
Abstract
The biosynthesis of integric acid, a secondary metabolite of the wood-decay fungus Xylaria feejeensis strain 2FB-PPM08M, has been studied. Labeling experiments using [1-13C], [2-13C] and [1,2-13C2] acetate and l-methionine (methyl-13C) were separately performed with fungal culture. The labeling patterns of these metabolites indicated the same origin, and determined that integric acid was formed through the condensation of a sesquiterpene and a polyketide. These experiments showed that side chain of compounds would be synthesized by the polyketide pathway, while the ring carbon indicated the biosynthesis of compounds via the mevalonate pathway.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Integric acid was isolated from the culture of Xylaria sp. Integric acid, an acylated eremophilane sesquiterpenoid, is defined by two distinct structural units: a bicyclic terpenoid unit consisting of a free carboxylic acid, a vinyl aldehyde, and a conjugated ketone; and a 2,4-dimethyloctanoic acyl chain. Selective chemical and enzymatic modifications of the functional groups of integric acid were carried out. This compound and its derivatives have reportedly shown HIV-1 integrase inhibitory activity [1, 2]. Furthermore, it also has been displayed antimalarial activity against Plasmodium falciparum [3]. We recently isolated integric acid from the wood-decay fungus X. feejeensis 2FB-PPM08M, and became interested in establishing the biosynthetic pathway. The biosynthesis of integric acid isolated from this fungus has not yet been investigated. In particular, we aimed to verify that integric acid was formed through the condensation of a sesquiterpene and a polyketide from 13C-labeled acetate and methyl. To the best of our knowledge, this is the first report on the biosynthesis of integric acid.
Materials and Methods
Fungal Materials
The 2FB-PPM08M fungal strain was collected from the forest surrounding the Phu Pha Man National Park in Khon Kaen province, Thailand, and classified as Xylaria sp. based on its morphological features [4]. The Xylaria isolate was identified as X. feejeensis based on its nuclear ribosomal RNA sequences (ITS). Sequence analysis of strain 2FB-PPM08M revealed a 99 % sequence similarity to X. feejeensis (GU322454.1) from GenBank (data not shown), and therefore was further identified as X. feejeensis. The partial sequence of the ITS1–ITS4 region of Xylaria sp. 2FB-PPM08M was deposited in the DNA Data Bank of Japan (DDBJ accession number AB809464). A voucher specimen was deposited in the National Center for Genetic Engineering and Biotechnology (BIOTEC) Culture Collection in Thailand (voucher no. BCC63553).
Fermentation and Isolation
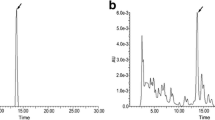
Xylaria feejeensis 2FB-PPM08M was grown at 30 °C for 21 days under static conditions into two 1,000-mL Erlenmeyer flasks, each containing 500 mL of potato dextrose broth (PDB; potato 200 g and dextrose 20 g/L, with 10 potato pieces for each flask). Metabolites were extracted from mycelial culture, after cultured mycelia were dried at 50 °C for 5 days and ground to a powder. Dried mycelial powder was extracted with ethyl acetate (EtOAc) with shaking for 24 h at 25 °C. Mycelia were removed by filtration, and the solvent was evaporated to dryness to give a residue (0.45 g). This residue was subjected to silica gel column chromatography (<0.063 mm mesh size; Merck) and eluted successively with 250 mL of EtOAc:n-hexane (50:50, v/v) and 250 mL of methanol (MeOH) to yield four fractions (F1–F4). The fourth fraction (F4: 0.38 g) had the most activity and was purified by HPLC. A pure compound was identified from the HPLC chromatogram of the residue by evaluating the retention times (t R) and UV spectra (acquired with a diode array detector). The HPLC system included a Waters separations module and UV/Vis detector, and the analytical column used was a Mightysil Si60 (250 × 3.0 mm; Kanto, Taiwan). The mobile phase consisted of 2-propanol:n-hexane (10:90, v/v) and 0.1 % (v/v) acetic acid. The flow rate was set at 1 mL/min at a temperature of 40 °C, and the detector was set at 254 nm. Under these conditions, the compound of interest had a t R of 6.3 min.
General Experimental Procedures
FTIR spectra were recorded for a thin film using a Bruker Tensor 27 spectrophotometer (Bruker, Billerica MA). 1H and 13C NMR spectra were recorded in CDCl3 at 25 °C on a Bruker Avance II 600 MHz NMR spectrometer and referenced to δ H 7.24 and δ C 77.0, respectively. Mass spectrometry (MS) was performed by positive- and negative-mode electrospray ionization on a Waters LCT Premier mass spectrometer (Waters, Milford MA). For high-precision measurements, the spectra were obtained by voltage scanning over a narrow mass range at 10,000 resolution. High-performance liquid chromatography (HPLC) system was employed, consisting of a Waters e2695 separations module and 2489 UV/Visible detector. Optical rotation was measured with a Horiba SEPA-500 polarimeter (Horiba, Kyoto, Japan). All labeled precursors were purchased from Sigma-Aldrich (St. Louis MO).
