
Abstract
Conjugating green algae of the genus Zygnema (Zygnematophyceae, Streptophyta) are dominant eukaryotic components of hydro-terrestrial microbial mats in the Arctic. Considering the harsh environmental conditions, the aim of this study was to elucidate mechanisms that enable Zygnema spp. to thrive in this habitat. We hypothesized that changes in morphology, physiological performance, and stress tolerance take place during the annual life cycle of the algae. We thus selected four natural populations of Zygnema spp. on Svalbard and investigated them throughout the vegetation season by means of light microscopy and chlorophyll a fluorescence. Additionally, we also investigated one overwintering population. No formation of specialized resting stages (e.g., dormant zygospores) was observed. Markedly, Zygnema spp. survived harsh periods as modified vegetative cells, i.e., pre-akinetes. Pre-akinetes tolerate both desiccation during summer and freezing in winter. These cells are not dormant and therefore recover their physiological activity immediately after transfer to favorable conditions, undergoing rapid growth in the early spring. Nevertheless, once pre-akinetes begin to grow, these newly produced vegetative cells lose stress tolerance. Such rapid dehardening explains their high mortality due to frequent freeze–thaw cycles in the early spring. Arctic Zygnema spp. thus face a phenological trade-off between missing the early growing season and experiencing frost damage.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The lack of liquid water is the major stress factor affecting living organisms in polar regions. Frozen water is present most of the year, and during the short summers, the precipitation level is usually low (Robinson et al. 2003; Rautio et al. 2011). Nevertheless, many terrestrial habitats are supplied by meltwater from snow and glacier-fed streams or melting permafrost throughout nearly the entire vegetation season. Such environments, which are referred to as hydro-terrestrial, include areas such as wetlands, shallow lakes, pools, streams, seepages, and springs (Elster 2002). Hydro-terrestrial habitats are generally small, shallow, and unstable. These habitats can partially or fully dry out during the summer, undergo diurnal freeze–thaw cycles in the spring and autumn, and freeze solid in the winter. Therefore, any inhabitants of such habitats must be well adapted to these annual cycles of unfavorable conditions.
Due to specific climatic conditions including the lack of liquid water, the biodiversity and ground cover of vascular plants is low in high Arctic desert and/or semidesert environments (Thomas et al. 2008). On the other hand, these environments are usually rich in microbial life. In many extreme polar habitats, eukaryotic microalgae and cyanobacteria, together with lichens and mosses, represent the most important phototrophs (Aleksandrova 1988; Vincent 2000). Numerous reports show that eukaryotic microalgae are less stress-resistant than cyanobacteria (Davey 1988, 1989; Šabacká and Elster 2006) and that they undergo typical annual development, while cyanobacteria form rather perennial mats (Tang et al. 1997). Winter is regarded as the major stressful season, causing significant declines in algal populations (Hawes1988, 1990; Davey 1991a; Davey and Rothery 1992; Šabacká and Elster 2006). New biomass grows rapidly from small inoculum in the early spring, sometimes even under snow (Davey 1988).
Filamentous conjugating green algae (class Zygnematophyceae), typically represented by the genus Zygnema, comprise the most common eukaryotic components of Arctic hydro-terrestrial mats (Sheath et al. 1996; Kim et al. 2008; Holzinger et al. 2009; Pichrtová et al. 2014a). These algae are closely related to land plants (Embryophyta; Leliaert et al. 2012) and are characterized by a special type of sexual reproduction, the process of conjugation, which leads to the formation of highly specialized, stress-resistant zygospores containing algaenan (a sporopollenin-like substance) in their cell walls (Kadlubowska 1984; Poulíčková et al. 2007). Despite the abundance of polar Zygnema, the formation of zygospores in these algae has only very rarely been observed (Elster et al. 1997). In addition, other types of specialized cells were described in the genus Zygnema, for example akinetes—asexual rectangular cells with cell walls of the same structure as that of the zygospores (Kadlubowska 1984; Stancheva et al. 2012).
However, polar Zygnema can survive various stress conditions, e.g., osmotic stress (Pichrtová et al. 2014a), desiccation (Pichrtová et al. 2014b), and freezing (Hawes 1990), by forming modified mature vegetative cells that are formed directly from vegetative cells via the gradual accumulation of storage materials and cell wall thickening (McLean and Pessoney 1971; Herburger et al. 2015). Their chloroplasts have reduced lobes, and their physiological activity is diminished. Here, we call such cells pre-akinetes (Fuller 2013; Pichrtová et al. 2014b) to distinguish them from true akinetes with characteristic cell wall features. The formation of pre-akinetes was previously observed in the Arctic at the end of the vegetation season (Pichrtová et al. 2014a). Thus, pre-akinetes might play an important role in the survival of Zygnema in polar hydro-terrestrial environments, but to date, no comprehensive study has been conducted on the annual development of these algae. The aim of the current study was to characterize the physiological performance of Zygnema during its entire annual life cycle in hydro-terrestrial habitats of Svalbard (High Arctic), with a focus on the production of specialized cells (zygospores, akinetes), which are hypothesized to assure survival of these algae during unfavorable periods.
