
Abstract
Urbanization exposes wildlife to unfamiliar environments, including novel structures and food sources. Adapting to such anthropogenic conditions may require superior innovation and problem-solving skills (e.g., for navigating, foraging). Human presence in urban areas is a particular biotic challenge that may impact problem-solving capabilities in wildlife. Urban animals may be superior problem solvers in the face of human disturbance, due to familiarity with—and reduced fear of—humans. Alternately, rural animals may be better innovators if heightened fear responses from unfamiliar humans facilitate problem solving. Here, we studied the relationship between human disturbance, urbanization, and the ability to solve a novel foraging problem in wild-caught juvenile house finches (Haemorhous mexicanus). This songbird is a successful urban dweller and native of the American desert southwest. In captivity, juvenile finches from urban and rural populations were presented with a novel foraging task (sliding a lid covering their typical food dish) and then exposed to regular periods of high or low human disturbance over several weeks before again being presented with the task. We found that rural, high-disturbance birds showed reduced task performance compared to highly disturbed urban finches. This is consistent with the hypothesis that urban birds are less affected by human disturbance due to habituation or adaptation. Additionally, we found that the best behavioral predictor of solving success was related to visual inspection of the problem and that urban low-disturbance birds exhibited this behavior more than rural high-disturbance birds. Overall, these findings suggest that urbanization and habituation to humans predict avian response to novel problems.
Significance statement
As Earth’s landscapes become more urbanized, wildlife is exposed to many new anthropogenic challenges. In particular, animals in cities may have to locate food in unique ways and/or cope with increased human presence. We tested the effects of urbanization and human disturbance on problem solving in juveniles of a common urban and rural songbird (the house finch, H. mexicanus). We found that, when subjected to high levels of human disturbance, urban birds outperformed rural conspecifics on a novel foraging task (sliding open a lid to uncover food). We also found that task focus (i.e., average bout length of inspecting the dish) was an important predictor of problem solving. Our results suggest that acclimation to humans may be important for the superior behavioral performance of urban house finches and ultimately their colonization of many North American cities.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Animals encounter many types of problems during their lifetime, from locating food to avoiding predators to securing mates. Changing environments can further complicate these challenges by rapidly creating new problems for wildlife but can also introduce opportunities for innovation. One increasingly common form of environmental change is the transformation of natural landscapes to urban human habitat. Urban environments can present animals with unique structural environments to navigate (e.g., homes, yards, buildings; Klem 1990), food sources to locate/exploit, and potential threats to avoid (e.g., pollution, moving cars, domestic cats; Baker et al. 2008). Some species overcome these problems and persist in anthropogenic habitats, while others do not (Beissinger and Osborne 1982; Marzluff 2001; Mikami and Mikami 2014). Birds are one such taxon that contains a number of species that have successfully conquered the novel obstacles of urban life (Marzluff 2001). Urban tolerators/thrivers exhibit many traits that allow them to inhabit cities, and these can include enhanced innovation and problem-solving ability (Sol et al. 2002; Audet et al. 2016). Such traits can be especially relevant when foraging in urban environments. Urban bird feeders and human food waste (often in novel containers or underneath lids) provide foraging contexts that are comparatively rare in rural habitats. This is well-illustrated by the famous example of blue tits (Cyanistes caeruleus; family Paridae) in Europe that learned to open milk bottles left on doorsteps and drink the milk inside. This behavior quickly swept through a number of bird species in the UK and Ireland during the first half of the twentieth century (Lefebvre 1995).
