
Abstract
Sea ice now forms later and melts earlier than it did 4 decades years ago, and it now melts completely in all parts of the Chukchi Sea. This decline in sea ice is expected to have repercussions on the trophic structure in this environment, and there are indications that changes already have taken place in the seabird community. We compared boat-based densities of seabirds in the eastern Chukchi Sea between July and October during 1975–1981 (historical data) with densities during 2007–2012 (recent data). We related the composition of the seabird community to sea-ice cover to explore how the community may be responding to changes in oceanography. The seabird community historically was composed predominantly of piscivorous Black-legged Kittiwakes (Rissa tridactyla) and murres (Uria spp.). In contrast, the seabird community now is composed predominantly of planktivorous seabirds such as Crested Auklets (Aethia cristatella) and Short-tailed Shearwaters (Puffinus tenuirostris). Total abundance of seabirds declined in three of four strata in the eastern Chukchi Sea, largely due to declines in densities of piscivorous and omnivorous species. These changes in the abundance and community composition of seabirds were associated with changes in ice cover. Earlier ice retreat appears to contribute to an environment that is more favorable to the sustained production of large oceanic copepods and euphausiids. We propose that long-term changes (4 decades) in the abundance and composition of the seabird community reflect an increase in the availability of large zooplankton prey in the region.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Summer ice cover in the Arctic Ocean reached the lowest minimal extent on record in 2012 (Laxon et al. 2013). Because sea-ice cover strongly influences habitat in the Arctic (Mueter and Litzow 2008; Hunt et al. 2013), this decline is anticipated to have repercussions on the trophic structure of this environment. Pelagic ecosystems in polar regions are particularly sensitive at multiple trophic levels to changes in ice cover (Kitaysky and Golubova 2000; Ainley et al. 2005; Beaugrand 2009; Kwasniewski et al. 2012). The effects of climate change on seabirds are mostly indirect, operating through changes in local and regional food webs and within pelagic habitats (Sydeman et al. 2012).
The timing and patterns of ice melt can alter the productivity of polar shelf seas by influencing water column structure and the availability of light for supporting photosynthesis (Hunt et al. 2011; Palmer et al. 2013). For example, in the northern Bering Sea, there has been a shift over the past 3 decades from a benthic-dominated system to one dominated numerically by pelagic organisms (Grebmeier et al. 2006b). The Chukchi Sea relies on water masses advected from the Bering Sea for much of its primary and secondary productivity (Grebmeier et al. 2006a), suggesting that changes observed in the northern Bering Sea also will affect the ecology of the Chukchi Sea. A second factor influencing productivity in the Chukchi Sea is environmental conditions during the open-water season that promote further growth of Pacific zooplankton once they reach the Chukchi Sea. We anticipate that as the open-water season begins earlier, the increase in light and water column heating in regions formerly covered with ice will promote further development of zooplankton biomass (Questel et al. 2013) that then is available as prey for higher trophic levels such as seabirds.
Historical studies of the avifaunal communities of the northeastern Chukchi Sea conducted in the late 1970s and early 1980s provided a snapshot of the community composition and density of seabirds (Divoky 1987, unpublished report) but did not address the variability of this community or link species to their habitat. Recent efforts to describe the circumpolar species diversity and distribution of marine birds (Bluhm et al. 2011; Huettmann et al. 2011) do not include regionally important taxa such as auklets and phalaropes that are critical to understanding energy flow in this ecosystem (Piatt and Springer 2003). Fortunately, many of the historical seabird surveys were conducted as part of the Outer Continental Shelf Environmental Assessment Program (OCSEAP) following protocols that are similar to those currently used to survey seabirds. The data from those historical surveys were archived by the U.S. Geological Survey (USGS) in the North Pacific Pelagic Seabird Database (NPPSD) and now offer a rare opportunity to combine historical data with surveys that have been conducted recently to assess the effects of climate change in Alaskan Arctic waters on a multi-decadal scale.
We compared boat-based survey data collected in the Chukchi Sea during 1975–1981 (historical period) with surveys conducted during 2007–2012 (recent period) to determine how the seabird community has responded to changes in the physical and biological oceanography of the Chukchi Sea. Specifically, our objectives were to (1) compare species composition of historical seabird communities with recent ones; (2) assess changes in the relative abundance of focal species between the two time periods; and (3) examine relationships between the seabird community and changes in the timing and extent of sea ice in the region over the past 4 decades that have driven changes in the open-water pelagic habitat.
Methods
Study area
In the Chukchi Sea, the net flow of water is northward through Bering Strait and toward the Arctic Ocean (Coachman et al. 1975). The broad northward flow through Bering Strait is steered by bathymetry into three main branches—one east of Hanna Shoal that feeds into Barrow Canyon, one west of Herald Shoal that feeds into Herald Valley, and one between the two shoals, referred to as the Central Channel flow (Fig. 1b; Weingartner et al. 1998, 2005). This separation also is evident in water mass properties (Weingartner et al. 2005, 2013) that define the oceanic habitat used by seabirds. Within the Chukchi Sea, the Alaska Coastal Current (ACC) lies east near the Alaska coastline and flows northward, carrying Alaskan Coastal Water (ACW), a warm (>2 °C), low-salinity (<32.2 psu) water mass that originates south of Bering Strait. The currents farther offshore move Bering Sea Water (BSW; Coachman et al. 1975), a warm (>2 °C), high-salinity (>32.4 psu) water mass, northward through the Central Channel and Herald Valley (Weingartner et al. 2005). This BSW is a mixture of Anadyr Water and Bering Shelf Water from south of Bering Strait that has a higher nutrient content and transports greater numbers of oceanic zooplankton, especially larger zooplankton, than does ACW (Walsh et al. 1989; Springer and McRoy 1993).

The Chukchi Sea, showing (a) oceanographic strata and survey locations and (b) currents and water flow
In addition to the warm, Bering-derived summer water masses advected northward through Bering Strait, Chukchi shelf waters are modified during the fall and winter by ice formation and during the spring by ice melt. Ice formation rejects brine that sinks to the sea floor and forms cold (−2 to +1 °C), salty (>32 psu) Winter Water (WW) below the ice layer. In the spring, ice melts at the surface and forms cold (−1 to +2 °C), low-salinity (<30 psu) Meltwater (MW) that is depleted of nutrients. This MW eventually becomes mixed with waters below by summer winds. In the summer, the bottom layer of the water column often still contains WW left over from the previous winter, whereas the surface layer consists of either MW or BSW, depending on location. Stratification increases from spring to summer and typically erodes in the fall as strong winds, cooling, and freezing enhance vertical mixing (Weingartner et al. 2005).
