
Abstract
Hydrological processes drive the ecological functioning and sustainability of cottonwood-dominated riparian ecosystems in the arid southwestern USA. Snowmelt runoff elevates groundwater levels and inundates floodplains, which promotes cottonwood germination. Once established, these phreatophytes rely on accessible water tables (WTs). In New Mexico’s Middle Rio Grande corridor diminished flooding and deepening WTs threaten native riparian communities. We monitored surface flows and riparian WTs for up to 14 years, which revealed that WTs and surface flows, including peak snowmelt discharge, respond to basin climate conditions and resource management. WT hydrographs influence the composition of riparian communities and can be used to assess if potential restoration sites meet native vegetation tolerances for WT depths, rates of recession, and variability throughout their life stages. WTs were highly variable in some sites, which can preclude native vegetation less adapted to deep drawdowns during extended droughts. Rates of WT recession varied between sites and should be assessed in regard to recruitment potential. Locations with relatively shallow WTs and limited variability are likely to be more viable for successful restoration. Suitable sites have diminished greatly as the once meandering Rio Grande has been constrained and depleted. Increasing demands on water and the presence of invasive vegetation better adapted to the altered hydrologic regime further impact native riparian communities. Long-term monitoring over a range of sites and hydroclimatic extremes reveals attributes that can be evaluated for restoration potential.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Hydrological processes driven by groundwater and surface water interactions strongly influence the ecological functioning and sustainability of riparian ecosystems (Webb and Leake 2006). In arid regions where evapotranspiration (ET) greatly exceeds annual rainfall, these systems are clearly groundwater dependent (Eamus and Froend 2006; Newman et al. 2006; Cleverly et al. 2015). Occasional overbank flooding and reliable access to groundwater regenerate and sustain native riparian vegetation (Stromberg et al. 1996; Eamus et al. 2006). In the western United States, seeds released by cottonwood (Populus spp.) and willow (Salix spp.) in late spring have a brief opportunity to germinate following flows sufficient to scour favorable sites and maintain moisture as floodwaters subside (Crawford et al. 1996; Naiman et al. 2005; Braatne et al. 2007; Burke et al. 2009; Benjankar et al. 2014; Shafroth 2010). Snowmelt runoff from montane regions can promote regeneration by inundating floodplains and elevating water tables (WTs) with favorable timing, magnitude, and duration. Once established, phreatophytes depend on sustained access to shallow WTs and their associated capillary fringe.
Altered hydrologic regimes are a primary cause for native riparian vegetation loss or replacement (Howe and Knopf 1991; Poff et al. 1997; Stromberg 2001; Braatne et al. 2007; Burke et al. 2009; Merritt and Poff 2010). Over the past century in New Mexico’s Middle Rio Grande (MRG) corridor, sub-water table canals, diversions, dams, and levees were constructed to promote drainage, irrigation, and to control flooding (Crawford et al. 1996), greatly altering the river’s natural flow regime (Poff et al. 1997). Diminished flood pulses and deepening WTs greatly attenuated river–floodplain connectivity and threaten the recruitment and sustainability of native cottonwood-dominant communities (Braatne et al. 1996; Scott et al. 1999; Cowley 2006; Rood et al. 2008; Smith and Finch 2016), which are already stressed by frequent droughts and invasive vegetation.
Groundwater alterations often take years to become evident, leading to unforeseen problems that might be avoided by informed resource management (Alley et al. 1999). Long-term monitoring of WTs reveals the cumulative effects of altered groundwater availability and can provide insights into ecosystem adaptability (Eamus and Froend 2006). Important attributes include normal patterns of change in groundwater depth, including spatial and temporal records of minimum and maximum depths, and the rate of change in depth (Eamus et al. 2006). Water level hydrographs are a useful means to demonstrate range, seasonality, and hydrologic stresses, and their value and reliability are augmented by high measurement frequency (Taylor and Alley 2002). Long-term groundwater data also help explain why levels change (e.g. management, land-use, and climate) and improve forecasts of future water levels (Molles et al. 1998; Taylor and Alley 2002). In New Mexico’s MRG, the degree to which regeneration and sustainability of cottonwoods and willows adaptability can be maintained in response to changing surface and groundwater flows is not clearly defined (Dello Russo 2007; Smith and Finch 2016).
Regeneration of native cottonwood floodplain forests occurs in response to the river’s natural flow regime (Howe and Knopf 1991; Poff et al. 1997; Braatne et al. 2007). This requires overbanking flows of sufficient energy to scour exposed sites, the timing, duration and frequency of such flows to coincide with seed dispersal, and recession rates of the elevated water table to meet root growth and extension requirements of seedlings (Rood et al. 2005; Braatne et al. 2007). Cottonwoods and willows are commonly the dominant vegetation along perennial and free-flowing rivers, while saltcedar (Tamarix spp.) is more dominant along intermittent and flow-regulated streams (Stromberg et al. 2007). With some exceptions (e.g., Coble and Kolb 2013), cottonwood recruitment is severely limited by even minor flow alterations, while saltcedar can successfully recruit in heavily regulated, as well as free flowing streams (Merritt and Poff 2010; Mortenson and Weisberg 2010). Peak river flows in the MRG commonly occur during snowmelt runoff, when cottonwoods are dispersing their seeds. Occasionally, peak flows result from monsoonal storms later in the growing season, especially in the southern reaches of the MRG (Cleverly et al. 2006). Unlike the typical peak flows during snowmelt runoff, later peak flows favor the establishment of saltcedar, which produce seeds throughout the growing season (Warren and Turner 1975; Stromberg 1997; Smith and Finch 2016).
In this study, we investigate how alluvial riparian groundwater levels respond to surface flows influenced by regional climate conditions along the MRG in New Mexico. Our objectives were (1) to describe the hydrological regimes (surface flows/flooding and groundwater depths) that affect riparian vegetation during the snowmelt period coinciding with cottonwood germination and the full growing season, when cottonwoods of all ages depend on access to WTs; (2) to assemble a rigorous, high-frequency collection of water table data and assess the variability in their hydrographs (annual, daily drawdown) important to native vegetation recruitment and sustainability, (3) to quantify an ecosystem response, ET, to surface and groundwater levels; and (4) to maintain where possible a long-term data collection to capture variability in climate, river flow, and WTs over several years. Long-term hydrological monitoring of flood- and groundwater-dependent ecosystems contributes to the management and rehabilitation of these endangered ecosystems. The MRG is characterized by snowmelt-driven surface flows (influenced by climate conditions and water resource management) with native and non-native dominated riparian reaches and with sites containing varying degrees of hydrologic connectivity between the river and floodplain. The data from this study will contribute to a growing knowledge base attempting to interpret how climate and water availability affect dryland riparian communities and offer insights into how they might respond to shifts in these drivers.
Methods
Site Descriptions
We monitored WTs at four sites along the Rio Grande floodplain in central New Mexico for up to 14 years with accompanying ET data for up to 12 years. Research sites are located within the semi-arid MRG corridor of New Mexico, often defined (e.g., for water budget analyses; Dahm et al. 2002) as the 320 km reach between the northern, upstream Otowi, and southern, downstream Elephant Butte U.S. Geological Survey gages (Fig. 1). Full growing season data are presented. We also collected concomitant data from a hydrological climate index and Rio Grande discharge data to determine relationships between regional climate conditions, surface flow, and WTs. Within-season snowmelt conditions and discharge critical to cottonwood germination were also considered. Snowmelt generally provides the majority of the streamflow in the basin (Llewellyn et al. 2013), although summer flash floods can contribute substantially to surface flows south of the confluence with the Rio Puerco (Vivoni et al. 2006). Groundwater flow in the alluvial aquifer generally follows the southerly flow of the river and is controlled by the river, artificial pathways (conveyance channels, drains), and adjacent, subsurface inflow (Anderholm 1987; Barroll and Reiter 1995). The floodplain was historically composed of native vegetation dominated by Rio Grande cottonwood (Populus deltoides ssp. wislizeni) and understory willows (Salix ssp.). Non-native vegetation, in particular saltcedar (Tamarix chinensis) and Russian olive (Elaeagnus angustifolia), were introduced to North America in the 19th century, eventually escaping cultivation to become the third and fourth most common riparian trees in the western U.S. (Friedman et al. 2005). More tolerant of the altered hydrologic regime of the 20th century, these species became well established along the Rio Grande (Knopf and Olson 1984; Crawford et al. 1996; Di Tomaso 1998; Everitt 1998; Katz and Shafroth 2003).

