
Abstract
Among birds exhibiting conspecific brood parasitism (CBP), parasites demonstrate a variety of intriguing tactics for selecting a host nest, including preference for safe nests (i.e., avoiding nests depredated during the previous season). Brood parasites of birds that do not reuse nest sites, however, are limited to nest-site information available to them during the current breeding season. This study explores cues used by brood parasites in a population of red-breasted mergansers nesting in upland vegetation and sandy substrate that prohibits reuse of nest sites in consecutive years. Nest-site and host traits were measured for 33 parasitized and 23 non-parasitized nests. There was no association between CBP and nest-site traits potentially providing cues about nest safety from avian predators (concealment and density of nesting larids), likely due to very low rates of egg predation. Distance to shore for parasitized nests was slightly greater than for unparasitized nests. Parasites did not select nests in relation to host age, mass, date of nest initiation, or stage of nesting (laying or incubation). Artificial nests were used to simulate natural nests without a host and to assess whether host presence serves as a cue for parasites. The proportion of natural nests receiving ≥ 1 foreign egg (60% of 15 nests) was thrice that for artificial nests (21% of 14 nests). Some aspect of host presence may therefore be an important, but not necessary, cue for brood parasites targeting nests that are heavily concealed in uplands.
Significance statement
Whenever brood parasitism is a well-developed component of reproduction, selection is expected to favor brood parasites that make use of environmental or social information allowing them to discriminate among host nests and maximize reproductive success. We assessed cues used by brood parasites in ground-nesting red-breasted mergansers for which conspecific brood parasitism (CBP) is common, and nests are well concealed in upland vegetation. Observations at parasitized and unparasitized nests revealed that nest-site safety and visibility from habitat edges are not central to laying decisions by parasites in this population. Brood parasites, however, were considerably more likely to lay their eggs in natural nests with a host than in artificial nests without a host, suggesting that some aspect of host presence is important in the nest-site selection process for parasites. The ability of brood parasites to discriminate among nests based on host presence is expected to affect parasite success because individuals can avoid nests in which their eggs are doomed to fail (e.g., abandoned nests).
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Conspecific brood parasitism (CBP) is an alternative breeding strategy wherein a female lays her eggs in the nest of a second female (the host) of the same species. CBP occurs in birds, fish, and insects (Brockman 1993; Yom-Tov 2001; Zink 2003), and it is particularly well developed in waterfowl, among which at least 76 of 161 species use the breeding tactic (Yom-Tov 2001). Brood parasitism offers females an opportunity to gain fitness when nesting is not possible (Lyon and Eadie 2008) and allows nesting females to increase their reproductive output (Åhlund and Andersson 2001). The success of the parasitic tactic is nonetheless dependent on the fate of the host nest because the host provides parental care for the parasite’s eggs and young. Thus, there should be strong selection pressure for parasitic females to discriminate among available nests and to increase the probability of survival of their offspring (Lyon and Eadie 2017).
Brood parasites may select host females that are of highest quality. Females that achieve high reproductive success often nest early in the breeding season (Eadie 1989; Forslund and Pärt 1995; Forest and Gaston 1996; Blums and Clark 2004) and indeed, early-initiated nests are often parasitized more frequently than those initiated later (Clawson et al. 1979; Sorenson 1991; Petrželková et al. 2013). Females that are in good physical condition (i.e., with sufficient stored nutrients) are less likely to abandon their nest than females in poor condition (Blums et al. 1997). Success of parasitic eggs may therefore be maximized when they are laid in early nests of hosts in good condition.
Parasites can make fine-scale assessments of available nests and select those that have a high probability of being successful (Lyon and Everding 1996; Pöysä 2006). Parasitic cliff swallows (Hirundo pyrrhonota) target nests containing few ectoparasites, and they sometimes transfer their eggs to nests that obtain high reproductive success (Brown and Brown 1988, 1991). The nest-searching tactics of brood parasites may also reflect the importance of minimizing predation of offspring. For example, common goldeneye (Bucephala clangula) and Barrow’s goldeneye (Bucephala islandica) parasites select nest boxes that are not depredated during the previous breeding season when risk of predation is predictable (Pöysä 1999a, 2006; Pöysä and Pesonen 2007). The ease in finding a host nest, however, may be more important than selecting safe nests for parasites of other birds. Brood-parasitic common eiders (Somateria mollissima) target highly visible nest shelters, despite shelter nests having lower nest success than those in dense vegetation (Lusignan et al. 2010). Likewise, high visibility of nest sites from habitat edges is an important correlate of CBP in cavity-nesting species (Heusmann et al. 1980; Andersson and Eriksson 1982; Semel and Sherman 1986).
CBP occurs in birds that nest on the ground in upland habitat (reviewed in Sayler 1992; Eadie and Savard 2015). However, unlike some nests in cavities and in open habitat, those in uplands are often concealed by vegetation and not seen from habitat edges. Locating concealed nests by searching randomly through habitat is likely not an efficient tactic (Weller 1959). Rather, brood parasites could use cues associated with the presence of a host, such as nest visits by the host, for finding and selecting a nest (Weller 1959; Sayler 1992). If host activity is an important cue for parasites, unattended nests (i.e., abandoned) are not expected to be parasitized (Pöysä et al. 2014). Despite this, little is known about cues used by parasites in uplands, so we still do not fully understand the full range of cognitive abilities of brood parasites across the different ecological contexts in which CBP occurs.