This study was conducted in the eastern Chukchi Sea, in an area bounded by Bering Strait to the south, the U.S.–Russia maritime boundary (168°58′37″W) to the west, and 73°N to the north and included a total area of 231,868 km2 (Fig. 1a). We divided the study area into 4 strata to account for the effects that latitude, water masses, and currents can have on determining oceanic habitat (Piatt and Springer 2003). The Southern Chukchi stratum has an area of 53,631 km2 and is influenced by strong flows through Bering Strait of Bering Shelf Water and Anadyr Water to the west and ACW to the east. The Nearshore stratum has an area of 35,673 km2 and is influenced by coastal flows that carry predominantly ACW, although there are episodic flow reversals in the northern half of this stratum that transport slope waters up Barrow Canyon (Aagaard and Roach 1990; Weingartner et al. 1999). This stratum also receives waters that flow eastward from the Central Channel that eventually enter Barrow Canyon (Weingartner et al., 2005). The Offshore stratum has an area of 81,159 km2 and is influenced by both the Central Channel flow that entrains Bering Sea Water along the western edge and resident waters over Hanna Shoal that are characterized by a two-layer water column with MW on top and WW on the bottom. The Northern stratum has an area of 61,405 km2 and includes the continental slope into the Arctic Basin. This stratum is influenced by waters flowing north from the Chukchi shelf and upwelling along the continental shelf-break from the Arctic Ocean.
Data collection
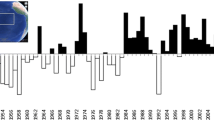
Historical data were collected by researchers during 1975–1981 (Fig. 2) from ships of opportunity following protocols developed by the U.S. Fish and Wildlife Service (USFWS; Tasker et al. 1984; Gould and Forsell 1989). These data were archived in the NPPSD, which is maintained by the USGS. We included historical data that were collected on the Russian side of the maritime boundary because the agreement establishing this boundary was not signed until 1990 (U.S. Department of State 1990). We conducted research cruises during 2007–2012 that covered much of the open-water period (roughly July–October) of the northeastern Chukchi Sea (Fig. 2) and followed a sampling protocol that was comparable to methods used to collect the historical data. We combined data sets from 4 recent studies that we conducted in the region to obtain coverage in all strata (Table 1): the Chukchi Sea Environmental Studies Program (CSESP; 2008–2012), the Alaska Monitoring and Assessment Program (AKMAP; 2010–2011), surveys conducted for the USFWS near Barrow (2009–2010); and surveys conducted by the USFWS on ships of opportunity (2007–2012; Kuletz and Labunski, unpubl. data).

Timing of ship-based surveys for marine birds in the Chukchi Sea, 1975–2012, by year and stratum. Horizontal dashed line separates historical data from recent data
We surveyed continuously when the ship was moving along a straight-line course at a minimal velocity of 9 km/h, recording environmental conditions every 10 min. We collected data 9–12 h/day during daylight hours, weather and sea-ice conditions permitting. We generally stopped surveys when sea height was Beaufort 6 (seas ~2–3 m) or higher, although we occasionally continued to sample if observation conditions were still acceptable (e.g., if seas were at the lower end of Beaufort 6 and we were traveling with the wind and seas). One observer stationed on the bridge of the vessel recorded all birds seen within a radius of 300 m in a 90° arc from the bow to the beam on one side of the ship (the count zone), locating and identifying seabirds with 10× binoculars as needed. For each bird or group of birds, we recorded species, total number of individuals, and behavior.
In the count zone, we counted all birds seen on the water, taking care to avoid recounting the same individuals. For flying birds, we conducted scans ~once/min (the exact frequency varied with ship’s speed) and recorded an instantaneous count (or “snapshot”) of all birds flying within the count zone. This snapshot method reduces the bias of overestimating the density of flying birds (Tasker et al. 1984; Gould and Forsell 1989). We counted only those flying birds that entered the count zone from the sides or front and excluded flying birds that entered from behind the ship (i.e., an area that already had been surveyed) to avoid the possibility of counting ship-following birds.
We entered all data directly into a computer connected to a Global Positioning System (GPS) with DLog software (R. G. Ford Consulting, Portland, OR) or TigerObserver software (TigerSoft, Las Vegas, NV). These computer programs time-stamped and georeferenced every observation and recorded the survey track line in ~20-s intervals.
Data analysis
Community analysis
We used multivariate analyses and descriptive statistics to explore seasonal and interannual changes in the structure of the seabird community. For the multivariate analysis, we included all bird observations that were identified to species. We grouped the data into sample units by stratum and year and used ordination for analysis. We used the species information to calculate a Bray–Curtis similarity matrix (Bray and Curtis 1957) to which we applied non-metric multidimensional scaling (nMDS; Clarke and Green 1988). This ordination technique then visualizes the similarity in overall community composition among samples as distances mapped in two-dimensional space (Clarke and Green 1988). The stress coefficient of the 2D ordinations was 0.177, indicating adequate fit to the data (Clarke and Ainsworth 1993). We examined the variation in species composition among strata and time periods with permutational multivariate analysis of variance (PERMANOVA), which partitions variation and requires no assumptions about the distribution or correlations among the variables (Anderson 2001). We used the package vegan (Oksanen et al. 2011) for the MDS analysis; all statistical analyses were conducted in R 3.0.1 (R Development Core Team 2013).
We included all bird observations that were identified at least to family in summaries of changes in species assemblages by stratum and year (Magurran 2004). We aggregated species into 4 ecological groups that represented foraging guilds (Table 2): planktivores, omnivores, piscivores, and benthivores. We examined differences between historical and recent periods to examine the hypothesis of a shift in community structure between periods.
Density analysis
We compared the density of birds during the historical period with density during the recent period in each of 4 strata of the eastern Chukchi Sea to determine whether there have been long-term changes in the abundance of the seabird community. For this comparison, we used total birds and 8 focal species from among the 10 most abundant species that represented the 4 foraging guilds, thereby providing an overview of functional ecological groups within the seabird community.
We pooled years within the 2 time periods and evaluated changes in abundance on each transect by using generalized linear models with period, region, and the interaction between these factors as explanatory variables (package MASS; Venables and Ripley 2002). We modeled the error terms with a negative binomial distribution to account for overdispersion in count data and used the area (km2) of each transect as an offset to account for survey effort. We excluded the Northern region in comparisons for species that were not recorded there during the historical time period. Because distance data were not available for the historical observations, we could not correct the density estimates for detection probability.
We summarized sea-ice conditions during the years included in our study to evaluate the direction and magnitude of change in ice cover. For the years 1979–2012, we used Special Sensor Microwave Imagery (SSM/I) passive microwave satellite sensor data from the National Snow and Ice Data Center (NSIDC; www.nsidc.org) to quantify the timing of ice advance, ice retreat, and changes in percent sea-ice cover in each stratum. These data are available from the fall and winter of 1978–1979 to the present. We use the daily data product from the standard 25-km grid derived with the NASA Team algorithm (Cavalieri et al. 2008) for consistent data resolution throughout the analysis period. Prior to 1979, data from satellites were less reliable, so, for the years 1975–1978, we used data from aerial surveys of ice cover in the Chukchi Sea. These original ice charts were scanned and summarized 4 times/month (1st, 8th, 15th, and 23rd days of each month) to provide ice cover (in tenths) on a grid 0.25°N × 0.25°W. Data are available from the Alaska Center for Climate Assessment and Policy (http://seaiceatlas.snap.uaf.edu/).
The date of sea-ice retreat was defined as the date when the sea-ice cover within a stratum was <10 %. Similarly, the date of sea-ice advance was defined as the date when sea-ice cover within a stratum was ≥10 %. The number of ice-free days was defined as the number of days elapsed between the date of sea-ice retreat and the date of sea-ice advance.
