
Abstract
Song by female birds is rare in some species, particularly at north-temperate latitudes. Nevertheless, female song can carry important signal content and may be used in functional ways, both when sung solo and when combined into partner duets. Evidence supports the idea that duets reflect elevated threat levels because they indicate partner cooperation, but this comes from species with frequent female song and duetting. Here we asked the following questions about infrequently given female song in canyon wrens (Catherpes mexicanus) in Western North America. Do female birds use song in a context that implies functional significance? Do canyon wren mates combine their songs to form duets? Are duets a more threatening signal than non-overlapping dual-sex solo songs? To address these questions, we challenged canyon wrens with paired simulated intruders singing alternating solo songs or duets formed by overlapping male and female songs with consistent timing. Results indicated that female canyon wrens approached and sang in response to conspecific song playback as quickly as males did and increased their song rates significantly. Partners did not overlap their songs to create duets more often than expected by chance, and neither sex responded more strongly to duets than to solo songs. Outcomes match the prediction that duets only carry different signal content from solo song in species that frequently duet, and suggest that canyon wren female song is highly functional despite being rarely used outside of contest situations.
Significance statement
Animal signals mediate social interactions in myriad ways. The majority of studies of avian song have focused on conspicuous, frequently given, signals but infrequently given signals may also have important fitness consequences. We examined how the usage of a rare signal, canyon wren female song, changed during contest situations and in response to a coordinated duet versus non-coordinated solo songs. Rates of female song increased significantly during contest situations, but duets did not provoke stronger responses than solos. Although duets are threatening to species that duet (including many wren species), they do not appear to be universally salient. Canyon wren female song, in contrast, carries strong signal content despite being used infrequently outside of contest situations. Results highlight the potential functionality of rare signals, as well as the variability in signaling strategies across avian species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Studies of animal communication generally focus on frequently given signal types, with the understanding that such signals play an important role in social organization and behavior (Dawkins and Guilford 1991; Bradbury and Vehrencamp 2011; Laidre and Johnstone 2013). Rarely given signals are less often studied, in part because they are difficult to sample, but may also provide reliable information to receivers and can have important fitness consequences (Hauser and Nelson 1991). In many bird species, females sing less frequently than males (Slater and Mann 2004; Odom and Benedict 2018). Historically, infrequent female song was sometimes considered “aberrant” and researchers have assumed that rarely given songs are unimportant or non-functional (Nice 1943; Byers and King 2000). Recent advances, however, have demonstrated that in many species female bird song is elaborate, learned, and functional, much as it is among males (Langmore 1998; Riebel et al. 2005; Hall and Langmore 2017). Females from a range of avian species use song in territory defense and/or mate attraction (Hoelzel 1986; Cooney and Cockburn 1995; Langmore et al. 1996; Cain and Langmore 2015; Krieg and Getty 2016). Furthermore, female bird song is more taxonomically widespread than historically thought and is likely ancestral among songbirds (Garamszegi et al. 2007; Benedict 2008; Odom et al. 2014; Tobias et al. 2016). Thus, it has become evident that if we want to understand avian vocal communication systems, we need to understand the form and function of female song across a wide range of species with varying rates of the behavior (Riebel et al. 2005; Odom and Benedict 2018). In particular, species with infrequent female song offer a valuable opportunity to test the potential functionality and timing of a rarely given signal.
Much of the recent work on female bird song has focused heavily on understanding song function, correlated traits, and evolutionary history (Price 2009; Odom et al. 2014; Cain and Langmore 2015; Krieg and Getty 2016; Najar and Benedict 2016; Webb et al. 2016). To complement this functional focus, the field needs studies on the timing and context of female song use. In any communication system, individuals may adjust usage to optimize signal efficacy (Gil and Gahr 2002). While some signal types are general or multi-functional, others are restricted or situation-specific (Catchpole and Slater 2008; Laidre and Johnstone 2013). Female birds of many species are known to sing in more restricted contexts than males of the same species, raising questions about what situations provoke female song. Two contexts in which females sing regularly are (1) during sex-specific resource defense challenges and (2) when combining their songs with a partner’s song to form a duet (Langmore 1998; Hall 2009). Thus, among the many bird species in which females do not sing frequent broadcast song throughout the breeding season, those same females can selectively time their song use in relation to rivals or mates.
By singing only in certain situations, female birds may increase signal efficacy via targeting of particular receivers. Existing research shows that female birds often sing more aggressively in response to other females, but less so in response to males during contests (or simulated contests) over resources (Elekonich 2000; Koloff and Mennill 2011; Spencer 2012; Krieg and Getty 2016). Male birds, on the other hand, often sing in response to both male and female conspecifics (Levin 1996; Appleby et al. 1999; Logue and Gammon 2004). In many temperate-zone species, males perform the majority of the resource defense behaviors; as a result, female song could become relatively rare, even when it is functional in that context.
The amount of coordination between male and female partners during a singing bout varies widely, even among closely related species (Mann et al. 2009; Kovach et al. 2014). In some species, males and females sing only solo songs, not predictably alternating song parts or overlapping them in time. On the other end of the spectrum, some species sing extremely precise, antiphonal duets where the male and female alternate singing with only milliseconds separating their parts (Mann et al. 2003; Hall 2004, 2009; Templeton et al. 2013). In between these two extremes are species that frequently create duets where males and females overlap their songs in predictable ways to varying degrees (Hall 2004, 2009; Benedict 2010). It has been hypothesized that duets create a more threatening signal than solo songs by demonstrating cooperation between mates working together to defend resources (Hall 2009; Dahlin and Benedict 2013). The majority of support for this idea comes from the many species in which duetting individuals actively modify the timing of their songs to promote duet coordination and increase duetting rates in response to rivals (Rivera-Cáceres 2015; reviewed in Hall 2009). Coordinated duets may allow birds to present a united front, but may still indicate sex-specific resource defense when males and females vary song and duet rates in different ways (Levin 1996; Appleby et al. 1999; Logue and Gammon 2004; Templeton et al. 2011). In some species, duet coordination has been shown to indicate pair quality and predict mating fidelity (Hall and Magrath 2007; Baldassarre et al. 2016). In contrast, a study of three wren species which manipulated duet coordination did not find that more tightly coordinated duets were most threatening (Kovach et al. 2014).
