
Abstract
Ichthyological studies are an important part of the research effort on the water ecosystems of the Arctic. However, data on the changes in the ichthyofauna of the Arctic seas are scarce and contradictory. We studied species composition of trawl catches made in 1980–2015 in the coastal and the open part of the Pechora Sea (south-eastern Barents Sea). We determined the relative abundance (individuals per hour of trawling) of the dominant fish species in the coastal and the open zone and described the taxonomic status of fish and their ecological grouping by distribution area and predominant type of feeding. Cluster analysis revealed four groups of study years similar in respect of the proportion of dominant fish species in trawl catches. The species diversity of the ichthyofauna increased over the study period and so did the number of predominantly Arctic species in the catches. On the contrary, the proportion of Arctic species such as polar cod decreased. In our opinion, this was caused by the changes in the mean water temperature, which showed a tendency to increase over the study period. A comparative analysis of the ichthyofauna of the Pechora Sea and the Baydaratskaya Bay of the Kara Sea revealed differences in the dominance of Arctic and boreal fishes, which might have been associated with the differences in the mean annual temperature in these water bodies. Our results can be used as background data for further monitoring of climate change.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Water ecosystems of the Arctic are unique and extremely vulnerable systems with a low restorative capacity. They require close scientific attention, especially taking into account the potential impact of climatic changes. Fish inventories are often needed both for the analysis of processes occurring in these ecosystems and for biodiversity conservation programs. Therefore, the evaluation of the current status of the ichthyofauna and its changes influenced by climatic factors are important research directions.
The ichthyofauna of the Barents Sea has been extensively studied (Gratsianov 1907; Knipovich 1897, 1926; Esipov 1937; Andriashev 1954; Andriashev and Chernova 1994). These studies, having a special historical significance, are the main sources of information on species composition and taxonomy of fish in the Barents Sea. Current estimates of the number of fish species in the Barents Sea differ: Dolgov (2004) reports 207 species, while Karamushko (2007, 2008) who considers the Barents Sea in restricted geographical boundaries, reports 182 species and subspecies of fish, mainly from the boreal complex.
Western areas of the Barents Sea are the most productive. They are also the best studied because research and fishing vessels can operate there all the year round. Studies of the south-eastern part of the Barents Sea have been sporadic due to its low productivity, remoteness, and complex ice conditions. Some publications describe the ichthyofauna of distinct areas such as the Choshskaya Bay, the Pechora Bay or smaller bays (Kornilova 1970; Karamushko et al. 1996; Chernova 2000; Karamushko and Aleksandrov 2003; Pogrebov et al. 2003; Karamushko 2005). Chernova (2000) clarified the data on the ichthyofauna of the Pechora Sea published by Karamushko et al. (1996) and described 71 taxa of fish and fish-like species and their habitat, including marine (46 species), anadromous, and semi-anadromous fish, as well as freshwater species, which regularly or occasionally occur in the brackish waters of the estuarine zones.
The Pechora Sea is a conventional name of a water area in the south-eastern part of the Barents Sea between the islands of Kolguev and Vaygach. It is influenced by the Pechora River, which provides about 80% of the total river inflow. The largest bays along the shoreline of the mainland are the Pechora Bay and the Khaypudyrskaya Bay. Two straits in the south-eastern part of the Pechora Sea, the Kara Strait and the Yugorsky Strait, connect the Barents Sea and the Kara Sea. The area is heavily influenced by the Nordkapp branch of the warm current from the north and north-west, cold currents from the north along the coast of Novaya Zemlya such as Litke current, and the freshwater flow of the Pechora river from the south (Girdyk et al. 1990; Romankevich et al. 2003). The Pechora Sea has unique abiotic conditions, a high level of biological productivity, and a great diversity of biotopes, which are important for maintaining the abundance of fish.
Climate change in the Arctic, resulting in the increase of average annual water and air temperatures and the shrinking of the ice cover (Almarov 1990; Karsakov et al. 2000; Matishov et al. 2011), affects the functioning of water ecosystems. In particular, it affects the distribution of commercial marine fish species (Berg 1939, 1962; Gasconskiy 1963; Borkin and Grigoriev 1986; Smirnov et al. 2000; Chernova 2000; Semushin and Novoselov 2009). In the light of this, ichthyological studies of these areas are potentially important for fishery. However, the available data on the changes in the ichthyofauna of the Arctic seas are scarce and contradictory.