We also assessed the influence of changing sea-ice conditions on the density of seabirds directly by modeling the influence of days of ice-free water on the density of 6 species of seabirds that had sufficient sample size in both historical and recent periods. Because the 3 metrics of ice are highly correlated (r > 0.80), we selected only the number of days of ice-free water as the proxy for oceanographic change to avoid issues of collinearity. For this analysis, the number of ice-free days was calculated individually for each 25-km grid cell.
We built logistic regression models in which the presence of seabirds within a grid cell was the dependent variable and the duration of ice-free water and stratum (Southern, Nearshore, Offshore, and Northern) were independent variables. We started with full models, including the interaction of terms DAYS ICE-FREE and STRATUM and used backwards-stepwise removal of terms with P > 0.05 from Wald tests (Hosmer et al. 2013). Because the area sampled varied among grid cells, we included sampling area as an offset. We evaluated strength of the models with Akaike’s Information Criterion (AIC) and likelihood ratio tests comparing them to the null model. We drew inference from the model best supported by the data (Burnham and Anderson 2002). Detailed model selection results are provided in Online Resource 1.
Results
Community composition
We compiled seabird data collected in the eastern Chukchi Sea on 1,675 km2 of surveys during 1975–1981 and on 15,157 km2 of surveys during 2007–2012 (Table 1). These surveys recorded 58,905 birds during the historical period and 225,948 birds during the recent period. Mean densities ranged 0–19.9 birds/km2 in the historical period and 0–9.3 birds/km2 in the recent period (Table 2).
Over the past 37 years, the seabird community has included a total of 50 species, of which 39 were recorded in both historical and recent time periods (Table 2). Four species (Iceland Gull, Red-legged Kittiwake, Harlequin Duck, and Black Scoter) were recorded only during 1975–1981, and 7 species (Ancient Murrelet, Pelagic Cormorant, Mew Gull, Aleutian Tern, Pigeon Guillemot, Marbled Murrelet, and Fork-tailed Storm Petrel) were recorded only during 2007–2012.
Multivariate analyses of the seabird community indicated that species composition varied primarily between the two periods (Fig. 3a), shifting from a community dominated by piscivorous birds to one dominated by planktivorous birds (Fig. 4); this shift was evident in all strata (Fig. 3b). The MDS ordination separated into 2 groups, with overlap caused primarily by differences among strata (Table 3). The Northern stratum showed the biggest shift in community structure over time (Fig. 3b), converging in composition with the other regions in the recent years in which it was sampled.

Non-metric multidimensional scaling (nMDS) ordination plot of Bray–Curtis similarities for seabirds recorded in the Chukchi Sea during 1975–2012. Each point represents one stratum–year, and distance between points represents similarity in composition. Points are identified (a) by year, with period centroids labeled; and (b) by stratum, with arrows indicating the vector of change in composition by year for each stratum

Relative abundance of foraging guilds that compose the seabird community in the Chukchi Sea during historical (1975–1981) and recent (2007–2012) periods, by stratum
The patterns in species composition identified in the multivariate analyses were reflected in changes in the relative abundance of each of the 4 feeding guilds among strata and years (Figs. 4, 5). Most notably, the relative abundance of piscivores in all regions decreased from the historical period to the recent period (Fig. 4). The Northern stratum was dominated numerically by planktivores in 1976 and 2009–2012, but planktivores were absent from there in 1977 and rare in 2007, suggesting high variability in species composition (Fig. 5). The relative abundance of planktivores in the Offshore stratum increased from the 1970s to the 1980s, with planktivores dominating numerically in 2007–2012. In the historical period, the Nearshore and Southern strata were dominated numerically by planktivores in some years and by piscivores and benthic-feeding birds in other years; however, in the recent period, species composition was less variable, with planktivores dominating numerically in both strata in most years from 2007 to 2012. Benthic-feeding birds (primarily eiders) composed 2–85 % of all birds, depending on the stratum and year, because they sometimes occurred in large groups (up to 1,500 birds). Similarly, planktivores composed 67–90 % of all birds in some strata and historical years because Short-tailed Shearwaters also occurred sporadically and in large groups.

Relative abundance of foraging guilds that compose the seabird community in the Chukchi Sea, 1975–2012, by year and stratum
Changes in seabird density
The total density of seabirds has declined significantly in 3 of the 4 strata in the eastern Chukchi Sea over the past 37 years (P < 0.0001 for PERIOD*STRATUM), with the largest declines occurring in the Nearshore stratum (Fig. 6). Only the Northern stratum showed no significant difference in density between the two time periods. These declines are driven primarily by declines in the density of piscivorous and omnivorous species. Piscivorous Black-legged Kittiwakes and omnivorous Glaucous Gulls were less abundant in all strata during the recent period than during the historical period (Fig. 7). Declines of both species were greatest in the Nearshore stratum, with mean decreases of 16.5 birds/km2 for Black-legged Kittiwakes and 20.2 birds/km2 for Glaucous Gulls. Northern Fulmars were present in low abundance (<0.6 birds/km2) both historically and recently. Their density increased in the Northern, Nearshore, and Southern strata but showed no significant change in the Offshore stratum. Thick-billed Murres now are less abundant in the Offshore and Southern strata than they were in the historical period and showed no significant change in abundance in the Nearshore stratum during the recent period.

Density (birds/km2) of all seabirds in the eastern Chukchi Sea during historical (1975–1981) and recent (2007–2012) periods, by stratum. These values are predicted from generalized linear models that account for geographic stratum, period, and the negative binomial distribution of the data. Error bars represent 95 % confidence intervals

Density (birds/km2) of eight species of seabirds in the eastern Chukchi Sea during historical (1975–1981) and recent (2007–2012) periods, by stratum. These values are predicted from generalized linear models that account for geographic stratum, period, and the negative binomial distribution of the data. Error bars represent 95 % confidence intervals. Thick-billed Murres, Least Auklets, and Short-tailed Shearwaters in the Northern stratum could not be modeled because they were not recorded in the historical period
Planktivorous Crested and Least auklets and Short-tailed Shearwaters were more abundant in recent years than they were in historical years (Fig. 7), with the largest increases for Crested Auklets in the Offshore stratum (estimated mean increase 6.1 birds/km2), for Least Auklets in the Southern stratum (estimated mean increase 3.6 birds/km2), and for Short-tailed Shearwaters in the Nearshore stratum (estimated mean increase 7.1 birds/km2). Further, Least Auklets and Short-tailed Shearwaters were not recorded in the Northern Stratum in the historical period. In contrast, phalaropes were less abundant in the Southern stratum in recent years than they were in historical years (Fig. 7) and showed no significant change in abundance in the other strata.

Linear trends in (a) date of ice retreat, (b) date of ice advance, and (c) number of days with <10 % ice cover in the Chukchi Sea, 1975–2012, by year and stratum. Lines are the least-squares linear fit, and the gray shading is the 95 % confidence interval around the fit of the line
Changes in sea-ice cover
The duration of open water (defined as 25-km2 grid cells with <10 % ice cover) in the Northern, Offshore, and Nearshore strata of the eastern Chukchi Sea has increased by ≥50 days over the past 3 decades (Fig. 8). Sea ice now forms later and melts earlier than it did 30 years ago, and it now melts completely in all strata of the eastern Chukchi Sea. Prior to 1990, there were years in which only the Southern stratum would be considered ice-free. During the 1990s, the ice retreated from all but the Northern stratum in every year; since 2002, the entire eastern Chukchi Sea has been ice-free for at least 50 days each year.