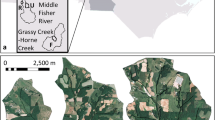
Map of the riparian research sites along the Middle Rio Grande corridor of New Mexico, located between the USGS Otowi and Elephant Butte river gages. ABQ: mature cottonwood forest, non-flooding, Albuquerque. LJY: Young Russian olive/willow forest, flooding, La Joya State Game Refuge. SEV: mature saltcedar mixed species forest, non-flooding, Sevilleta National Wildlife Refuge. BDA: mature saltcedar monotypic forest, flooding, Bosque del Apache National Wildlife Refuge
WT and ET data were collected between 2000 and 2013. The four sites with the longest record of WT and ET monitoring are included in this study (Fig. 1; Table 1). They are, from upstream to downstream: (1) a cottonwood-dominated site near Albuquerque (ABQ), which had its saltcedar and Russian olive understory removed in 2004 (Cleverly et al. 2006; Martinet et al. 2009) and was partially burned by a wildfire in 2007 (Cleverly et al. 2015); (2) an emerging Russian olive-dominated and willow-dominated site at the La Joya State Game Refuge (LJY); (3) a site of saltcedar thickets interspersed with saltgrass (Distichlis spicata) and halophytes at the Sevilleta National Wildlife Refuge (SEV) and part of the Sevilleta Long Term Ecological Research (LTER) program; and (4) a dense saltcedar-dominated site within the Bosque del Apache National Wildlife Refuge (BDA). Given sufficient discharge during snowmelt and summer monsoonal storms, the river inundates the floodplains at LJY and BDA. SEV flooded only once across the entire record in September 2013 for a single day, and ABQ remained dry. Near-well hydraulic conductivities (from slug tests) range from 1.3 m day−1 in the silty sand and clay lenses at BDA to 41 m day−1 in the sandy sediments at ABQ. We also measured leaf area index (LAI) to estimate canopy structure at ABQ, SEV, and BDA, described in Dahm et al. (2002) and Cleverly et al. (2006). The Rio Grande corridor and sites are described in more detail by Crawford et al. (1996), Dahm et al. (2002), Cleverly et al. (2006), and Martinet et al. (2009).
Monitoring and Datasets
WT monitoring wells were located in the historical floodplain between existing, constructed levees and the main channel. Wells were installed ~1 m below the baseflow WT. Wells were constructed of 5 cm inner diameter PVC pipe with 1 m long, 0.25 mm slot screen lengths, capped at the bottom. Each site had a network of five wells: a central well (C) and four wells 40 m from the center well in the four cardinal directions (N, S, E, W). Pressure transducers logged WT levels at 30 min intervals in most of the wells at various times (i.e., gaps in measurements were inevitable from time to time in some wells). Along with all data collection, water levels were determined manually to calibrate transducer measurements.
Growing season and daily rates of ET were measured with eddy covariance flux towers placed within the internal boundary layer of the riparian corridor (for detailed descriptions see Cleverly et al. 2006, 2015). Briefly, water vapor density and vertical wind speed were measured at a frequency of 10 Hz, and their covariance was computed every 30 min. Corrections were then applied to account for the physical setting of the measurements. These corrections included: two-dimensional coordinate rotation to align the wind with the sensors (Wesely 1970) and to eliminate lateral flux divergence from the conservation equations (Stull 1988); frequency-response corrections to account for the physical orientation of sensors and path averaging (Massman and Clement 2004); conversion of heat flux based on virtual temperature to that of actual air temperature (Schotanus et al. 1983); and the Webb–Pearman–Leuning correction for flux effects on density measurements (i.e., density of water vapor in air; Webb et al. 1980).
Approved Rio Grande surface discharge data from the USGS Albuquerque gage were obtained from the agency’s National Water Information System web interface (USGS, gage #08330000, waterdata.usgs.gov/nwis). Approved data were released by the USGS following quality control. This gage is the closest in proximity to our sites with reliable, long-term data and captures snowmelt runoff into our sites. Climate conditions during the 2000–2013 study were characterized by the Palmer Hydrologic Drought Index (PHDI). Mean monthly data were collected from the National Oceanic and Atmospheric Administration National Climatic Data Center (NOAA). PHDI data were averaged from the regions that affect the MRG: NOAA New Mexico Climate Divisions 2 (Northern Mountains) and 5 (Central Valley).
Reductions in streamflow, storage, and groundwater elevations are indicators of hydrological drought, where impacts can linger long after the effects of a meteorological drought have subsided (Heim 2002; Keyantash and Dracup 2002). PHDI is a variation of the Palmer Drought Severity Index (PDSI) that uses the index from longer term, established spells rather than incipient indices (Guttman 1991). Thus, it is considered a hydrological rather than a meteorological drought index. Dahm et al. (2003) found that for this region, PHDI and PDSI provide comparable characterizations of drought over a century. The Palmer indices are critiqued in Alley (1984) and Guttman (1991).
Data Analysis
We assessed correlations between river discharge and PHDI using simple linear regressions with change in discharge as the dependent variable and change in PHDI as the independent variable (SigmaPlot, Systat Software, Inc., San Jose, CA). We also performed regressions to assess relationships between annual mean WT level change and total growing season ET, with change in ET as the dependent variable and change in WT depth as the independent variable. We introduced a simple parameter to compare the annual range in WT fluctuations to mean WT depth as a measure of variability between sites, the range to depth ratio (R/D). Coefficients of variation were also calculated to compare annual WT variability between sites. To determine if riparian communities respond to flooding we analyzed ET rates at three sites over several years during the snowmelt period, which coincides with the longest inundations at sites that flood (LJY and BDA). We included the site located between them, SEV, as a non-flooding control. We first performed three separate, one-way analyses of variance (ANOVA, SAS Institute, Cary, NC) within each individual site to assess if ET differed among years. Post-hoc Fisher’s least-square differences (LSD) tests were used to compare ET rates within each site during each year’s snowmelt period to determine if ET was affected by flooding. Above average snowmelt flows resulting in long-duration floods occurred in 2005 and 2008.
Results
Climate and River Flow
Climate conditions and Rio Grande discharge during the 14-year study were expressed during snowmelt (mean of the months of April through June, inclusively) and over the full growing season (mean of the months of April through October, inclusively) periods (Fig. 2). PHDI averaged −1.6 (moderate drought) in both periods. Severe drought (−3) occurred during snowmelt and full growing seasons in 5 years, three of which were below −4 (extreme drought). Unusually wet periods (PHDIs > 2) occurred during snowmelt in 3 years and throughout the growing season in a single year (2005). PHDI showed a marginally stronger correlation to growing season discharge than to snowmelt discharge (r 2 = 0.63 and 0.52, respectively; p < 0.05 for both). Growing season PHDI and ET were well correlated at LJY, SEV, and BDA (Pearson’s correlation coefficients from 0.7 to 0.8, data not shown).

Palmer Hydrological Drought Index (PHDI) using the average for NOAA New Mexico regions 2 and 5 (top), and Rio Grande daily discharge at the USGS Albuquerque gage (bottom). For both PHDI and discharge, snow melt is the April–June mean for each year, and growing season is the April–October mean for each year. Mean long-term discharge (1942–2013) for snowmelt (sm, 35 m3 s−1, solid line) and growing season (gs, 65 m3 s−1, dashed line) periods are indicated
Rio Grande snowmelt flows averaged 45.3 m3 s−1 during the study period, 69% of the long-term (1942–2013) mean of 65.4 m3 s−1. Growing season flows averaged 27.4 m3 s−1, or 71% of the long-term mean of 38.3 m3 s−1. Snowmelt and growing season flows were less than 75% of the long-term mean in 10 years and less than 50% in 7 and 5 years, respectively. The years 2009 and 2010 were closest to long-term means during both flow periods (all within 97–110%). In 2005 and 2008, respectively, snowmelt discharge was 88 and 54% higher than the long-term mean, and growing season discharge was 61 and 40% above average. The year 2005 also had the wettest PHDIs both during snowmelt (3.53) and over the full growing season (2.62), far exceeding the snowmelt and full growing season means of all years (−1.55 and −1.61, respectively). Despite the two wet years, the maximum snowmelt flows that historically co-occur with cottonwood seedling germination were well below normal in most of the study years, greatly limiting flooding and deepening WTs, particularly at BDA, where WTs remained >3 m below the ground for most of the 2002–2004 growing seasons (Fig. 3). Even if high flows in 2005 had been capable of scouring sites suitable for future seedling establishment, snowmelt flows and WTs plummeted in 2006, detrimentally affecting survivorship of seedlings or saplings that might have germinated in 2005.

Mean daily WT depths for LJY, ABQ, SEV, and BDA (upper four hydrographs, left axis) and Rio Grande discharge (single lower hydrograph, right axis)
Peak daily discharge at the USGS Albuquerque gage upstream of the sites occurred during the snowmelt period (1 April–30 June) in 76% of the years in the 1942–2013 record. Peak flows associated with snowmelt generally have occurred in mid to late May. The US Army Corps of Engineers completed Cochiti Dam upstream of this gage in 1974, after which peak flows diminished by 10% and occurred 11 days later (Fig. 4). Prevailing dry conditions from 2000 through 2013 were associated with a 29% reduction in peak flows during the post-dam era (1975–2013, Fig. 4). Peak snowmelt flows capable of forming open germination sites through erosion and deposition are now constrained by the dam. The shift in peak flow timing further decouples spring runoff from the brief window of cottonwood germination.