The aim of this study was to describe the ichthyofauna of the Pechora Sea in respect of species diversity, taxonomic and faunal composition, ecological characteristics and the dynamics of changes over the study period and to compare it with the ichthyofauna of the neighboring Baydaratskaya Bay of the Kara Sea.
Materials and methods
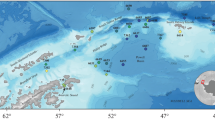
Our study was based on the results of the annual trawl surveys of the Northern Branch of Polar Research Institute of Marine Fisheries and Oceanography (PINRO). We analyzed fish species composition of trawl catches in the Pechora Sea made in 1980–2015. The data were collected in the course of regular research voyages (July–September) during the ice–free period. The grid and the number of trawling stations differed from year to year, depending on the voyage aims and ice and weather conditions, but they were always confined to the same areas to ensure comparability of the data (Fig. 1).

Standard grid of PINRO sampling stations in the Pechora Sea
Ichthyological material was collected by a bottom trawl with a horizontal opening of 14 m, a vertical opening of 5 m and a mesh size of 16 mm. Trawling was performed at an average speed of 3.2 knots with a time period of 15–60 min. Studies were not conducted in 1997–1999 and 2009–2012. Data were analyzed as a single body and in two groups: coastal trawls (down to the isobath of 15–20 m, including the coastal strip and the entire water area of the Pechora and the Khaypudyrskaya Bay) and trawls in the open part of the Pechora Sea. In total, the composition of 1226 trawls during the period of 29 years was analyzed.
Data on the average monthly temperature of the surface water layer were obtained from the reports of "Cape Konstantinovsky" meteostation and the readings of satellite imagery (Monthly Sea Surface Temperature from NOAA NCEP EMC CMB GLOBAL; Karsakov et al. 2000). The maximum sea depth in our study area was 60m, while in 80% of area the depths were lower than 30m. Water mixing in the Pechora Sea is fairly intense, especially in autumn. As a result, there is almost no thermocline and no vertical stratification there, particularly in the coastal shallow zone (Girdyk et al. 1990). Therefore, in our opinion, the surface water temperature is an adequate index of climate change in the study area and can be applied to the analysis of distribution of both pelagic and bottom fish.
Previously published data on the ichthyofauna of the Baydaratskaya Bay of the Kara Sea (Semushin and Novoselov 2009) were used for the comparative analysis of the Pechora Sea and the Baydaratskaya Bay in this paper. Overall composition of ichthyofauna of the Baydaratskaya Bay was derived from literature sources and PINRO data from trawl surveys. Anadromous and semi-anadromous species in the catches were considered as an element of the marine community. The trophic status of the studied species was categorized after Karamushko (2008). Their geographical range and ecological status were defined after Andriashev and Chernova (1994). Distribution areas were defined based on zoogeographical approach (Dolgov 2011; Mecklenburg et al. 2018). Classification and species names were used according to the Catalog of Fishes (https://www.calacademy.org/scientists/projects/catalog-of-fishes).
Linear correlations between the abundance of fish species in catches by year and the average water temperature for individual months (May to October), as well as the entire vegetation season (May–October) were calculated. The ecological parameters and the composition of the ichthyocenosis in different years were assessed with the use of the Shannon and the Berger-Parker indices of diversity (Bigon et al. 1989; Shilov 1997).
The Shannon index (H) was calculated using the formula
where \({n}_{i}\) the abundance of a given species; \(N\) the abundance of all species in the biocenosis;
The Berger–Parker index was calculated using the formula
where \({N}_{{\text{max}}}\) the number of individuals of the most abundant species, \(N\) the number of individuals of all species.