In addition to being exposed to novel feeding challenges, urban birds are typically faced with greater proximity to and density of humans and thus are exposed to more regular human disturbances than rural birds. Humans may initially be perceived as threatening to wild animals, but due to our generally predictable and benign actions, wildlife may eventually habituate to our presence (Jimenez et al. 2011). In fact, there is some evidence that urban birds show less of a fear response towards humans (i.e., less likely to perceive us as a predatory threat) than their rural counterparts (Sol et al. 2011; Myers and Hyman 2016; Vincze et al. 2016). Fear of predators can hinder problem solving and innovation, as it increases vigilance and thus reduces foraging time (Breviglieri and Romero 2016; Klett-Mingo et al. 2016). This generates the hypothesis that human disturbance may negatively influence avian problem-solving ability but more so in animals for which humans are perceived as highly threatening (i.e., in rural compared to urban animals).
Problem-solving ability is often measured and tested in adult animals (Werdenich and Ludwig 2006; Sol et al. 2011; Griffin et al. 2014), but a great deal of learning and exposure to novel problems and challenges may occur at a young age. Song learning, for example, has a finite developmental window in some bird species. In these cases, juveniles must learn to sing before they reach adulthood because adults lack the ability to learn new songs (Marler 1970). Juveniles may also be superior innovators and problem solvers than adults (Morand-Ferron et al. 2011), as they are still developing and focused on learning skills for adulthood. This can be especially true in the face of novel stimuli. For example, in a study of adult and juvenile Chimango caracaras (Milvago chimango), juveniles were less neophobic, more exploratory, and better problem solvers than adults when faced with a novel foraging task (Biondi et al. 2010). Such neophilic and exploratory responses to novelty can be adaptive due to the benefit of a potential new resource. However, this can conflict with the risk of harmful consequences such as predation or the ingestion of noxious food items. Therefore, fear responses may also be adaptive, depending on the given situation (Sol et al. 2011). There is conflicting literature on exploration and neophobia in urban versus rural birds. Some studies have shown that urban birds are less neophobic and more exploratory than rural counterparts, likely due to more frequent exposure to novel situations and disturbance (Sol et al. 2011; Atwell et al. 2012). If urban birds are more acclimated to human presence, they may experience less fear in their presence than rural birds would in the same situation due to decreased perceived threat. However, other studies have found that urban birds are less exploratory and more neophobic than rural birds, perhaps as a defense against the comparatively increased complexity and risks of urban environments (Echeverria and Vassallo 2008; Bókony et al. 2012; Miranda et al. 2013). As all of these studies have involved adult animals, studies of juveniles may offer key insights into problem solving, human disturbance, and urbanization because young animals are at a stage when they must learn to balance the conflicts of exploratory behavior and neophobia in order to survive.
One bird species that has been particularly successful in both urban and rural habitats is the house finch (Haemorhous mexicanus). It is a songbird native to Mexico and the southwestern USA; they were also introduced to New York in 1940 and have expanded their range across North America in accordance with the spread of urbanization from both of these points of origin (Badyaev et al. 2012). Their natural diet is almost exclusively plant based, including seeds, fruits, and buds, and in urban environments, they frequent backyard feeders (Hill 1993). In their native range, there are well-established populations in both urban and rural habitats, making them an ideal subject for examining urban-rural differences in feeding behavior (Giraudeau et al. 2014). Also, a great deal of avian problem-solving research has involved highly “intelligent” species, such as corvids or parrots, but relatively little work has been done to explore these abilities in other avian taxa, including finches, and particularly in the context of urbanization (Emery 2006; Bókony et al. 2014; Templeton et al. 2014; Papp et al. 2015; Audet et al. 2016).
Previously, Arnold (2013) investigated the ability of wild-caught adult male house finches to solve a novel foraging problem in a lab setting, as a function of variation in urbanization and sexually selected plumage coloration. This foraging problem consisted of finding food hidden within a container by moving a sliding lid left slightly ajar with no food visible. Arnold (2013) found that there were no differences in problem-solving ability between urban and rural males. In this study, however, we used this same foraging task to further explore the problem-solving ability of juvenile wild-caught house finches, specifically as a function of an experimental manipulation of recent human disturbance as well as of urbanization. We tested birds two times, first immediately after capture (at ca. 1–2 months of age) and then after consistently subjecting them to either high or low levels of human exposure for several weeks (see “Methods”). This study is one of the first to explore novel problem-solving ability and associated behaviors (e.g., level of activity, visual inspection of the problem) of a juvenile animal, not only in an urban-rural context but also in the context of human disturbance.
