea. Parasitol Res 97:141–149. https://doi.org/10.1007/s00436-005-1407-6
Klimpel S, Palm H, Rückert S, Piatkowski U (2004) The life cycle of Anisakis simplex in the Norwegian Deep (northern North Sea). Parasitol Res 94:1–9. https://doi.org/10.1007/s00436-004-1154-0
Klimpel S, Kuhn T, Münster J, Dörge DD, Klapper R, Kochmann J (2019) Parasites of marine fish and cephalopods, 1st edn. Springer, Cham
Klotz D, Hirzmann J, Bauer C, Schöne J, Iseringhausen M, Wohlsein P, Baumgärtner W, Herder V (2018) Subcutaneous merocercoids of Clistobothrium sp. in two Cape fur seals (Arctocephalus pusillus pusillus). Int J Parasitol Parasites Wildl 7:99–105. https://doi.org/10.1016/j.ijppaw.2018.02.003
Køie M (1979) On the morphology and life-history of Derogenes varicus (Muller, 1784) Looss, 1901 (Trematoda, Hemiuridae). Z Parasitenkd 59:67–78. https://doi.org/10.1007/BF00927847
Krupenko D, Kremnev G, Skobkina O, Gonchar A, Uryadova A, Miroliubov A (2022) Lecithaster (Lecithasteridae, Digenea) in the White Sea: an unnoticed guest from the Pacific? J Helminthol 96:E43. https://doi.org/10.1017/S0022149X22000281
Kuchta R, Brabec J, Kubáčková P, Scholz T (2013) Tapeworm Diphyllobothrium dendriticum (Cestoda)—neglected or emerging human parasite? PLoS Negl Trop Dis 7:e2535. https://doi.org/10.1371/journal.pntd.0002535
Kudersky LA (2005) Acclimatization of pink salmon Oncorhynchus gorbuscha (Walbaum, 1972) in Russia. The study, sustainable use and conservation of natural resources of the White Sea. In: Alimov AF, Berger VYa, Iyeshko YeP, Nemova NN (eds) Materialy IX mezhdunarodnoy konferentsii. Izdatel’skiy dom PIN, Petrozavodsk, pp 172–183 (In Russian)
Kulachkova VG (2001) Dynamics of Anisakis sp. (Nematoda) larvae infestation of pink salmon acclimated in the White Sea. In: Tezisy dokladov VIII regional'noy nauchno-prakticheskoy konferentsii "Problemy izucheniya, ratsional'nogo ispol'zovaniya i okhrany prirodnykh resursov Belogo morya". “Pravda Severa”, Arkhangel'sk, pp 227–229 (In Russian)
Kuris AM, Jaramillo AG, McLaughlin JP, Weinstein SB, Garcia-Vedrenne AE, Poinar GO, Pickering M, Steinauer ML, Espinoza M, Ashford JE, Dunn GLP (2015) Monsters of the sea serpent: Parasites of an oarfish, Regalecus russellii. J Parasitol 101:41–44. https://doi.org/10.1645/14-581.1
Love MS, Moser M (1983) A checklist of parasites of California, Oregon, and Washington marine and estuarine fishes. In: NOAA Technical Report NMFS SSRF, U.S. Department of Commerce, vol 777, Rockville, Maryland, pp 1–576
Lozano-Cobo H, Gómez del Prado-Rosas MC, Silva-Segundo CA, Oceguera-Figueroa A, Gómez-Gutiérrez J (2021) Molecular identification of plerocercoids of Clistobothrium montaukensis (Cestoda: Phyllobothriidea) parasitizing the King of Herrings Regalecus glesne. Acta Parasit 66:1586–1592. https://doi.org/10.1007/s11686-021-00400-9
Lymbery AJ, Morine M, Kanani HG, Beatty SJ, Morgan DL (2014) Co-invaders: the effects of alien parasites on native hosts. Int J Parasitol Parasites Wildl 24:171–177. https://doi.org/10.1016/j.ijppaw.2014.04.002