We compared data on the composition of ichthyofauna for each study year to identify trends and similarities between individual research years. Statistical processing involved values of the relative numbers of fish in trawls (individuals per hour of trawling) for 10 most common species: Gymnocanthus tricuspis (Reinhardt, 1830), Arctic staghorn sculpin, Liopsetta glacialis (Pallas, 1776), Arctic flounder, Osmerus dentex Steindachner & Kner, 1870, Pacific rainbow smelt, Lumpenus fabricii Reinhardt, 1836, slender eelblenny, Eleginus nawaga (Walbaum, 1792), navaga, Melanogrammus aeglefinus Linnaeus, 1758, haddock, Coregonus sardinella Valenciennes, 1848, sardine cisco, Boreogadus saida (Lepechin, 1774), polar cod, Clupea pallasii Valenciennes, 1847, Pacific herring and Gadus morhua Linnaeus, 1758, Atlantic cod. Normal distribution of the abundance of species in the catches was analyzed using the Kolmogorov–Smirnov test. The Kruskal–Wallis H criterion showed no significant differences (p > 0.05) between the data from 2004 and 2005 and between the data from 2013, 2014 and 2015, which made it possible to form two unified groups of study periods, respectively (“2005” and “2015”). Thus, the results for 2004 and 2005, as well as for 2013, 2014 and 2015 were processed separately from the rest of the data.
At the first stage, the samples were compared pairwise (325 pairs of comparisons) with the use of the non-parametric Mann–Whitney U test for independent samples. Significant differences were found for each pair of comparisons within each of the 10 analyzed species. For each pair of studied years, a total number of significant differences was calculated. The results were arranged in the matrix of paired comparisons.
At the second stage, all study years were classified into groups using Ward's method for hierarchical cluster analysis (Ward 1963), based on the resulting matrix of paired comparisons. The optimal number of clusters was determined on the basis of a stepwise growth of the coefficient characterizing the change in distance between clusters (the square of the Euclidean distance). The differences were considered reliable at the significance level of p < 0.05. Mathematical and statistical processing was performed with the use of Microsoft Excel 2013 and SPSS 22.0 for Windows.
Results
The total list of fish and fish-like species from the trawl catches in the Pechora Sea during the study period contained 50 species belonging to 19 families of marine, anadromous, semi-anadromous and non-migratory fish (Table 1). Forty-seven species were found in trawl catches. Three representatives of the Salmonidae family were not recorded in the catches, but the archival results of net catches indicated that they were present in the ichthyocenosis.
The analysis of the generalized sample on the ecological status showed that marine near bottom group and marine cryo-pelagic group contained only one species each (Greenland halibut and polar cod, respectively; 2.0% of all caught fish in both cases) (Fig. 2a). Three species (6.0%) belonged to the group of freshwater fishes. The group of marine nerito-pelagic fish contained 4 species (8.0%), the groups of semi-anadromous and marine bottom-pelagic fish were represented by five species each (10.0%). Anadromous fishes were represented by 6 species (12%) All other fish noted in the trawl catches (25 species, 50.0%) were marine bottom species.

Proportion of species (%) by ecological status (a), geographical distribution (b), and trophic status (c)
The fish inhabiting the Pechora Sea can be attributed to ten groups by their geographical distribution (Andriashev and Chernova 1994). Fifteen species (30.0%) were Arctic species (Fig. 2b). There were 12 mostly boreal-Atlantic species (24.0%) and 8 mostly boreal-European species (16.0%). Five species (10.0%) belonged to mostly boreal group. The latter three groups are typical of boreal waters but are also common in the marginal regions of the Arctic (for example, in the eastern part of the Barents Sea). Mostly Arctic fishes were represented much more poorly (2 species, 4.0%). The Arctic-boreal faunal complex contained three species (6.0%). The boreal group comprised 2 species (4.0%). The boreal-European, boreal-Atlantic and boreal-Pacific groups were represented by one species each (2.0%) (Fig. 2b).
Four groups were identified according to the predominant feeding type (Karamushko 2008). Only one species, anadromous Arctic lamprey (2.0%), was characterized by a predatory-parasitic type of feeding (Fig. 2c). Eleven species (22.0%) were typical predators. Ten species (20.0%) fed on plankton. The remaining fish (28 species, 56.0%) were marine bottom species, which fed on benthos (Table 1, Fig. 2c).