Due to increased exposure to novel foraging challenges, we hypothesize that innovative problem solving is advantageous in urban birds. Thus, we predict that urban juveniles are better problem solvers and innovators than rural juveniles. We also predict that, because of more regular exposure to benign humans, urban birds should be more tolerant of human presence and thus be more likely to solve a problem in the face of human disturbance than rural birds. Additionally, we hypothesize that urban juveniles will benefit from greater neophilia and exploration as compared to rural juveniles. Therefore, we predict that urban juveniles will exhibit more behaviors in relation to the novel task and will approach the dish sooner than their rural counterparts. We also predict that the human-disturbance treatment will have less of an effect on these behaviors in urban juveniles than rural juveniles.
Methods
Field work
From 21 to 24 May 2014, we captured 82 juvenile house finches across four study sites (see more below) using seed-baited basket traps in the Phoenix metropolitan area (Giraudeau et al. 2014). Juveniles were distinguished from adults by plumage pattern but could not be sexed because they had not yet produced their sexually dimorphic adult coloration. Though we are uncertain of the exact age of the juveniles, young of the year begin fledging in mid-April and continue to fledge until August (Badyaev et al. 2012), leaving up to a month and a half of time for juveniles to familiarize with their given habitat. Two of the study sites were urban (the Arizona State University (ASU) campus in Tempe (n = 19 birds) and a residential front yard in downtown Phoenix (n = 20)) and two were rural (South Mountain Regional Park (n = 22) and Estrella Mountain Regional Park (n = 21)). These urban and rural sites were distinguished in our previous work (Giraudeau et al. 2014) based on differences in human population density and land use-land cover metrics. At capture, we banded birds with a numbered United States Geological Survey-issued metal leg band and measured their body mass to the nearest 0.01 g using a digital scale.
Housing
We housed the birds in captivity for the duration of the study. Each bird was housed individually in small wire cages (60 cm × 40 cm × 30 cm) in three indoor animal-approved rooms on the ASU campus. Birds from the four sites were spread randomly in space among the housing rooms, and the cages (on rolling racks) were rotated among the three rooms every 2 weeks to minimize room and social-contact effects. Rooms were set to a constant temperature of 25 °C and to the current outdoor light cycle. Birds were provided with tap water and a diet of black oil sunflower seeds ad libitum.
Behavioral trials
As in Arnold (2013), we removed food from the test birds’ cages at 1900 h on the evening prior to testing, to increase motivation to feed during the trial. To minimize disturbance of birds during testing, we conducted the trials in two identical animal-approved rooms separate from those used for housing. The size and configuration of the cages used for testing remained the same as those used for housing, but atop the food dish (now empty), we attached a small tin container (4 cm × 6.5 cm × 1.25 cm) that was half-filled with black oil sunflower seeds and covered with a sliding lid (Fig. 1). This lid was left slightly ajar (ca. 7 mm) to permit the bird to attempt to open it and feed, and all seeds were pushed away from the opening so as to be out of view when the bird began the trial. The experimenter set up the novel feeding dish prior to placing the bird in the cage. All birds were tested singly in their own cage, and each testing room contained two cages; thus, four total birds were tested during each trial period. The two cages in each room were separated from each other by a hanging sheet to minimize disturbance and to prevent them from observing each other’s behavior during the trial. The two birds were still able to hear each other, but from our video observations, it did not appear that vocal communication between test subjects influenced problem-solving success or other behaviors during the trial.