Materials and methods
Study site
Green algal mats growing in small pools in the western part of Petunia Bay (Billefjorden, central Svalbard, High Arctic) were selected for this study (Online Resource 1). These mats are dominated by conjugatophytes of the genus Zygnema and occur in close proximity to the Petuniahytta field research station (Elster and Rachlewicz 2012). All investigated pools occupy an area of up to 5 m2 and are very shallow (<5 cm). Therefore, the water temperature can reach as high as 16 °C during sunny summer days. The main sources of water in the summer are melting permafrost and streamlets that bring meltwater from snowfields. The water chemistry of the shallow pools is influenced by limestone inclusions in their catchments and sometimes by seawater. Therefore, these pools have neutral to slightly alkaline pH levels of between 7.2 and 8.4, and conductivity between 502 and 1584 µS cm−1 (Komárek et al. 2012). The vegetation season lasts from June to September. The monthly mean air temperature in the area ranges from –17 to 7.2 °C, and during winter, the temperature can occasionally fall below –30 °C or exceed 0 °C (Láska et al. 2012).
Field sampling of natural populations
The seasonal development of algal mats was investigated in the summer seasons of 2010 and 2011. Altogether, four populations forming hydro-terrestrial mats were selected for investigation. Twenty permanent spots that were marked per population served as replicates. The morphology of the cells was studied under an Olympus BX53 light microscope during early (June 28–July 12) and late (August 26–September 8) summer.
In addition, a winter sampling of one of the populations (referred to as population 2) was performed in April 2012 when the biomass was still frozen solid in ice and covered by 30 cm of ice-snow. Population 2 formed the most extensive mats during the summer which made it easy to be localized even under snow and ice cover.
Laboratory experiments with population 2 collected in winter
The frozen winter samples were transported to the university laboratory in Prague for further investigation. After melting, one portion of the biomass was transferred into fresh liquid BBM medium (Bischoff and Bold 1963) and stored at 5 or 20 °C at a PAR (photosynthetic active radiation) irradiance of 39 µmol photons m−2 s−1.
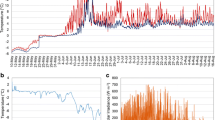
The other portion of the frozen biomass was melted, placed onto agar plates, and cultivated in crossed gradients of light and temperature (Labio, Czech Republic; Fig. 1; Kvíderová and Lukavský 2001) for 1 week. This method enables various parameters (growth rate, pigment content, and chlorophyll fluorescence parameters) to be monitored simultaneously in factorial combinations of light and temperature. Twenty-five positions were defined: five levels of temperature (T1–T5; mean values 7.1–25.0 °C) and five levels of irradiance (I1–I5; mean values 26–368 µmol photons m−2 s−1; Fig. 1).

Temperature (a) and irradiance (b) measured at different positions of the cultivation unit for a crossed gradient of light and temperature
At the end of the cultivation period, biomass grown under a crossed gradient of light and temperature was desiccated together with freshly melted material. The algal filaments were evenly spread on glass fiber filters (Whatman GF/C) without additional moistening and transferred to a closed glass chamber over a saturated sodium chloride solution (relative air humidity of 75 %). After 24 h, the samples were transferred to liquid BBM medium, and after additional 48 h, the percentage of living cells was estimated using light microscopy observation.
Chlorophyll a fluorescence
Chlorophyll a fluorescence was measured using an imaging modulated fluorometer FluorCam (PSI, Czech Republic). The field-collected samples were transferred to the research station and dark-acclimated for 30 min prior to measurement of F V/F M, the maximum quantum yield of photosystem II (PSII). The samples were then kept in darkness for an additional 8 h, followed by a second measurement of F V/F M to assess the recovery rate after possible photoinhibitory stress under field conditions.
Chlorophyll a fluorescence was also measured on the melted winter material during 80-h recovery in fresh medium and cultivation in crossed gradients of light and temperature. Two parameters were selected, F V/F M and steady-state quantum yield of PSII in the light (Φ PSII; measured at the actinic light of 100 µmol photons m−2 s−1). Briefly, F V/F M characterizes the overall physiological state of PSII, whereas Φ PSII serves as a good estimation of photosynthetic activity (Maxwell and Johnson 2000).
Pigment content
After cultivation, the biomass from nine representative positions was harvested, filtered on glass fiber filters, and extracted in 80 % acetone (combination of positions T1, T3, and T5, and I1, I3, and I5; Fig. 1). The contents of chlorophylls a and b and total carotenoids were estimated spectrophotometrically according to the equations provided by (Lichtenthaler and Buschmann 2001).