Peak discharge (highest daily mean) during snowmelt periods (1 Apr–30 Jun) from 1942 to 2013. Mean peak discharge and the peak flow’s average date are indicated for the periods before Cochiti Dam was complete (1942–1974) and after its completion (1975–2013). A subset of post-dam discharge (2000–2013) is included to indicate the lower flows that characterized most of the snowmelt flows during our monitoring. Discharge data are from USGS gage 0833000 at Albuquerque, NM
Water Tables
For WT depths and ET we defined an extended growing season as 1 April through 15 November, which captures the earliest start of ET (6 April 2004 at SEV) and the latest termination of ET (10 November in both 2004 and 2010 at BDA; Cleverly et al. 2006). Figure 5 illustrates growing season patterns of surface flow and WTs at the sites in 3 years characterized by (a) drought, (b) intermediate conditions, and (c) unusually wet conditions. In all cases, surface flow and WT elevations peak during snowmelt. WTs at the non-flooding sites with low R/D (ABQ and SEV) were much less influenced by surface flows than at the more connected sites with high R/D (LJY and BDA; Table 1). Flood-prone sites were not inundated and experience deep WTs during drought years when flows are <60 m3 s−1 (Fig. 5a). By contrast, a much larger discharge of water (>100 m3 s−1) over most of the snowmelt period in 2005 led to extensive overbank flooding and shallow growing season WTs at LJY and BDA (Fig. 5c). Surface flow and WTs were closest to their means in 2009, which resulted in restricted flooding at LJY and especially BDA (where flooding was not reflected in mean WT depth; Fig. 5b). The range in WT depths across sites (but especially for the flood-prone sites LJY and BDA) was attenuated in the wet year, enhanced in the drought year, and intermediate in the intermediate years (Fig. 5). Overall, WTs ranged from ~4 m depth to more than 0.5 m flood height (Table 1; Figs. 3 and 5). Mean WTs ranged from ~1 m depth at LJY to 2 m depth at SEV and BDA. Like peak river flows, minimum WT depths were most frequent during snowmelt. Maximum WT depths typically occurred late in the growing season. WT levels were highly variable at the more hydrologically connected flood-prone sites (BDA, LJY), where the mean range exceeded mean depth. Minimum WT depths all occurred in the two wet years (2005 and 2008). Maximum WT depths occurred in dry years (Fig. 5a) at all sites except in the wet year (2005) at LJY, where the WT dropped from its highest peak (flooded) to its maximum depth in fewer than 4 months (Fig. 5c, Table 1).

WT hydrographs at the four study sites and Rio Grande discharge at the USGS Albuquerque gage. Half-hourly data WT and flow collected during three growing seasons characterized by different PHDIs: a severe drought (2003), b normal range (2009), and c unusually wet (2005). Mean PHDI and percent of long-term (1942–2013) average river discharge (Q) for snowmelt (SM, Apr–Jun) and full growing season (GS, Apr–Oct) are indicated
More than 80% of minimum WT depths occurred during the 1 April–30 June snowmelt period that is critical for native cottonwood establishment. Mean minima were observed during this period, excluding the less common years when short-lived minima occurred later in the summer following intense monsoonal storms. Mean minimum WT levels were measured in May, occurring progressively later upstream from BDA to ABQ, where they nearly coincided with peak river flow at the nearby USGS stream gage (Fig. 3; Table 1). For 60 days following peak elevations, WTs declined an average of 2.1 cm day−1 at BDA, with 6 of the 14 years exceeding rates of 2.5 cm day−1 (cf. declining WTs during each year’s snowmelt season in Fig. 3). WTs descended 1.8 cm day−1 at LJY and much more slowly at ABQ and SEV (0.5 cm day−1). Seedling roots at BDA and LJY are thus less likely to keep pace with the rapidly declining capillary fringe than at ABQ and SEV.
ET and Flooding
Mean growing season ET ranged from 79 cm at SEV to 119 cm at LJY. ET was highest at the sites that were flooded during wet years. ET was lowest during dry years at all sites, although the particular dry year in which ET was lowest varied across sites (Table 1). On average, ET rates were lower in the saltcedar-dominated sites, SEV and BDA, particularly at SEV, the site with the lowest leaf area index (Dahm et al. 2002; Cleverly et al. 2006). ET within sites varied by 30–40 mm annually except at ABQ (16 mm), which also exhibits the least ranging WT depths (Table 1).
Growing season ET declined with deeper WTs within each site across years (Fig. 6). Lower rates of ET during years associated with drought and deeper WTs at BDA may be indicative of water stress, although saltcedars can tolerate these conditions more readily than native species. Stronger correlations between ET and WT were observed at BDA and LJY, sites with dynamic WT fluctuations and larger range to depth ratios than at ABQ or SEV. Relationships between growing season WT and ET were very weak at sites that do not have a recent history of flooding (ABQ and SEV; Fig. 6). In contrast to within sites, WT depth itself is not a reliable ET indicator across sites. As an example, SEV and BDA have similar mean WT depths but very different ET rates (Table 1).

Relationships between mean growing season WT depth and ET over several years. Lines are least-squares regressions (p < 0.01 at LJY and BDA). CV = mean of yearly coefficients for variation in WTs. The year 2008 was excluded at BDA due to incomplete ET data
Spring snowmelt floods were larger in magnitude and duration than flashier summer monsoonal floods. To examine the effect of sustained flooding on the ecosystem, we compared mean daily ET rates within the flood-prone sites (BDA, LJY) during each year’s peak historic spring runoff period (1 May–15 June) over the period when LJY was mostly operational (2003 through 2008). Complete inundation occurred over the entire period in 2005 and 2008 at BDA and in 2005 at LJY, which was inundated in 2008 > 60% of the time and the soil remained saturated despite a mean WT elevation just below the ground surface (Fig. 7). Within each of these sites as well as non-flooding SEV, ET during snowmelt varied significantly across years (ANOVA, for LJY F = 13, df = 4217, p < 0.0001; for SEV F = 6, df = 5271, p < 0.0001; for BDA p = 24, df = 5268, p < 0.0001). Fisher’s LSD post-hoc tests revealed that ET was significantly higher at LJY in flood years vs. dry years (p < 0.05). By contrast, significant differences in ET across years at BDA did not coincide with flooding patterns, including the extremely dry snowmelt period in 2006 (Figs. 2 and 7). In fact, ET was significantly lower in one of the flood years (2005) than in several dry years at this site. Thus, riparian communities along the river do not respond uniformly to flooding.

Mean WT depth and mean daily ET during peak snowmelt runoff (1 May–15 June). Average inundation over the complete peak snowmelt period occurred at BDA in 2005 and 2008 and at LJY in 2005. LJY was flooded for most of 2008, although lower WTs at the furthest two wells from the channel resulted in a slightly higher subsurface mean. LJY was excluded in 2004 due to limited ET data. SEV is included here as non-flooding control site. Within each site, yearly ET means are compared to determine if they exhibit annual differences in response to flooding. ET rates with the same letter (BDA: a, b, c; SEV: p, q, r; LJY: x, y, z) are not significantly different at the 0.05 level (ANOVA with post-hoc Fisher’s LSD tests, SAS Institute 2015)
Discussion
The Drought Index
In general, river flow, WTs, and ET reflected the dry climatic conditions during the period of study (Figs. 2 and 3). Prevailing hydroclimatic conditions during the 14 years of monitoring were moderately dry, with several severe to extreme droughts encompassing both the snowmelt period and the overall growing season (Fig. 5a). Wet conditions and high river flows (Fig. 5c) were much less frequent. Droughts covered much of the western U.S. during this period. From 2000 to 2014, annual Upper Basin Colorado River flows averaged over a 19% reduction from the 1906 to 1999 mean (Udall and Overpeck 2017). High flows in 2008 did not follow particularly wet conditions and were more likely driven by management decisions to store and move water downstream following a Rio Grande Compact agreement early in 2008 (New Mexico Interstate State Stream Commission 2009). The high 2008 discharge despite a normal PHDI (Figs. 2 and 3) illustrates that water resource management can exert greater influence on flows than climate conditions. Low flows were aligned well with PHDI during the driest years, particularly during the severe droughts in 2002–2003 and again in 2011–2013 (Fig. 2), which included severe droughts across the continental USA in 2012 (Wolf et al. 2016) and across the southwestern United States in 2011 (Moore et al. 2016; van Gestel et al. 2016). Although growing season PHDI and ET were well correlated at three of the four sites, no correlation was observed at ABQ, located just downstream from the Albuquerque wastewater reclamation plant’s outflow. A substantial and steady contributor to the MRG, this facility likely maintains the shallow, stable, low ranging WT and mitigates the impact of drought at this site.
The interaction of atmospheric moisture and the hydrology of the basin drive riparian ecosystem functioning in the MRG (Crawford et al. 1996). Restoration is therefore dependent on the extent to which climate and anthropogenic management influence the flow regime. As streams respond to a variable and changing climate, riparian plant life cycles are likely to exhibit various responses to hydrological changes (Smith and Finch 2016). Long-term monitoring of surface water and groundwater in tandem with plant bioindicators can help explain the dominant vegetation observed at riparian zones, help assess how species respond to changes, and instruct flow management (Stromberg et al. 2006). PHDI can be a useful operational tool for ecological and water resource management as a hydrological index influenced by streamflow, reservoir storage, and groundwater levels and with a recovery that lags behind more immediate meteorological indices (Guttman 1991; Keyantash and Dracup 2002; Svoboda et al. 2002). Snowpack measurements and indices like PHDI help to assess and project riparian moisture conditions at the onset of and throughout growing seasons, but their effectiveness might be compromised by water management and increasing temperatures (Woodhouse et al. 2016; Udall and Overpeck 2017). Although we focused on snowmelt and full growing season periods, winter droughts in the snowmelt-driven western US also can significantly affect entire growing seasons that follow, where deficits can linger to the following winter (Redmond 2002).