We examined the behavior of brood-parasitic red-breasted mergansers (Mergus serrator) in upland nesting habitat at Kouchibouguac National Park, New Brunswick, Canada. Here, red-breasted mergansers exhibit high rates of CBP (34–60% nests annually; Young and Titman 1988; Craik et al. 2018). Nest sites are located up to 40 m from shore and in dense stands of grass, and they are concealed under a dome of old and new vegetative growth (Craik and Titman 2009). Thus, it is unlikely that brood parasites are readily able to detect a nest from a distance. Unlike birds that reuse nest sites, parasitic red-breasted mergansers do not have access to information from previous breeding attempts because scouring of substrate by ice and shifts in the distribution of vegetation preclude the reuse of nest bowls in consecutive years. Red-breasted Mergansers at this site form a nesting association with a large colony of common terns (Sterna hirundo), which actively defend their nests (and indirectly those of mergansers) against avian egg predators (Young and Titman 1986).
The objective of our study was to identify cues used by parasitic red-breasted mergansers in finding and selecting a nest. Specifically, we examined whether parasites use (a) host presence, (b) nest-site characteristics reflecting level of risk to egg predation and visibility of the nest (e.g., concealment, distance to shore), (c) density of red-breasted merganser nests, or (d) host age, body mass, date of nest initiation, and stage of nesting (laying or incubation) in their nest-site selection decisions.
Methods
Study site
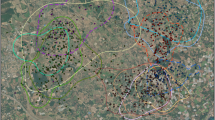
We studied red-breasted mergansers nesting on Tern Islands, a 3-ha archipelago in the Saint-Louis Lagoon of Kouchibouguac National Park (Fig. 1). The islands are composed of sand stabilized by marram grass (Ammophila breviligulata) and sea lyme grass (Leymus mollis) and are protected from the Northumberland Strait by the South Kouchibouguac Dune (Thimot 2018). Tern Islands support the largest common tern (Sterna hirundo) colony in Canada (annually ~ 6000 pairs). The majority of common tern nests are initiated about a week before the earliest red-breasted merganser nests (SRC, unpubl. data). Common terns aggressively mob avian egg predators, notably common ravens (Corvus corax), American crows (Corvus brachyrhynchos), and great blacked-backed gulls (Larus marinus; Young and Titman 1986). Mammalian egg predators do not have access to the islands during the breeding season.

Tern Islands at Kouchibouguac National Park, New Brunswick. Regions in light gray represent intertidal flats
Nest monitoring
It was not possible to record data blind because our study involved focal animals in the field. We monitored nests from late May to mid-August during 2014–2016. In each year, we systematically searched the vegetation to find nests. Nest searching was limited to 2 h per day to reduce disturbance to breeding birds. Nests were marked with a numbered wooden lath placed 1 m north of the bowl, and coordinates of each nest were recorded using a global positioning system with an error of ≤ 5 m (Etrex, Garmin Ltd.). Nests were visited about every other day during the laying period and every ~ 6 days during incubation. During each nest visit, we recorded egg status (warm/cold) and noted any general differences in egg color and shape within a clutch (Young and Titman 1988). We floated 1–2 eggs in a container filled with sea water to determine hatch date based on a 30-day incubation period (Westerskov 1950; Craik et al. 2015). A nest was considered parasitized when the laying rate exceeded 1 egg per 1.5 day (i.e., the laying rate for a red-breasted merganser; Palmer 1976; Craik et al. 2015), or if eggs were added after the onset of incubation (Young and Titman 1988). Variation in egg color and shape within parasitized nests was generally greater than that in non-parasitized nests. To identify parasitism as accurately as possible, we only considered active nests discovered during the earliest stages of egg laying (≤ 4 eggs; median 2 eggs) and that reached incubation. We omitted nests abandoned during egg laying because they were typically abandoned upon discovery, and thus, it was difficult to assess parasitism status. We may have missed some nest parasitism at the beginning of the host laying cycle prior to nest discovery. Analyses of DNA microsatellites from a sample of nests nevertheless indicated that our field method for detecting CBP was accurate; parasitism status of 9 of 10 nests were classified similarly by genetic and field data (SRC, unpubl. data).
Nest-site traits
Red-breasted mergansers access nests by walking from shore through vegetation (Young and Titman 1986) or by flying over the islands and subsequently landing at or near nesting habitat. We thus measured five nest-site traits that could affect decisions by parasites attempting to find a host nest by either walking through vegetation or flying over nesting habitat: red-breasted merganser nest density, common tern nest density, distance to shore, overhead concealment, and lateral concealment. Red-breasted merganser nest density was measured by plotting the coordinates of all merganser nests on a map of Tern Islands and counting the number of conspecific nests within a 20-m radius of each nest. Common tern nest density was calculated as the number of tern nests within 4 m of a red-breasted merganser nest. Distance to shore was measured as the shortest distance from a nest to the spring high-tide level (wrack line). Overhead concealment was estimated by placing a black disc with five 6.5 cm2 white squares over the nest bowl and estimating the percentage of the white squares that was covered by vegetation to the nearest 10% (Traylor et al. 2004). The disc’s diameter was that of a typical red-breasted merganser nest (29.5 cm; Craik et al. 2015). We used the number of nest entrances (i.e., tunnels in vegetation used by females to access the nest) as an index of lateral concealment; nests with many entrances were the least concealed. Nest-site characteristics were measured during each of the 3 years, except for density of common tern nests and overhead concealment, which were measured in 2015 and 2016 only.
Host traits
In each year since 2002, female red-breasted mergansers were captured at their nest with a dip net or automated nest trap (Weller 1957) and issued a standard metal leg band. Captures occurred during the final week of incubation. During this study (2014–2016), ~ 70% of captured hosts had been banded in a previous year. We estimated age (experience) of the host by considering unbanded females as 2 years of age (Craik et al. 2015). The age of previously captured females was estimated as two plus the number of years since their first capture. We weighed (± 5 g) females with a Pesola scale. To minimize the chance a host would abandon her nest, we avoided the capture of females during egg laying and early incubation. Nonetheless, body mass during late incubation has been shown to be similar to that during early incubation for a number of waterfowl populations (e.g., Hepp et al. 2005), including for some hooded mergansers Lophodytes cucullatus (Sayler 1992; Dugger et al. 2009), so our measure of host body mass during late incubation may not have been considerably different to host body mass earlier during the nesting cycle, when most parasitism occurred. We estimated dates of nest initiation by backdating, assuming one egg is laid every 1.5 day (Craik et al. 2015). Nest initiation dates were standardized to account for differences in nesting schedule among the 3 years.