Relationship between number of days with <10 % ice cover and predicted probability of occurrence for 6 species of seabirds in the eastern Chukchi Sea, 1975–2012. Values and 95 % confidence intervals are predicted from logistic regression models of the effects of the number of days with <10 % ice cover and geographic stratum on the probability of seabird occurrence
These changes in ice cover are associated with changes in the presence of seabirds, although the relationships between ice cover and seabird abundance varied among taxa and strata (Fig. 9). Black-legged Kittiwakes, Glaucous Gulls, and phalaropes were more likely to occur in years with few ice-free days. The negative association with ice-free days was strongest for these three species in the Southern and Nearshore strata. In the Northern stratum, Black-legged Kittiwakes and Glaucous Gulls also were more likely to occur in years with few ice-free days, whereas phalaropes had a low probability of occurrence, regardless of ice conditions. In the Offshore stratum, however, there was little evidence of a negative relationship with ice-free days for any of these species.
Crested Auklets, Northern Fulmars, and Short-tailed Shearwaters were more likely to occur in years with many ice-free days; this relationship was consistent in all strata (Fig. 9). The probability of occurrence for these three species generally was highest in the Offshore stratum. In the Northern stratum, the probability of occurrence for Crested Auklets was similar to that in the Offshore stratum in years with >50 ice-free days, whereas Short-tailed Shearwaters had a lower probability of occurrence than in the Offshore and Northern Fulmars were least likely to occur there. In the Nearshore stratum, probabilities of occurrence for all three species paralleled and were lower than probabilities in the Offshore stratum. In the Southern stratum, Northern Fulmars and Short-tailed Shearwaters had similar probabilities of occurrence, whereas Crested Auklets were least likely to occur there.
Discussion
Over the past 40 years, we found an increase in the proportion of subarctic seabirds, and particularly planktivorous species, in the eastern Chukchi Sea during the open-water period of late July to early October. Many species that breed or summer in the Bering Sea historically have moved northward into the Chukchi Sea during periods of maximal ice retreat (G. Divoky, unpubl. data). Our recent data indicate that, as summer ice cover has declined, a suite of planktivorous species, including Short-tailed Shearwaters, Least and Crested auklets, and Northern Fulmars, now occurs farther north and in higher abundance than it did historically. Although they penetrated beyond Cape Lisburne only in late August and early September in the 1970s and 1980s, we now see these species regularly as far as 72°N and remaining into late October. Other species such as Kittlitz’s and Ancient murrelets are more episodic in their use, showing up in large numbers in some years but rarely or not at all in others. Nonetheless, all of these Bering Sea species have become common throughout the eastern Chukchi Sea, suggesting that seabird distributions have changed in response to a warming and increasingly ice-free Arctic.
Changes in community composition
The seabird community of the Chukchi Sea contains a list of species that has changed little over the past 40 years, but the relative abundance of species has shifted toward planktivorous seabirds that are primarily coming north from the Bering Sea. Based on only the historical data, phalaropes were thought to replace auklets as the dominant planktivorous birds north of Bering Strait (Piatt and Springer 2003); in fact, planktivorous seabirds in general were assumed to be insignificant consumers in the Chukchi Sea as a whole (Piatt and Springer 2003; Hunt et al. 2013). The data presented here and in other recent studies indicate that, although planktivorous seabirds did not dominate the seabird community in the 1970s and early 1980s, they have become the numerically dominant feeding guild of the 2000s and 2010s, particularly those species common in the Bering Sea such as Crested Auklets and Short-tailed Shearwaters (Sigler et al. 2011; Gall et al. 2013; Wong et al. 2014; Kuletz et al. 2015).
Species typically associated with the high Arctic and sea ice include Ivory and Ross’s gulls and Black Guillemots. Historically, they occurred consistently in the Chukchi Sea, but in low numbers (Watson and Divoky 1970 [unpubl. report]; Divoky 1987 [unpubl. report]). They rarely have been recorded in recent years, despite a large increase in effort relative to historical surveys. Ice has retreated entirely from the Chukchi Shelf in every summer since 1996 and is delayed in re-forming in the fall, restricting the habitat available for these birds.
Species recorded in the eastern Chukchi Sea only recently are ones that either nest in the Bering Sea and/or North Pacific or have made the transit from the Atlantic Ocean. The Short-tailed Albatross, Rhinoceros Auklet, Ancient Murrelet, Mew Gull, and Fork-tailed Storm Petrel all are Pacific species that have been able to extend their ranges northward into the Chukchi, presumably in response to reductions in ice and warming climatic conditions (Day et al. 2013a). Incidental observations recently recorded off-transect included the Northern Gannet (Morus bassanus), an Atlantic species that was recorded in the Pacific for the first time in 2010 (Day et al. 2013a). It presumably traversed the Northwest Passage during a period of open water. If so, it is one of three seabird species from the North Atlantic recorded in the Pacific in recent years (Atlantic Puffin [Fratercula arctica]—Kharitonov 2009; Great Black-backed Gull [Larus marinus]—Day et al. 2013a). Of the species recorded only during the historical period, Red-legged Kittiwakes are considered endemic to the Bering Sea and Iceland Gulls are a northwestern Atlantic species. These species may have been vagrant individuals or may have been misidentified: Red-legged Kittiwakes are very similar to the more common Black-legged Kittiwake, and Iceland Gulls are very similar to Thayer’s Gulls, which migrate through the Chukchi Sea (Snell 2002). We anticipate that, as summer ice cover continues to decline and ship traffic increases, incidental observations of Bering Sea and Atlantic species will become more common in the Chukchi Sea.
Changes in abundance
As species that breed in the Bering Sea (e.g., Crested and Least auklets, Northern Fulmars) or even farther south in Australia (Short-tailed Shearwaters) have become more abundant in the eastern Chukchi Sea over the past 40 years, species that breed along the Chukchi coast (e.g., Black-legged Kittiwakes, Thick-billed Murres) and inland on the Arctic Coastal Plain (e.g., Glaucous Gulls, phalaropes) appear to be less abundant. These latitudinal shifts in abundance are consistent with the retreat of sea ice opening up habitat and creating conditions in the Chukchi Sea that more closely resemble the productive areas of the northern Bering Sea (Sigler et al. 2011). Species that rely primarily on prey of Pacific origin (Crested Auklets, Northern Fulmars, and Short-tailed Shearwaters) are more likely to occur as sea-ice cover declines, whereas those that forage in Arctic waters (Black-legged Kittiwakes, Glaucous Gulls, and phalaropes) are less likely to occur, suggesting that ice cover is an ecological driver that affects variations in seabird abundance interannually and over longer timescales.
Birds that are surveyed at sea include both breeding and non-breeding individuals, providing an ecosystem perspective that includes free-ranging animals not necessarily restricted by central-place foraging (Ainley et al. 2012). The distribution and abundance of seabirds within 40 km of large breeding colonies such as the Diomede Islands, Cape Lisburne, and Cape Thompson may be influenced by the availability of nesting habitat (Swartz 1967; Piatt and Springer 2003; Wong et al. 2014). Our data, however, spanned the entire open-water season, including September and October—after birds have left the breeding colonies. The birds that we surveyed included non-breeding birds, birds that may have attempted to breed but failed, and birds that bred successfully. Regardless of their breeding status in a given year, most birds are free of their land-bound constraints by September, so their presence and abundance in the study area implies that food resources are available and energetically profitable to acquire.