Integric Acid
Integric acid was isolated as a colorless powder, [α] 24D +38.0° (c 0.40, MeOH):1H NMR (600 MHz, CDCl3) δ H 9.53, 6.54, 6.35, 6.24, 6.09, 1.83, 1.51, 0.99 and 0.86; 13C NMR (77 MHz, CDCl3) δ C196.9, 193.2, 178.1, 166.8, 159.0, 149.7, 147.6, 136.5, 129.6, 125.8, 72.6, 53.3, 43.1, 43.0, 38.2, 36.5, 33.3, 29.7, 29.6, 22.7, 20.0, 19.9, 19.5, 14.0 and 12.6; IR 3040, 1720, 1719, 1688 and 1645 cm−1;HRMS m/z ([M-H]−): calcd. for C25H34O6, 429.2272; found 429.2277. The spectral data for this compound closely resembled those for the compound reported by Singh et al. [1] and were determined by comparing with reported data [5] as shown in (Fig. 1).

Structure of integric acid isolated from X. feejeensis 2FB-PPM08M
The Production of Integric Acid by X. feejeensis 2FB-PPM08M
To investigate for integric acid production profile, the X. feejeensis 2FB-PPM08M was cultured in PDB medium and incubated at 30 °C. The fungal mycelia were sampled after 3, 6, 9, 12, 15, 18, and 21 days of cultivation. After cultivation, mycelia were harvested, dried at 50 °C for 5 days, and ground, and dried mycelial powder was extracted with EtOAc and shaking for 24 h at 25 °C. Mycelia were removed by filtration, and the solvent was evaporated to produce a crude extract. One hundred microliters of the mycelia extracts were filtered and applied to HPLC for identification of changes in the integric acid product spectrum.
Incorporation of Labeled Acetates
A total volume of 1.5 L of PDB was distributed into fifteen 300-mL Erlenmeyer flasks, each containing 100 mL of PDB, in triplicate for each incorporation experiment. X. feejeensis 2FB-PPM08M was then inoculated in each flask. The fungus was grown without shaking at 30 °C for 21 days. Acetic acid Na salts—[1-13C], [2-13C] and [1,2-13C2], and l-methionine (methyl-13C)—were used as precursors. These labeled acetates and methyl were separately and successively added to the cultures at a final concentration of 1 mg/mL on 8, 9, 10, 11, and 12 days of fungal growth. A separate culture without the addition of any precursor was incubated as a control. Nine days after the last supply of labeled acetate and methyl, the cultured mycelia were harvested, dried, ground, and extracted with EtOAc with shaking for 24 h at room temperature. Mycelia were removed by filtration; the filtrate was evaporated to afford a residue, which was then subjected to silica gel column chromatography. The column was developed successively with EtOAc:n-hexane (50:50, v/v) and EtOAc:2-propanol (50:50, v/v) to give a pure compound, which was analyzed by 13C NMR [6, 7].
Results and Discussion
Production of Integric Acid in X. feejeensis 2FB-PPM08M
The investigation of the correlation between growth stage and production of integric acid was carried out to determine the optimum growth stage of the fungi, and investigated for their mycelial biomass and integric acid production. The optimum growth stage where organisms showed the optimal production of integric acid was chosen according to this study. The production of integric acid of X. feejeensis 2FB-PPM08M can be seen in (Fig. 2). Within 3–9 days of cultivation, the fungus was in lag phase and on day 12, initiated in exponential phase and stationary phase until 21 days of fermentation. Therefore, the continuous higher production of integric acid starting after 12 days of cultivation was investigated.

Fungal growth and integric acid production during cultivation of X. feejeensis 2FB-PPM08M for 21 days  indicates integric acid production;
indicates integric acid production;  indicates mycelial biomass
indicates mycelial biomass
Biosynthesis of Integric Acid from X. feejeensis 2FB-PPM08M
To clarify the biosynthetic origins of integric acid in X. feejeensis 2FB-PPM08M, [1-13C], [2-13C], and [1,2-13C2] acetates, and l-methionine (methyl-13C) were fed to cultures of X. feejeensis 2FB-PPM08M in separate experiments. 13C-Labeled precursors were added at the 8, 9, 10, 11, and 12 days of growth, at a final concentration of 1 mg/mL for each precursor, using previously optimized growth conditions. Separate growth experiments were performed without the addition of labeled precursors as control. Cultures were harvested after 9 days for each growth experiment using 13C-labeled precursors. After the cultured mycelia were harvested, dried, ground in a mortar, then extracted with EtOAc, and filtered on vacuum system. Mycelia filtrate was evaporated and subjected to silica gel column chromatography to give an integric acid, which was analyzed by 13C NMR.