Although research results have varied across species, the bulk of existing evidence suggests that mates can increase the efficacy of song as a resource defense signal by creating duets. Thus far, studies testing this idea have generally done so in species that regularly duet, and almost all have compared birds’ responses to duets with their responses to either male or female song (see Kovach et al. 2014 for an exception). No studies that we are aware of have tested the question of whether song coordination carries signal content in a non-duetting species or in a species that duets rarely because females sing rarely. Few studies have asked whether birds respond to duets differently from both male and female songs presented in the same playback stimulus but without the species-typical timing of a duet.
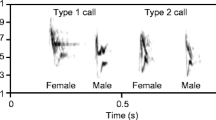
Canyon wrens present an interesting opportunity to investigate the contextual use of female song and the effect of male–female song coordination on its perceived quality. Canyon wrens are resident in temperate Western North America where pairs maintain year-round territories, providing long-term resource-defense opportunities for both sexes (Jones and Dieni 1995). Although not well studied, evidence suggests that pairs remain together across multiple breeding seasons, a trait typical of duetting species (Jones and Dieni 1995; Benedict 2008; Tobias et al. 2016). Male broadcast song is common, and males aggressively respond to playback of both male and female song (Benedict et al. 2012; Spencer 2012). Female song in canyon wrens is widespread, but infrequent (Spencer 2012; Benedict et al. 2013). Female canyon wrens do not respond to male song playback but do respond readily to female song playback (Spencer 2012; LB unpublished data). Canyon wrens are not reported to duet regularly; however, multiple instances have been recorded where female songs overlapped male songs (Fig. 1) (Fig. 4 in Spencer 2012; Benedict et al. 2013). These observations led Spencer (2012) to hypothesize that female canyon wrens time their songs to create overlapping duets similar in structure to those of Carolina wrens, a duetting temperate North American species (Shuler 1965). These natural history traits provide opportunities to test multiple ideas. First, we investigated how rates of female song production varied in broadcast singing versus contest situations by simulating intrusion via playback of male and female song. Second, we assessed whether canyon wrens duet in response to simulated intrusion and whether they perceive duets as more threatening than solo male and female songs. By studying a species that is not known to be a frequent duetter, we tested for the possibility that duets carry different signal contents than solo song alone, even in a species that has not evolved to duet frequently. By studying a species with rare female song, we tested for context-specific functional use of that infrequent signal type.

Spectrograms of a male canyon wren song (a), a female canyon wren song (b), and overlapped songs with a hypothesized duet form (c)
Methods
We conducted playback experiments on 29 canyon wren territories in Colorado between May 5 and July 15, 2016. Twenty-five of the 29 territories were along the Front Range of the Rocky Mountains at GIS locations ranging from 39.5° to 40.8° N and approximately − 105° W. Four territories were found in Picture Canyon on the Comanche National Grassland (37.0° N, − 102.7° W). Canyon wrens in this area actively defend large (5–6 ha), widely spaced territories (Jones and Dieni 1995; Warning and Benedict 2015). Individual subjects were not banded, but all territories were at least 0.65 km apart, and whenever a focal territory had an immediate neighbor, birds on neighboring territories were observed simultaneously to ensure that all were separate individuals. While canyon wrens are monomorphic, it was not difficult to distinguish between the male and female due to the high rates of singing, especially by males, and their easily distinguished sex-specific songs (Fig. 1) (Jones and Dieni 1995; Benedict et al. 2013). On seven territories, a female was never seen. Accordingly, those pairs were excluded from analyses involving females, and male pairing status was accounted for in analyses involving males.
Playback tracks
All subjects were tested with two types of playback: (1) a track containing solo male and female canyon wren songs spaced evenly apart and (2) a “duet” track containing male and female canyon wren songs overlapping with a short and consistent response time. We created ten sets of playback tracks using recordings from ten males and ten females, with each set containing a solo track and a duet track. In each set, the solo playback consisted of five repeats of one female song and five repeats of one male song. Female and male songs alternated and were separated by 10 s from the start of each song, a natural rate of singing for canyon wrens (Benedict et al. 2013). This structure includes much longer periods of silence between songs than is found in species with alternating duets, which typically have tight (often only a few milliseconds) temporal coordination (Hall 2009). The duet playback consisted of the same five male and five female songs, but the songs were overlapped to form five duets, each separated by 20 s from the start of the previous duet. The duet tracks were created by starting the female song approximately halfway through the male song. This replicates the “duet” form observed by Spencer (2012, Fig. 4).