Decline of piscivorous species
The decline in the density of seabirds in the eastern Chukchi Sea was driven primarily by declines in density of Black-legged Kittiwakes and Glaucous Gulls in the Nearshore stratum (i.e., within 40 km of the coast). The Southern stratum also has experienced a decline in density of total birds, primarily due to declines of Black-legged Kittiwakes, Thick-billed Murres, and phalaropes. Black-legged Kittiwakes and Thick-billed Murres are more cosmopolitan in their distribution than are Glaucous Gulls and phalaropes, with their summer ranges including the Pacific Arctic and subarctic. Like Glaucous Gulls, however, Black-legged Kittiwakes and murres are flexible foragers that also take prey other than oceanic zooplankton and often are associated with ACW (Swartz 1967; Piatt and Springer 2003). Phalaropes are planktivorous, but they are restricted to foraging at or near the surface and on Calanus copepods that are characteristic of ACW (Brown and Gaskin 1988; Haney and Stone 1988). The declines in abundance of these 4 species that rely on prey resources associated with ACW suggest a possible reduction in the abundance of fish species in that water mass as annual sea-ice cover has declined. Alternatively, declines in the density of these birds may be related to the expansion of available habitat as sea ice retreats earlier. With an increase in the amount of available habitat, the birds can disperse over a larger area than they could previously, decreasing at-sea densities while the total abundance in the region remains similar. Although the decline in density would account for the apparent decline in abundance of piscivorous birds in the Nearshore stratum, it does not account for the increase in density of planktivorous birds, particularly Short-tailed Shearwaters.
Population trends measured at one of the two biggest seabird colonies in the eastern Chukchi Sea provide some additional insight into the long-term change in foraging conditions in ACW. The productivity of Black-legged Kittiwakes shows a long-term decline (chicks/nest start) during the 40 years that nests have been monitored at the Cape Lisburne breeding colony, with productivity below the long-term (1976–2012) colony mean in all but 2 years since 2000 (Dragoo et al. 2012). Despite this local decline in productivity, reproductive success of colonies in the Chukchi Sea remains higher than productivity at colonies in the central and southern Bering Sea (Dragoo et al. 2015). Counts of adult kittiwakes and Thick-billed Murres present at the colony have increased during the same period, possibly because of immigration from colonies in the Bering Sea. This decline in productivity coupled with an increase in the number of birds loafing at the colony rather than engaging in chick-rearing suggests that food availability may be limiting breeding activity (Golet et al. 2004). Because they are long-lived species, these birds may be foregoing breeding to maximize their own survival (Golet et al. 1998). Rather than traveling north to forage in the Chukchi Sea after the breeding season, piscivorous species may be heading south to foraging locations in the Bering Sea where fish and other preferred prey may be more readily available (Sigler et al. 2011; Stabeno et al. 2012, Orben et al. 2015a, b).
Increase of planktivorous species
Shearwaters have been among the most abundant seabirds in the Chukchi Sea since the earliest shipboard surveys were conducted (Swartz 1967; Watson and Divoky 1970 [unpubl. report]; Divoky 1987 [unpubl. report]). In the historical period, they were not recorded in the Northern stratum of the Chukchi Sea and occurred irregularly in the Beaufort Sea (Divoky 1987 [unpubl. report]), indicating that the Chukchi represented the northernmost limit of annual migration from breeding colonies in Australia for most of these birds. In the past 40 years, their abundance in the Chukchi Sea has increased, and they now are being recorded nearly annually in the Beaufort Sea (USGS 2014). Recent surveys determined that they continue to be among the most abundant birds in the Chukchi Sea and that they are most abundant within 40 km of the coast, where they are foraging in ACW (Gall et al. 2013; Wong et al. 2014; Kuletz et al. 2015). This increase is a contrast to the decline in abundance of piscivorous birds within the same stratum.
The Offshore stratum that historically was dominated numerically by Black-legged Kittiwakes and Thick-billed Murres has been taken over by Crested Auklets, which are now the most abundant seabirds in August and September (Gall et al. 2013; Kuletz et al. 2015) despite not nesting north of Bering Strait. The increase in abundance of Crested Auklets is consistent with the hypothesis that large numbers of euphausiids now are transported through the Central Channel of the Chukchi Sea (Berline et al. 2008; Ashjian et al. 2010). Not only are Crested Auklets feeding in the Offshore stratum during and after the breeding season, it appears that they may be flightless for a few weeks in the area as they complete their molt (AEG, RHD, KJK, pers. obs.). The limited mobility of these birds and the energetic demands associated with molt suggest that their food supply is abundant and reliable enough to support these physiological demands.
There are, of course, challenges in making comparisons with historical data, particularly when those data were collected opportunistically and by different researchers. The historical data were collected from ships of opportunity where the tracks were nonrandom and the survey coverage differed among years and strata. The only stratum that was surveyed in all 6 years of the historical period was the Nearshore stratum, whereas survey coverage in the Northern stratum was limited due to heavy ice cover. The reduction in ice cover of the past 40 years means that there has been an increase in the availability of open water and, therefore, greater opportunity for ship-based surveys in the northern parts of the Chukchi Sea. Survey coverage also was greatly increased in the Offshore stratum in recent years because of interest in areas that are currently leased for oil and gas exploration. Despite the uneven sampling coverage, the trends in all areas of the eastern Chukchi Sea consistently indicate a northward expansion of subarctic seabirds, especially planktivorous ones, into the Chukchi Sea.
System-wide changes and implications
Earlier ice retreat and a longer ice-free season (Wang and Overland 2009; Wang et al. 2012) may contribute to an environment that is more favorable to primary and secondary production. The increase of light availability through thinner winter ice and the earlier ice retreat can lead to higher production by ice algae along the coast and at ice edges (Brown and Arrigo 2012; Palmer et al. 2013). So far, models base on satellite observations suggest there was a 65 % increase in primary productivity in the Chukchi Sea associated with the increase in open water and the longer growing season in 2007 than there was in 1998–2002 (Arrigo et al. 2008). In a shallow Chukchi Sea with less ice cover, increased easterly winds could intensify shelf-break upwelling (Carmack and Chapman 2003; Mathis et al. 2012) that will enhance the nutrient supply and support larger phytoplankton and zooplankton stocks throughout the open-water season (Lane et al. 2008, Ershova et al. 2015).
In addition to an increase of in situ production in the Chukchi Sea, there also is evidence that northward flow through Bering Strait is increasing (Woodgate et al. 2012), a change that should increase the advection of phytoplankton and zooplankton into the Chukchi Sea (Springer et al. 1989). These Pacific zooplankters are being advected into a region where warming summer waters can sustain higher growth rates and develop greater zooplankton biomass (Questel et al. 2013) than they historically could. Although the historical zooplankton community is not as well documented as the seabird community, there is evidence of higher zooplankton abundance and biomass along the Chukchi shelf-break in the 2000s than historically (Lane et al. 2008) and of increased biomass and abundance of Bering-Pacific zooplankton over the northeastern Chukchi shelf from 1946 to 2012 (Matsuno et al. 2011, Ershova et al. 2015). Concurrently, there has been an increase in the bowhead whale population of 3.7 %/year during 1978–2011 (Givens et al. 2013). These whales prey on zooplankton, especially euphausiids, that are the primary prey of Crested Auklets and Short-tailed Shearwaters, and they transit the Chukchi Sea to feed in and near Barrow Canyon (Berline et al. 2008; Citta et al. 2015). The steady annual increase in the bowhead population indicates that their preferred prey is readily available.