Statistical analyses
The seasonal changes in F V/F M were tested by general linear model (GLM) repeated-measures analysis of variance (ANOVA) with nested design. The time of dark acclimation was set as a repeated factor. The other factors tested, “population,” “day,” and “season,” were regarded as fixed factors, with “day” nested in “season.” “Season” had two levels: early and late summer. For each population, 20 independent replicate measurements were taken.
The recovery of F V/F M and Φ PSII values, measured after melting on eight replicate samples, was described by the asymptotic function of hyperbolic growth: P = P max × slope × time/(P max + slope × time) + P min, where P is the chlorophyll fluorescence parameter, P max is its maximum increase, P min is its minimum value (y-intercept), and slope is its initial (maximum) recovery rate in time (maximum slope of the curve). The three parameters of the model were estimated, and the differences between both cultivation temperatures were statistically tested using GraphPad Prism 6 software (GraphPad Software, USA).
Data obtained from the one-week cultivation under a crossed gradient of light and temperature (dry weight and pigment content) were not analyzed statistically, because the very slow growth in the lowest temperature did not provide enough biomass for replicate samples.
Results
Seasonal changes in morphology
In the early summer (late June), the investigated sites were full of dead Zygnema biomass from the previous year. Nevertheless, new biomass was produced rapidly, and within a few weeks, the pools were filled with new Zygnema mats. Light microscopy revealed that the cells had typical vegetative morphology, with two clear, stellate chloroplasts and large, hyaline vacuoles (Fig. 2a).

Typical morphology of field-collected Zygnema sp. (population 2) over the course of the season: a vegetative cells with stellate chloroplasts (July 6), b pre-akinetes (August 24), and c overwintering pre-akinetes immediately after melting (April 22). Scale bars 20 µm
In the late summer (August), the growth ceased and the cells began to accumulate storage materials. Their morphology gradually changed, becoming mature, stationary-phase-like cells usually referred to as pre-akinetes (Fig. 2b). The filaments also tended to break into short fragments or even single cells.
In April, we collected frozen biomass from under the ice-snow. The biomass was frozen solid in ice but still appeared bright green. After melting, the biomass consisted of viable pre-akinetes that appeared similar to those observed in late summer (Fig. 2c).
We never observed conjugation or zygospore formation at these particular sites. True akinetes with zygospore-like cell walls were also not produced by the investigated populations.
Seasonal changes in chlorophyll fluorescence
A significant rise in F V/F M values was detected between 30 min and 8 h of dark acclimation in all cases (p < 0.0001), indicating that 30 min was not sufficient for the relaxation of PSII ( Fig. 3). However, the significance of the interaction between the dark acclimation time and the season (p < 0.0001) indicates that the dynamics of F V/F M differed between both parts of the summer season, and that the increase in F V/F M after 8 h in the dark was much more pronounced in the early summer.

Maximum quantum yield of photosystem II (F V/F M) measured in four natural Zygnema sp. populations. Measurements were taken after 30 min and 8 h of dark acclimation. Each value represents the mean of 20 sites on 3 (a) and 4 (b) different days. Weighted mean ± standard error, n = 60 (a) and 80 (b)
Notably, population 2 exhibited different seasonal development of F V/F M compared to the three other populations (Fig. 3). The values after 30 min and 8 h of dark acclimation were both significantly lower in late summer than in early summer in populations 1 (p < 0.0001), 3 (p < 0.0001), and 4 (p = 0.0001, respectively, p < 0.0001). On the other hand, the 8-h dark values remained the same throughout the summer (p = 1.00) in population 2, and after only 30 min of darkness, the F V/F M was even higher at the end of the season (p < 0.0001). A complete list of the results of the GLM ANOVA and Tukey’s post hoc tests can be found in Online Resource 2.
Viability and stress tolerance of frozen cells from population 2
Photosynthetic activity began to recover immediately after melting and transfer to fresh liquid BBM medium. At 24 h following rehydration, cell division was already observed. The biomass growth was evident at both temperatures (5 and 20 °C). However, at 20 °C, the recovery rate of both F V/F M and Φ PSII (slopes of the hyperbolae; p = 0.0004, respectively, 0.0061) as well as their estimated maxima (asymptotes of the hyperbolae; p = 0.0001, respectively, 0.042) were higher (Fig. 4; Online Resource 2). There were no significant differences in the starting minimum values of the parameters. Fitted parameters of the hyperbolae and complete results of the tests can be found in Online Resource 2.

Recovery of parameters of chlorophyll fluorescence after melting during cultivation at 5 and 20 °C: a maximum quantum yield of photosystem II, and b steady-state quantum yield of PSII in the light; n = 8, mean ± SD. The parameters were fitted by hyperbolic growth function; the initial slope of the curve denotes the maximum recovery rate of the parameters. The fitted curves are shown as dashed (5 °C) and solid (20 °C) lines, respectively
After 1 week of cultivation, biomass growth and cell division were apparent at all positions of the crossed gradients of light and temperature. At all combinations of conditions, the pre-akinetes transformed back into vegetative cells. The highest values of the fluorescence parameters were reached under various combinations of low irradiance and high temperature (Fig. 5).