Host presence
We made artificial red-breasted merganser nests to examine whether host presence is a cue for nest selection by parasites. Fourteen artificial nests were established on Tern Islands from 3 to 18 June 2016. This date range coincided with the greatest egg-laying activity, and so it ensured that exposure time of artificial nests to parasites was similar to that of most natural nests. At least one active natural nest was located within 15 m of each artificial nest. We chose where to place artificial nests by selecting a random distance (10 m, 20 m, 30 m, or 40 m) and direction (N, E, W, S, SE, SW, NE, or NW) from a randomly selected active nest. We chose the nearest site from that location where habitat was considered suitable based on nest-site requirements for red-breasted mergansers at the study site (Craik and Titman 2009). Artificial nests were created in dense stands of marram grass by digging bowls in the sand that had identical dimensions as those of a typical red-breasted merganser nest at the site: ~ 7 cm deep with an inner diameter of ~ 20 cm and an outside diameter of ~ 30 cm (Craik et al. 2015). Vegetation was removed from inside the bowl. If there was no natural entrance leading into the nest (i.e., an area void of vegetation), 1–2 openings were created to adjacent open areas to provide nest access to prospecting females. Natural nests typically have 1–3 openings (Thimot 2018).
In 11 artificial nests, we placed three large chicken eggs that were painted with glossy, buff-colored paint to resemble red-breasted merganser eggs. In each of the other three artificial nests, we placed three red-breasted merganser eggs collected from early-abandoned clutches. We chose three artificial eggs because brood parasites may be more likely to lay in nests with few eggs (Odell and Eadie 2010). Mean chicken egg length was 58.4 mm (range 55.1–62.6 mm) and mean width was 44.7 mm (range 43.7–45.6 mm). Mean red-breasted merganser egg length was 63.9 mm (range 58.4–65.4 mm), and mean width was 44.3 mm (range 35.2–46.9 mm). Thus, chicken eggs were similar in size to many red-breasted merganser eggs. Artificial nests were visited every 3–6 days throughout the nesting season.
Statistical analyses
We combined data from all 3 years because samples sizes were limited in each year. Seven females were each captured during 2 years. We thus used generalized linear mixed models (GLMMs) to identify characteristics that differed between parasitized and non-parasitized nests. Parasitism status was a binary response variable (0 = non-parasitized; 1 = parasitized). For fixed effects, we considered the three host traits (date of nest initiation, age, body mass) and five nest-site characteristics (distance to shore, red-breasted merganser nest density, common tern nest density, and overhead and lateral concealment). Female identity was treated as a random effect. We performed univariate GLMMs to assess the level of association of each fixed effect with parasitism status (parasitized or non-parasitized), and the magnitude of effect for each variable was determined with a likelihood ratio test (LRT). Overhead and lateral concealment, common tern nest density, and host age and body mass for parasitized nests were similar to non-parasitized nests (Table 1).
A model set was generated by first fitting a global GLMM that included date of nest initiation, distance to shore, red-breasted merganser nest density, and lateral concealment. Ideally, the model set would have included all eight fixed effects, but this was not possible given that overhead concealment and common tern nest density were not studied in 2014 (i.e., 41% of nests lacked data for these two variables) and host age and body mass were unknown for 16 nests. Our model set nonetheless included those variables of interest in potentially discriminating between parasitized and non-parasitized nests, given the poor fit of overhead concealment, common tern nest density, and host age and body mass (Table 1). Next, we generated a set of 16 models from the global model and that contained all combinations of single and additive fixed effects, and an intercept-only model with random effects (Grueber et al. 2011). Fixed effects were standardized to a mean of 0 and a SD of 0.5 prior to model analyses (Gelman 2008). We examined the level of collinearity of fixed effects by calculating variance inflation factors (VIF). All VIFs were < 2.1, indicating little collinearity among variables.
We used an information-theoretic approach, based on Akaike’s information criterion (corrected for small sample size; AICc), to select among the 16 competing models (Akaike 1973; Burnham and Anderson 2002). Models were fit using a logit link function. Selection of the best approximating model(s) was based on the values of ΔAICc, calculated as the difference in values of AICc between the model of interest and the model with the lowest AICc value (Burnham and Anderson 2002). For each model, we calculated its weight (w); models with larger weights better approximate the data. The potential of each fixed effect to discriminate between parasitized and non-parasitized nests was assessed in two ways. First, we summed weights of all models with a particular fixed effect to assess its relative importance; the larger the sum of model weights, the more important the variable is relative to other fixed effects (Burnham and Anderson 2002). Second, parameter estimates and their standard errors were obtained from the best approximating models (ΔAICc < 2; Burnham and Anderson 2002). For each parameter, we model-averaged slopes and generated unconditional standard errors and 95% confidence intervals to account for model uncertainty. We concluded lack of fit of fixed effects when their 95% confidence interval included zero.
We examined whether parasites non-randomly choose nests based on (1) date of nest initiation by hosts and (2) stage of nesting by hosts (egg laying versus incubation) at the time a parasitic egg was laid. To accomplish this, randomization tests were used to examine whether observed patterns of CBP deviated from random expectation (Emlen and Wrege 1986). For dates of nest initiation, we used a sample of 47 parasitic eggs in which date of laying was known. All eggs were assumed to have been laid by different females because we were unable to associate parasitic eggs to particular individuals. For each parasitic egg, we calculated the observed difference between the (standardized) date of nest initiation of the chosen nest and the median date of nest initiation of all nests available to the parasite at the time the egg was laid. A simulation was then performed in which a host nest was chosen at random from the pool of hosts available to the parasite. We calculated the difference between the date of nest initiation of the randomly chosen nest and the median date of initiation for all available nests. The process was repeated 1000 times to yield a distribution of nest initiation dates, and from which we estimated the probability that the differences in initiation dates between chosen and available nests occurred by chance.