The Chukchi Sea is a transition zone between the highly productive waters of the northern Bering Sea and the low productivity of the Arctic Ocean. As recently as 2003, the boundary between these two systems was found to lie in the southern Chukchi Sea (Piatt and Springer 2003). As sea ice retreats earlier in the summer and the Chukchi remains ice-free for longer each year, the boundary between these two systems appears to be shifting northward. These connections have stretched the biogeographic province characterized by zooplankton, surface fish, and seabirds associated with Bering Sea Water so that it now extends from the Chirikov Basin north of St. Lawrence Island to include the eastern Chukchi Sea as far north as Barrow Canyon (Sigler et al. 2011).
We propose that the changes in the abundance and composition of the seabird community in the eastern Chukchi Sea reflect an increase in the availability of zooplankton prey in the region (Ershova et al. 2015). The increase in abundance of planktivorous species of seabirds in the Chukchi Sea suggests that large zooplankton persist in the water column of the Chukchi Sea for longer now than they did 40 years ago. This longer persistence allows for the seabird community, especially planktivorous alcids, to remain in the Chukchi Sea throughout the summer and fall to exploit this food resource that now is available over the wide and shallow Chukchi shelf. As long as the Chukchi Sea continues to freeze in winter, however, the process of ice formation will maintain a pool of cold Winter Water that acts a thermal barrier to many Bering Sea species of fishes, particularly in the offshore area (Sigler et al. 2011; Hunt et al. 2013; Day et al. 2013b). Without a diverse community of plankton-feeding fishes, seabirds and marine mammals may remain the primary predators on zooplankton in the Chukchi Sea.
The reduction in ice cover of the Chukchi Sea is not only opening up habitat for birds, whales, and zooplankton, it is providing humans the opportunity to explore for oil and gas, develop commercial shipping routes, and consider the viability of a commercial fishery. This study provides a long-term perspective on the environment of the Chukchi Sea to help inform policy-makers and managers responsible for spatial planning, permitting, and ecosystem-based management of marine resources (Schwemmer et al. 2011; Ainley et al. 2012). The possible effects of anthropogenic stressors must be considered in light of the shifting baseline of a seabird community and an entire marine ecosystem that is actively responding to a warming and increasingly ice-free Arctic.
References
Aagaard K, Roach AT (1990) Arctic ocean-shelf exchange: measurements in Barrow Canyon. J Geophys Res 95:18163–18175. doi:10.1029/JC095iC10p18163
Ainley DG, Clarke ED, Arrigo K, Fraser WR, Kato A, Barton KJ, Wilson PR (2005) Decadal-scale changes in the climate and biota of the Pacific sector of the Southern Ocean, 1950s to the 1990s. Antarct Sci 17:171–182. doi:10.1017/S0954102005002567
Ainley D, Ribic C, Woehler E (2012) Adding the ocean to the study of seabirds: a brief history of at-sea seabird research. Mar Ecol Progr Ser 451:231–243. doi:10.3354/meps09524
Anderson MJ (2001) A new method for non parametric multivariate analysis of variance. Aust Ecol 26:32–46. doi:10.1111/j.1442-9993.2001.01070
Arrigo KR, van Dijken G, Pabi S (2008) Impact of a shrinking Arctic ice cover on marine primary production. Geophys Res Lett 35:L19603. doi:10.1029/2008GL035028
Ashjian CJ, Braund SR, Campbell RG, Geroge JC, Kruse J, Maslowski W, Moore SE, Nicolson CR, Okkonen SR, Sherr BF, Sherr EB, Spitz YF (2010) Climate variability, oceanography, bowhead whale distribution, and Iñupiat subsistence whaling near Barrow, Alaska. Arctic 63:179–194
Beaugrand G (2009) Decadal changes in climate and ecosystems in the North Atlantic Ocean and adjacent seas. Deep Sea Res Part II Top Stud Oceanogr 56:656–673. doi:10.1016/j.dsr2.2008.12.022
Berline L, Spitz YH, Ashjian CJ, Campbell RG, Maslowski W, Moore SE (2008) Euphausiid transport in the western Arctic Ocean. Mar Ecol Progr Ser 360:163–178. doi:10.3354/meps07387
Bluhm BA, Gebruk AV, Gradinger R, Hopcroft RR, Huettmann F, Kosobokova KN, Sirenko BI, Weslawski JM (2011) Arctic marine biodiversity: an update of species richness and examples of biodiversity change. Oceanography 24:232–248
Bray JR, Curtis JT (1957) An ordination of the upland forest communities of southern Wisconsin. Ecol Monogr 27:325–349
Brown Z, Arrigo K (2012) Contrasting trends in sea ice and primary production in the Bering Sea and Arctic Ocean. ICES J Mar Sci 69:1180–1193
Brown RGB, Gaskin DE (1988) The pelagic ecology of the Grey and Red-necked Phalaropes phalaropes Phalaropus fulicarius and P. lobatus in the Bay of Fundy, eastern Canada. Ibis 130:234–250. doi:10.1111/j.1474-919X.1988.tb00974.x
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information–theoretic approach, 2nd edn. Springer, New York 496 pp
Carmack E, Chapman DC (2003) Wind-driven shelf/basin exchange on an Arctic shelf: the joint roles of ice cover extent and shelf-break bathymetry. Geophys Res Lett 30:1778. doi:10.1029/2003GL017526
Cavalieri D, Parkinson CL, Gloersen P, Zwally HJ (2008) Sea ice concentrations from Nimbus-7 SMMR and DMSP SSM/I passive microwave data, 1998–2009. http://nsidc.org/data/nsidc-0051
Citta JJ, Quakenbush LT, Okkonen SR, Druckenmiller ML, Maslowski W, Clement-Kinney J, Goerge JC, Brower H, Small RJ, Ashjian CJ, Harwood LA, Heide-Jorgensen MP (2015) Ecological characteristics of core-use areas used by Bering–Chukchi–Beaufort (BCB) bowhead whales, 2006–2012. Progr Oceanogr 136:201–222. doi:10.1016/j.pocean.2014.08.012
Clarke K, Ainsworth M (1993) A method of linking multivariate community structure to environmental variables. Mar Ecol Progr Ser 92:205–219
Clarke K, Green R (1988) Statistical design and analysis for a“biological effects” study. Mar Ecol Progr Ser 46:213–226
Coachman LK, Aagaard K, Tripp RB (1975) Bering Strait: the regional physical oceanography. University of Washington Press, Seattle
Day RH, Gall AE, Morgan TM, Rose JR, Plissner JH, Sanzenbacher PM, Fenneman JD, Kuletz KJ, Watts BH (2013a) Seabirds new to the eastern Chukchi and Beaufort seas, Alaska: response to a changing climate? Western Birds 44:174–182