Fluorescence parameters measured on biomass cultivated for 1 week at different positions of the crossed gradient of light and temperature. Average values of temperature and irradiance were as follows: T1 = 7.1, T2 = 12.8, T3 = 15.6, T4 = 20.5, and T5 = 25.0 °C; and I1 = 26, I2 = 50, I3 = 89, I4 = 181, and I5 = 368 µmol photons m−2 s−1; n = 4, mean + SD
Accordingly, the highest growth rate (estimated as an increase in dry weight) was also detected at the highest temperature tested combined with intermediate irradiance (Fig. 6a). In general, the relative pigment content also increased with increasing temperature and decreasing irradiance (Fig. 6b–e). Increases in irradiance led to lower chlorophyll-to-carotenoid ratios as well (Fig. 6f).

Dry weight and pigment content of biomass grown for 1 week under a crossed gradient of light and temperature. For T1, n = 1, and for T3 and T5, n = 4, mean + SD
The chlorophyll fluorescence parameters decreased to zero within 30 min after the transfer into 75 % relative air humidity, indicating that physiological activity ceased due to the loss of intracellular water. Notably, no single cell from liquid culture survived 24 h of desiccation, whereas in the freshly melted material consisting of pre-akinetes, approximately 5 % of the cells survived these conditions.
Discussion
In this study, we showed that Arctic Zygnema spp. are able to survive throughout an entire annual cycle in a vegetative state without forming stress-resistant zygospores or akinetes. Vegetative cells gradually accumulate storage compounds and reduce their physiological activity during the vegetation season. Such modified vegetative cells (referred to as pre-akinetes) that were naturally hardened by slow desiccation were tolerant to osmotic stress (Pichrtová et al. 2014a) and desiccation (Pichrtová et al. 2014b), and were also able to survive the Arctic winter (this study). These cells are not dormant and recover quickly under favorable conditions, which is important for the rapid biomass growth that occurs soon after snowmelt.
The morphological characteristics of pre-akinetes differ from those of young vegetative cells in several ways. Pre-akinetes contain granules of storage substances and are slightly brownish. Ultrastructural investigations confirmed that pre-akinetes accumulate high levels of lipids and have reduced chloroplast lobes (McLean and Pessoney 1971; Holzinger et al. 2009; Pichrtová et al. 2014b; Herburger et al. 2015). Pre-akinete formation was also accompanied by a significant change in fatty acid composition (Pichrtová et al. 2016). Pre-akinetes also possess thicker cell walls than young vegetative cells but lack the three-layered walls of zygospores containing algaenan (Fuller 2013).
Pre-akinetes formed gradually from young vegetative cells during late summer. Their formation was not induced by desiccation under natural (Pichrtová et al. 2014a) or experimental (Pichrtová et al. 2014b) conditions. However, these cells formed more readily under nitrogen-limited culture conditions, indicating the important effect of starvation (Pichrtová et al. 2014b). We thus hypothesize that nitrogen (or other nutrients) is depleted during extensive growth in polar seepage pools, which in turn causes the cessation of growth and pre-akinete formation. In general, nutrient cycling is relatively slow in polar habitats (Robinson et al. 2003), and nutrient availability is usually a limiting factor (Davey and Rothery 1992; Arnold et al. 2003; Hogan et al. 2014), although exceptions exist, such as near bird colonies (Zwolicki et al. 2013).
In addition, other physicochemical environmental factors that change during the season (e.g., temperature, light intensity/spectral composition, and electric conductivity) can also lead to the accumulation of storage products and pre-akinete formation. For example, the lipid content in the eustigmatophyte Nannochloropsis oculata increased in autumn, even with constant nutrient availability (Olofsson et al. 2012). The cessation of growth in dense populations has also been observed in microalgal mass cultures. In addition to nutrient depletion, this observation may reflect the reduced photosynthesis caused by self-shading (Richmond and Hu 2013) or the lack of CO2; the biomass itself also produces and accumulates growth inhibitors (Park et al. 2014).
Recent studies on polar Zygnema spp. also showed that pre-akinetes become more stress-resistant after hardening, e.g., by slow desiccation (Pichrtová et al. 2014a, b). The physiological effects of freezing and desiccation are similar because they both lead to lower intracellular water potential (Bisson and Kirst 1995). Thus, acclimation during exposure to desiccation stress at the end of the summer is expected to result in resistance to freezing. Conversely, we showed that naturally frozen pre-akinetes were able to survive desiccation immediately after melting.
The biochemical nature of such hardening is currently under investigation. Green microalgae accumulate various substances with protective functions against desiccation and freezing stress (Holzinger and Karsten 2013). For example, the Antarctic trebouxiophyte green alga Prasiola crispa accumulates proline before winter (Jackson and Seppelt 1995). Similarly, the intracellular concentrations of glycerol and erythritol in Antarctic soil microalgae increase at the end of summer (Arnold et al. 2003). Moreover, changes in cell wall structure and composition accompany pre-akinete formation. The secondary pectin layer of the cell wall allows water to be retained due to its physicochemical properties (Fuller 2013).