For timing of CBP relative to host nesting stage, we identified a sub-sample of the 47 parasitic eggs (n = 26) for which a parasite had the option of laying their egg in ≥ 1 host nest in each of the laying and incubation stages (Lyon 1993a). For each parasitic egg, we chose a host nest at random from the pool of host nests available at the time the egg was laid. We calculated the percent of randomly chosen host nests in the laying stage (out of the 26 parasitic eggs). The process was performed 1000 times to produce a distribution of percent choices in the laying stage, from which we estimated the probability that the observed percentage of eggs laid in nests during the laying stage occurred by chance. We would have missed CBP that occurred prior to nest discovery and between the last visit in which a nest was in the laying stage and the subsequent visit. This would have made it more difficult to detect non-random patterns of nest selection, so any non-random effects that were detected were likely real.
A Fisher’s exact test was computed to compare the occurrence of parasitism between artificial and natural red-breasted merganser nests. We set significance levels at P < 0.05. Statistical analyses were performed with software R 3.4.1 (R Core Team 2017).
Data availability
Datasets analyzed in the current study are available from the corresponding author on reasonable request.
Results
Rates of CBP and nesting synchrony
We found a total of 44 nests in 2014 of which 22 were used to assess parasitism, 31 nests in 2015 of which 19 were used, and 38 nests in 2016 of which 15 were used. Nearly 60% of nests were parasitized each year (Table 2). Of the 33 parasitized nests, daily laying rate exceeding that for a red-breasted merganser (1 egg per 1.5 day) was detected at 32 nests and the addition of an egg following the onset of incubation was detected at two nests (n = 3 eggs in total). Nest initiation was synchronous in each year as nearly 65% of nests were initiated within the first 2 weeks of the breeding season (Fig. 2).

Standardized dates of nest initiation for parasitized (n = 33) and non-parasitized (n = 23) red-breasted merganser nests
Nest-site and host traits
There was some uncertainty in model selection because two models containing fixed effects on CBP probabilities were plausible (ΔAICc < 2; Table 3). Three lines of evidence indicated that date of nest initiation and distance to shore were predictors of nest parasitism, whereas there was little support for red-breasted merganser nest density and lateral concealment. First, nest initiation date and distance to shore were the only variables to be included in both of the top models, which together represented 0.75 of total model weight. Second, the sum of model weights for date of nest initiation (1.0) and distance to shore (0.96) were considerably higher than for red-breasted merganser nest density (0.52) and lateral concealment (0.23). Finally, model-averaged parameter estimates showed that the probability of CBP decreased with later dates of nest initiation (β = − 5.3, 95% CI = − 8.4 to − 2.2) and increased with greater distance between a nest and shore (β = 2.6, 95% CI = 0.5 to 4.7; Fig. 3a). Parasitized nests were initiated on average 10 days earlier than non-parasitized nests (Fig. 2). Red-breasted merganser nest density had poor fit in the top model (β = 0.8, 95% CI = − 1.2 to 2.7); thus, there was unlikely any real difference in nearby nest density between parasitized and unparasitized nests (Fig. 3b).

Boxplots showing summary statistics for a distance (m) from a nest to shore and b number of conspecific nests within 20 m of non-parasitized (n = 23) and parasitized (n = 33) red-breasted merganser nests. The dark horizontal line represents the median, and the horizontal lines below and above the median represent the first and third quartiles, respectively. The lower vertical line projecting downwards from the first quartile extends to the smallest value that is no more than 1.5 * the distance between the first and third quartiles. The upper vertical line projecting upwards from the third quartile extends to the largest value that is no more than 1.5 * the distance between the first and third quartiles
Randomization tests revealed that parasitic red-breasted mergansers chose nests at random with respect to a host’s date of nest initiation from those available to them at the time they laid the parasitic egg (two-tailed test; P = 0.68). Twenty-four of 26 (92%) parasitic eggs were deposited into host nests during the laying stage, but this pattern did not deviate from random expectation (two-tailed test; P = 0.12), at least in part due to the availability of very few incubated nests during time of parasitic egg laying.
Host presence
The proportion of natural nests receiving ≥ 1 foreign egg (60% of 15 nests) was thrice that for artificial nests (21% of 14 nests), although the difference in rates of nest parasitism between natural and artificial nests was marginally insignificant (Fisher’s exact test; P = 0.06). Of the three artificial nests receiving ≥ 1 foreign egg, two contained experimental red-breasted merganser eggs and one nest contained chicken eggs. Two of these artificial nests were incubated (n = 9 and 10 eggs). The large number of foreign eggs laid in each of these two nests suggests that they were added by one female only (i.e., possible nest takeovers). Since we were interested in CBP, we performed a second analysis in which we only considered nests that were clearly parasitized. Rate of CBP at artificial nests (1 of 12 nests) was lower than that at natural nests (Fisher’s exact test; P = 0.01).
Discussion
Our study examined the role of nest-site and host traits and host presence as cues used by a brood-parasitic bird for finding and selecting a nest. We found little evidence that nest-site characteristics related to nest-site safety (e.g., concealment), conspecific nest density, or visibility from habitat edges play a central role in decisions made by brood-parasitic red-breasted mergansers choosing host nests in upland vegetation. Rather, host presence, seasonal timing of nest initiation, and distance to shore were correlates of CBP. A key assumption is that our observations reflect patterns of nest choice by parasites rather than the ability of some hosts to defend their nest against brood parasitism. Red-breasted mergansers spend no more than about 10% of the egg-laying period at their nest (i.e., when almost all CBP occurs) and only rarely do females react aggressively to another individual at the nest (SRC, unpubl. data). Thus, the patterns of CBP observed in our study reflected, at least in part, decisions made by parasites (Pöysä et al. 2014).