Seventeen songs (10 male, seven female) for the playbacks came from our own wave files made using a Sennheiser MKH-70 shotgun microphone connected to a Marantz PMD 661 MKII solid-state digital recorder, using a sampling rate of 48 kHz, and 24-bit depth. All recordings were made in 2012 and were played in locations at least 10 km from the recording sites. Three female songs came from recordings of Colorado female canyon wrens available on Xeno-canto (XC100999, XC102224, XC102260), used with permission of the recordist and converted from mp3 to wave files with properties matching the other playback stimuli. We were limited to ten sets of playback stimuli based on how few high-quality recordings of canyon wren female song were available. Female canyon wrens produce only a single song type that was represented in all tracks (Benedict et al. 2013). Individual males in our population sing up to six song types (Benedict et al. 2013) and deliver them in bouts with eventual variety. All male songs used in playback tracks were of a single type that is widely distributed and sung by all previously assessed local individuals (type 5 in Benedict et al. 2013). To ensure similarity across stimuli, we included only the descending cascade and not the harsh buzzy “cheet” notes that males sometimes append in variable numbers to their songs (visible at the end of the song in Fig. 1a) (Benedict et al. 2013). Playback tracks were created in Audacity (https://www.audacityteam.org/) by repeating and overlapping songs and adding periods of silence between them. Tracks were not filtered or amplified, but for some recordings, individual background sounds were removed.
Playback trials
All playback trials were conducted between 0700 and 0830 h Mountain Standard Time. Each trial was continuously recorded using a Sennheiser MKH-70 shotgun microphone connected to a Marantz PMD 661 MKII solid-state digital recorder using a sampling rate of 48 kHz and 24-bit depth. Playback tracks were played over an Anchor AN-Mini speaker held by the experimenter located near the center of the subject’s territory and played at 80 dB(C) (re. 20 μPa) peak amplitude at one meter. Previous work has verified that this approximates natural male singing volumes and that responding birds readily approach and sing in response to playback even with a person present (Benedict et al. 2012). Within tracks, male and female song amplitudes differed. Measurements conducted in Raven Pro 1.3 software (http://www.birds.cornell.edu/brp/raven) indicated that female song peak amplitudes were 3.57 ± 3.36 (SD) dB (range = − 0.4–8.9) lower than male song peak amplitudes. Bird song amplitude can differ to a receiver based on factors including environmental conditions, distance to the sender, and the sender’s head orientation (Zollinger et al. 2012). Our within-track differences in amplitude fall well within the range of normal biological variation and would indicate that the female was farther away from or facing away from the receiver, relative to the male. Sound levels were set and verified offsite prior to playback trials using a Radio Shack 33-2055 digital sound level meter with a C frequency weighting curve and fast time weighting.
Each playback trial included three sections and lasted for 1 h and 30 min. First, we conducted 30 min of pre-playback recording and observation, followed by two 30-min sections consisting of a 90-s playback followed by 28.5 min of silence. Playbacks were begun at these set times within the trial, regardless of where the birds were located. This meant that sometimes birds were out of sight at the start of playback. Given the playback amplitudes, we expect that birds within territory boundaries would hear them, and we have no reason to expect that birds would be consistently farther away from the speaker in one treatment condition versus the other. Each pair heard both playback treatments: a dual sex solo playback and a duet playback separated by 30 min with presentation order randomly determined for the first pair of subjects and then alternating in successive trials. Previous work has shown that male canyon wrens return to natural singing levels approximately 20 min after playback, making this interval long enough for birds to resume normal behavior between playbacks (Benedict et al. 2012).
During each 30 min segment of a playback experiment, the experimenter dictated behavioral observations of both the male and the female that included data regarding (1) the latency for each sex to first approach the playback speaker and (2) the closest approach to the playback speaker, both measured in meters and estimated visually. All recordings were inspected and analyzed in Raven Pro 1.3 software. From the recordings, we extracted the latency to first approach, closest approach, and latency to first song for the male and female from the start of each playback segment. We collected similar data during the first 30 min, the pre-playback section, to use as a baseline for comparison. To assess song rate, we counted the number of male and female songs sung during the pre-playback period and during each post-playback period (solo and duet). We recorded when each male and female song began and ended in order to calculate rates and timing of song overlap. It was not possible to analyze response or song recording data blind because treatment type was audible to the observer in both contexts.
Previous research has shown that male canyon wrens respond to conspecific territorial challenges by shortening the cascade portions of their songs and adding harsh cheet notes to the ends of their songs (Benedict et al. 2012). If this is a generalized aggressive response, and if duet songs are more threatening than solo songs, then we would expect to see that males (and potentially females) respond to duets versus solo songs in this way. We therefore measured song properties from songs produced prior to and following solo and duet playback. Only high-quality song examples were included in the analysis; we did not measure songs that were low in amplitude or obscured when visualized. A single observer (TGH) measured the duration (sec) of male and female songs and counted the number of cheet notes at the end of male songs using methods similar to those in Benedict et al. (2012). All female song syllables have a harsh buzzy quality similar to male cheet notes. Therefore, following the motivation-structural and non-linearity hypotheses, which suggest that such sounds indicate arousal or aggression, we counted the total number of syllables in female songs (Morton 1977; Blumstein and Recapet 2009). Song duration was collected from waveforms and associated Hanning spectrograms. Spectrograms were generated with a discrete Fourier transform of 512 samples and a time resolution of 5.8 ms with a 50% overlap in the time grid in Raven Sound Analysis Software. Resulting data allowed us to ask whether both sexes adjusted song length and used more harsh sounds in response to solos versus duets. We examined whether males altered song form following playback but were unable to do the same for females because so few sang prior to playback.