Snowmelt Runoff, Groundwater Drawdown, and Cottonwood Establishment
Gradual WT decline subsequent to scouring floods promotes root elongation in cottonwoods (Segelquist et al. 1993; Amlin and Rood 2002; Sher and Marshall 2003). Cottonwood seedling roots have been shown to grow up to 1 cm day−1 and 60–100 cm year−1 (Mahoney and Rood 1998). Mahoney and Rood (1998) found that a WT decline of ~2.5 cm day−1 is likely the maximum rate at which the seedlings can survive, although this rate is probably too rapid for survival in the typically coarse sediment substrate and hot, dry environment typical of the MRG prior to monsoon season (Braatne et al. 2007). For example, during a WT drawdown of more than 2 cm day−1 over 20 days in 2002, depleted soil moisture in the root zone led to widespread cottonwood seedling mortality at a site near BDA (Bhattacharjee et al. 2006). In another study, seedling mortality increased greatly after 3 weeks at a recession rate of 0.7 cm day−1 (Segelquist et al. 1993). Thus, WTs at BDA and possibly LJY may recede too quickly for successful germination of cottonwood (Amlin and Rood 2002) after a spring inundation in wet years and may remain too deep at BDA in dry years. Even though the sites continue to flood, their dynamic WTs may be a disadvantage for cottonwood establishment, particularly in the presence of saltcedar. WT depths at the non-flooding sites were more amenable to cottonwood root growth, but they lacked overbanking spring flood pulses and thus would require extensive management (e.g., diversions of relatively high-energy water, clearing, excavation, and planting or seeding) for establishment of cottonwood communities.
Similar to the reduction in overall river flows, peak snowmelt runoff in the MRG during our 2000–2013 monitoring period was substantially lower than the post-dam mean, likely as a result of prevailing drought (Fig. 4). Cottonwoods are more susceptible than saltcedar to reduced flow and altered timing of peaks (Braatne et al. 2007; Merritt and Poff 2010), which are typical hydrological responses downstream of dams (Webb and Leake 2006; Burke et al. 2009). Although Cochiti Dam operations may have extended peak flow timing later into the growing season (Fig. 4), many free-flowing, snowmelt-driven rivers are experiencing earlier peak flows, partially uncoupling seasonal flow from seedling establishment (Rood et al. 2008). Stella et al. (2006) found that early season temperatures influence snowmelt runoff and the development and release of seeds in Fremont cottonwoods (Populus fremontii). In the MRG, the shift towards managed, later peak flows could be further detrimental to cottonwood regeneration, especially if seeds are released earlier in response to warming temperatures.
Despite two wet years with extended snowmelt overbank flooding and favorably elevated WTs for germination (2005 and 2008), discharge peaks of up to 191 m3 s−1 did not attain the magnitude necessary to scour substantive new seedling sites. Floods (defined by monitoring well water columns ascending above the ground surface) did endure at LJY and BDA; however, for 60% (55 days) and 100% (91 days) of the snowmelt period in 2005 and 2008, respectively. This was in part due to operational decisions related to upstream and downstream water storage. Inundation of this duration can result in a hypoxic root environment detrimental to seedlings and mature plants (Crawford et al. 1996; Cleverly et al. 2006; Auchincloss et al. 2012; Cleverly 2013). However, some riparian species of cottonwoods, willows, and alders tend to be moderately tolerant of inundation and saturated soils (Harrington 1987; Mahoney and Rood 1992; Scott et al. 1999; Auchincloss et al. 2012), particularly those species located in low elevation, near-channel habitats (Amlin and Rood 2001). Effects of hypoxia are likely attenuated to some extent by the well-oxygenated water overbanking the channel near our sites, which has also been shown to increase dissolved oxygen in groundwater (Valett et al. 2005) and to stimulate production of adventitious roots near the high-water mark (Cleverly 2013). In the Rio Grande corridor, a series of frequent, high intensity, short duration flood pulses might provide an influx of more oxygenated water and interrupt periods of hypoxia and therefore be more beneficial to native riparian vegetation, provided WT levels maintain capillary soil moisture for seedling roots (Crawford et al. 1996; Follstad Shah and Dahm 2008; Auchincloss et al. 2012).
Water Table Depths and Fluctuations
WTs tracked river discharge quite well over the course of 14 growing seasons (Fig. 3). Compared to the first 7 years, mean WTs from 2007 to 2013 were 36% shallower (0.8 m) at BDA and ≤6% shallower or deeper at SEV and ABQ. The change in the 7-year mean is likely due to higher river flows and elevated WTs during the first three consecutive years of the latter period (2008–2010), whereas 6 of the 7 years of the reference period (2000–2006) were drier than normal (Fig. 2). The greater effect at BDA may have also been due to flow redirected toward the site when the channel was altered by the high discharge in 2008 (personal communication, Bosque del Apache NWR staff). WTs responded to flows in dry, wet, and normal years, particularly at the more connected, higher ranging, flooding sites (LJY and BDA, Fig. 5). BDA groundwater averages four times the annual range of WT levels at ABQ and SEV (Table 1, Fig. 5). Surface water and groundwater interactions near SEV and ABQ were likely to be more stable due to water that accumulates behind the US Bureau of Reclamation’s San Acacia Diversion Dam 2 km downstream of SEV and Albuquerque’s wastewater treatment outflow 8 km upstream of ABQ. Thus, we found that the effects of drought on WT levels at some of our study sites were partially ameliorated by water resource management in the MRG.
Surface/subsurface connectivity indicates that groundwater availability to alluvial riparian ecosystems is susceptible to changes in Rio Grande surface flows. Saltcedar is a facultative, deep-rooted plant capable of growing across a wide range of WT depths (e.g., BDA hydrographs, Fig. 5), whereas cottonwoods are restricted to shallower WT depths (e.g., ABQ hydrographs, Fig. 5; Robinson 1958; Howe and Knopf 1991; Busch and Smith 1995; Mahoney and Rood 1998; Horton et al. 2001; Lite and Stromberg 2005). Because cottonwoods and other native vegetation are vulnerable to cavitation caused by drought stress, their distribution in arid regions is limited to locations with reliable WTs, often near streams (Pockman and Sperry 2000). Cottonwoods at ABQ were mostly established prior to the construction of Cochiti Dam. They are confined by levees to a limited zone adjacent to the river. Though the site no longer floods and sees little regeneration, the mature cottonwood community has access to a shallow and low ranging WT.
Cottonwoods and willows exhibit reduced stomatal conductivity and photosynthesis and increased canopy dieback and mortality with greater WT depth (Scott et al. 1999; Rood et al. 2000). Horton et al. (2001) observed water stress in cottonwoods when depth exceeded 3 m, while saltcedars, although showing some water stress with depth, did not experience mortality in the full 8 m depth range. Mean growing season WT depths at all of our sites were shallower than 3 m. However, WTs at BDA from 2002 to 2004 remained below 3 m depth for 50–61% of those three growing seasons (Figs. 3 and 5a), likely limiting cottonwood and willow viability in this saltcedar-dominated site, despite the reliable influx of flood water.
In addition to WT depth, cottonwoods are susceptible to rapid WT fluctuations and rates of decline (Mahoney and Rood 1992; Stromberg et al. 1996; Scott et al. 1999; Lite and Stromberg 2005). WTs at LJY and BDA were much more variable than at ABQ and SEV (Figs. 3 and 5; Table 1). At the dynamic, flood-prone sites, the seasonal range in WTs usually exceeded the mean depth with range to depth (R/D) ratios exceeding unity. Thus, the mean R/D of 1.5 and an annual depth of 2 m at BDA would be expected to produce a range of 3 m, and sites with larger values of R/D could result in flooding or relatively deep WTs (or both, in the case of LJY in 2005, Fig. 5c; Table 1). Growing season WT ranges can be similar in wet and dry years. The largest R/Ds occurred at LJY and BDA in the substantial flood years (2005 and 2008), when mean WT depths were shallowest. The stable, non-flooding sites had more limited ranges, maintaining R/Ds of less than 0.5 with similarly shallow depths (Table 1). Variations in range and depth of WTs are important indicators of riparian restoration potential (Segelquist et al. 1993; Scott et al. 1999; Shafroth et al. 2008) and likely are indicative of sites that better support native species (ABQ) or invasive species (BDA). In locations with high R/D ratios, cottonwoods may be at a disadvantage. Saltcedars are more flexible phreatophytes with the potential to produce deep roots that can more effectively extract water from unsaturated soil and thus are less impacted by variable depths (Busch and Smith 1995; Cleverly et al. 2006). In two southwestern U.S. studies cottonwood abundance was greater at sites with less fluctuation in WTs (Lite and Stromberg 2005), and sapling survival was more dependent on limited range than depth (Shafroth et al. 2000). Along the San Pedro River in Arizona, Lite and Stromberg (2005) found that native trees dominated at sites with small WT fluctuations (<0.5 m) and mean depths of less than 2.6 m, which equates to an approximate R/D ratio of 0.2. At ABQ, our most cottonwood-dominant site, R/D was 0.4. At another cottonwood-dominated flooding site ca. 30 km downstream of ABQ (with limited ET data but 9 years of WT data), R/D averaged 0.8 (>1 in the 2 wet years). This lower R/D ratio was largely driven by proximity of that site to irrigation ditches. At a mean depth capable of supporting cottonwoods, there may be a dynamic range whereby inundation and drawdown maintain an optimal R/D (e.g., 1 or less). Conditions at LJY appear reasonably favorable. The moderately high range in WT is tempered by the shallow depth, and the site occasionally floods. A small number of young cottonwoods exist within the forest, where willows and Russian olives make up the majority of the riparian plant community.