a A caged house finch during a problem-solving trial. All cages were configured with the metal dish left slightly ajar in the bottom left corner, an adjacent perch, and a cup of water in the bottom right corner. b Metal dish with sliding lid which concealed sunflower seed during the trials. Finches were tasked with inserting the bill in the opening and then sliding the lid towards the right. Visit http://youtu.be/4WrV3eAX0nk for video examples of finches successfully solving the problem
We conducted the first set of problem-solving trials (i.e., pre-human-disturbance) from 31 May–4 June 2014, ca. 1.5 weeks after capture from the wild (mean = 10.17 days, SD = 2.317 days). There was a slight delay in post-capture testing because, during the first week after capture, all birds participated in another behavioral trial for a concurrent study (behavioral response to an approaching human); however, each bird received the same testing procedure during that time. Our problem-solving trials lasted 55 min each and were run from 0730 and 1330 h each day (i.e., when the birds are most active; sensu Arnold 2013). The time between capture from the subject’s home cage and placement in the testing cage was approximately 15 min, as birds were weighed immediately prior to testing. Humans were never present in the room during problem-solving trials. We video-recorded each individual bird during each trial to quantify its behavior and to record the success and speed of solving the problem (see more below). After testing, we returned each bird and cage to its proper housing location and restored ad libitum access to food and water.
The second set of trials was conducted 5 weeks later (7–12 July 2014), after birds had been subjected to our human-disturbance treatments. The time between the last disturbance treatment and trial 2 for each individual varied from 5 to 10 days. Half the birds were randomly assigned to receive a treatment that consisted of 1 hour of a person silently walking in a circle around the housing room, administered every other day until the week before the second set of trials was run. The birds that did not receive the treatment still received some human disturbance (i.e., the same amount of regular animal husbandry that the human-disturbed birds also received) and thus we consider them to be a low-disturbance (as opposed to “no disturbance”) group. Standard husbandry consisted of daily feeding, watering, and cleaning, resulting in roughly 30 min of human presence in each housing room per day; also, each bird was captured and handled for ca. 5 s every other week as they were transferred to a clean cage. During the human-disturbance treatment, we observed that birds would generally hop and fly around the cage with high frequency, make attempts to escape the cage (i.e., by probing between the cage bars with the head), and rarely stopped to eat or drink, suggesting that the treatment created an effective human disturbance to the birds. The person conducting the problem-solving tests was involved in the aforementioned, separate human-approach experiment but was not the “approaching human.” They were also one of nine people conducting husbandry and administering the human-disturbance treatments.
During this year and season of study, there was an unusually large outbreak of canarypox virus (genus Avipoxvirus) in wild finches, and if birds showed visible signs of illness (i.e., lesions on exposed skin) at capture, we did not include them in the study, meaning that no birds in the pre-disturbance trials showed pox symptoms. However, at the time of post-disturbance testing, seven birds (two rural, five urban) showed signs of mild infection (i.e., small pox lesions). We found (using Cox’s proportional hazards models; see below for further explanation) that pox infection had no significant effect on problem-solving ability in these trials (likelihood ratio test: χ 2 = 1.44, df = 1, p > 0.23); thus, this variable was not further considered in statistical analyses.
Behavioral data
From the video recordings of each trial, we used the software program CowLog (Hänninen and Pastell 2009) to quantify the number, duration, or latency of various behaviors relevant to the novel feeding problem (e.g., eyeing the food dish, pecking at the dish, stepping on the dish, standing next to the dish, and hopping/flying in the cage; Table 1). MOC was the sole video observer and watched all of the videos in a random order and blind to the site of origin of each bird. Most central to our study hypotheses, we examined success and latency to solve the problem, where “solving” was defined as the bird actively sliding the dish lid laterally and consuming the hidden food inside. Solving latency was calculated from the start of the trial, just after the experimenter placed the bird in the cage and left the room. We had no instances where a bird unintentionally opened the dish (i.e., by colliding with or stepping on it), as the lid was too difficult to open unless the bird directly pushed it with its beak. We also quantified each bird’s proportion of time spent hopping and flying in the cage (i.e., general level of activity), which may indicate an animal’s level of fear or ability/willingness to focus on the foraging task (Table 1). For videos of trials in which the bird solved the problem, behavioral data were collected from the start of the trial up to the time the problem was solved, regardless of how long that took. For videos of trials in which the bird did not solve the problem, we collected data from the first 35 min of each trial because the vast majority of birds that solved the task did so within this time period (trial 1 mean = 22 min, four birds after 35 min; trial 2 mean = 24 min, two birds after 35 min). This meant that the amount of time observing birds that solved the problem was roughly similar to that spent observing birds that failed to solve the problem.