The species composition of trawl catches in the coastal zone and the open part of the Pechora Sea is shown in Table 2. There were no representatives of the Anarhichadidae family in the coastal zone but more species of the Salmonidae family were found there (6 species, 14.0%). Catches in the open part of the sea did not contain representatives of the Lotidae (burbot) and the Percidae (ruffe) families, which were sporadically observed in the coastal, more freshened zone. Two species of the Anarhichadidae family (spotted and Atlantic wolffishes) were found in the open zone of the sea. In the trawl catches at depths less than 15–20 m 43 species of fish belonging to 18 families were identified. In the open area (trawl surveys at depths more than 20 m), 37 species of 17 families were found.
E. nawaga dominated in the catches of commercial fish both in the coastal zone (56.5%) and the open zone (46.9%) (Fig. 3). B. saida and C. pallasii were common subdominant species in the open zone (19.2 and 16.6%, respectively), occurring in the coastal zone in smaller numbers (4.3% and 11.9%, respectively). M. aeglefinus did not occur in the coastal zone at all but was quite common in the open zone (8.4%). O. dentex (16.0%) and L. glacialis (7.2%) were quite common in the coastal zone, while in the open zone their relative abundance was rather low (4.4% and 1.4%, respectively). C. sardinella was found in small numbers (1.4%) in trawl catches in the coastal zone and in even smaller numbers (a few individuals) in the open zone. G. morhua individuals were found sporadically both in the coastal zone (0.2%) and in the open zone (0.7%). Gymnocanthus tricuspis and L. fabricii occurred in very small numbers in the coastal zone (0.5 and 0.7%, respectively) and in somewhat greater numbers in the open zone (2.5 and 2.6%, respectively).

Proportion of fish in trawl catches (percentage of individuals per hour of trawling) in the coastal and the open part of the Pechora Sea
The study years were combined into four clusters as a result of the statistical analysis. Each of these four clusters comprised years characterized by the smallest number of reliable differences in 10 most common fish species. Figure 4 displays the results of cluster analysis including two combined groups (“2005”, “2015”).

Dendrogram of studied time periods by the degree of similarity in the composition of trawl catches. Asterisk, combined year groups
The mean water temperature increased during the study period from 5.1 to 6.3°C in the coastal waters (data of "Cape Konstantinovsky" meteostation) and from 3.4 to 4.8°C in the open waters (the satellite data) (Table 3). Five dominant fish species whose numbers showed the greatest variation during different time periods are shown in Table 3.
The indices of fish species diversity increased, except in the last period (IV) covering 2007, 2008 and “2015” (Table 3). The trend towards an increase of average monthly water temperatures in the Pechora Sea during the vegetation season was observed (Fig. 5).

Changes of the water temperature in the Pechora Sea during the study period
Discussion
Our analysis revealed four time periods characterized by a similar species diversity and the proportion of dominant fish species in trawl catches. Our results showed that the relative abundance of mostly Arctic species (MA) in trawl catches increased over the past 36 years. For instance, the relative abundance of Liopsetta glacialis increased from 1.0 to 11.0%, and that of Lumpenus fabricii, from 0.9 to 2.8% (Fig. 6). At the same time, the relative abundance of an Arctic species (A) B. saida decreased from 20.7 to 3.6%, while that of another Arctic species, E. nawaga, remained approximately at the same level, varying from 44.1% in period III to 64.9% in period IV.

Fluctuation in a relative (catch per effort) abundance of dominant species in the Pechora Sea by clusters
The proportion of a mostly boreal species (MB) O. dentex increased from 6.1% during period I to 14.2 and 13.6% during period II and period III, respectively, and decreased to the initial level (5.9%) during period IV (Fig. 6).
The correlation between the average annual abundance of dominant fish species and the surface water temperature in both coastal and open parts of the Pechora Sea did not reach significant levels in a linear correlation analysis. The correlation coefficients for all the compared variables were below 0.6. This might be explained by the fact that the abundance of fish and the biodiversity of the ichthyocenosis is determined not only by abiotic factors (temperature, gas regime, salinity, hydrodynamic processes) but also by a number of biotic ones (spawning and feeding migrations, intra- and interspecies trophic relations).