While cottonwoods depend on the natural flow regime and shallow WTs, saltcedars tolerate a wider spectrum of flows and WT depths (Di Tomaso 1998; Dahm et al. 2002; Stromberg et al. 2006; Merritt and Poff 2010) and dominate sites with R/Ds from 0.3 (SEV) to 1.5 (BDA; Table 1). As with cottonwoods, open, exposed sites facilitate saltcedar recruitment (Merritt and Poff 2010), and saltcedars disperse seeds over a much longer portion of the growing season. Saltcedars occupy active floodplains, as well as terraces unsuitable for mesic riparian trees, such as cottonwoods (Nagler et al. 2005). Vegetation at SEV has access to relatively shallow, low-ranging WTs but is mostly limited to drought-adapted saltcedar (Cleverly et al. 1997, 2006) and other xeric species, such as salt grass, halophytes, and a few mesquite (Prosopsis ssp.). Groundwater at this site is also affected by an upwelling of deep brackish groundwater. Consequently, it has a much higher concentration of salts than the other sites (unpublished data), supports plants with higher salt tolerances, and would be a candidate for restoration to upland species instead of cottonwood and willow (Shafroth et al. 2005). Like ABQ, flooding does not generally occur at SEV (1 day in the 14- year study).
Russian olives also exhibit generalist traits and can establish in undisturbed (e.g., non-flooding) sites, in drier, elevated bottomlands, and under dense canopies given their tolerance for shade (Katz and Shafroth 2003; Nagler et al. 2005; Mortenson and Weisberg 2010; Reynolds and Cooper 2010; Hultine and Bush 2011). Russian olives thrived in the highly variable WT (R/D = 1.6) and flooding regime at LJY, rejecting in this case the hypothesis that Russian olive is favored by the reduced levels of disturbance found downstream of dams (Katz and Shafroth 2003). Cottonwoods are thus subjected both to altered hydrologic regimes and to introduced competitors with more plastic traits.
ET Responses to Water Table Depth and Flooding
Plant community water requirements can be measured by ET responses (Stromberg et al. 2006), the effects of which can be seen in sinusoidal diel WT hydrographs (Martinet et al. 2009). A quantifiable relationship between WT depth and ET is a challenge to assess at fine-scale intervals (daily or sub-daily) in riparian sites (Butler et al. 2007; Lautz 2008; Loheide 2008; Martinet et al. 2009). As in other phreatophyte investigations, ET in the MRG declined with deeper WTs on a coarse temporal scale (growing season: Devitt et al. 1998; Cooper et al. 2006). However, mean WT depths between our sites were not a reliable indicator of groundwater consumption. For example, SEV and BDA both exhibited mean WT depths ca. 2 m, but ET averaged nearly 30% greater at the latter site (Table 1). The dynamic, flooding sites (BDA and LJY, with differing vegetation) exhibited much more robust correlations between groundwater depth and ET (Fig. 6). Plants able to establish in widely ranging sites may have a more extensive network of roots that more readily utilize groundwater and available soil moisture. Higher R/D sites expose and inundate a greater extent of the biogeochemically active saturated and vadose zones throughout the growing season (Vinson et al. 2007) and may influence ET more than depth itself (Cleverly et al. 2006). We did observe an ecosystem response to flooding when ET was higher when comparing individual dates of flooding vs. dry conditions at BDA (Cleverly et al. 2015). We also examined the relationship between ET and elevated WTs and flooding at these sites during high runoff in the two wet years.
Extensive flooding in the 2005 and 2008 snowmelt periods coincided with significantly higher ET compared to dry snowmelt periods at LJY but not at BDA (Fig. 7). It is likely that standing flood water in the more open canopy at LJY had a greater effect on the evaporation component of ET and total ET than at the densely covered BDA site. This has implications for managed floods during snowmelt as a restoration strategy. While flooding may temporarily increase ET rates in more open, emerging mixed stands, inundation into mature saltcedar thickets would not likely result in excessive water consumption in this water-scarce region.
Restoration Strategies and Challenges
The differing riparian communities at the four sites we investigated are likely a product of their hydrological regimes (Fig. 5). Based on the average age determined by Howe and Knopf (1991), cottonwoods along the Rio Grande in Albuquerque were mostly established in the 1940s, when snowmelt runoff and flooding were less regulated by dams, diversions, and levees. Yet they now benefit from managed and return flows that maintain shallow, low-ranging WTs, including at the ABQ site. Prior to understory clearing in 2004, ABQ in addition to LJY contained Russian olives, illustrating the large range in suitable hydrological regimes (Katz and Shafroth 2003). The exclusion of Russian olives at SEV and BDA may be due to high concentrations of salts and the exceedingly deep WTs during droughts, respectively. Saltcedars at BDA can tolerate wide WT fluctuations, including lengthy drops in WTs during droughts. The lack of flooding at SEV is a competitive advantage for saltcedars over native riparian species.
Groundwater regimes necessary to regenerate and sustain aridland cottonwood-dominated riparian ecosystems need to be well characterized and ecosystem response to changes in groundwater availability carefully monitored (Eamus and Froend 2006; Webb and Leake 2006; Stromberg et al. 2007). Ideally, informed flow management could enable cottonwood seedling recruitment (Mahoney and Rood 1998). Given favorable WT drawdown rates, cottonwood seedlings can outcompete saltcedar in open canopy areas (Sher and Marshall 2003). Cottonwoods that can establish in suboptimal conditions at the drier end of the fluctuating moisture gradient are more likely to avoid mortality due to flooding and scouring in subsequent years and may have the best chance of surviving to maturity (Segelquist et al. 1993). In the MRG, the fringes of the floodplain at LJY might be such a site, with occasional flooding and a reasonable R/D ratio where depth remains relatively shallow. In years when snowpack is well above normal managed flooding would be a useful tool in locations targeted for recruitment and restoration of cottonwood and other native vegetation (Mahoney and Rood 1998; Molles et al. 1998; Stromberg 2001; Rood et al. 2003; Taylor et al. 2006; Follstad Shah and Dahm 2008; Merritt and Poff 2010; Benjankar et al. 2014).
In the MRG, the absence of flooding of sufficient magnitude to scour fresh substrate might require manual clearing to mimic erosional scouring and deposition in suitable sites during the narrow window of cottonwood seed dispersal (Howe and Knopf 1991; Burke et al. 2009). Flooding in high runoff years, even if lacking the energy to scour, could provide elevated WTs and drawdown rates conducive to restoration. Areas largely dominated by dense, mature saltcedar, such as BDA that are inundated with low-energy water would likely not recruit new seedlings (Merritt and Poff 2010; Reynolds and Cooper 2010) nor see increased water consumption during years with flooding (Fig. 7). Optimal site conditions for native recruitment and sustainability are challenging to locate in the regulated, water-limited Rio Grande corridor. If the latent causes of species decline are not addressed, restoration, including passive restoration to remove stressors, is often unsuccessful (Rood et al. 2003; Lite and Stromberg 2005; Shafroth et al. 2008) and squanders time and funds. A flood-prone site like BDA might appear promising for restoration (Fig. 5c), but long-term monitoring revealed that its high ranging WT may be too great to support native species during extended droughts (Fig. 5a). Despite the ability of invasive species to compete in varying hydrologic conditions, a river that can be managed to mimic its natural flow regime to the extent possible is a key component of native species restoration (Merritt and Poff 2010; Yarnell et al. 2015). In some cases the pattern of flow is more critical to native recruitment than the quantity of flow (Rood et al. 2003). Reorganizing and stabilizing disconnected ecosystems within the floodplain will likely depend on managed flooding, some level of invasive species control and long-term WT monitoring (Stromberg et al. 1996; Poff et al. 1997; Molles et al. 1998; Rood et al. 2003, 2005; Shafroth et al. 2008; Merritt and Poff 2010).
Regional temperatures in the catchment have increased over the past few decades, depleting snowpack and reducing seasonal flows. This shifts peak runoff earlier by weeks, increases ET, and escalates consumption by crops (Gutzler 2004; Lukas et al. 2014; Dettinger et al. 2015). In the MRG, irrigation storage and river management since the completion of Cochiti Dam have counteracted earlier snowmelt, thus maximum snowmelt flows now occur an average of 10–11 days later (Fig. 4). Shallow alluvial aquifers will likely be the groundwater component most vulnerable to climate change (Alley et al. 1999). Groundwater is already replacing surface water supplies for use by humans and is rapidly diminishing in the western US and globally (Castle et al. 2014; Famiglietti 2014). The hydroclimatic regime in the western US is expected to see a decrease in surface water and groundwater availability and an increase in drought frequency (McCabe and Wolock 2007; Barnett et al. 2008; Rood et al. 2008; Perry et al. 2012; Castle et al. 2014; Woodhouse et al. 2016; Udall and Overpeck 2017) particularly in the Southwest (Cayan et al. 2010; Williams et al. 2013; Cook et al. 2015) and in the upper Rio Grande watershed (Hurd and Coonrod 2012; Llewellyn et al. 2013; Dettinger et al. 2015; Elias et al. 2015). Consequently, it is imperative to optimize restoration strategies within a changing climate.