Not only do pre-akinetes differ from young vegetative cells in terms of morphology and ultrastructure, but (above all) they also differ in terms of physiological characteristics and stress tolerance. The sustained decline in F V/F M indicates either photoinhibitory damage to PSII or downregulation at PSII due to sink limitation, i.e., reduced requirements for CO2 fixation during, e.g., winter dormancy or nutrient starvation (Adams and Demmig-Adams 2004). The latter interpretation of sink limitation fits our observation that old, stationary-phase cells (pre-akinetes) are usually physiologically less active than young vegetative cells (Coleman 1983; Herburger et al. 2015). Indeed, in the majority of populations, the F V/F M values were significantly lower in late summer than in early summer. However, population 2 showed no clear trend throughout the summer season, suggesting that pre-akinetes in this population were exceptionally well adapted to utilizing light throughout the growing season, in contrast to the others whose pre-akinetes lost their PSII potential in late summer. The same population was included in a previous study by Pichrtová et al. (2014a; also referred to as “population 2”); although it formed pre-akinetes like other populations, its F V/F M values were also markedly higher than those of the other populations investigated. This difference in populations is likely associated with genetic variability rather than acclimation to local conditions (Pichrtová et al. 2014a).
The significant increase in F V/F M after 8 h of dark acclimation indicates strong photoinhibition under natural conditions, particularly in young vegetative cells (i.e., in early summer). The populations are exposed to continuous daylight throughout the summer without the possibility of full dark recovery of the photosynthetic apparatus. The strong photoinhibition may be thus understood as a constitutive photoprotective mechanism that prevents excess chlorophyll excitation at PSII, and thus reduces oxidative stress and uncontrolled photodamage (Adams and Demmig-Adams 2004). Another excitation-reducing mechanism involves self-screening of the cells by aggregating filaments into multilayered structures. Mat-forming growth is a common strategy used by microorganisms for protection against multiple stresses (Bischof et al. 2002; De los Ríos et al. 2004; Knowles and Castenholz 2008; Holzinger and Pichrtová 2016). In addition, Zygnema produce a wide range of phenolic substances that provide screening not only in the UV range, but (partially) also in the PAR region, which can provide an additional layer of protection against excessive irradiation (Pichrtová et al. 2013).
Accordingly, when the algae were cultivated in crossed gradients of light and temperature, the highest values of both fluorescence parameters and the highest biomass production occurred under low irradiance. Low-light adaptation and photoinhibition at higher irradiance were previously reported in Arctic and Antarctic Zygnema (Kaplan et al. 2013). Moreover, an Antarctic Zygnema sp. was able to grow under very low PAR flux (as low as 1 µmol photons m−2 s−1; Davey 1991b). Low-light adaptation is a typical feature of soil and biofilm microalgae that grow under soil particles or under upper biomass layers (Davey 1991b; Gray et al. 2007; Karsten et al. 2013).
In contrast to low irradiance levels, optimal Zygnema growth occurred at the highest temperature tested (ca. 25 °C), and recovery was faster at high cultivation temperatures. Similarly, the maximum growth rate of an Antarctic Zygnema sp. was achieved at 20 °C (Davey 1991b). A similar phenomenon was previously observed in other Arctic and Antarctic microalgae, which were shown to be psychrotrophs rather than psychrophiles (Seaburg et al. 1981; Davey 1991b). A broad temperature range for growth (20–30 °C units) is important under the highly unstable conditions of polar wetlands (Seaburg et al. 1981).
Markedly, conjugation and zygospore formation was observed only very rarely in the Arctic (Elster et al. 1997). This is in contrast to the findings of Genkel and Pronina (1979) who investigated the populations of Zygnema stellinum in the temperate climate of Belarus and concluded that pre-akinetes are able to survive short-term stress events during the summer, but the formation of highly resistant cells (zygospores and parthenospores) is necessary for survival in the winter, the major annual stress event. Nevertheless, Arctic and Antarctic microalgae have repeatedly been shown to survive as vegetative cells with thick cell walls and accumulated reserves (Hawes 1990; Sheath et al. 1996).
The formation of pre-akinetes is advantageous because these cells are not dormant but are instead physiologically active immediately after melting. Similarly, Antarctic vegetative Prasiola also recovered photosynthetic activity immediately after collection during winter after being frozen for 3 months (Jackson and Seppelt 1995). Additionally, the above-mentioned adaptation of pre-akinetes to low-light levels enables them to grow even under snow cover. Rapid growth in early spring is an important competitive advantage, as these algae must compete with cyanobacteria and other microalgae in unstable habitats, with only a few weeks of favorable conditions (the presence of liquid water) per year (Davey 1988, 1991b). The survival of only modified vegetative cells may also be due to the low grazing pressure in the Arctic habitats. Potential grazers of microalgae in the Arctic include insects such as chironomids, caddisflies, and mayflies (Sheath et al. 1996), which are low in abundance compared to grazers in temperate regions. Finally, filaments consisting of pre-akinetes tend to break into single cells (Pichrtová et al. 2014a) and can thus serve as airborne propagules.