Malakhova RP (1972) Parasites of salmon Salmo salar L., trout Salmo trutta L., pink salmon Oncorhynchus gorbuscha (Wabb.) and whitefish Coregonus lavaretus n. pidschianoides Pravdin in the White Sea basin. In: Potapova OI, Smirnov YA (eds) Salmonidae of Karelia. Ecology, parasites, biochemistry, Karelian Branch of the USSR Academy of Sciences Institute of Biology, Petrozavodsk, pp 21–26 (In Russian)
Mamaev YL, Parukhin AM, Baeva OM, Oshmarin PG (1959) Helmintofauna of Far Eastern salmonids in relation to the issue of local stocks and migration ways of these fish. Primorskoye knizhnoye izdatel'stvo, Vladivostok (In Russian)
Mattiucci S, Nascetti G (2008) Chapter 2 Advances and trends in the molecular systematics of anisakid nematodes, with implications for their evolutionary ecology and host—parasite co-evolutionary processes. In: Advances in parasitology, pp 47–148. https://doi.org/10.1016/s0065-308x(08)00202-9
Mattiucci S, Cipriani P, Webb SC, Paoletti M, Marcer F, Bellisario B, Gibson DI, Nascetti G (2014) Genetic and morphological approaches distinguish the three sibling species of the Anisakis simplex species complex, with a species designation as Anisakis berlandi n. sp. for A. simplex sp. C (Nematoda: Anisakidae). J Parasitol 100:199–214. https://doi.org/10.1645/12-120.1
Mattiucci S, Cipriani P, Paoletti M, Levsen A, Nascetti G (2017) Reviewing biodiversity and epidemiological aspects of anisakid nematodes from the North-east Atlantic Ocean. J Helminthol 91:422–439. https://doi.org/10.1017/s0022149x1700027x
Mitenev VK, Karasev AB (1995) Parasites of salmonids of the Murmansk region. Polyarnyy Nauchno-Issledovatel’skiy Institut Morskogo Rybnogo Khozyaystva i Okeanografii Press, Murmansk (In Russian)
Mitenev VK, Shulman BS (1999) Fish parasites of water bodies of Murmansk region: Systematic katalog. Polyarnyy Nauchno-Issledovatel'skiy Institut Morskogo Rybnogo Khozyaystva i Okeanografii, Murmansk (In Russian)
Muzzall PM, Bowen CA (2002) Parasites of the Slimy Sculpin, Cottus cognatus Richardson, 1836, from Lake Huron, U.S.A. Comp Parasitol 69:196–201. https://doi.org/10.1654/1525-2647(2002)069[0196:POTSSC]2.0.CO;2
Nagasawa K, Urawa S, Awakura T (1987) A checklist and bibliography of parasites of Salmonids of Japan. Sci Rep Hokkaido Salmon Hatchery 41:1–75
Najda K, Kijewska A, Kijewski T, Plauška K, Rokicki J (2018) Distribution of ascaridoid nematodes (Nematoda: Chromadorea: Ascaridoidea) in fish from the Barents Sea. Oceanol Hydrobiol Stud 47:128–139. https://doi.org/10.1515/ohs-2018-0014
Nielsen J, Rosing-Asvid A, Meire L, Nygaard R (2020) Widespread occurrence of pink salmon (Oncorhynchus gorbuscha) throughout Greenland coastal waters. J Fish Biol 96:1505–1507. https://doi.org/10.1111/jfb.14318
Ninburg EM (1963) Parasites of Murmansk pink salmon and its features. Trudy Polyarnogo Nauchno-Issledovatel’skogo Instituta Morskogo Rybnogo Khozyaystva i Okeanografii 15:57–66 (In Russian)
Oliva M, Luque JL, Iannacone JA (1990) The metazoan parasites of Stellifer minor (Tschudi, 1844): an ecological approach. Rev Bras Biol 60:577–584. https://doi.org/10.1590/s0074-02761990000300003
Olson PD, Littlewood DTJ, Bray RA, Mariaux J (2001) Interrelationships and evolution of the tapeworms (Platyhelminthes: Cestoda). Mol Phylogenet Evol 19:443–467. https://doi.org/10.1006/mpev.2001.0930