ET rates declined with WTs during drought, although the correlations were weaker at sites with more stable WTs (ABQ and SEV). We observed increased ET rates during extended flooding at an emerging, open site, but concomitant flooding at a mature saltcedar stand had no effect on ET. This indicates that inundation into ubiquitous saltcedar stands that will occur when targeting other sites with floods for regeneration may have little effect on regional water budgets. Riparian restoration will not result in large water savings in ET (Shafroth et al. 2005; Moore and Owens 2012). Rates of ET scale to leaf area, and dense riparian stands of vegetation use more water than sparse stands. However, more drought-tolerant saltcedars can persist in habitats unable to support native species and increase canopy cover in these locations within the catchment (Hultine and Bush 2011). Russian olives can potentially occupy substantial areas of the western US. They can establish well above stream channels and subsist on (and thus intercept) precipitation for several years, subsequently tapping into the alluvial aquifer (Reynolds and Cooper 2010). This species was most commonly observed by Knopf and Olson (1984) on outer edges of riparian vegetation where it meets upslope vegetation and is observed on terraces absent of native phreatophytes (Hultine and Bush 2011). Similarly, saltcedars are facultative phreatophytes that can survive on soil water without reductions in transpiration due to very deep WTs (Busch et al. 1992; Gries et al. 2003). In the absence of some extent of native restoration, saltcedar and Russian olive thickets will likely dominate riparian woodlands and could spread further into the historic floodplain, exacerbating basin ET losses and affecting water budgets (Howe and Knopf 1991; Dahm et al. 2002; Webb and Leake 2006). The impact is greater in smaller watersheds, such as the San Pedro River in Arizona (Hultine and Bush 2011).
Conclusions
Riparian ecosystems in the MRG’s semi-arid climate are dependent on groundwater and surface flows derived largely from upstream montane catchments. These flows correlate reasonably well with PHDI, a tool that can help project seasonal discharge. In addition to climatic influences, surface and groundwater flows are also impacted by irrigation dams, diversions, and wastewater discharge. We found that the magnitude and timing of snowmelt-driven peak discharge in the MRG is diminished and occurring later, respectively, following completion of a large upstream dam. River management aligned to the extent possible with a more natural functional flow regime would better meet the needs of the native riparian communities (Poff et al. 1997; Yarnell et al. 2015). Long-term monitoring of WTs in riparian ecosystems revealed widely varying annual and daily hydrographs that can indicate the requirements of existing communities and assess if potential restoration sites meet cottonwood and other native vegetation tolerances for WT depths, rates of recession, and variability throughout their life stages. Highly variable hydrographs were observed (BDA) that can preclude native vegetation that is less adapted to deep drawdowns during extended droughts. We also found that WT drawdown rates vary between sites and must be assessed in regard to recruitment potential. Sites with relatively shallow WTs and limited annual variability (R/D ≤ 1) might be ideal. A location with a lower R/D situated further into the floodplain from the channel (e.g., away from the active river channel at LJY), and occasional diversions of flood water into a non-flooding, low R/D site (ABQ) could be considered. As well, monitoring reveals that sites like SEV and BDA, where invasive species are likely to out-compete natives, can be avoided or targeted for rehabilitation using native, xeric-adapted riparian plants (Shafroth et al. 2005). Suitable restoration sites have diminished greatly as the once meandering, freely flowing Rio Grande has been constrained and depleted. Increasing demands on water and the presence of invasive vegetation that is less susceptible to the altered hydrologic regime can further impact the floodplain. Long-term monitoring at various sites over a range of hydroclimatic extremes reveals attributes that can be evaluated for restoration potential. These include the likelihood of flooding, timing of peak WT elevations, rate of WT decline following peaks, and WT depths and variability associated with snowmelt and the overall growing season that are critical to successful native plant germination and sustainability.
References
Alley WM (1984) The Palmer drought severity index: limitations and assumptions. J Clim Appl Meteorol 23:1100–1109
Alley WM, Reilly TE, Franke OL (1999) Sustainability of ground-water resources. US Geological Survey Circular 1186
Amlin NA, Rood SB (2001) Inundation tolerances of riparian willows and cottonwoods. J Am Water Resour Asssoc 37:1709–1720
Amlin NA, Rood SB (2002) Comparative tolerances of riparian willows and cottonwoods to water-table decline. Wetlands 22:338–346
Anderholm SK (1987) Hydrogeology of the Socorro and La Jencia Basins, Socorro County, New Mexico. USGS Water Resources Investigation Report 84-4342
Auchincloss LC, Richards JH, Young CA, Tansey MK (2012) Inundation depth, duration, and temperature influence Fremont cottonwood (Populus fremontii) seedling growth and survival. West N Am Nat 72:323–333
Barnett TP, Pierce DW, Hildago HG, Bonfils C, Santer BD, Das T, Bala G, Wood AW, Nozawa T, Mirin AA, Cayan DR, Dettinger MD (2008) Human-induced changes in the hydrology of the western United States. Science 319:1080–1083
Barroll MW, Reiter M (1995) Hydrogeothermal investigation of the Bosque del Apache, New Mexico. N M Geol 17:1–6.
Benjankar R, Burke M, Yager E, Tonina D, Egger G, Rood SB, Merz N (2014) Development of a spatially-distributed hydroecological model to simulate cottonwood seedling recruitment along rivers. J Environ Manag 145:277–288
Bhattacharjee J, Taylor JP, Smith LM (2006) Controlled flooding and staged drawdown for restoration of native cottonwoods in the Middle Rio Grande Valley, New Mexico, USA. Wetlands 26:691–702
Braatne JH, Jamieson R, Gill KM, Rood SB (2007) Instream flows and the decline of riparian cottonwoods along the Yakima River, Washington, USA. River Res Appl 23:247–267
Braatne JH, Rood SB, Heilman PE (1996) Life history, ecology, and conservation of riparian cottonwoods in North America. In: Stettler RF, Bradshaw Jr HD, Heilman PE, Hinckley TM (eds) Biology of Populus and its implications for management and conservation. NRC Research Press, Ottawa, ON, pp 57–85
Burke M, Jorde K, Buffington J (2009) Application of a hierarchical framework for assessing environmental impacts of dam operation: changes in streamflow, bed mobility and recruitment of riparian trees in a western North American river. J Environ Manag 90:S224–S236
Busch DE, Ingraham NL, Smith SD (1992) Water uptake in woody riparian phreatophytes of the Southwestern United States: a stable isotope study. Ecol Appl 2:450–459
Busch DE, Smith SD (1995) Mechanisms associated with decline of woody species in riparian ecosystems of the southwestern U.S. Ecol Monogr 65:347–370
Butler JJ, Kluitenberg GJ, Whittemore DO, Loheide II SP, Jin W, Billinger MA, Zahn X (2007) A field investigation of phreatophyte-induced fluctuations in the water table. Water Resour Res 43:W02404. https://doi.org/10.1029/2005WR004627
Castle SL, Thomas BF, Reager JT, Rodell M, Swenson SC, Famiglietti JS (2014) Groundwater depletion during drought threatens future water security of the Colorado River Basin. Geophys Res Lett 41. https://doi.org/10.1002/2014GL061055
Cayan DR, Das T, Pierce DW, Barnett TP, Tyree M, Gershunov A (2010) Future dryness in the southwest US and the hydrology of the early 21st century drought. Proc Natl Acad Sci USA 107:21271–21276
Cleverly J (2013) Water use by Tamarix. In: Sher A, Quigley MF (eds) Tamarix: a case study of ecological change in the American west. Oxford University Press, New York, pp 85–98
Cleverly JR, Dahm CN, Thibault JR, McDonnell DE, Coonrod JEA (2006) Riparian ecohydrology: regulation of water flux from the ground to the atmosphere in the Middle Rio Grande, New Mexico. Hydrol Process 20:3207–3225
Cleverly JR, Smith SD, Sala A, Devitt DA (1997) Invasive capacity of Tamarix ramosissima in a Mojave Desert floodplain: the role of drought. Oecologia 111:12–18
Cleverly JR, Thibault JR, Teet SB, Tashjian P, Hipps LE, Dahm CN, Eamus D (2015) Flooding regime impacts on radiation, evapotranspiration, and latent energy fluxes over groundwater-dependent riparian cottonwood and saltcedar forests. Adv Meteorol 2015:935060. https://doi.org/10.1155/2015/935060