Even though frozen filaments consist of viable pre-akinetes during the winter, the spring always brings high mortality to the biomass of the previous year; the reason for this remains unknown. A similar phenomenon was observed for other Arctic and Antarctic eukaryotic algae (Davey 1991a; Elster 2002). Here, we showed that immediately after transfer to fresh liquid medium, the pre-akinetes began to grow, but the dehardened young cells lost their stress tolerance. Therefore, we propose that the early-spring diurnal, short-term cycles of freeze–thaw and/or desiccation–rehydration (Láska et al. 2012) represent a bottleneck in the life cycle of Arctic populations of Zygnema. For the population to survive, it is crucial that at least a few cells retain their resistance (Hawes 1988, 1990; Tashyreva and Elster 2012).
We conclude that naturally hardened pre-akinetes play a key role in the survival of Zygnema spp. in the polar hydro-terrestrial environment. However, populations of these species exhibit a phenological trade-off between missing the early growing season and experiencing frost damage. Also, the scarcity of sexual reproduction indicates that Arctic Zygnema populations live at their physiological limits.
References
Adams WW III, Demmig-Adams B (2004) Chlorophyll fluorescence as a tool to monitor plant response to the environment. In: Papageorgiou GC (ed) Chlorophyll a fluorescence: a signature of photosynthesis. Advances in photosynthesis and respiration, vol 19. Springer, Berlin, pp 583–604
Aleksandrova VD (1988) The Arctic and Antarctic: their division into geobotanical areas. Cambridge University Press, Cambridge
Arnold RJ, Convey P, Hughes KA, Wynn-Williams DD (2003) Seasonal periodicity of physical factors, inorganic nutrients and microalgae in Antarctic fellfields. Polar Biol 26:396–403
Bischof K, Peralta G, Kräbs G, van de Poll WH, Pérez-Lloréns JL, Breeman AM (2002) Effects of solar UV-B radiation on canopy structure of Ulva communities from southern Spain. J Exp Bot 53:2411–2421
Bischoff HW, Bold HC (1963) Phycological studies IV. Univ Texas Publ No, Some soil algae from enchanted rock and related algal species 6318
Bisson MA, Kirst GO (1995) Osmotic acclimation and turgor pressure regulation in algae. Naturwissenschaften 82:461–471
Coleman AW (1983) The roles of resting spores and akinetes in chlorophyte survival. In: Fryxell GA (ed) Survival strategies of the algae. Cambridge University Press, Cambridge, pp 1–21
Davey MC (1988) Ecology of terrestrial algae of the fellfield ecosystems of Signy Island, South Orkney Islands. Brit Antarct Surv B 81:69–74
Davey MC (1989) The effects of freezing and desiccation on photosynthesis and survival of terrestrial Antarctic algae and Cyanobacteria. Polar Biol 10:29–36
Davey MC (1991a) The seasonal periodicity of algae on Antarctic fellfield soils. Holarctic Ecol 14:112–120
Davey MC (1991b) Effects of physical factors on the survival and growth of Antarctic terrestrial algae. Brit Phycol J 26:315–325
Davey MC, Rothery P (1992) Factors causing the limitation of growth of terrestrial algae in maritime Antarctica during late summer. Polar Biol 12:595–601
De los Ríos A, Wierzchos J, Sancho LG, Ascaso C (2004) Exploring the physiological state of continental Antarctic endolithic microorganisms by microscopy. FEMS Microbiol Ecol 50:143–152
Elster J (2002) Ecological classification of terrestrial algal communities in polar environments. In: Beyer L, Bölter M (eds) Geoecology of Antarctic ice-free coastal landscapes, ecological studies, vol 154. Springer, Berlin, pp 303–326
Elster J, Rachlewicz G (2012) Petuniabukta, Billefjorden in Svalbard: Czech-Polish long term ecological and geographical research. Pol Polar Res 33:289–295
Elster J, Svoboda J, Komárek J, Marvan P (1997) Algal and cyanoprocaryote communities in a glacial stream, Sverdrup Pass, 79°N, Central Ellesmere Island, Canada. Algol Stud 85:57–93
Fuller C (2013) Examining morphological and physiological changes in Zygnema irregulare during a desiccation and recovery period. M. Sc. thesis, California State University San Marcos
Genkel PA, Pronina ND (1979) Ecology of Zygnema stellinum Vauch. during desiccation of a shallow body of water. Biol Bull Acad Sci USSR 6:504–509