Statistical analyses
Seven video files from trial 2 were lost prior to data analysis, and 13 birds were unable to participate in trial 2 due to death or illness, meaning that we had to exclude data from 20 birds total from repeated-measures analyses (i.e., we had behavioral data from two trials for 62 of the 82 total birds - 36 rural birds and 26 urban birds).
All statistical analyses were performed in the R computing environment (version 3.1.2). To test the hypothesis that human disturbance, urbanization, and their interaction are related to problem-solving success, we used mixed Cox’s proportional hazards models, found in the R package “coxme” available from the Comprehensive R Archive Network (R Core Team 2013). In the model-building process, we set solve latency (in seconds) as the response variable and the interaction between three variables and associated lower-order terms as predictors: urbanization, human-disturbance level, and trial (pre- vs. post-treatment). We also controlled for the variation in time between trials 1 and 2, variation in the time of day during testing and percent change in mass (both of which might indicate possible variation in hunger). We controlled for individual effects (due to re-testing across trials) by adding individual identity as a random effect in the model. Birds that did not solve the problem were censored at the end of the trial (3300 s). Because no high disturbance, rural birds solved the foraging problem during trial 2, this caused separation in the data, which forced the full model to unreliably estimate the hazards. However, in this scenario, the creators of this statistical package assure that the model log likelihood estimates remain reliable (Therneau and Grambsch 2000; Therneau 2015). This allowed us to test the significance of the highest-order term (habitat*trial*disturbance) with a likelihood ratio test against a model containing all predictors except this three-way interaction.
To examine potential behavioral correlates of our observed patterns of problem-solving success, we conducted a principal component analysis (PCA) on the proportion of time, average bout duration, and frequency of aforementioned focal behaviors (e.g., dish eyeing, pecking, stepping). After examining scree plots (Jackson 1993), we retained two PCs that together explained 55.4% of all variation in measured behavior (see Table 2 for variable loadings and descriptions). PC1 loaded strongly and positively with the proportion and frequency of many behaviors associated with the dish, but not bout durations of the behaviors. Additionally, PC1 loaded strongly negatively with latency to approach the dish, and we therefore considered this to be a “dish visitation” PC (i.e., higher values indicate that birds approached the dish sooner and visited the dish frequently but did not stay long or display long bouts of dish-inspection behaviors such as eyeing; Table 2). PC2 loaded strongly and negatively on the proportion of time and average bout duration of hopping/flying, and the frequency of dish visits (Table 2). However, it was strongly and positively loaded with the average bout duration of many behaviors, such as time at the dish, stepping on the dish, dish peck frequency, and most strongly eyeing the dish. Latency to approach the dish did not load strongly in this component. Therefore, we considered this the “task focus” PC (i.e., higher values indicate more focus on the task). We then carried out two linear mixed model regressions, with PCs as response variables. The initial models contained urbanization, trial, human-disturbance level, and solving success as predictors, and we removed non-significant highest-order (p > 0.05) interaction terms. We controlled for the effects of individuals across trials by including bird ID as a random effect in each model. We followed a similar procedure for assessing how body mass differed among trials, treatments, and habitats. Finally, to test the behavioral predictors of solving success, we created a mixed Cox model with PC1 and PC2 as predictors and with individual ID as a random effect. All post hoc comparisons were performed with Tukey contrasts performed on the final model. All tests were two tailed.