Paggi L, Nascetti G, Cianchi R, Orecchia P, Mattiucci S, D’Amelio S, Berland B, Brattey J, Smith J, Bullini L (1991) Genetic evidence for three species within Pseudoterranova decipiens (Nematoda, Ascaridida, Ascaridoidea) in the North Atlantic and Norwegian and Barents seas. Int J Parasitol 21:195–212. https://doi.org/10.1016/0020-7519(91)90010-5
Paterson RA, Rauque CA, Fernandez MV, Townsend CR, Poulin R, Tompkins DM (2013) Native fish avoid parasite spillback from multiple exotic hosts: consequences of host density and parasite competency. Biol Invasions 15:2205–2218. https://doi.org/10.1007/s10530-013-0445-8
Paulsen T, Sandlund OT, Østborg G, Thorstad EB, Fiske P, Muladal R, Tronstad S (2022) Growth of invasive pink salmon (Oncorhynchus gorbuscha) at sea assessed by scale analysis. J Fish Biol 100:218–228. https://doi.org/10.1111/jfb.14937
Potter ECE, Crozier WW (2000) A perspective on the marine survival of Atlantic salmon. In: Mills D (ed) The ocean life of Atlantic salmon—environmental and biological factors influencing survival. Fishing News Books, Oxford, pp 19–36
R Core Team. R: a language and environment for statistical computing. R version 4.1.2. (2021–11–01). Vienna: R Foundation for statistical computing. Available online: http://www.r-project.org/. Accessed 1 Nov 2022
Rambaut A (2018) FigTree v. 2.0. http://tree.bio.ed.ac.uk/software/
Randhawa HS, Brickle P (2011) Larval parasite gene sequence data reveal cryptic trophic links in life cycles of porbeagle shark tapeworms. Mar Ecol Prog Ser 431:215–222. https://doi.org/10.3354/meps09108
Rullestad I (2021) Parasites found in pink salmon (Oncorhynchus gorbuscha) caught in the feeding areas in the Norwegian Sea. MS thesis. The University of Bergen.
Sandlund OT, Berntsen HH, Fiske P, Kuusela J, Muladal R, Niemelä E, Uglem I, Forseth T, Mo TA, Thorstad EB, Veselov AE, Vollset KW, Zubchenko AV (2019) Pink salmon in Norway: the reluctant invader. Biol Invasions 21:1033–1054. https://doi.org/10.1007/s10530-018-1904-z
Sarabeev V, Balbuena JA, Morand S (2017) Testing the enemy release hypothesis: abundance and distribution patterns of helminth communities in grey mullets (Teleostei: Mugilidae) reveal the success of invasive species. Int J Parasitol 47:687–696. https://doi.org/10.1016/j.ijpara.2017.05.006
Shitikov VK, Rosenberg GS (2013) Randomization and bootstrap: statistical analysis in biology and ecology using R. Cassandra, Tol'yatti (In Russian)
Sokolov SG, Frenkel SE, Gordeev II (2016) Metacercerae of Brachyphallus crenatus Rudolphi, 1802 (Trematoda: Heniuridae) in plankton crustaceans from Prostor Gulf (Iturup island, Russia). Parazitologiya 50:150–155
Strelkov YuA (1960) Endoparasitic worms of marine fish of eastern Kamchatka. Tr Zool Inst 28:147–196 (In Russian)
Tompkins DM, Poulin R (2006) Parasites and biological invasions. In: Allen RB, Lee WG (eds) Biological invasions in New Zealand. Springer, Berlin, pp 67–86. https://doi.org/10.1007/3-540-30023-6_5
Torchin ME, Lafferty KD (2009) Escape from parasites. In: Rilov G, Crooks JA (eds) Biological invasions in marine ecosystems, vol 204. Springer, Heidelberg, pp 203–214. https://doi.org/10.1007/978-3-540-79236-9_11
Utne KR (2021) Bycatches of pink salmon in the Norwegian Sea and along the coast of Norway. In: International seminar on pink salmon in The Barents Region and in Northern Europe, NIBIO, Svanhovd, Norway, pp 8–9