Host presence and timing of breeding
Host activity may serve as a cue for parasites selecting nests in concealed upland habitat (Weller 1959). Our experimental nest study assessed the role of host presence in parasite decisions by comparing the occurrence of CBP at natural nests with a host to artificial nests without a host. Three artificial nests received ≥ 1 foreign red-breasted merganser egg, so at least some females can find and will lay eggs in nests that lack host activity, as is the case with some other waterfowl (e.g., Odell and Eadie 2010). However, artificial nests were three times less likely to receive a foreign egg (three of 14 nests) than natural nests (nine of 15 nests). Furthermore, only one of the 14 artificial nests was clearly parasitized because for each of the other two artificial nests receiving foreign eggs, large clutches were added and incubated, suggesting that they were nest takeover events rather than CBP. The ability of parasites to discriminate among nests based on host presence is expected to affect parasite success because parasites can avoid nests in which their eggs are doomed to fail (e.g., abandoned nests). Natural red-breasted merganser nests on Tern Islands also rarely receive parasitic eggs after a period of inactivity, despite naturally high rates of CBP in the population (up to 60% of nests/year; Craik et al. 2018). The much higher rate of nest parasitism in active, natural nests coupled with the seemingly inefficient tactic of randomly searching for nests throughout large tracts of habitat are consistent with the hypothesis that host behavior is an important cue for parasitic red-breasted mergansers, as may be the case with other birds nesting in uplands (Weller 1959). Equally consistent with these observations, however, is the thesis that parasites can find nests and avoid those without a host. Clearly, field observations of host and parasite behavior are needed to identify aspects of host presence that serve as proximate cues for brood parasites.
Considering nests available to a parasite at the time a parasitic egg was laid, patterns of nest choice with respect to a host’s date of nest initiation and stage of host nesting (laying vs incubation) did not deviate from random expectation. Parasites tended to lay their eggs early in the season, a period when synchrony in nest initiation was greatest. Accordingly, parasites had little choice other than to lay their eggs in some of the earliest-initiated nests, most of which had yet to reach incubation. Synchronizing the laying of parasitic eggs with egg laying by hosts is crucial in waterfowl because broods leave the nest shortly after hatching; any parasitic eggs laid after the onset of incubation may not be incubated to full term (Sayler 1992). Although parasitic red-breasted mergansers did not actively select nests in which the host was in the laying cycle, the overall trend for parasites to target host nests early in the season led to most parasitic eggs (26 of 28) being laid during the host laying stage.
The high rates of CBP observed early in the season are consistent with previous reports for red-breasted mergansers on Tern Islands (Young and Titman 1988; Craik and Titman 2009) and for many other waterfowl (Clawson et al. 1979; Dow and Fredga 1984; Sorenson 1991; Robertson et al. 1992; Pöysä 1999b; Paasivaara et al. 2010). Parasites may have an easier time finding nests early in the season when new vegetation around nests is limited (Sayler 1992). Although the overall density of marram grass on Tern Islands early in the season is lower than later on, red-breasted mergansers place their nests under a concealed dome of old growth (Craik and Titman 2009), so it is unlikely that the amount lateral concealment at a nest varies considerably across the season. Individual variability in nesting chronology has also been linked to the experience and/or quality of nesting females. Late nesting hens may be in relatively poor condition or are inexperienced, and generally have reduced reproductive success in comparison to earlier nesting females with more experience (Weller 1959; Spurr and Milne 1976; Dow and Fredga 1984; Devries et al. 2008). Our study did not reveal any association between timing of nest initiation and host age or body mass (see below), so other factors must underlie the tendency for brood parasites to lay their eggs in the earliest-initiated nests. A female may lay parasitically prior to initiating her own nest (Heusmann et al. 1980; Semel and Sherman 1986; Eadie 1989; Sorenson 1991; Lyon 1993b; Andersson and Åhlund 2001; Reichart et al. 2010; Lyon and Eadie 2017) as a way to increase reproductive output compared to parasite-only and non-parasitic nesting strategies (Andersson and Åhlund 2001). Alternatively, the greater tendency for early-initiated nests to be parasitized may reflect limited nest-site availability (Sayler 1985; McRae 1997) or a greater availability of hosts early on (Lyon 2003).
The highest rates of nest parasitism for red-breasted mergansers on Tern Islands coincide with a period of relatively frequent nest abandonment. During their 2-year study on Tern Islands, Craik and Titman (2009) found that abandonment of nests initiated during the first 2 weeks of the season was about 30% higher than nests initiated later on. Some early nests are most likely abandoned by the host in response to heavy parasitism during the egg-laying period. Indeed, Craik et al. (2018) showed that red-breasted merganser nests receiving ≥ 2 experimental parasitic eggs in a single day were more likely to be abandoned than unparasitized control nests, suggesting that parasitism can be costly. Heavily parasitized nests, however, represented < 25–30% of all parasitized nests (Craik et al. 2018). When all parasitized nests are considered, nests reaching the incubation stage are just as likely to be parasitized as those abandoned during egg laying (NJT, unpubl. data), and nest success and hatching success at parasitized nests are similar to unparasitized nests (Craik et al. 2018). These observations, coupled with those of timely laying of parasitic eggs (i.e., most foreign eggs laid before host incubation), show that CBP has the potential of being a successful mean by which red-breasted mergansers can gain fitness.