Separate X. feejeensis 2FB-PPM08M growth experiments in a total volume of approximately 1.5 L were performed using acetic acid Na salts—[1-13C], [2-13C], and [1,2-13C2], and l-methionine (methyl-13C) (see “General Experimental Procedures” section). After growth in the presence of the labeled precursors, mycelia extraction, and purification of integric acid, 13C NMR analyses clearly showed the incorporation of the labeled precursors. Results of the incorporation experiments with 13C-labeled precursors are given in (Table 1; Figs. 3, 4).

Isotopically labeled precursors were created by biosynthesis a obtained from [1-13C] acetic acid Na salt, b obtained from [2-13C] acetic acid Na salt. Filled circles indicate 13C-enriched atoms

13C-Labeling patterns of integric acid. Bold lines indicate 13C-labeled isotopomers with directly adjacent 13C atoms; arrows indicate [1,2-13C2] acetic acid Na salt
Starting from the labeling pattern of integric acid from cultures of X. feejeensis 2FB-PPM08M growing on [1-13C] acetate as the sole carbon and energy source, a retrobiosynthetic scheme led us to deduce the labeling pattern of intermediates involved in the biosynthesis of the compound. This shows clearly that, starting from [1-13C] acetate, since a combination of acetyl starter and molonyl extender would give a straight-chain poly-β-ketoester, the polyketide synthase (PKSs) catalyze a sequence of decarboxylative condensation reactions between acyl thioester and malonyl-CoA followed by a cycle of ketoreduction, dehydration, and enoyl reduction activities by the polyketide pathway. While the terpenoid pathway, acetyl-CoA, the precursor of isoprenic units, has been synthesized by the reduction of HMG-CoA synthase (HMGS), HMG-CoA reductase (HMGR), Mevalonate kinase (MK), Phosphomevalonate kinase (PMK), and Mevalonate diphosphate decarboxylase (MDPD), the condensation of isopentenyl pyrophosphate (IPP) arising from the activated acetate pool, the addition of a further C5 IPP unit to geranyl diphosphate (GPP) in an extension of the GPP synthase reaction leads to the fundamental sesquiterpene precursor farnesyl diphosphate (FPP). FPP can then give rise to linear and cyclic sesquiterpenes, [2-13C] acetate (i.e., with an inverted labeling by comparison with that of the starting material) (Fig. 3), and [1,2-13C2] acetate, indicating the presence of 13C–13C couplings (Fig. 4). In addition to using l-methionine (methyl-13C), methyl groups of the side chain were labeled (Table 1).
The 13C NMR spectrum of integric acid enriched with [1-13C] acetate gave, after isolation, a compound with significant incorporation at seven carbon atoms (C-2, C-4, C-6, C-8, C-1′, C-5′, and C-7′) (Fig. 3a), while five carbon atoms (C-1, C-3, C-9, C-12, and C-15) were enriched by [2-13C] acetate (Fig. 3b). An additional feeding experiment with [1,2-13C2] acetate confirmed this result, indicating that acetate units were incorporated into the molecule (C-1 to C-15 and C-1′to C-10′). The incorporation of l-methionine (methyl-13C) indicates that the methyl group was clearly methionine derived, confirmed after a feeding experiment with l-methionine (methyl-13C) that showed a very significant incorporation at C-9′ and C-10′ (Table 1). Therefore, the side chain of the compound (C1′ to C8′) would be synthesized by the polyketide pathway (Fig. 3a), [6, 7], while the ring carbon indicated that biosynthesis of the compound was probably via the mevalonate (MVA) or 2-C-methyl-D-erythritol 4-phosphate (MEP) pathway. Because acetate is utilized as a carbon source, it is metabolized via the glyceraldehyde 3-phosphate (GAP)/pyruvate pathway [8]. However, based on this approach, it has been established that archaea, certain bacteria, yeasts, fungi, and some protozoa and animals use only the MVA pathway, whereas many bacteria, green algae, and some protists rely on the MEP pathway [9].
13C-labeled acetate incorporation experiments to confirm terpenoid biosynthesis of the compound were performed in PDB medium, and the results of the experiments support the terpenoid pathway, as shown in (Table 1; Fig. 3). The detected 13C signal satellites in the NMR spectrum of integric acid enriched with [1,2-13C] acetate were attributed to 8 adjacent 13C2 units, from a detailed analysis of coupling constants. More specifically, three 13C2 units (C-1′to C-2′, C-3′ to C-4′, and C-7′ to C-8′) were formed in the polyketide side chain of the compound. Five 13C2 units (C-1 to C-2, C-3 to C-4, C-5 to C-6, C-7 to C-8, and C-11 to C-12) in the terpenoid moieties are observed in (Fig. 4).