Statistical analysis
To assess whether canyon wrens actively overlap their songs, we used the SONG package implemented in R version 3.4 (https://www.r-project.org/) (Masco et al. 2016). In SONG, we ran both the resampling randomization procedure and the duty cycle procedure of Ficken et al. (1974). These methods offer two ways to calculate whether song overlap occurs more often than expected by chance for each pair (Masco et al. 2016). Similar procedures have been used to verify the presence of duetting in rufous-and-white-wrens, a species with overlapping duets (Masco et al. 2016). Input data were song start and end times for all male and female songs during the two playback response periods. We included songs with poor recording quality and those obscured by other sounds because leaving songs out of the analysis would provide incorrect patterns of song usage. To do so, we estimated the length of songs with poor recording quality as the average length of the measured songs for the appropriate individual. Songs with estimated lengths comprised 9.85 ± SD 8.79% of the songs of each male and 6.56 ± 7.12% of the songs of each female. Although we hypothesized that females generally form duets by overlapping males, we also asked whether males formed duets by overlapping females, and we asked whether each sex avoided overlapping the other, as might be expected if the species creates alternating duets (Hall 2009). To describe potential duet timing, we calculated the latency for males and females to begin singing after their partner sings for all instances in which a bird sang following a song from its partner and for all instances where songs overlapped. For overlapping songs, we report what percentage of the first song was overlapped by the second and the associated standard deviation. If each sex overlaps the songs of the other with predictable duet timing, we would expect to see small standard deviations. Throughout, all means are reported with standard deviations unless otherwise noted.
Playback response measures (including the number of songs given, latency to sing, closest approach and latency to approach) were compared for members of each sex between pre-playback, solo response, and duet response periods using repeated measures ANOVAs. We followed these with Wilcoxon signed rank tests comparing responses to solo and duet playback, matched by individual. For male analyses, we included all individuals, even those without mates. All behavioral measures were also compared between males and females for each period using Wilcoxon signed-rank tests. For male–female comparisons, territories without females were excluded to allow for matching by pair. All tests were two-tailed. Because we assessed four response variables, we report significances not only at p = 0.05 but also at a critical value of α = 0.0125, following Bonferroni correction.
To validate the inclusion of unmated males, we verified that all males responded to playback similarly using Wilcoxon rank-sum tests for our four behavioral variables. Mated (n = 22) and unmated (n = 7) males showed no difference in their number of songs given (Z1 = − 18, p = 0.86), latency to sing (Z1 = 0.33, p = 0.74), latency to approach (Z1 = 0.38, p = 0.70), or closest approach (Z1 = 0.33, p = 0.72) in response to playback (solo and duet responses averaged). We validated the use of female song stimuli downloaded as mp3 files from xeno-canto by verifying that the seven females who heard playback converted from the mp3 format did not differ from the 15 who heard female songs recorded in the wave format in any of the following response measures (solo and duet responses averaged): number of songs given (Z1 = − 1.24, p = 0.21), latency to sing (Z1 = 1.35, p = 0.18), latency to approach (Z1 = − 0.035, p = 0.97), and closest approach (Z1 = 0.035, p = 0.97). We tested for a potential confounding effect of female song sound pressure level variation using linear regression of our response variables on the difference in male and female playback peak amplitude and found no suggestion that peak amplitude affected the number of songs given (F1 = 1.11 p = 0.30), latency to sing (F1 = 0.60, p = 0.44), latency to approach (F1 = 0.95, p = 0.33), and closest approach (F1 = 0.41, p = 0.53) of female responders.
To compare the structural features of male and female songs given in response to solo and duet playback, we constructed linear mixed models for duration and numbers of harsh, broadband notes using the standard least squares method. For both sexes, playback order and Pair ID were included as random factors. For males, pairing status was also included as a random factor, nested within Pair ID. We constructed similar models to evaluate whether male song duration and number of cheet notes differed in the pre-playback and post-playback periods. That model included playback type, Pair ID, and pairing status nested within Pair ID as random factors.
All procedures were conducted following ethical guidelines found in Fair et al. (2010) and in accordance with applicable international and national guidelines. Work was overseen by the University of Northern Colorado’s Institutional Animal Care and Use Committee (Protocol No. 1506C-LB-Birds-18).
Results
Songs and duets
Only one female sang (21 songs) during the pre-playback period and she did not overlap any songs with her mate’s 53 songs during this period. Conversely, 28 of the 29 males sang during the pre-playback period, and all males sang following playback (Fig. 2). Following playbacks, 17 of 22 (77%) females sang and 13 of those 17 (76%) females overlapped their song(s) with their mate at least once. Combined, we recorded 119 instances of overlapping songs, which is 21% of all female songs given during the post-playback period and 7% of all male songs. The majority of female songs did not overlap with or immediately follow male songs. On average, female songs were given 17.08 ± 82.12 (median 3.20) s after the ends of the preceding male songs, and male songs were given 18.93 ± 69.33 (median 1.63) s after the ends of the preceding female songs. It is worth noting that means are inflated by skewed distributions, and the median values might be more representative of typical song timing during aggressive encounters.

Average number (± SD) of songs given by males (blue) and females (red) during the 30-min pre-playback period, the 30 min following alternating solo playback, and the 30 min following duet playback
SONG results offer no evidence that canyon wren males and females avoid overlapping their mates’ songs, as would be expected of a species that creates alternating duets. SONG results offer very limited evidence that they consistently overlap their mates’ songs (Table 1). The randomization resampling approach identified one female and one male that overlapped partner songs more than expected by chance (Table 1). The duty cycle method identified three females and no males that overlapped partner songs more than expected by chance (Table 1). None of the potentially duetting individuals heard the same playback stimuli. The four stimuli associated with duetting included tracks in which female song amplitude was a mean of 4.88 ± 3.99 dB lower than male song amplitude (range = − 0.4–8.9). Females overlapped male songs 26 times, and they initiated singing a mean of 1.27 ± 1.00 s after the start of the male songs. In doing so, females sang during 45 ± 33% of the duration of each overlapped male song. Males overlapped female songs 93 times, initiating their songs a mean of 1.90 ± 1.79 s after the start of the female songs. In doing so, males sang during 44 ± 36% of the duration of each overlapped female song.