Results
Problem-solving success and temporal variation
Overall, finches solved the foraging problem with similar success in the pre- (26% success; 16/62 birds) and post-disturbance (18%; 11/62 birds) trials (Table S1). Thus, there was no evidence of a change in solving success with age/experience. Of the 62 birds that participated in both trials, 10 solved it only during trial 1, 5 solved it only during trial 2, 6 solved it both times, and 42 never solved it. Also, out of the six birds that solved the problem twice, only three of them solved the problem faster the second time, indicating that previous experience with solving the task did not improve solving speed.
Effects of urbanization and human disturbance on problem-solving success and its behavioral predictors
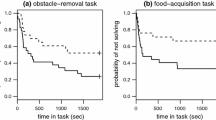
We found that solving success was significantly affected by the human-disturbance treatment and the interaction between disturbance, urbanization, and trial (Fig. 2; Table 3). In particular, rural birds that experienced high disturbance performed much worse on the problem-solving task; in fact, unlike urban birds, not a single rural bird (out of 18 tested) solved the novel foraging problem after they were subjected to the high human-disturbance treatment (Fig. 2). It should be noted that, despite randomization of groups prior to treatment, we found a bias in the solving ability of the high- and low-disturbance groups in trial 1 prior to any disturbance treatment, with the high-disturbance group performing worse than the low-disturbance group. However, our analysis nevertheless indicated a significant three-way interaction, indicating that urbanization and human disturbance still had a significant effect on solving success, even with the initial natural bias (Fig. 2, Table 3). None of the other variables (time of day, time between trials 1 and 2, percent mass change) significantly predicted problem-solving success (Table 3)
.

Problem-solving success for a trial 1 and b trial 2 as a function of urbanization (rural = black, urban = gray) and human-disturbance level (low disturbance = dotted lines, high disturbance = solid lines) in house finches. High-disturbance rural birds were far less likely to solve the task than low-disturbance rural birds. Numbers at the tips of lines show the number of birds that solved the problem (numerator) and the total number of birds in the group (denominator)
We found that PC1 (“dish visitation”) was not significantly associated with solving success (Cox’s proportional hazards: hazard ratio = 0.94, z = −0.58 ± 0.1, p = 0.56; Table 4). However, birds with higher PC2 (“task focus”) values were significantly more likely to solve the problem (hazard ratio = 1.51 ± 0.13, z = 3.19, p = 0.001).
Variation in behavioral parameters and mass based on trial, habitat, and disturbance treatment
We found a significant habitat*trial effect on body mass (Table S2). Urban and rural birds did not differ in their body mass in the pre-disturbance trials (Tukey’s HSD: p = 0.23), but urban birds were less massive than rural birds at the time that post-disturbance trials were run (p = 0.005; urban trial 1 = 18.78 ± 0.22 (mean ± SE), rural trial 1 = 18.43 ± 0.15, urban trial 2 = 17.46 ± 0.23, rural trial 2 = 18.27 ± 0.20). There was no significant three-way interaction, suggesting that the mass change of urban vs. rural birds was not sensitive to disturbance treatment type (χ 2 = 0.47, df = 1, p = 0.49). High- and low-disturbance birds did not differ in body mass at any time point (Table 4). All other effects were non-significant (all p > 0.05).
We found several other differences in finch behavior during trials as a function of habitat, trial number, and/or disturbance treatment (Table S2). Birds had higher PC1 values (i.e., spent more time at the dish) during trial 1 compared to trial 2 (Fig. 3, Table 4). Additionally, urban birds had higher “dish visitation” scores than rural birds (Fig. 3, Table 4). There was a significant three-way trial*habitat*disturbance interaction on PC2 values (“task focus”), suggesting that the human-disturbance treatment had different effects on task focus of urban and rural birds during the experiment (Fig. 3, Table 4). However, the effect was small and pairwise post hoc analyses show no significant differences among those groups (Fig. 3).