Nest-site and host traits
The safety of a nest is among the most important evolutionary drivers of nest-site selection behavior (Ricklefs 1969), and evidence suggests that safety from predators also shapes decisions made by brood parasites (see Pöysä et al. 2014). For birds having the option of reusing nest sites annually (e.g., cavity nesters), brood parasites can maximize their fitness by targeting nests that were not depredated the previous season (Pöysä 1999a), provided that nest success is predictable across years (Pöysä 2006). Females gain information on nest success by prospecting nests immediately following the previous season (Pöysä 2006). In contrast, parasites of birds that do not reuse nests annually, such as red-breasted mergansers in our study, only have access to nest-specific cues available to them while prospecting during the current season. We failed to find an association between CBP and nest-site variables potentially providing cues about the level of nest safety from avian predators, notably vegetative concealment and nearby nest density of common terns. On Tern Islands, the level of overhead and lateral concealment at unsuccessful nests is similar to successful nests (Craik and Titman 2009), so there is likely little selective pressure for parasites to choose host nests that are relatively well concealed. The equally high number of common tern nests surrounding parasitized and unparasitized nests (~ 10 tern nests within 4 m) was reflected in part by the widespread distribution of tern nests across the islands; in other words, nest sites for red-breasted mergansers were always in close proximity to many tern nests (Thimot 2018). Rates of nest predation for red-breasted mergansers on Tern Islands (< 5% nests) are significantly lower than those for merganser nests on islands without terns in the Kouchibouguac region (30–40%), at least partially because terns defend their own nests by driving gulls and corvids away from the islands (Young and Titman 1986). Red-breasted mergansers gain indirect fitness advantages from the anti-predator behavior of common terns (i.e., little egg predation), and regardless of where they nest on the islands. The nesting environment at our study site may thus be so safe in terms of nest depredation risk that selection between a risky and safe nest site may not be an option for parasites. This may also be the case for other waterfowl nesting with large groups of larids.
Waterfowl nests that are located close to shore or that are otherwise highly visible are often more likely to be parasitized than nests farther inland or that are more concealed (Giroux 1981; Sayler 1985; Eadie 1989; Pöysä et al. 1999). These observations support the premise that the ease by which nests can be detected from a distance is central in the nest-site selection process of brood parasites (Payne 1977; Semel et al. 1988; Lusignan et al. 2010). We found that parasitized red-breasted merganser nests were on average slightly farther from shore than non-parasitized nests. The high degree of lateral concealment around nests may have acted as a screen and prevented brood parasites from detecting even the closest nests from shore or other habitat edges (Young and Titman 1986). Indeed, a number of nests within a few meters of shore in each year were not parasitized. The distribution of parasitized nests, however, may reflect the availability of nests during periods of peak prospecting by parasites (Eadie and Savard 2015). Brood-parasitic red-breasted mergansers often targeted the earliest-initiated nests, but the distance between a nest and water was not correlated with timing of breeding (see “Methods”). Thus, selection of nests far from shore did not reflect a greater availability of inland nests than nests closer to water early in the season. It is unclear why parasites at our study site would select nests relatively far from shore. Parasitized nests were on average only 2–3 m farther from shore than unparasitized nests. Although this difference was statistically significant, we question whether it actually reflects parasites that prefer to target inland nests on small islands.
Host experience and condition were not cues for parasitic red-breasted mergansers. Host quality has seldom been tested as a cue for brood-parasitic birds (Brown and Brown 1991; Lyon 1993a; Pöysä et al. 2014). Two recent waterfowl studies have shown that host quality is not strongly correlated with parasite decisions (Paasivaara et al. 2010; Waldeck et al. 2011). Brood parasites may be unable to identify fine-scale differences in host quality (Åhlund 2005). Success of red-breasted merganser nests on Tern Islands is generally high (e.g., 60–70%; Craik and Titman 2009), and regardless of host age or body mass (NJT, unpubl. data). Thus, even if host age and condition can be assessed by parasites, they may not be relevant cues for females looking to maximize their fitness on Tern Islands. Nevertheless, we caution that host characteristics other than body mass and age may be important to brood parasite decisions. For waterfowl and other birds in which females demonstrate natal philopatry, individuals have an opportunity to lay eggs in the nest of close kin (Andersson 1984, 2017). Indeed, brood parasites in some waterfowl select nests of kin more often than by chance (e.g., Jaatinen et al. 2009), possibly because host and parasite can recognize each other (Andersson et al. 2019).
Our study supports the hypothesis that at least some birds exhibiting CBP use cues relating to host presence to find and select a nest to parasitize. However, unlike for some other waterfowl, there is little selective advantage for parasitic red-breasted mergansers to choose host nests based on nest-site traits potentially affording protection from avian predators (e.g., nest concealment), due at least in part to very low rates of egg predation at our study location.