Behavioral measures
Canyon wrens of both sexes responded to playback of male and female conspecific song by singing more songs (Fig. 2) and approaching the playback speaker. Repeated measures ANOVAs found strong differences in behavioral response, driven by differences between the control pre-playback period and the two playback treatments (Table 2). The only response measure not strongly affected by playback was the male latency to sing, which becomes non-significant after Bonferroni correction. This reflects the fact that even prior to playback males sang frequently. To complement these results, we ran a similar analysis on female song data collected as part of the study reported in Benedict et al. (2012). In that study, we used only playback of male song. Males increased song rates significantly in response to playback, but females showed no difference in song rates prior to and following playback of male solo song without female solo song (W1 = 2.00, p = 0.63) (Benedict et al. 2012).
Males showed no differences in number of songs sung, latency to sing, closest approach, or latency to approach in response to the solo versus duet playbacks (Table 2). Female singing rates, latencies to sing, and latencies to approach the speaker did not differ following solo versus duet playback (Table 2). Females approached the speaker more closely during solo playback than during duet playback, but the significance of this difference goes away after Bonferroni correction (Table 2).
In all treatment periods, males sang significantly more songs than females (pre-playback W1 = 126.50, p < 0.0001, solo W1 = 112.50, p < 0.0001, duet W1 = 112.50, p < 0.0001) (Fig. 2). During the pre-playback period, males showed significantly lower latencies to sing (W1 = 125.50, p < 0.0001) and approach (W1 = 25.50 p = 0.0059) than females, and they approached the speaker more closely (W1 = 20.00 p = 0.041). In contrast, during the solo response (W1 = 22.50, p = 0.48) and duet response (W1 = 36.50, p = 0.25) periods, females and males did not differ in their latencies to sing (Fig. 3). Similarly, males and females did not differ in their latencies to approach following solo (W1 = 22.00, p = 0.43) and duet (W1 = 25.00, p = 0.36) playback (Fig. 3). Males came closer to the speaker than females did following duet (W1 = 33.50, p = 0.034), but not solo (W1 = 21.50, p = 0.28) playback. Differences in closest approach values between males and females in the pre-playback and duet response periods lose significance after Bonferroni correction to a critical value of α < 0.0125.

Two measures (± SD) of latency to respond to solo and duet playback for male (blue) and female (red) canyon wrens
Song form
In the pre- versus post-playback comparisons, males significantly shortened song cascade durations and increased the number of harsh-sounding cheet notes per song following playback (Table 3). In response to solo versus duet playbacks, males did not sing songs with measurably different durations or numbers of cheet notes (Table 4). Females also showed no differential responses to solo versus duet playbacks (Table 4).
Discussion
Our results support existing observations that female canyon wrens rarely produce spontaneous song. In 11 h of recording on 22 territories that housed female canyon wrens, we documented only one singing female (5% of individuals). Parallel observations over a comparable time period documented spontaneous singing by 28 of 29 males (97%). Although canyon wren females sing infrequently, a previous study suggested that they use this signal in sex-specific resource defense (Spencer 2012). To test this hypothesis, we simulated territory intrusions by a pair of conspecifics using playback of male and female canyon wren songs. Existing work shows that females do not respond to male song alone (Benedict et al. 2012), but in the study reported here, they did respond to combined male and female song. When exposed to a same-sex territorial intruder, female canyon wrens overwhelmingly responded by approaching and singing towards the threat. This reliable use of female song in a resource defense context suggests that this signal, although rare in unprovoked settings, is highly functional. Like many other species of birds (including other wrens), female canyon wrens appear to use song as a resource defense signal, and the sex-specific form of the song allows for targeted communication between members of the same sex (Levin 1996; Appleby et al. 1999; Logue and Gammon 2004; Logue 2005; Templeton et al. 2011). These results raise the possibility that female song might be functional in other species where it has been understudied or rarely observed (Byers and King 2000; Taff et al. 2012)
It has been previously shown that male and female canyon wrens react aggressively to songs of their own sex (Benedict et al. 2012; Spencer 2012), but the responses of the two sexes have not been compared with each other. Although female canyon wrens sing less frequently than males, we found that they responded to same-sex song as aggressively as males did based on behavioral measures. Females approached just as quickly, approached just as closely, and sang just as quickly as males did when challenged by a simulated intruder. Importantly, these responses did not covary with pairing status, playback stimulus recording type (mp3 versus wave) and female song peak amplitude relative to male song peak amplitude. It has long been known that some of these variations, notably pairing status and amplitude, can affect behavioral responses to playback, and we recommend that future studies standardize these variables (Catchpole 1977; Searcy 1996). Nevertheless, even with these sources of variation in playback stimuli, we documented rapid responses, suggesting that male and female canyon wrens are attentive to conspecific signals in their environment and recognize them even in the face of signal variations.
Other species of birds show a wide range of variability in sex-biased playback response, with females sometimes being less responsive than males and sometimes more responsive than males (Mennill 2006; Illes and Yunes-Jimenez 2009; Koloff and Mennill 2011; Hall et al. 2015). Typically, the species with the strongest female responses are the ones in which females sing most often (Illes and Yunes-Jimenez 2009). In at least one species with infrequent female song, females show weak responses to conspecific intruders (Hall et al. 2015). Given the rarity of canyon wren female song, the rapid female responses that we documented are perhaps surprising and likely indicate that female song is highly salient to other females. Females of other species use song to defend both territories and mates, resources that are critical to individual fitness, and often target these songs towards other females (Cooney and Cockburn 1995; Seddon and Tobias, 2006). Canyon wrens fit this pattern; the rarity of female song in our study population likely reflects a low rate of female–female confrontation which arises because territories are widely spaced (Warning and Benedict 2015).