Least square means ± SE of PC1 (“dish visitation”) and PC2 (“task focus”) values as a function of human-disturbance treatment and urbanization during both trials. a PC1 values for both urban and rural groups significantly decreased from trial 1 to trial 2, and urban birds had greater PC1 values (lower dish attentiveness, i.e., interactions with the dish) than rural birds. b PC2 values were similar for all groups during trial 1. High levels of human disturbance caused urban birds to generally decrease in PC2 values, whereas low levels of disturbance caused them to increase in PC2 values
Discussion
Here, we show that human disturbance negatively impacts problem solving in juvenile house finches and does so differentially in rural- v. urban-caught juveniles. Specifically, high levels of human disturbance differentially impaired problem-solving ability in rural birds. This suggests that city birds have habituated and/or adapted to typically benign human presence but that rural birds (with less frequent interactions with humans) may still perceive humans as threatening and that the resulting disturbance interferes with their ability to solve a problem. This result is consistent with previous studies in which urbanization predicted differences in neophobia and exploratory behavior (Sol et al. 2011; Myers and Hyman 2016; Vincze et al. 2016) and also supports the idea that fear inhibits problem solving (Breviglieri and Romero 2016; Klett-Mingo et al. 2016). A recent study by Papp et al. (2015) found no overall difference between urban and rural house sparrows (Passer domesticus) in problem-solving ability, which we found here as well in juvenile house finches (also see Arnold 2013 in adults). However, in a recent study, urban bullfinches (Loxigilla barbadensis) exhibited greater problem-solving success than their rural counterparts (Audet et al. 2016), and the same has been found in common mynas (Acridotheres tristis; Sol et al. 2011) and great tits (Parus major; Preiszner et al. 2017). Thus, it may be that urban-rural variation in problem solving is species dependent or task dependent. However, our study differed from all of these studies by including a human-disturbance treatment, which evoked differences between urban and rural finches in problem-solving behavior under a potentially stressful situation. However, we didn’t measure physiological stress (e.g. corticosterone) per se in this study, so we suggest that an avenue for further study is the relationship between stress physiology and problem solving, urbanization, and human disturbance.
We also quantified various behaviors exhibited by our tested birds, with the aim of uncovering potential behavioral correlates of human disturbance and problem-solving ability. We especially considered the behaviors that occurred in association with our novel food dish/lid (e.g., approaching, pecking, eyeing), with the idea that overcoming neophobia and increasing exploratory behavior may be linked to solving a novel foraging problem (Biondi et al. 2010). Although we did not directly quantify neophobia and exploration per se, we did find that some dish-related behaviors were associated with a novel object in trial 1 and a still highly unfamiliar object in trial 2. We considered the foraging task still highly unfamiliar in trial 2 because our results indicate that previous exposure to the dish in trial 1 did not affect problem solving in trial 2. This required that the birds overcome any neophobia they might experience in order to explore the novel foraging opportunity. In both trials, we found that urban birds associated with the dish significantly more so than rural birds, regardless of disturbance level, which is consistent with the hypothesis that urban birds (that were more likely to solve the problem following disturbance) are more exploratory and less neophobic than rural birds. We also found that rural birds had lower PC1 scores than urban birds, which generally loaded with behaviors suggestive of neophobia (e.g., frequency of hopping/flying and eyeing the dish). However, this component was also strongly and negatively loaded with approach latency, suggesting that rural birds were slower to approach the novel foraging problem. Thus, there is only some indication that urban birds are more neotolerant and exploratory overall than rural birds. One might suspect that hunger level could also affect a bird’s interest in the novel foraging problem, but we found that body mass (a proxy for nutritional state) had no effect on solving success (also see prior evidence in Arnold (2013) that fasting time did not affect problem-solving success in finches).