Day RH, Weingartner TJ, Hopcroft RR, Aerts LAM, Blanchard AL, Gall AE, Gallaway BJ, Hannay DE, Holladay BA, Mathis JT, Norcross BL, Questel JM, Wisdom SS (2013b) The offshore northeastern Chukchi Sea, Alaska: a complex high-latitude ecosystem. Cont Shelf Res 67:147–165. doi:10.1016/j.csr.2013.02.002
Dragoo DE, Kettle AB, Bolin DE (2012) Biological monitoring at Cape Lisburne, Alaska in 2012. U.S. Fish and Wildlife Service report AMNWR2012/06. Homer, Alaska. https://absilcc.org/science/amnwr/sitepages/library.aspx
Dragoo DE, Renner HM, Irons DB (2015) Breeding status and population trends of seabirds in Alaska, 2014. U.S. Fish and Wildlife Service report AMNWR2015/03. Homer, Alaska. https://absilcc.org/science/amnwr/sitepages/library.aspx
Ershova EA, Hopcroft RR, Kosobokova KN, Matsuno K, Nelson RJ, Yamaguchi A, Eisner LB (2015) Long-term changes in summer zooplankton communities of the western Chukchi Sea, 1945–2012. Oceanography 28:100–115
Gall AE, Day RH, Weingartner TJ (2013) Structure and variability of the marine-bird community in the northeastern Chukchi Sea. Cont Shelf Res 67:96–115. doi:10.1016/j.csr.2012.11.004
Golet GH, Irons DB, Estes JA (1998) Survival costs of chick rearing in Black-legged Kittiwakes. J Anim Ecol 67:827–841. doi:10.1046/j.1365-2656.1998.00233.x
Golet GH, Schmutz JA, Irons DB, Estes JA (2004) Determinants of reproductive costs in the long-lived Black-legged Kittiwake: a multiyear experiment. Ecol Monogr 74:353–372
Gould PJ, Forsell DJ (1989) Techniques for shipboard surveys of marine birds. Fish and Wildlife Technical Report 25, US Fish and Wildlife Service, Anchorage, AK. 24 p. http://alaska.usgs.gov/science/biology/seabirds_foragefish/products/protocols/gould_seabird_protocol.pdf
Grebmeier J, Cooper L, Feder H, Sirenko B (2006a) Ecosystem dynamics of the Pacific-influenced Northern Bering and Chukchi seas in the Amerasian Arctic. Progr Oceanogr 71:331–361. doi:10.1016/j.pocean.2006.10.001
Grebmeier JM, Overland JE, Moore SE, Farley EV, Carmack EC, Cooper LW, Frey KE, Helle JH, McLaughlin FA, McNutt SL (2006b) A major ecosystem shift in the northern Bering Sea. Science 311:1461–1464. doi:10.1126/science.1121365
Haney JC, Stone AE (1988) Littoral foraging by Red Phalaropes during spring in the northern Bering Sea. Condor 90:723–726
Hosmer DW, Lemeshow S, Sturdivant RX (2013) Applied logistic regression, 3rd edn. Wiley, New York
Huettmann F, Artukhin Y, Gilg O, Humphries G (2011) Predictions of 27 Arctic pelagic seabird distributions using public environmental variables, assessed with colony data: a first digital IPY and GBIF open access synthesis platform. Mar Biodivers 41:141–179. doi:10.1007/s12526-011-0083-2
Hunt GL, Coyle KO, Eisner LB et al (2011) Climate impacts on eastern Bering Sea foodwebs: a synthesis of new data and an assessment of the Oscillating Control Hypothesis. ICES J Mar Sci 68:1230–1243. doi:10.1093/icesjms/fsr036
Hunt GL, Blanchard AL, Boveng P, Dalpadado P, Drinkwater KF, Eisner L, Hopcroft RR, Kovacs KM, Norcross BL, Renaud P, Reigstad M, Renner M, Skoldjal HR, Whitehouse A, Woodgate RA (2013) The Barents and Chukchi seas: comparison of two Arctic shelf ecosystems. J Mar Syst 109–110:43–68. doi:10.1016/j.jmarsys.2012.08.003
Kitaysky AS, Golubova EG (2000) Climate change causes contrasting trends in reproductive performance of planktivorous and piscivorous alcids. J Anim Ecol 69:248–262. doi:10.1046/j.1365-2656.2000.00392.x
Kuletz KJ, Ferguson MC, Hurley B et al (2015) Seasonal spatial patterns in seabird and marine mammal distribution in the eastern Chukchi and western Beaufort seas: identifying biologically important pelagic areas. Progr Oceanogr 136:175–200
Kwasniewski S, Gluchowska M, Walkusz W, Karnovsky NJ, Jakubas D, Wojczulanis-Jakubas K, Harding AMA, Goszczko I, Cisek M, Beszczynska-Moller A, Walczowski W, Weslawski JM, Stempniewicz L (2012) Interannual changes in zooplankton on the West Spitsbergen Shelf in relation to hydrography and their consequences for the diet of planktivorous seabirds. ICES J Mar Sci 69:890–901. doi:10.1093/icesjms/fss076
Lane P, Llinas L, Smith S, Pilz D (2008) Zooplankton distribution in the western Arctic during summer 2002: hydrographic habitats and implications for food chain dynamics. J Mar Syst 70:97–133. doi:10.1016/j.jmarsys.2007.04.001
Laxon SW, Giles KA, Ridout AL et al (2013) CryoSat-2 estimates of Arctic sea ice thickness and volume. Geophys Res Lett 40:1–6. doi:10.1002/GRL.50193
Magurran AE (2004) Measuring biological diversity. Blackwell Publishing, Malden
Mathis JT, Pickart RS, Byrne RH, McNeil CL, Moore GWK, Juranek LW, Liu X, Ma J, Easley RA, Elliot MM, Cross JN, Reisdorph SC, Bahr F, Morison J, Lichendorf T, Feely RA (2012) Storm-induced upwelling of high pCO2 waters onto the continental shelf of the western Arctic Ocean and implications for carbonate mineral saturation states. Geophys Res Lett 39:L07606. doi:10.1029/2012GL051574
Matsuno K, Yamaguchi A, Hirawake T, Imai I (2011) Year-to-year changes of the mesozooplankton community in the Chukchi Sea during summers of 1991, 1992 and 2007, 2008. Polar Biol 34:1349–1360. doi:10.1007/s00300-011-0988-z
Mueter FJ, Litzow MA (2008) Sea ice retreat alters the biogeography of the Bering Sea continental shelf. Ecol Appl 18:309–320
Oksanen J, Guillaume Blanchet F, Kindt R et al (2011) Vegan: community ecology package. http://vegan.r-forge.r-project.org/
Orben RA, Irons DB, Paredes R, Roby DD, Phillips RA, Shaffer SA (2015a) North or South? Niche separation of endemic Red-legged Kittiwakes and sympatric Black-legged Kittiwakes during their non-breeding migrations. J Biogeogr 42:401–412
Orben RA, Paredes R, Roby DD, Irons DB, Shaffer SA (2015b) Body size affects individual winter foraging strategies of Thick-billed Murres in the Bering Sea. J Anim Ecol 84:1589–1599. doi:10.1111/1365-2656.12410
Palmer MA, van Dijken GL, Mitchell BG, Seegers BJ, Lowry KE, Mills MM, Arrigo KR (2013) Light and nutrient control of photosynthesis in natural phytoplankton populations from the Chukchi and Beaufort seas, Arctic Ocean. Limnol Oceanogr 58:2185–2205. doi:10.4319/lo.2013.58.6.2185
Piatt JF, Springer AM (2003) Advection, pelagic food webs and the biogeography of seabirds in Beringia. Mar Ornithol 31:141–154