Canyon wren females have been observed to overlap some songs with those of their mates (Spencer 2012; Benedict et al. 2013). Because female song is rare, it has been difficult to quantify whether song overlap is frequent enough to conclude that members of this species actively produce duets by coordinating song timing. By stimulating female song via playbacks, we were able to address this question. Overall, we found that most females did not overlap mate songs more often than expected by chance. Most males also did not overlap mate songs, and neither sex avoided song overlapping. However, there was a large amount of variability in song overlapping among pairs, and our two approaches indicated that either two (resampling) or three (duty cycle) out of 34 individuals did tend to initiate songs during the songs of their partners (Table 1). When they did so, however, there was variability in their timing; song initiation times and overlap percentages of males and females had high variation. The literature does not contain perfect criteria to conclusively distinguish duets as different from overlapping songs, but researchers generally agree that partners creating duets must regularly coordinate their songs with temporal precision (Hall 2004, 2009). The variation that we documented in song overlap percentages, and the inconsistent interval lengths between female and male songs do not fit this expectation. Moreover, duetting is considered to be a species-level trait exhibited by all individuals (Hall 2004, 2009). Our results do not fulfill either of those criteria, and therefore, we do not consider canyon wrens to be a duetting species. Nevertheless, four individuals did overlap partner songs more often than the null expectation, leaving open the possibility that some individuals do intentionally initiate songs while their mates are singing.
A main objective of this study was to test whether canyon wrens differentiate between overlapping (duet) and non-overlapping (solo) playbacks. We confirmed that both male and female canyon wrens responded strongly to conspecific song, but neither sex behaviorally distinguished between the duetting and solo playback tracks. Within each sex, there was no difference in behavioral responses including the time to approach, time to first song, or number of songs produced (Table 2; Figs. 1 and 2). Females approached speakers playing solo songs more closely than duets (although the effect was minor). If anything, this suggests that solo songs are more threatening to females than duets are. Solos might provoke stronger female responses because they represent a threat to the pair bond or because females are reluctant to directly confront larger-bodied males (Langmore 1998; Jones and Dieni 1995).
Results suggest that our overlapping playback structure neither reduced the salience of songs via masking (Brumm and Naguib 2009) nor increased their salience by demonstrating coordination. Much research has examined this latter question in duetting species. The majority of relevant studies have presented birds with duet playbacks, male solos, and female solos separately. Under these conditions, some duetting species respond more strongly to duets, while others respond more strongly to solos (Hall 2000, 2009; Seddon and Tobias 2006; Rogers et al. 2007; Illes and Yunes-Jimenez 2009). Importantly, however, almost all duetting species do behaviorally distinguish between these types of signals. The same holds true for studies with designs similar to ours where birds are presented with both male and female songs of varying coordination levels (Hall and Magrath 2007; Kovach et al. 2014). Thus, existing research indicates that song coordination is a salient signal for many duetting species, but our data suggest that it carries no information that alters behavior during conspecific contests in a non-duetting species. Future work should test this question in other groups to see how coordinated versus uncoordinated singing varies across families and species with different rates of female song and song coordination between mates.
In addition to examining how male and female canyon wrens vary their approach behaviors and song rates during simulated conspecific intrusion, we asked whether males and females altered song duration and structure in response to solo and duet playback. Following playback, males reduced the mean duration of their song by 0.19 s and increased the mean number of broadband harsh-sounding notes by 0.50 notes per song (Table 3). These results replicate a previous study which concluded that brief, harsh-sounding songs indicate aggression in canyon wrens (Benedict et al. 2012). The same features indicate aggression in other species, and general theories about acoustic signal structure suggest that animals should emphasize these song properties during contest situations (Morton 1977; Blumstein and Recapet 2009; Benedict et al. 2012; Geberzahn and Aubin 2014). Songs with these “aggressive” features were not, however, used more often by either sex in response to duet versus solo playback (Table 4), reinforcing the conclusion that our two stimuli did not convey different information to canyon wrens.
Taken together, our data suggest that canyon wrens do not perceive overlapping duets as a more threatening display than solo songs, but both sexes do consider these signal types to be threatening. We established that canyon wrens do not consistently overlap their songs in a way that would warrant labeling them as “duets,” and we showed that a non-duetting species does not act more aggressively towards overlapped than non-overlapped songs. This result counters the idea that coordination of songs into duets is a universally aggressive signal. We also found that canyon wren female song, despite being rare in our population, is used in ways consistent with a sex-specific resource defense function and provokes strong, rapid reactions in territory-holding females. Thus, this infrequently given signal is highly salient within the right context. Rare events in the life of an individual can have important fitness consequences. For example, loss of a territory or mate might cause complete reproductive failure for an individual bird. Across avian species, females regularly use song to prevent such losses (as well as in other ways with fitness consequences) (Halkin 1997; Langmore 1998; Cain et al. 2015; Brunton et al. 2016; Krieg and Getty 2016). Our results add to the growing consensus that although avian female song is less conspicuous than male song, it is no less functional (Hall and Langmore 2017).
Data availability
The datasets generated and analyzed during the current study are available from the corresponding author on reasonable request.