A deeper understanding of the relationship between the composition of fish assemblages and the temperature regime of marine areas can be gained by comparing the ichthyofauna of the Pechora Sea with that of the Baydaratskaya Bay. This bay is located in the southwest of the Kara Sea and is characterized by a lower annual water temperature. Ichthyofauna of the Pechora Sea is richer (50 species) than that of the Baydaratskaya Bay (32 species). Out of the total number of species recorded in these two areas (53 species), about a half (29 species) were found in trawl and net catches in both areas. Twenty-one species found in catches in the Pechora Sea were absent in the Baydaratskaya Bay, while only three species present in the Baydaratskaya Bay were absent in the Pechora Sea (Table 1).
The analysis of composition of the fish groups by geographical distribution revealed the predominance of Arctic species (A) and mostly Arctic species (MA) in the Baydaratskaya Bay compared to the Pechora Sea (A: 46.8% and 30.0%, respectively; MA: 6.3 and 4.0%, respectively) (Fig. 7). An opposite situation was observed for the species of the boreal complex. Proper boreal species (B) accounted for 4.0% of the total identified ichthyofauna in the Pechora Sea but were absent in the Baydaratskaya Bay.

The proportion of species from different faunal complexes by geographic distribution
Mostly boreal (MB), mostly boreal-Atlantic (MBA), mostly boreal-European (MBE), and Arctic-boreal (AB) species predominated in the Pechora Sea compared to the Baydaratskaya Bay, accounting, respectively, for 24.0%, 16.0%, 10.0%, and 6.0% in the Pechora Sea and 9.4%, 18.8%, 9.4%, and 3.1% in the Baydaratskaya Bay. Boreal-Arctic species (BA) were found only in the Pechora Sea (2.0%). One boreal-European (BE) species (Agonus cataphractus) and one boreal-Pacific (BP) species (acclimatized Oncorhynchus gorbuscha) were identified in both geographical areas, accounting, respectively, for 2.0% in the Pechora Sea and 3.1% in the Baydaratskaya Bay (Fig. 7).
The predominance of boreal species in the Pechora Sea and the Arctic species in the Baydaratskaya Bay can be explained by the significant difference in mean annual temperatures of these neighboring water bodies. In the Pechora Sea, according to the satellite survey, the mean surface water temperature for the period from May to October was in the range 1.9–5.7 (mean 4°C), while in the Baydaratskaya Bay it fluctuated in different years from 0.2 to 4°C (mean 2.3°C) (Fig. 8).

Mean annual temperatures in the Pechora Sea of the Barents Sea and the Baydaratskaya Bay of the Kara Sea (satellite data)
Conclusion
The analysis of the composition of trawl catches in the Pechora Sea over a 36-year period revealed 50 fish species belonging to 19 families. The highest species diversity in the coastal zone was observed in the families Salmonidae and the Cottidae. The most diverse families in the open part of the sea were the Gadidae, the Cottidae, and the Pleuronectidae. The analysis of the generalized sample showed that more than half of the identified fish species were marine bottom species. Arctic species represented 30% of the ichthyofauna, while mostly boreal-European and mostly boreal-Atlantic species occurred in lower numbers. About 56% of the ichthyofauna were benthophagous species, and about 25% were plankton-feeders and predators.
Four time periods characterized by the similarity of the variables—species diversity and proportion of dominant fish species in trawl catches—were revealed by clustering analysis of the generalized data. During the study period the mean water temperatures had a pronounced tendency to increase both in the coastal waters of "Cape Konstantinovsky" meteostation and the surface waters according to the satellite data. The coefficients of species diversity of the ichthyofauna increased. A long-term increase of the water temperature apparently resulted in a significant decrease in the proportion of some Arctic species such as polar cod in catches but had almost no effect on the abundance of other Arctic species such as navaga. At the same time, the relative numbers of the mostly Arctic species (slender eelblenny) and the Arctic species (Arctic flounder) increased, the reasons being unclear.