Coble AP, Kolb KE (2013) Native riparian tree establishment along the regulated Dolores River, Colorado. West N Am Nat 73:41–53
Cook BJ, Ault TR, Smerdon JE (2015) Unprecedented 21st century drought risk in the American Southwest and Central Plains. Sci Adv 1:e1400082. https://doi.org/10/1126/sciadv.1400082
Cooper DJ, Sanderson JS, Stannard DI, Groeneveld DP (2006) Effects of long-term water table drawdown on evapotranspiration and vegetation in an arid region phreatophyte community. J Hydrol 325:21–34
Cowley DE (2006) Strategies for ecological restoration of the Middle Rio Grande in New Mexico and recovery of the endangered Rio Grande silvery minnow. Rev Fish Sci 14:169–186
Crawford CS, Ellis LM, Molles Jr MC (1996) The Middle Rio Grande bosque: an endangered ecosystem. N M J Sci 36:277–299
Dahm CN, Baker MA, Moore DI, Thibault JR (2003) Coupled biogeochemical and hydrological responses of streams and rivers to drought. Freshw Biol 48:1219–1231
Dahm CN, Cleverly JR, Coonrod JEA, Thibault JR, McDonnell DE, Gilroy DJ (2002) Evapotranspiration at the land/water interface in a semi-arid drainage basin. Freshw Biol 47:831–843
Dello Russo G (2007) Opportunities for long-term bosque preservation in the San Acacia reach. In: Price LG, Johnson PS, Bland D (eds) Water resources of the Middle Rio Grande. San Acacia to Elephant Butte. New Mexico Bureau of Geology and Mineral Resources, Socorro, NM, pp 84–88
Dettinger M, Udall B, Georgakakos A (2015) Western water and climate change. Ecol Appl 25:2069–2093
Devitt DA, Sala A, Smith SD, Cleverly J, Shaulis LK, Hammett R (1998) Bowen ratio estimates of evapotranspiration on the Virgin River in southern Nevada. Water Resour Res 34:2407–2414
Di Tomaso JM (1998) Impact, biology, and ecology of saltcedar (Tamarix spp.) in the southwestern United States. Weed Technol 12:326–336
Eamus D, Froend R (2006) Groundwater-dependent ecosystems: the where, what and why of GDEs. Aust J Bot 54:91–96
Eamus D, Froend R, Loomes R, Hose G, Murray B (2006) A functional methodology for determining the groundwater regime needed to maintain the health of groundwater-dependent vegetation. Aust J Bot 54:97–114
Elias EH, Rango A, Steele CM, Mejia JF, Smith R (2015) Assessing climate change impacts on water availability of snowmelt-dominated basins of the Upper Rio Grande basin. J Hydrol 3:525–546
Everitt BL (1998) Chronology of the spread of tamarisk in the central Rio Grande. Wetlands 18:658–668
Famiglietti JS (2014) The global groundwater crisis. Nat Clim Change 4:945–948
Follstad Shah JJ, Dahm CN (2008) Flood regime and leaf fall determine soil inorganic nitrogen dynamics in semiarid riparian forests. Ecol Appl 18:771–788
Friedman JM, Auble GT, Shafroth PB, Scott ML, Merigliano MF, Freehling MD, Griffin ER (2005) Dominance of non-native riparian trees in western USA. Biol Invasions 7:747–751
Gries D, Zeng F, Foetzki A, Arndt S, Bruelheide H, Thomas F, Zhang X, Runge M (2003) Growth and water relations of Tamarix ramosissima and Populus euphratica on Taklamakan desert dunes in relation to depth to a permanent water table. Plant Cell Environ 26:725–736
Guttman NB (1991) A sensitivity analysis of the Palmer Hydrologic Drought Index. Water Resour Bull 27:797–807
Gutzler DS (2004) New Mexico’s changing climate. N M Earth Matters 4:1–4.
Harrington CA (1987) Responses of red alder and black cottonwood seedlings to flooding. Physiol Plant 69:35–48
Heim Jr RR (2002) A review of twentieth-century drought indices in the United States. Bull Am Meteorol Soc 83:1149–1165
Horton JL, Kolb TE, Hart SC (2001) Physiological responses to groundwater depth varies among species and with river flow regulation. Ecol Appl 11:1046–1059
Howe WH, Knopf FL (1991) On the imminent decline of Rio Grande cottonwoods in central New Mexico. Southwest Nat 36:218–224
Hultine KR, Bush SE (2011) Ecohydrological consequences of non-native riparian vegetation in the southwestern United States: a review from an ecophysiological perspective. Water Resour Res 47:W07542. https://doi.org/10.1029/2010WR010317
Hurd BH, Coonrod J (2012) Hydro-economic consequences of climate change in the upper Rio Grande. Clim Res 53:103–118
Katz GL, Shafroth PB (2003) Biology, ecology and management of Elaeagnus angustifolia L. (Russian olive) in western North America. Wetlands 23:763–777
Keyantash J, Dracup JA (2002) The quantification of drought: an evaluation of drought indices. Bull Am Meteorol Soc 83:1167–1180
Knopf FL, Olson TE (1984) Naturalization of Russian-Olive: implications to Rocky mountain wildlife. Wildl Soc B 12:289–298
Lautz LK (2008) Estimating groundwater evapotranspiration rates using diurnal water-table fluctuations in a semi-arid riparian zone. Hydrogeol J 16:483–497
Lite SJ, Stromberg JC (2005) Surface water and ground-water thresholds for maintaining Populus-Salix forests, San Pedro River, Arizona. Biol Conserv 125:153–167
Llewellyn DS, Vaddey S, Roach J, Pinson A (2013) West-wide climate risk assessment: Upper Rio Grande Impact Assessment. Executive Summary. US Bureau of Reclamation, Upper Colorado Region, Albuquerque Area Office
Loheide II SP (2008) A method for estimating subdaily evapotranspiration of shallow groundwater using diurnal water table fluctuations. Ecohydrology 1:59–66
Lukas J, Barsugli J, Doesken N, Rangwala I, Wolter K (2014) Climate change in Colorado: a synthesis to support water resources management and adaptation. A report for the Colorado Water Conservation Board. University of Colorado, Boulder, CO
Mahoney JM, Rood SB (1992) Response of a hybrid poplar to water-table decline in different substrates. Forest Ecol Manag 54:141–156
Mahoney JM, Rood SB (1998) Streamflow requirements for cottonwood seedling recruitment-an integrative model. Wetlands 18:634–645
Martinet MC, Vivoni ER, Cleverly JR, Thibault JR, Schuetz JF, Dahm CN (2009) On groundwater fluctuations, evapotranspiration, and understory removal in riparian corridors. Water Resour Res 45:W05425. https://doi.org/1029/2008WR007152
Massman W, Clement R (2004) Uncertainty in eddy covariance flux estimates resulting from spectral attenuation. In: Lee X, Massman W, Law B (eds) Handbook of micrometeorology: a guide for surface flux measurement and analysis. Kluwer Academic Publishers, Dordrecht, Boston, London, pp 67–100
McCabe GJ, Wolock DM (2007) Warming may create substantial water supply shortages in the Colorado River basin. Geophys Res Lett 34:L22708. https://doi.org/1029/2007GL031764
Merritt DM, Poff NL (2010) Shifting dominance of riparian Populus and Tamarix along gradients of flow alteration in western North American rivers. Ecol Appl 20:135–152
Molles Jr MC, Crawford CS, Ellis LM, Valett HM, Dahm CN (1998) Managed flooding for riparian ecosystem restoration. BioScience 48:749–756
Moore GW, Edgar CB, Vogel JG, Washington-Allen RA, March OG, Zehnder R (2016) Tree mortality from an exceptional drought spanning mesic to semiarid ecoregions. Ecol Appl 26:602–611
Moore GW, Owens MK (2012) Transpirational water loss in invaded and restored semiarid riparian forests. Restor Ecol 20:346–351
Mortenson SG, Weisberg PJ (2010) Does river regulation increase the dominance of invasive woody species in riparian landscapes? Global Ecol Biogeogr 19:562–574
Nagler PL, Hinojosa-Huerta O, Glenn EP, Garcia-Hernandez J, Romo R, Curtis C, Huete AR, Nelson SG (2005) Regeneration of native trees in the presence of invasive saltcedar in the Colorado River delta, Mexico. Conserv Biol 19:1842–1852
Naiman RJ, Décamps H, McClain ME (2005) Riparia: ecology, conservation, and management of streamside communities.. Elsevier Academic Press, Burlington, MA
New Mexico Interstate Stream Commission (2009) 2007–2008 Annual report. Office of the State Engineer, Santa Fe, NM
Newman BD, Wilcox BP, Archer SA, Breshears DD, Dahm CN, Duffy CJ, McDowell NG, Phillips FM, Scanlon BR, Vivoni ER (2006) Ecohydrology of water-limited environments: a scientific vision. Water Resour Res 42:W06302. https://doi.org/10.1029/2005WR004141
NOAA National Oceanic and Atmospheric Administration Climate Data Center. New Mexico Climate Divisions 2 and 5. http://www1.ncdc.noaa.gov/pub/data/cirs/climdiv/climdiv-phdidv-v1.0.0. Accessed 1 Aug 2016
Perry LG, Andersen DC, Reynolds LV, Nelson SM, Shafroth PB (2012) Vulnerability of riparian ecosystems to elevated CO2 and climate change in arid and semiarid western North America. Global Change Biol 18:821–842
Pockman WT, Sperry JS (2000) Vulnerability to xylem cavitation and the distribution of Sonoran desert vegetation. Am J Bot 87:1287–1299
Poff NL, Allan JD, Bain MB, Karr JR, Prestegaard KL, Richter BD, Sparks RE, Stromberg JC (1997) The natural flow regime. BioScience 47:769–784