References
Appleby BM, Yamaguchi N, Johnson PJ, MacDonald DW (1999) Sex-specific territorial responses in tawny owls Strix aluco. Ibis 141:91–99
Baldassarre DT, Greig EI, Webster MS (2016) The couple that sings together stays together: duetting, aggression and extra-pair paternity in a promiscuous bird species. Biol Lett 12:20151025
Benedict L (2008) Occurrence and life history correlates of vocal duetting in North American passerines. J Avian Biol 39:57–65
Benedict L (2010) California towhee vocal duets are multi-functional signals for multiple receivers. Behaviour 147:953–978
Benedict L, Rose A, Warning N (2012) Canyon wrens alter their songs in response to territorial challenges. Anim Behav 84:1463–1467
Benedict L, Rose A, Warning N (2013) Small song repertoires and high rate of song-type sharing among canyon wrens. Condor 115:874–881
Blumstein DT, Recapet C (2009) The sound of arousal: the addition of novel non-linearities increases responsiveness in marmot alarm calls. Ethology 115:1074–1081
Bradbury JW, Vehrencamp SL (2011) Principles of animal communication, 2nd edn. Sinauer, Sunderland
Brumm H, Naguib M (2009) Environmental acoustics and the evolution of bird song. Adv Stud Behav 40:1–33
Brunton DH, Roper MM, Harmer AM (2016) Female song rate and structure predict reproductive success in a socially monogamous bird. Front Ecol Evol 4:13
Byers BE, King DI (2000) Singing by female chestnut-sided warblers. Wilson Bull 112:547–550
Cain KE, Langmore NE (2015) Female and male song rates across breeding stage: testing for sexual and nonsexual functions of female song. Anim Behav 109:65–71
Cain KE, Cockburn A, Langmore NE (2015) Female song rates in response to simulated intruder are positively related to reproductive success. Front Ecol Evol 3:119
Catchpole CK (1977) Aggressive responses of male sedge warblers (Acrocephalus schoenobaenus) to playback of species song and sympatric species song, before and after pairing. Anim Behav 25:489–496
Catchpole CK, Slater PJ (2008) Bird song: biological themes and variations. Cambridge University Press, Cambridge
Cooney R, Cockburn A (1995) Territorial defense is the major function of female song in the superb fairy-wren. Anim Behav 49:1635–1647
Dahlin CR, Benedict L (2013) Angry birds need not apply: a perspective on the flexible form and multifunctionality of avian vocal duets. Ethology 120:1–10
Dawkins MS, Guilford T (1991) The corruption of honest signalling. Anim Behav 41:865–873
Elekonich MM (2000) Female song sparrow, Melospiza melodia, response to simulated conspecific and heterospecific intrusion across three seasons. Anim Behav 59:551–557
Fair J, Paul E, Jones J (eds) (2010) Guidelines to the use of wild birds in research, 3rd edn. Ornithological Council, Washington DC
Ficken RW, Ficken MS, Hailman JP (1974) Temporal pattern shifts to avoid acoustic interference in singing birds. Science 183:762–763
Garamszegi LZ, Pavlova DZ, Eens M, Møller AP (2007) The evolution of song in female birds in Europe. Behav Ecol 18:86–96
Geberzahn N, Aubin T (2014) How a songbird with a continuous singing style modulates its song when territorially challenged. Behav Ecol Sociobiol 68:1–12
Gil D, Gahr M (2002) The honesty of bird song: multiple constraints for multiple traits. Trends Ecol Evol 17:133–141
Halkin SL (1997) Nest-vicinity song exchanges may coordinate biparental care of northern cardinals. Anim Behav 54:189–198
Hall ML (2000) The function of duetting in magpie-larks: conflict, cooperation, or commitment? Anim Behav 60:667–677
Hall ML (2004) A review of hypotheses for the functions of avian duetting. Behav Ecol Sociobiol 55:415–430
Hall ML (2009) A review of vocal duetting in birds. Adv Stud Behav 40:67–121
Hall ML, Langmore NE (2017) Fitness costs and benefits of female song. Front Ecol Evol 5:48
Hall ML, Magrath RD (2007) Temporal coordination signals coalition quality. Curr Biol 17:406–407
Hall ML, Rittenbach MRD, Vehrencamp SL (2015) Female song and vocal interactions with males in a neotropical wren. Front Ecol Evol 3:12
Hauser MD, Nelson DA (1991) ‘Intentional’ signaling in animal communication. Trends Ecol Evol 6:186–189
Hoelzel A (1986) Song characteristics and response to playback of male and female robins Erithacus rubecula. Ibis 128:115–127
Illes AE, Yunes-Jimenez L (2009) A female bird out-sings male conspecifics during simulated territorial intrusions. Proc R Soc Lond B 276:981–986
Jones S, Dieni J (1995) Canyon wren (Catherpes mexicanus). In: Rodewald PG (ed) The birds of North America. Cornell Lab of Ornithology, Ithaca. https://doi.org/10.2173/bna.197
Koloff J, Mennill D (2011) Aggressive responses to playback of solos and duets in a neotropical antbird. Anim Behav 82:587–593
Kovach KA, Hall ML, Vehrencamp SL, Mennill DJ (2014) Timing isn’t everything: responses of tropical wrens to coordinated duets, uncoordinated duets and alternating solos. Anim Behav 95:101–109
Krieg CA, Getty T (2016) Not just for males: females use song against male and female rivals in a temperate zone songbird. Anim Behav 113:39–47
Laidre ME, Johnstone RA (2013) Animal signals. Curr Biol 23:R829–R833
Langmore NE (1998) Functions of duet and solo songs of female birds. Trends Ecol Evol 13:136–140