The Pechora Sea, being a relatively isolated south-eastern part of the Barents Sea, combines two large zoogeographic complexes of the ichthyofauna: the Arctic and the boreal. Significant differences revealed in the ichthyofauna composition of the Pechora Sea and Baydaratskaya Bay of the Kara Sea were mainly determined by the temperature factor. Changes in the ichthyofauna structure of the Arctic seas should be expected taking into account the long-term gradual warming trend of the water area under study. Based on the obtained results, we can assume the possibility of structural changes in the ichthyocenoses, with a tendency towards decreasing abundance of Arctic fish and increasing abundance of boreal fish.
References
Almarov GL (1990) Global warming of the Earth climate and the prospects for fishing. Fisheries 12:27–29 (in Russian)
Andriashev AP (1954) Fishes of northern seas of the USSR. ANSSSR, Leningrad (in Russian)
Andriashev AP, Chernova NV (1994) Annotated list of fish like species and fishes of the Arctic seas and adjacent waters. Voprosy Ikhtiologii 4:435–456 (in Russian)
Berg LS (1939) Appearance of boreal fishes in the Barents Sea. In: A collection devoted to the scientific work of NM Knipovich. Moscow, Leningrad, pp 207–208 (in Russian)
Berg LS (1962) Recent climatic fluctuations and their effect on fish migration. Selected works 5:149–159 (in Russian)
Bigon M, Harper D, Townsend K (1989) Ecology: individuals, populations, and communities. Mir, Moscow (in Russian)
Borkin IV, Grigoriev GV (1986) On capture of maurolicus off Novaya Zemlya. Voprosy Ikhtiologii 5:857–859 (in Russian)
Chernova NV (2000) The fishes of the Pechora Sea and potential impact of petroleum industry. Materials of symposium on biological resources of coastal areas of the Russian Arctic. VNIRO, Moscow, pp 179–191 (in Russian)
Dolgov AV (2004) Species composition of the ichthyofauna and structure of ichthyocenes of the Barents Sea. Izvestiya TINRO 137:177–195 (in Russian)
Dolgov AV (2011) Atlas identifier of fish of the Barents Sea. PINRO, Murmansk (in Russian)
Eschmeyer WN, Fong JD (2017) Species by family/subfamily in the Catalog of fishes https://www.calacademy.org/scientists/projects/catalog-of-fishes.
Esipov VK (1937) Commercial fishes of the Barents Sea. Leningrad, Moscow (in Russian)
Gaskonskiy AA (1963) Warm water fishes in northern seas. Priroda 12:118 (in Russian)
Girdyk GV, et al. (eds) (1990) Hydrometeorology and Hydrochemistry of the USSR Seas, the Barents Sea. Gidrometeoizdat, Leningrad (in Russian)
Gratsianov VI (1907) An attempt at review of fishes of the Russian empire in systematic and geographic aspect. In: Works of the Ichthyology Department of the Imperial Russian Society for the Acclimatization of Animals and Plants, vol 4, pp 1–558 (in Russian)
Karamushko LI, Chernetskii AG, Karamushko OV (1996) Ecosystems, bioresources and anthropogenous pollution of the Pechora Sea. KNTs RAN, Apatity, pp 72–79 (in Russian)
Karamushko OV (2005) Life conditions and some formation patterns of fish communities in the Barents Sea. In: Fundamental bases of management of biological resources. KMK, Moscow pp 279–284 (in Russian)
Karamushko OV (2007) Life conditions and some formation of patterns of fish communities in the Barents Sea. In: Recent investigations of the ichthyofauna of Arctic and southern Seas. KNTs RAN, Apatity, pp 33–55 (in Russian)
Karamushko OV (2008) Species composition and structure of the ichthyofauna of the Barents Sea. J Ichthyol 4:277–291
Karamushko OV, Aleksandrov DI (2003) Diversity of fish communities of the southeastern part of the Barents Sea in summer 2001. J Ichthyol 7:511–515
Karsakov AL, Sentyabov EV, Bochkov YA (2000) North Atlantic surface temperature and long-term prediction of abiotic and biotic parameters of the ecosystem of the northern basin. In: Materials of the final session of PINRO on results of research in 1998–1999. PINRO, Murmansk, pp 188–199 (in Russian)