Utne KR, Thomas K, Jacobsen JA, Fall J, Maoileidigh NO, Broms CT et al (2021) Feeding interactions between Atlantic salmon (Salmo salar) postsmolts and other planktivorous fish in the Northeast Atlantic. Can J Fish Aquat Sci 78:255–268. https://doi.org/10.1139/cjfas-2020-0037
Villesen P (2007) FaBox: an online toolbox for FASTA sequences. Mol Ecol Notes 7:965–968. https://doi.org/10.1111/j.1471-8286.2007.01821.x
Waeschenbach A, Littlewood DTJ (2017) A molecular framework for the Cestoda. In: Caira JN, Jensen K (eds) Tapeworms from the vertebrate bowels of the Earth, Planetary Biodiversity Inventory (2008–2017). Natural History Museum, Lawrence, pp 431–451
Waeschenbach A, Webster BL, Littlewood DT (2012) Adding resolution to ordinal level relationships of tapeworms (Platyhelminthes: Cestoda) with large fragments of mtDNA. Mol Phylogenet Evol 63:834–847. https://doi.org/10.1016/j.ympev.2012.02.020
Waeschenbach A, Brabec J, Scholz T, Littlewood DTJ, Kuchta R (2017) The catholic taste of broad tapeworms—multiple routes to human infection. Int J Parasitol 47:831–843. https://doi.org/10.1016/j.ijpara.2017.06.004
Yamasaki H, Muto M, Yamada M, Arizono N, Rausch RL (2012a) Validity of the bear tapeworm Diphyllobothrium ursi (Cestoda: Diphyllobothriidae) based on morphological and molecular markers. J Parasitol 98:1243–1247. https://doi.org/10.1645/GE-3063.1
Yamasaki H, Ohmae H, Kuramochi T (2012b) Complete mitochondrial genomes of Diplogonoporus balaenopterae and Diplogonoporus grandis (Cestoda: Diphyllobothriidae) and clarification of their taxonomic relationships. Parasitol Int 61:260–266. https://doi.org/10.1016/j.parint.2011.10.007
Zdzitowiecki K (1988) Occurrence of digenetic trematodes in fishes of South Shetlands (Antarctic). Acta Parasitol Pol 33:155–167
Zueva KD (1963) Smoltification of pink salmon Oncorhynchus gorbuscha (Walb.) without “parr” stage. Voprosy Ikhtiologii 5:324–330 (In Russian)
Acknowledgements
The authors thank Olga Kislova and Natalia Lentsman for language editing.
Funding
The current study was funded by the Russian Ministry of Science and Education: FFER-2021-0005 376 (SS) and FMEN-2022-0005 (EI and AP).
Author information
Authors and Affiliations
Contributions
SS identificated parasites. SS, EI, and AP collected samples. VG conducted statistical analyses. NG conducted phylogenetic analyses. SS, EI, and VG wrote the original draft. SS, EI, VG, NG, and AP edited the draft. All authors have read and approved the published version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
The authors assert that all procedures contributing to this work comply with the ethical standards of the relevant national and institutional guides on the collecting, care, and dissection of animals.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Sokolov, S., Ieshko, E., Gordeeva, N. et al. Parasites of invasive pink salmon, Oncorhynchus gorbuscha (Walbaum, 1792) (Actinopterygii: Salmonidae), in the Kandalaksha Bay of the White Sea. Polar Biol 47, 101–113 (2024). https://doi.org/10.1007/s00300-023-03214-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-023-03214-9