Redmond KT (2002) The depiction of drought. Bull Am Meteorol Soc 83:1143–1147
Reynolds LV, Cooper DJ (2010) Environmental tolerance of an invasive riparian tree and its potential for continued spread in the southwestern US. J Veg Sci 21:733–743
Robinson TW (1958) Phreatophytes. US Geological Survey Water Supply Paper 1423
Rood SB, Gourley CR, Ammon EM, Heki LG, Klotz JR, Morrison ML, Mosley D, Scoppettone GG, Swanson S, Wagner PL (2003) Flows for floodplain forests: a successful riparian restoration. BioScience 647:647–656
Rood SB, Pan J, Gill KM, Franks CG, Samuelson GM, Shepherd A (2008) Declining summer flows of Rocky Mountain rivers: changing seasonal hydrology and probable impacts on floodplain forests. J Hydrol 349:397–410
Rood SB, Patino S, Coombs K, Tyree MT (2000) Branch sacrifice: cavitation-associated drought adaptation of riparian cottonwoods. Trees-Struct Funct 14:248–257
Rood SB, Samuelson GM, Braatne JH, Gourley CR, Hughes FMR, Mahoney JM (2005) Managing river flows to restore floodplain forests. Front Ecol Environ 3:193–201
SAS Institute (2015) Version 9.3. SAS Institute, Inc., Cary, NC
Schotanus P, Nieuwstadt FTM, Debruin HAR (1983) Temperature-measurement with a sonic anemometer and its application to heat and moisture fluxes. Bound-Layer Meteorol 26:81–93
Scott ML, Shafroth PB, Auble GT (1999) Responses of riparian cottonwoods to alluvial water table declines. Environ Manag 23:347–358
Segelquist CA, Scott ML, Auble GT (1993) Establishment of Populus deltoides under simulated alluvial groundwater declines. Am Midl Nat 130:274–285
Shafroth PB (2010) Background and introduction. In: Shafroth PB, Brown CA, Merritt DM (eds) Saltcedar and Russian olive control demonstration act science assessment. US Geological Survey Scientific Investigations Report 2009-5247, pp 1–6
Shafroth PB, Beauchamp VB, Briggs MK, Lair K, Scott ML, Sher AA (2008) Planning riparian restoration in the context of Tamarix control in western North America. Restor Ecol 16:97–112
Shafroth PB, Cleverly JR, Dudley TL, Taylor JP, Van Riper C, Weeks EP, Stuart JN (2005) Control of Tamarix in the Western United States: implications for water salvage, wildlife use, and riparian restoration. Environ Manag 35:231–246
Shafroth PB, Stromberg JC, Patten DT (2000) Woody riparian vegetation response to different alluvial water table regimes. West N Am Nat 60:66–76
Sher AA, Marshall DL (2003) Seedling competition between native Populus deltoides (Salicaceae) and exotic Tamarix ramosissima (Tamaricaceae) across water regimes and substrate types. Am J Bot 90:413–422
Smith DM, Finch DM (2016) Riparian trees and aridland streams of the southwestern United States: an assessment of the past, present, and future. J Arid Environ 135:120–131
Stella JC, Battles JJ, Orr BK, McBride JR (2006) Synchrony of seed dispersal, hydrology and local climate in a semi-arid river reach in California. Ecosystems 9:1200–1214
Stromberg JC (1997) Growth and survivorship of Fremont cottonwood, Goodding willow, and salt cedar seedlings after large floods in central Arizona. Gt Basin Nat 57:198–208
Stromberg JC (2001) Restoration of riparian vegetation in the south-western United States: importance of flow regimes and fluvial dynamism. J Arid Environ 49:17–34
Stromberg JC, Lite SJ, Marler R, Paradzick C, Shafroth PB, Shorrock D, White JM, White MS (2007) Altered stream-flow regimes and invasive plant species: the Tamarix case. Global Ecol Biogeogr 16:381–393
Stromberg JC, Lite SJ, Rychener TJ, Levick LR, Dixon MD, Watts JM (2006) Status of the riparian ecosystem in the upper San Pedro River, Arizona: application of an assessment model. Environ Monit Assess 115:145–173
Stromberg JC, Tiller R, Richter B (1996) Effects of groundwater decline on riparian vegetation of semiarid regions: the San Pedro, Arizona. Ecol Appl 6:113–131
Stull RB (1988) An introduction to boundary layer meteorology. Kluwer Academic Publishers, Boston, MA
Svoboda M, LeComte D, Hayes M, Heim R, Gleason K, Angel J, Rippey B, Tinker R, Palecki M, Stooksbury D, Miskus D, Stephens S (2002) The drought monitor. Bull Am Meteorol Soc 83:1181–1190
Taylor CJ, Alley WM (2002) Ground-water-level monitoring and the importance of long-term water-level data. US Geological Survey Circular 1217
Taylor JP, Smith LM, Haukos DA (2006) Evaluation of woody plant restoration in the Middle Rio Grande: ten years after. Wetlands 26:1151–1160
Udall B, Overpeck J (2017) The twenty-first century Colorado River hot drought and implications for the future. Water Resour Res 53:2404–2418
USGS United States Geological Survey National Water Information System: Web interface. www.waterdata.usgs.gov/nwis/uv/?site_no=08330000. Accessed 26 Aug 2016
Valett HM, Baker MA, Morrice JA, Crawford CS, Molles Jr MC, Dahm CN, Moyer DL, Thibault JR, Ellis LM (2005) Biogeochemical and metabolic responses to the flood pulse in a semiarid floodplain. Ecology 86:220–234
van Gestel NC, Dhungana N, Tissue DT, Zak JC (2016) Seasonal microbial and nutrient responses during a 5-year reduction in the daily temperature range of soil in a Chihuahuan Desert ecosystem. Oecologia 180:265–277
Vinson DS, Block SE, Crossey LJ, Dahm CN (2007) Biogeochemistry at the zone of intermittent saturation: field-based study of the shallow alluvial aquifer, Rio Grande, New Mexico. Geosphere 3:366–380
Vivoni ER, Bowman RS, Wyckoff RL, Jakubowski RT, Richards KE (2006) Analysis of a monsoon flood event in an ephemeral tributary and its downstream hydrologic effects. Water Resour Res 42:W03404. https://doi.org/10.1029/2005WR004036
Warren DK, Turner RM (1975) Saltcedar (Tamarix chinensis) seed production, seedling establishment, and response to inundation. J Arizona Acad Sci 10:135–144
Webb E, Pearman G, Leuning R (1980) Correction of flux measurements for density effects due to heat and water-vapor transfer. Q J Roy Meteor Soc 106:85–100
Webb RH, Leake SA (2006) Ground-water surface-water interactions and long-term change in riverine riparian vegetation in the southwestern United States. J Hydrol 320:302–323
Wesely ML (1970) Eddy correlation measurements in the atmospheric surface layer over agricultural crops. Dissertation, University of Wisconsin
Williams AP, Allen CD, Macalady AK, Griffin D, Woodhouse CA, Meko DM, Swetnam TW, Rauscher SA, Seager R, Grissino-Mayer HD, Dean JS, Cook ER, Gangodagamage C, Cai M, McDowell NG (2013) Temperature as a potent driver of regional forest drought stress and tree mortality. Nat Clim Change 3:292–297
Wolf S, Keenan TF, Fisher JB, Baldocchi DD, Desai AR, Richardson AD, Scott RL, Law BE, Litvak ME, Brunsell NA, Peters W, Van Der Laan-Luijkx IT (2016) Warm spring reduced carbon cycle impact of the 2012 US summer drought. Proc Natl Acad Sci USA 113:5880–5885
Woodhouse CA, Pederson GT, Morino K, McAfee SA, McCabe GJ (2016) Increasing influence of air temperature on upper Colorado River streamflow. Geophys Res Lett 43:2174–2181
Yarnell SM, Petts GE, Schmidt JC, Whipple AA, Beller EE, Dahm CN, Goodwin P, Viers JH (2015) Functional flows in modified riverscapes: hydrographs, habitats and opportunities. BioScience 65:963–972
Acknowledgements
The authors wish to thank the UNM Hydrogeoecology Group, in particular Julie Coonrod, Diane McDonnell, Jennifer Schuetz, Manuel Molles, Jr., John Craig, Stephen Teet, and David Gilroy for their assistance and insights over many years. The authors also thank the following agencies for granting access to the research sites: Sevilleta National Wildlife Refuge, Bosque del Apache National Wildlife Refuge, City of Albuquerque Open Space Division, the New Mexico State Land Office, the Middle Rio Grande Conservancy District, and New Mexico State Game and Fish. The authors also extend thanks to Salim Bawazir and New Mexico State University and to the Sevilleta Long Term Ecological Research program. Funding was provided by NASA Award NAG5-6999, the US Fish and Wildlife Bosque Initiative, the US Bureau of Reclamation Endangered Species Act research grant, the New Mexico Interstate Stream Commission, a National Science Foundation EPSCoR Research Infrastructure Improvement Award, and NSF DEB#0080529/DEB#0217774 to the Sevilleta LTER program. Finally, we thank two anonymous reviewers for improving this paper with their constructive, insightful inputs.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no competing interests.
Rights and permissions
About this article

Cite this article
Thibault, J.R., Cleverly, J.R. & Dahm, C.N. Long-term Water Table Monitoring of Rio Grande Riparian Ecosystems for Restoration Potential Amid Hydroclimatic Challenges. Environmental Management 60, 1101–1115 (2017). https://doi.org/10.1007/s00267-017-0945-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-017-0945-x