Gray DW, Lewis LA, Cardon ZG (2007) Photosynthetic recovery following desiccation of desert green algae (Chlorophyta) and their aquatic relatives. Plant, Cell Environ 30:1240–1255
Hawes I (1988) The seasonal dynamics of Spirogyra in a shallow maritime antarctic lake. Polar Biol 8:429–437
Hawes I (1990) Effects of freezing and thawing on a species of Zygnema (Chlorophyta) from the Antarctic. Phycologia 29:326–331
Herburger K, Lewis LA, Holzinger A (2015) Photosynthetic efficiency, desiccation tolerance and ultrastructure in two phylogenetically distinct strains of alpine Zygnema sp. (Zygnematophyceae, Streptophyta): role of pre-akinete formation. Protoplasma 252:571–589
Hogan EJ, McGowan S, Anderson NJ (2014) Nutrient limitation of periphyton growth in arctic lakes in south-west Greenland. Polar Biol 37:1331–1342
Holzinger A, Karsten U (2013) Desiccation stress and tolerance in green algae: consequences for ultrastructure, physiological and molecular mechanisms. Front Plant Sci 4:327
Holzinger A, Pichrtová M (2016) Abiotic stress tolerance in charophyte green algae: new challenges for omics techniques. Front Plant Sci 7:678
Holzinger A, Roleda MY, Lütz C (2009) The vegetative arctic freshwater green alga Zygnema is insensitive to experimental UV exposure. Micron 40:831–838
Jackson AE, Seppelt RD (1995) The accumulation of proline in Prasiola crispa during winter in Antarctica. Physiol Plantarum 94:25–30
Kadlubowska JZ (1984) Conjugatophyceae I: Chlorophyta VIII: Zygnemales. In: Ettl H, Gerloff J, Heynig H, Mollenhauer D (eds) Süsswasserflora von Mitteleuropa, vol 16. Gustav Fisher, Jena
Kaplan F, Lewis LA, Herburger K, Holzinger A (2013) Osmotic stress in Arctic and Antarctic strains of the green alga Zygnema (Zygnematales, Streptophyta): effects on photosynthesis and ultrastructure. Micron 44:317–330
Karsten U, Pröschold T, Mikhailyuk T, Holzinger A (2013) Photosynthetic performance of different genotypes of the green alga Klebsormidium sp. (Streptophyta) isolated from biological soil crusts of the Alps. Algol Stud 142:45–62
Kim GH, Klochkova TA, Kang SH (2008) Notes on freshwater and terrestrial algae from Ny-Ålesund, Svalbard (high Arctic sea area). J Environ Biol 29:485–491
Knowles EJ, Castenholz RW (2008) Effect of exogenous extracellular polysaccharides on the desiccation and freezing tolerance of rock-inhabiting phototrophic microorganisms. FEMS Microbiol Ecol 66:261–270
Komárek J, Kováčik L, Elster J, Komárek O (2012) Cyanobacterial diversity of Petunia-Bukta, Billefjorden, central Svalbard. Pol Polar Res 33:347–368
Kvíderová J, Lukavský J (2001) A new unit for crossed gradients of temperature and light. Nova Hedwig Beih 123:541–550
Láska K, Witoszová D, Prošek P (2012) Weather patterns of the coastal zone of Petuniabukta, central Spitsbergen in the period 2008–2010. Pol Polar Res 33:297–318
Leliaert F, Smith DR, Moreau H, Herron MD, Verbruggen H, Delwiche CF, De Clerck O (2012) Phylogeny and molecular evolution of the green algae. Crit Rev Plant Sci 31:1–46
Lichtenthaler HK, Buschmann C (2001) Chlorophylls and carotenoids: measurement and characterization by UV-VIS spectroscopy. In: Wrolstad RE, Acree TE, An H, Decker EA, Penner MH, Reid DS, Schwartz SJ, Shoemaker CF, Sporns P (eds) Current protocols in food analytical chemistry (CPFA). Wiley, New York, pp F4.3.1–F4.3.8
Maxwell K, Johnson GN (2000) Chlorophyll fluorescence—a practical guide. J Exp Bot 51:659–668
McLean RJ, Pessoney GF (1971) Formation and resistance of akinetes of Zygnema. In: Parker BC, Brown RM Jr (eds) Contributions in phycology. Allen, Lawrence, pp 145–152
Olofsson M, Lamela T, Nilsson E, Bergé JP, del Pino V, Uronen P, Legrand C (2012) Seasonal variation of lipids and fatty acids of the microalgae Nannochloropsis oculata grown in outdoor large-scale photobioreactors. Energies 5:1577–1592
Park JC, Choi SP, Hong M-E, Sim SJ (2014) Enhanced astaxanthin production from microalga, Haematococcus pluvialis by two-stage perfusion culture with stepwise light irradiation. Bioproc Biosyst Eng 37:2039–2047
Pichrtová M, Remias D, Lewis LA, Holzinger A (2013) Changes in phenolic compounds and cellular ultrastructure of arctic and antarctic strains of Zygnema (Zygnematophyceae, Streptophyta) after exposure to experimentally enhanced UV to PAR ratio. Microb Ecol 65:68–83