Questel JM, Clarke C, Hopcroft RR (2013) Seasonal and interannual variation in the planktonic communities of the northeastern Chukchi Sea during the summer and early fall. Cont Shelf Res 67:23–41. doi:10.1016/j.csr.2012.11.003
R Development Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org
Schwemmer P, Mendel B, Sonntag N et al (2011) Effects of ship traffic on seabirds in offshore waters: implications for marine conservation and spatial planning. Ecol Appl 21:1851–1860
Sigler M, Renner M, Danielson S, Eisner L (2011) Fluxes, fins, and feathers: relationships among the Bering, Chukchi, and Beaufort seas in a time of climate change. Oceanography 24:250–265
Snell RR (2002) Thayer’s Gull (Larus glaucoides). In: Poole A (ed) The birds of North America online. No. 699, Cornell Lab of Ornithology, Ithaca, NY. http://bna.birds.cornell.edu/bna/species/699b
Springer AM, McRoy CP (1993) The paradox of pelagic food webs in the northern Bering Sea—III. Patterns of primary production. Cont Shelf Res 13:575–599
Springer A, McRoy C, Turco KR (1989) The paradox of pelagic food webs in the northern Bering Sea—II. Zooplankton communities. Cont Shelf Res 9:359–386. doi:10.1016/0278-4343(89)90039-3
Stabeno PJ, Jr Farley E V, Kachel NB, Moore S, Mordy CW, Napp JM, Overland JE, Pinchuk AI, Sigler MF (2012) A comparison of the physics of the northern and southern shelves of the eastern Bering Sea and some implications for the ecosystem. Deep Sea Res Part II Top Stud Oceanogr 65–70:14–30. doi:10.1016/j.dsr2.2012.02.019
Swartz LG (1967) Distribution and movement of birds in the Bering and Chukchi seas. Pacific Sci 21:332–347
Sydeman WJ, Thompson SA, Kitaysky A (2012) Seabirds and climate change: roadmap for the future. Mar Ecol Progr Ser 454:107–117. doi:10.3354/meps09806
Tasker ML, Hope Jones P, Dixon T, Blake BF (1984) Counting seabirds at sea from ships: a review of methods employed and a suggestion for a standardized approach. Auk 101:567–577
U.S. Department of State (1990) Agreement with the Union of Soviet Socialist Republics on the maritime boundary. Senate Treaty Document 101–22. http://www.state.gov/e/oes/ocns/opa/c28187.htm
USGS (2014) North Pacific Pelagic Seabird Database. http://alaska.usgs.gov/science/biology/nppsd/index.php
Venables WN, Ripley BD (2002) Modern applied statistics with S, 4th edn. Springer, New York
Walsh J, McRoy C, Coachman L, Goering JJ, Nihoul JJ, Whitledge TE, Blackburn TH, Parker PL, Wirick CD, Shuert PG, Grebmeier JM, Speinger AM, Tripp RD, Hansell DA, Djenidi S, Deleersnijder E, Henriksen K, Lund BA, Andersen P, Müller-Karger FE, Dean K (1989) Carbon and nitrogen cycling within the Bering/Chukchi Seas: source regions for organic matter affecting AOU demands of the Arctic Ocean. Progr Oceanogr 22:277–359
Wang M, Overland JE (2009) A sea ice free summer Arctic within 30 years? Geophys Res Lett 36:L07502. doi:10.1029/2012GL052868
Wang M, Overland JE, Stabeno P (2012) Future climate of the Bering and Chukchi seas projected by global climate models. Deep Res Part II 65–70:46–57. doi:10.1016/j.dsr2.2012.02.022
Weingartner T, Cavalieri D, Aagaard K, Sasaki Y (1998) Circulation, dense water formation, and outflow on the northeast Chukchi shelf. J Geophys Res 103:7647–7661
Weingartner TJ, Danielson S, Sasaki Y, Pavlov V, Kulakov M (1999) The Siberian Coastal Current: a wind- and buoyancy-forced Arctic coastal current. J Geophys Res 104:29697. doi:10.1029/1999JC900161
Weingartner T, Aagaard K, Woodgate R, Danielson S, Sasaki Y, Cavalieri D (2005) Circulation on the north central Chukchi Sea shelf. Deep Sea Res Part II Top Stud Oceanogr 52:3150–3174. doi:10.1016/j.dsr2.2005.10.015
Weingartner T, Dobbins E, Danielson S, Winsor P, Potter R, Statscewich H (2013) Hydrographic variability over the northeastern Chukchi Sea shelf in summer–fall 2008–2010. Cont Shelf Res 67:5–22. doi:10.1016/j.csr.2013.03.012
Wong SN, Gjerdrum C, Morgan KH, Mallory ML (2014) Hotspots in cold seas: the composition, distribution, and abundance of marine birds in the North American Arctic. J Geophys Res Ocean 119:1691–1705. doi:10.1002/2013JC009198
Woodgate RA, Weingartner TJ, Lindsay R (2012) Observed increases in Bering Strait oceanic fluxes from the Pacific to the Arctic from 2001 to 2011 and their impacts on the Arctic Ocean water column. Geophys Res Lett. doi:10.1029/2012GL054092
Acknowledgments
Data collection and analysis conducted by ABR, Inc. was funded by ConocoPhillips Company, Anchorage, AK; Shell Exploration and Production Company, Anchorage, AK; and Statoil USA E&P, Inc., Anchorage, AK. Data collection by USFWS was funded with grants from the North Pacific Research Board (2007–2010), the Bureau of Ocean Energy Management (2010–2012), and the USFWS. The data collection, analysis, and interpretation, however, were conducted by the authors; the conclusions are ours and do not necessarily represent the views of the funding organizations. We particularly thank scientists Caryn Rea (ConocoPhillips), Michael Macrander (Shell), and Steinar Eldøy (Statoil) for support and feedback during all phases of this research. We also thank ABR and USFWS personnel for data collection in 2007–2012 and thank the many contributors to the North Pacific Pelagic Seabird Database over the past 4 decades. In particular, we thank Elizabeth Labunski (USFWS) for assisting with data collection and management. Gary Drew and John Piatt (US Geological Survey) maintain the NPPSD and provided much-appreciated assistance. Allison Zusi (ABR) provided invaluable GIS support. Bill Chapman (University of Illinois) and Lena Krutikov (Alaska Center for Climate Assessment & Policy) provided access to historical aerial surveys of ice cover. At the University of Alaska Fairbanks, we thank Arny Blanchard for statistical advice and Seth Danielson, John Walsh, and Peter Winsor for assistance with the summary of sea-ice data. This manuscript has been improved by the comments and suggestions of three anonymous reviewers.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Gall, A.E., Morgan, T.C., Day, R.H. et al. Ecological shift from piscivorous to planktivorous seabirds in the Chukchi Sea, 1975–2012. Polar Biol 40, 61–78 (2017). https://doi.org/10.1007/s00300-016-1924-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-016-1924-z