References
Åhlund M (2005) Behavioural tactics at nest visits differ between parasites and hosts in a brood-parasitic duck. Anim Behav 70:433–440
Åhlund M, Andersson M (2001) Female ducks can double their reproduction. Nature 414:600–601
Akaike H (1973) Information theory as an extension of the maximum likelihood principle. In: Petrov BN, Csaki F (eds) Second international symposium on information theory. Akademiai Kiado, Budapest, pp 267–281
Andersson M (1984) Brood parasitism within species. In: Barnard CJ (ed) Producers and scroungers. Strategies of exploitation and parasitism. Chapman and Hall, New York, pp 195–228
Andersson M (2017) Female-biased natal philopatry, social parallels, and conspecific brood parasitism in New World quails and waterfowl. Auk 135:25–28
Andersson M, Åhlund M (2001) Protein fingerprinting: a new technique reveals extensive conspecific brood parasitism. Ecology 82:1433–1442
Andersson M, Eriksson MOG (1982) Nest parasitism in goldeneyes Bucephala clangula: some evolutionary aspects. Am Nat 120:1–16
Andersson M, Åhlund M, Waldeck P (2019) Brood parasitism, relatedness and sociality: a kinship role in female reproductive tactics. Biol Rev 94:307–327
Blums P, Clark RG (2004) Correlates of lifetime reproductive success in three species of European ducks. Oecologia 140:61–67
Blums P, Mednis A, Clark RG (1997) Effect of incubation body mass on reproductive success and survival of two European diving ducks: a test of the nutrient limitation hypothesis. Condor 99:916–925
Brockman HJ (1993) Parasitizing conspecifics: comparisons between hymenoptera and birds. Trends Ecol Evol 8:2–4
Brown CR, Brown MB (1988) A new form of reproductive parasitism in cliff swallows. Nature 331:66–68
Brown CR, Brown MB (1991) Selection of high-quality host nests by parasitic cliff swallows. Anim Behav 41:457–465
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer, Berlin
Clawson EL, Hartman GW, Fredrickson LH (1979) Dump nesting in a Missouri wood duck population. J Wildl Manag 43:347–355
Craik SR, Titman RD (2009) Nesting ecology of red-breasted mergansers in a common tern colony in eastern New Brunswick. Waterbirds 32:282–292
Craik S, Pearce J, Titman R (2015) Red-breasted merganser (Mergus serrator). In: Poole AF (ed) The birds of North America online. Cornell Lab of Ornithology, Ithaca. https://doi.org/10.2173/bna.443
Craik SR, Titman RD, Savard J-PL, Kaouass M, Thimot N, Elliott KH, Tremblay É (2018) Costs and response to conspecific brood parasitism by colonial red-breasted mergansers. J Ethol 36:251–258
Devries JH, Brook RW, Howerter DW, Anderson MG (2008) Effects of spring body condition and age on reproduction in mallards (Anas platyrhynchos). Auk 125:618–628
Dow H, Fredga S (1984) Factors affecting reproductive output of the goldeneye duck Bucephala clangula. J Anim Ecol 53:679–692
Dugger BD, Dugger KM, Fredrickson LH (2009) Hooded Merganser (Lophodytes cucullatus). In: Poole AF (ed) The birds of North America online. Cornell Lab of Ornithology, Ithaca. https://doi.org/10.2173/bna.98
Eadie JM (1989) Alternative reproductive tactics in a precocial bird: the ecology and evolution of brood parasitism in goldeneyes. PhD Dissertation, University of British Columbia
Eadie JM, Savard J-PL (2015) Breeding systems, spacing behavior, and reproductive behavior of sea ducks. In: Savard JPL, Derksen DV, Esler D, Eadie JM (eds) Ecology and conservation of north American sea ducks. CRC Press, Boca Raton, FL, pp 365–415
Emlen ST, Wrege PH (1986) Forced copulations and intra-specific parasitism: two costs of social living in the white-fronted bee-eater. Ethology 71:2–29
Forest LN, Gaston AJ (1996) The effect of age on timing of breeding and reproductive success in the thick-billed murre. Ecology 77:1501–1511
Forslund P, Pärt T (1995) Age and reproduction in birds – hypotheses and tests. Trends Ecol Evol 10:374–378
Gelman A (2008) Scaling regression inputs by dividing by two standard deviations. Stat Med 27:2865–2873
Giroux J-F (1981) Interspecific nest parasitism by redheads on islands in southeastern Alberta. Can J Zool 59:2053–2057
Grueber CE, Nakagawa S, Laws RJ, Jamieson IG (2011) Multimodel inference in ecology and evolution: challenges and solutions. J Evol Biol 24:699–711
Hepp GR, Folk TH, Manlove CA (2005) Temperature, incubation period, and investment decisions of incubating wood ducks Aix sponsa. J Avian Biol 36:523–530
Heusmann HW, Bellville RH, Burrell RG (1980) Further observations on dump nesting by wood ducks. J Wildl Manag 44:908–915
Jaatinen K, Jaari S, O’Hara RB, Öst M, Merilä J (2009) Relatedness and spatial proximity as determinants of host-parasite interactions in the brood parasitic Barrow’s goldeneye (Bucephala islandica). Mol Ecol 18:2713–2721
Lusignan AP, Mehl KR, Jones IL, Gloutney ML (2010) Conspecific brood parasitism in common eiders (Somateria mollissima): do brood parasites target safe nest sites? Auk 127:765–772
Lyon BE (1993a) Tactics of parasitic American coots: host choice and the pattern of egg dispersion among host nests. Behav Ecol Sociobiol 33:87–100
Lyon BE (1993b) Conspecific brood parasitism as a flexible female reproductive tactic in American coots. Anim Behav 46:911–928
Lyon BE (2003) Ecological and social constraints on conspecific brood parasitism by nesting female American coots (Fulica Americana). J Anim Ecol 72:47–60
Lyon BE, Eadie JM (2008) Conspecific brood parasitism in birds: a life-history perspective. Annu Rev Ecol Evol Syst 39:343–363
Lyon BE, Eadie JM (2017) Why do birds lay eggs in conspecifics’ nests? In: Soler M (ed) Avian brood parasitism. Springer, New York, pp 105–123
Lyon BE, Everding S (1996) High frequency of brood parasitism in a colonial waterbird, the eared grebe Podiceps nigricollis. J Avian Biol 27:238–244
McRae SB (1997) A rise in nest predation enhances the frequency of intraspecific brood parasitism in a moorhen population. J Anim Ecol 66:143–153
Odell NS, Eadie JM (2010) Do wood ducks use the quantity of eggs in a nest as a cue to the nest’s value? Behav Ecol 21:794–801