Pichrtová M, Hájek T, Elster J (2014a) Osmotic stress and recovery in field populations of Zygnema sp. (Zygnematophyceae, Streptophyta) on Svalbard (High Arctic) subjected to natural desiccation. FEMS Microbiol Ecol 89:270–280
Pichrtová M, Kulichová J, Holzinger A (2014b) Nitrogen limitation and slow drying induce desiccation tolerance in conjugating green algae (Zygnematophyceae, Streptophyta) from polar habitats. PLoS ONE 9:e113137
Pichrtová M, Arc E, Stöggl W, Kranner I, Hájek T, Hackl H, Holzinger A (2016) Formation of lipid bodies and changes in fatty acid composition upon pre-akinete formation in arctic and Antarctic Zygnema (Zygnematophyceae, Streptophyta) strains. FEMS Microbiol Ecol 92:fiw096
Poulíčková A, Žižka Z, Hašler P, Benada O (2007) Zygnematalean zygospores: morphological features and use in species identification. Folia Microbiol 52:135–145
Rautio M, Dufresne F, Laurion I, Bonilla S, Vincent WF, Christoffersen KS (2011) Shallow freshwater ecosystems of the circumpolar Arctic. Ecoscience 18:204–222
Richmond A, Hu Q (2013) Handbook of microalgal culture: applied phycology and biotechnology, 2nd edn. Wiley-Blackwell, London
Robinson SA, Wasley J, Tobin AK (2003) Living on the edge—plants and global change in continental and maritime Antarctica. Global Change Biol 9:1681–1717
Šabacká M, Elster J (2006) Response of Cyanobacteria and algae from Antarctic wetland habitats to freezing and desiccation stress. Polar Biol 30:31–37
Seaburg KG, Parker BC, Wharton RA Jr, Simmons GM Jr (1981) Temperature-growth responses of algal isolates from Antarctic oases. J Phycol 17:353–360
Sheath RG, Vis ML, Hambrook JA, Cole KM (1996) Tundra stream macroalgae of North America: composition, distribution and physiological adaptations. Hydrobiologia 336:67–82
Stancheva R, Sheath RG, Hall JD (2012) Systematics of the genus Zygnema (Zygnematophyceae, Charophyta) from Californian watersheds. J Phycol 48:409–422
Tang EPY, Tremblay R, Vincent WF (1997) Cyanobacterial dominance of polar freshwater ecosystems: are high-latitude mat-formers adapted to low temperature? J Phycol 33:171–181
Tashyreva D, Elster J (2012) Production of dormant stages and stress resistance of polar cyanobacteria. In: Hanslmaier A, Kempe S, Seckbach J (eds) Life on Earth and other planetary bodies. Springer, Dordrecht, pp 367–386
Thomas DN, Fogg GE, Convey P, Fritsen CH, Gili J-M, Gradinger R, Laybourn-Parry J, Reid K, Walton DWH (2008) The biology of polar regions. Oxford University Press, Oxford
Vincent WF (2000) Cyanobacterial dominance in the polar regions. In: Whitton BA, Potts M (eds) The ecology of cyanobacteria—their diversity in time and space. Kluwer, Dordrecht, pp 321–340
Zwolicki A, Zmudczynska-Skarbek KM, Iliszko L, Stempniewicz L (2013) Guano deposition and nutrient enrichment in the vicinity of planktivorous and piscivorous seabird colonies in Spitsbergen. Polar Biol 36:363–372
Acknowledgments
We thank to Kamil Láska for providing map background for a figure showing site location. We also wish to thank Daria Tashyreva for her assistance during field expeditions. This work was supported by the Grant No. 15-34645L of the Czech Science Foundation (GAČR), by projects from the Czech Ministry of Education, Youth and Sports (KONTAKT ME 934, INGO LA 341 and LM2010009 CzechPolar), by Project Reg. No. CZ.1.07/2.2.00/28.0190 funded by the European Social Fund and from the government budget of the Czech Republic, and as a long-term research development project of the Czech Academy of Sciences RVO 67985939.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Online resource 1 (ESM1)
Location of the investigated Zygnema spp. populations 1–4 in Petuniabukta, Svalbard (TIFF 2709 kb)
Online resource 2 (ESM2)
Complete results of the statistical analyses used in the study: general linear model repeated-measures analysis of variance, Tukey’s post hoc test, and comparison of hyperbolae (PDF 206 kb)
Rights and permissions
About this article

Cite this article
Pichrtová, M., Hájek, T. & Elster, J. Annual development of mat-forming conjugating green algae Zygnema spp. in hydro-terrestrial habitats in the Arctic. Polar Biol 39, 1653–1662 (2016). https://doi.org/10.1007/s00300-016-1889-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-016-1889-y