The use of doubly 13C-labeled precursors provides information on the mode of assembly, cyclisation, and rearrangement of polyketide and terpenoid carbon skeletons [10]. A labeling experiment using [1-13C] acetate was determined by terpenoid biosynthesis. This revealed a biosynthetic pathway, starting from acetate, activated as acetyl-CoA and yielding IPP, which is the biological equivalent of isoprene and presents the basic branched C5 skeleton of the isoprenic unit. In particular, isotopically labeled MVA and acetate were always efficiently incorporated into the sterol, the triterpenoids, and quite often also into the sesquiterpenoids [11]. In addition, mevalonic acid originates from three molecules of acetyl-CoA, which therefore eventually contribute to the biosynthesis of IPP. By this exclusive use of acetyl-CoA, the MVA pathway should produce sesquiterpenoids that are significantly depleted in 13C isotoners [12]. For the microorganism, the presence of the MVA route was confirmed by the labeling pattern found after incorporation of [1-13C] acetate [8]. However, in further experiments, we will use 13C-labeled mevalonic acid to prove that it is a MVA pathway because mevalonic acid is in the downstream of the acetyl-CoA at MVA pathways.
References
Singh SB, Zink D, Polishook J, Valentino D, Shafiee A, Silverman K, Felock P, Teran A, Vilella D, Hazuda DJ, Lingham RB (1999) Structure and absolute stereochemistry of HIV-1 integrase inhibitor integric acid. A novel eremophilane sesquiterpenoid produced by Xylaria sp. Tetrahedron Lett 40:8775–8779
Singh SB, Felock P, Hazuda DJ (2000) Chemical and enzymatic modifications of integric acid and HIV-1 integrase inhibitory activity. Bioorg Med Chem Lett 10:235–238
Srisapoomi T, Ichiyanagi T, Nakajima H, Aimi T, Boonlue S (2014) Biological activities of integric acid isolated from the wood-decay fungus Xylaria feejeensis 2FB-PPM08M. Chiang Mai J Sci 42:xx–xx
Rogers JD (1985) Anamorphs of Xylaria: taxonomic considerations. Sydowia 38:255–262
Waalboer DCJ, Kalkeren HA, Schaapman MC, Delft FL, Rutjes FPJT (2009) Total synthesis and absolute stereochemistry of integric acid. J Org Chem 74:8878–8881
Furumoto T, Hamasaki T, Nakajima H (1999) Biosynthesis of phytotoxin neovasinin and its related metabolites, neovasipyrones A and B and neovasifuranones A and B, in the phytopathogenic fungus Neocosmospora vasinfecta. J Chem Soc Perkin Trans 1:131–135
Romminger S, Pimenta EF, Nascimento ES, Ferreira AG, Berlinck RGS (2012) Biosynthesis of two dihydropyrrole-polyketides from a marine-derived Penicillium citrinum. J Braz Chem Soc 23:1783–1788
Putra SR, Disch A, Bravo JM, Rohnmer M (1998) Distribution of mevalonate and glyceraldehyde 3-phosphate/pyruvate routes for isoprenoid biosynthesis in some Gram-negative bacteria and mycobacteria. FEMS Microbiol Lett 164:169–175
Wilson RJM (2002) Progress with parasite plastids. J Mol Biol 319:257–274
Simpson TJ (1998) Application of isotopic methods to secondary metabolic pathways. Top Curr Chem 195:1–48
Rohnmer M (1999) The discovery of a mevalonate-independent pathway for isoprenoid biosynthesis in bacteria, algae and higher plants. Nat Prod Rep 16:565–574
Jux A, Gleixner G, Boland W (2001) Classification of terpenoids according to the methylerythritol phosphate or the mevalonate pathway with natural 12C/13C isotope ratios: dynamic allocation of resources in induced plants. Angew Chem Int Ed 40:2091–2094
Acknowledgments
We would like to thank the Global COE Program (Tottori University, Japan) on Advanced Utilization of Fungus/Mushroom Resources for Sustainable Society in Harmony with Nature for their valuable support and analytical processes.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Srisapoomi, T., Ichiyanagi, T., Nakajima, H. et al. Biosynthesis of Integric Acid Isolated from the Wood-Decay Fungus Xylaria feejeensis 2FB-PPM08M. Curr Microbiol 70, 550–555 (2015). https://doi.org/10.1007/s00284-014-0759-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00284-014-0759-5