Knipovich NM (1897) List of fishes of the White and Murmansk Seas. Yearbook of the Zoological Museum. St Peterburg 2:144–158 (in Russian)
Knipovich NM (1926) Key to fishes of the Barents, White and Kara Seas. Works of Research Institute on North studies. St Peterburg 27:1–183 (in Russian)
Kobelev EA, Novoselov AP (2000) Composition of ichthyofauna and structure of populations of commercial fish in Baidaratskaya Bay of the Kara Sea. In: Proceedings of symposium on biological resources of the coastal zone of the Russian Arctic. VNIRO, Moscow, pp 57–63 (in Russian)
Kornilova VP (1970) Tthe ichthyofauna of the Pechora lower reaches and the Pechora Bay of the Barents Sea. Materials of fishery research of the northern basin. PINRO, Arkhangelsk, pp 5–44 (in Russian)
Matishov GG, Moiseyev DV, Lubina OS, Zhichkin AP, Djenjuk SL, Makarevitch PR, Frolova EA (2011) Hydrobiological indicators of cyclical climate changes in Western Arctic. Vestnik Yuzhnogo Nauchnogo Tsentra RAN 2:54–68 (in Russian)
Mecklenburg et al (2018) Marine fishes of the Arctic region. Akureyri, Iceland
Monthly Sea Surface Temperature from NOAA NCEP EMC CMB GLOBAL Reyn Smith OI v2. (2017) [Electronic resource]: National Oceanic and Atmospheric Administration, National Center for Environmental Prediction, Environmental Modeling Center, Climate Modeling Branch. https://iridl.ldeo.columbia.edu/SOURCES/.NOAA/.NCEP/.EMC/.CMB/.GLOBAL/.Reyn_SmithOIv2/.monthly/.sst/
Novoselov AP (2010) The significance of salmon-like fish in the ichthyofauna of the shelf zone of the Baydaratskaya Bay in the southwestern part of the Kara Sea. VI International contact forum on habitat conservation in the Barents region. NARFU, Arkhangelsk, pp 57 (in Russian)
Pogrebov VB, Kiyko OA, Stogov IA et al (2003) Composition, structure, and long-term dynamics of the ecosystem of the Pechora Bay.Monitoring of the Pechora Sea: present and future. Naryan-Mar, pp 35–39 (in Russian)
Romankevich EA, et al. (eds) (2003) The Pechora Sea. System Investigations, More (in Russian)
Semushin AV, Novoselov AP (2009) Species composition of the ichthyofauna of the Baidartskaya Gulf of the Kara Sea. J Ichthyol 5:362–375
Semushin AV, Sherstkov VS, Rukhlova VA (2011) Diversity of fish species in the trawl catches in the southeastern Barents Sea in 1980–2008. J Ichthyol 9:717–737
Shilov IA (1997) Ecology. Vysshaya shkola, Moscow (in Russian)
Smirnov OV, Dolgov AV, Guzenko VV (2000) New data on hydrological conditions and the ichthyofauna of the archipelagos Spitsbergen and Franz Josef Land. Materials of the final session of PINRO on results of research in 1998–1999. PINRO, Murmansk 1:79–92 (in Russian)
Ward JH Jr (1963) Hierarchical grouping to optimize an objective function. J Am Stat Assoc 301:236–244
Acknowledgements
We appreciate the help of SevPINRO colleagues, who collected fish samples during the expeditions. We are grateful to Dr. Alexey A. Sukhotin (Zoological institute RAS) for corrections and advice. Special thanks are due to Dr. Natalia I. Lavrinova (Northern [Arctic] Federal University named after M.V. Lomonosov), Dr. Natalia Lentsman and PhD Anna Zashikhina for the help with translation. We gratefully acknowledge the critical remarks made by anonymous referees.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
This article belongs to the special issue on the “Ecology of the Pechora Sea”, coordinated by Alexey A. Sukhotin.
Rights and permissions
About this article

Cite this article
Semushin, A.V., Novoselov, A.P., Sherstkov, V.S. et al. Long-term changes in the ichthyofauna of the Pechora Sea in response to ocean warming. Polar Biol 42, 1739–1751 (2019). https://doi.org/10.1007/s00300-018-2405-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-018-2405-3