Paasivaara A, Rutila J, Pöysä H, Runko P (2010) Do parasitic common goldeneye Bucephala clangula choose nests on the basis of host traits or nest site traits? J Avian Biol 41:662–671
Palmer RS (1976) Handbook of North American birds, vol. 3. Yale University Press, New Haven
Payne RB (1977) The ecology of brood parasitism in birds. Annu Rev Ecol Syst 8:1–28
Petrželková A, Klvana P, Albrecht T, Hořák D (2013) Conspecific brood parasitism and host clutch size in common pochards Aythya ferina. Acta Ornithol 48:103–108
Pöysä H (1999a) Conspecific nest parasitism is associated with inequality in nest predation risk in the common goldeneye (Bucephala clangula). Behav Ecol 10:533–540
Pöysä H (1999b) Association between conspecific nest parasitism and the timing of breeding in the common goldeneye Bucephala clangula: an alternative interpretation. Ornis Fenn 76:89–92
Pöysä H (2006) Public information and conspecific nest parasitism in goldeneyes: targeting safe nests by parasites. Behav Ecol 17:459–465
Pöysä H, Pesonen M (2007) Nest predation and the evolution of conspecific brood parasitism: from risk spreading to risk assessment. Am Nat 169:94–104
Pöysä H, Milonoff M, Ruusila V, Virtanen J (1999) Nest-site selection in relation to habitat edge: experiments in the common goldeneye. J Avian Biol 30:79–84
Pöysä H, Eadie JM, Lyon BE (2014) Conspecific brood parasitism in waterfowl and cues parasites use [special issue]. Wildfowl 4:192–219
R Core Team (2017) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org. Accessed 25 Aug 2019
Reichart LM, Anderholm S, Muñoz-Fuentes V, Webster MS (2010) Molecular identification of brood-parasitic females reveals an opportunistic reproductive tactic in ruddy ducks. Mol Ecol 19:401–413
Ricklefs RE (1969) An analysis of nesting mortality in birds. Smithson Contrib Zool 9:1–48
Robertson GJ, Watson MD, Cooke F (1992) Frequency, timing, and costs of intraspecific nest parasitism in the common eider. Condor 94:871–879
Sayler RD (1985) Brood parasitism and reproduction of canvasbacks and redheads on the delta marsh. PhD Dissertation, University of North Dakota
Sayler RD (1992) Ecology and evolution of brood parasitism in waterfowl. In: Batt BDJ, Afton AD, Anderson MG, Ankney CD, Johnson DH, Kadlec JA, Krapu GL (eds) Ecology and management of breeding waterfowl. University of Minnesota Press, Minneapolis, pp 290–322
Semel B, Sherman PW (1986) Dynamics of nest parasitism in wood ducks. Auk 103:813–816
Semel B, Sherman PW, Byers SM (1988) Effects of brood parasitism and nest-box placement on wood duck breeding ecology. Condor 90:920–930
Sorenson MD (1991) The functional significance of parasitic egg laying and typical nesting in redhead ducks: an analysis of individual behaviour. Anim Behav 42:771–796
Spurr EB, Milne H (1976) Factors affecting laying date in the common eider. Wildfowl 27:107–109
Thimot N (2018) Conspecific brood parasitism in red-breasted mergansers Mergus serrator: cues parasites use for selecting a nest. Thesis, McGill University
Traylor JJ, Alisauskas RT, Kehoe FP (2004) Nesting ecology of white-winged scoters (Melanitta fusca deglandi) at Redberry Lake, Saskatchewan. Auk 121:950–962
Waldeck P, Hagen JI, Hanssen SA, Andersson M (2011) Brood parasitism, female condition, and clutch reduction in the common eider Somateria mollisima. J Avian Biol 42:231–238
Weller MW (1957) An automatic nest-trap for waterfowl. J Wildl Manag 21:456–458
Weller MW (1959) Parasitic egg laying in the redhead (Aythya americana) and other North American Anatidae. Ecol Monogr 29:333–365
Westerskov K (1950) Methods for determining the age of game bird eggs. J Wildl Manag 14:56–67
Yom-Tov Y (2001) An updated list and some comments on the occurrence of intraspecific nest parasitism in birds. Ibis 143:133–143
Young AD, Titman RD (1986) Costs and benefits to red-breasted mergansers nesting in tern and gull colonies. Can J Zool 64:2339–2343
Young AD, Titman RD (1988) Intraspecific nest parasitism in red-breasted mergansers. Can J Zool 66:2454–2458
Zink AG (2003) Intraspecific brood parasitism as a conditional reproductive tactic in the treehopper Publilia concave. Behav Ecol Sociobiol 54:406–415
Acknowledgments
This manuscript was part of NJT’s MSc thesis. We are grateful to D. Bilodeau, K. Francis, J. Haché, F. Leblanc, K. S. Seaborn, B. Spinney, and E. Titman for their field assistance. We thank Kouchibouguac National Park, particularly É. Tremblay and B. Martin, for logistical support. M. Kaouass led analyses of microsatellites. We appreciate discussions on randomization tests with B. Lyon. J. Head, B. Lyon, and five anonymous reviewers provided very helpful comments on earlier versions of the manuscript.
Funding
Financial support was provided by the Natural Sciences and Engineering Research Council of Canada (Canada Graduate Scholarship to NJT and DDG-2019-06038 to SRC), New Brunswick Wildlife Trust Fund, McGill University, Bird Protection Québec, and Université Sainte-Anne.
Author information
Authors and Affiliations
Contributions
NJT, RDT, and SRC conceived and designed the study and collected data; NJT conducted statistical analyses, interpreted results, and drafted the manuscript; RDT, KHE, and SRC supervised the research; all the authors contributed to editing various versions of the paper and approved the final manuscript.
Corresponding author
Ethics declarations
Ethical statement
All applicable international, national, and institutional guidelines for the care and use of animals were followed. All procedures involving animals were in accordance with the ethical standards of the institution at which the study was conducted (McGill University Animal Care Committee; permits # 1926 and #7329).
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by M. Soler
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Thimot, N.J., Titman, R.D., Elliott, K.H. et al. Conspecific brood parasitism in an upland-nesting bird: cues parasites use to select a nest. Behav Ecol Sociobiol 74, 27 (2020). https://doi.org/10.1007/s00265-020-2808-3
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-020-2808-3