Langmore NE, Davies NB, Hatchwell BJ, Hartley IR (1996) Female song attracts males in the alpine accentor Prunella collaris. Proc R Soc Lond B 263:141–146
Levin RN (1996) Song behaviour and reproductive strategies in a duetting wren, Thryothorus nigricapillus: II. Playback experiments. Anim Behav 52:1107–1117
Logue DM (2005) Cooperative defence in duet singing birds. Cogn Brain Behav 9:497–510
Logue DM, Gammon DE (2004) Duet song and sex roles during territory defense in a tropical bird, the black-bellied wren, Thryothorus fasciatovetris. Anim Behav 68:721–731
Mann NI, Marshall-Ball L, Slater PJB (2003) The complex song duet of the plain wren. Condor 105:672–682
Mann NI, Dingess KA, Barker FK, Graves JA, Slater PJB (2009) A comparative study of song form and duetting in neotropical Thryothorus wrens. Behaviour 146:1–43
Masco C, Allesina S, Mennill DJ, Pruett-Jones S (2016) The Song Overlap Null model Generator (SONG): a new tool for distinguishing between random and non-random song overlap. Bioacoustics 25:29–40
Mennill DJ (2006) Aggressive responses of male and female rufous-and-white wrens to stereo duet playback. Anim Behav 71:219–226
Morton ES (1977) On the occurrence and significance of motivation-structural rules in some bird and mammal sounds. Am Nat 111:855–869
Najar N, Benedict L (2016) Female song in new world wood-warblers (Parulidae). Front Ecol Evol 3:139
Nice M (1943) Studies in the life history of the song sparrow. II. Trans Linn Soc NY 6:l–328
Odom KJ, Benedict L (2018) A call to document female bird songs: applications for diverse fields. Auk: Ornithol Adv 135:314–325
Odom KJ, Hall ML, Riebel K, Omland KE, Langmore NE (2014) Female song is widespread and ancestral in songbirds. Nat Commun 5:3379
Price JJ (2009) Evolution and life history correlates of female song in the new world blackbirds. Behav Ecol 20:967–977
Riebel K, Hall ML, Langmore NE (2005) Female songbirds still struggling to be heard. Trends Ecol Evol 20:419–420
Rivera-Cáceres KD (2015) Plain wrens Cantorchilus modestus zeledoni adjust their singing tempo based on self and partner’s cues to perform precisely coordinated duets. J Avian Biol 46:361–368
Rogers AC, Langmore NE, Mulder RA (2007) Function of pair duets in the eastern whipbird: cooperative defense or sexual conflict? Behav Ecol 18:182–188
Searcy WA (1996) Sound-pressure levels and song preferences in female red-winged blackbirds (Agelaius phoeniceus) (Aves, Emberizidae). Ethology 102:187–196
Seddon N, Tobias JA (2006) Duets defend mates in a suboscine passerine, the warbling antbird (Hypocnemis cantator). Behav Ecol 17:73–83
Shuler JB (1965) Duet singing in the Carolina wren. Wilson Bull 77:405–405
Slater PJB, Mann NI (2004) Why do the females of many bird species sing in the tropics? J Avian Biol 35:289–294
Spencer A (2012) Female song in canyon wrens, with notes on vocal repertoire. Colorado Birds 46:268–276
Taff CC, Littrell KA, Freeman-Gallant CR (2012) Female song in the common yellowthroat. Wilson J Ornithol 124:370–374
Templeton CN, Rivera-Cáceres KD, Mann NI, Slater PJB (2011) Song duets function primarily as cooperative displays in pairs of happy wrens. Anim Behav 82:1399–1407
Templeton CN, Mann NI, Rios-Chelen AA, Quiros-Guerrero E, Garcia CM, Slater PJB (2013) An experimental study of duet integration in the happy wren, Pheugopedius felix. Anim Behav 86:821–827
Tobias JA, Sheard C, Seddon N, Meade A, Cotton AJ, Nakagawa S (2016) Territoriality, social bonds, and the evolution of communal signaling in birds. Front Ecol Evol 4:74
Warning N, Benedict L (2015) Overlapping home ranges and microhabitat partitioning among canyon wrens (Catherpes mexicanus) and rock wrens (Salpinctes obsoletus). Wilson J Ornithol 127:395–401
Webb WD, Brunton DH, Aguirre JD, Thomas DB, Valcu M, Dale J (2016) Female song occurs in songbirds with more elaborate female coloration and reduced sexual dichromatism. Front Ecol Evol 4:22
Zollinger SA, Podos J, Nemeth E, Goller F, Brumm H (2012) On the relationship between, and measurement of, amplitude and frequency in birdsong. Anim Behav 84:e1–e9
Acknowledgments
The following agencies provided access to field sites: The U.S. Forest Service, CO Parks and Wildlife, Larimer Co., Boulder Co. OSMP, Denver Mountain Parks, and Fort Collins Natural Areas. We thank Andrew Spencer for allowing us the use of audio files. Henrik Brumm and two anonymous reviewers provided comments that improved the manuscript.
Funding
This work was supported by a grant from the University of Northern Colorado’s Provost Fund.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed. All procedures performed in studies involving animals were in accordance with the ethical standards of the University of Northern Colorado’s Institutional Animal Care and Use Committee (Protocol No. 1506C-LB-Birds-18).
Additional information
Communicated by H. Brumm
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Hathcock, T.J., Benedict, L. Conspecific challenges provoke female canyon wrens to sing but not to duet. Behav Ecol Sociobiol 72, 196 (2018). https://doi.org/10.1007/s00265-018-2625-0
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-018-2625-0