We found that, based on the two PCs used in the study, dish-visitation behaviors (PC1) were not necessarily predictive of problem-solving ability. Instead, variables that strongly loaded onto PC2, such as the average bout duration of dish eyeing, seemed more important (i.e., time spent focusing on the task in one sitting increased task solvability). First, we found that finches under low disturbance and that had solved the problem had greater task focus relative to high-disturbance solvers. This suggests that disturbance reduced investment into task assessment and ultimately problem solving. Also, urban solvers had much greater task focus than urban non-solvers, though this was not true among rural birds. From trial 1 to trial 2, finches changed their task focus as a function of both urbanization and human-disturbance treatment. However, post-hoc tests revealed no significant differences among individual test groups. This provides some evidence that rural birds seek to overcome problems in the face of stressors through different mechanisms than those used by urban birds but are perhaps unable to overcome such challenges (as indicated by their poor problem-solving success). Our results fit with those of Valcarcel and Fernandez-Juricic (2009), who showed that urban house finches, while more responsive to potential predation threats, foraged with greater pecking intensity than rural counterparts. Ultimately, these results show that exposure to perceived threats can alter the expression of behaviors necessary for innovative problem solving and assessment of foraging challenges. Moreover, this gives further evidence that urban finches are able to acclimate to human exposure during a very short and early developmental window.
When comparing our results to those in Arnold’s (2013) previous study of foraging problem solving in adult male house finches, we found similar rates of problem solving between adults and juveniles (23% in adults tested only once, 28% in trial 1 for the juveniles in this study), demonstrating that there was no improved performance with age/experience. The same was true when we compared problem-solving ability within individuals in this study and failed to find an increase in solving success in trial 2 v. 1. Though we cannot necessarily draw strong conclusions about age, given that we conducted the adult and juvenile studies in two different years and that our juveniles had only aged a few weeks at the point of re-testing, this result is consistent with other studies of wild avian and non-avian taxa that have found problem solving to be independent of age (or associated experience; Cole et al. 2010; Griffin et al. 2014; Hopper et al. 2014; but see Biondi et al. 2010; Morand-Ferron et al. 2011; Thornton and Lukas 2012). The lack of improvement in solving success between trials indicates that learning did not take place and that memory did not contribute to problem solving. This result may also be expected given the presumed limited recall capabilities of fringillid finches (e.g., which have a small hippocampus, the major brain region involved in long-term memory; Krebs et al. 1989; Harvey and Krebs 1990) and the fact that our study birds were exposed to the brief task only once 5 weeks earlier. Thus, birds that had previously solved the task would likely not have an advantage over those that had failed in the previous trial. Future studies may best examine urban-rural variation in learning/recall using an expanded task-training and multiple-testing paradigm, which will permit a thorough investigation of both problem-solving plasticity and adaptation in the city.
In summary, we found that urbanization can influence avian behavioral responses to novelty and to foraging challenges specifically in high-disturbance situations. When solving a foraging problem, urban birds were better able to cope with the effects of previous human presence than their rural counterparts. As urbanization rapidly increases, wildlife’s ability to acclimate and adapt to the challenges of urban living will become increasingly relevant, and an understanding of how urban dwellers succeed in these unique human environments will be key.
References
Arnold S (2013) Beauty and brains: redder rural finches have it all. Undergraduate Honor’s Thesis, Arizona State University
Atwell JW, Cardoso GC, Whittaker DJ, Campbell-Nelson S, Robertson KW, Ketterson EW (2012) Boldness behavior and stress physiology in a novel urban environment suggest rapid correlated evolutionary adaptation. Behav Ecol 23:960–969
Audet J, Ducatez S, Lefebvre L (2016) The town bird and the country bird: problem solving and immunocompetence vary with urbanization. Behav Ecol 27:637–644
Badyaev AV, Belloni V, Hill GE (2012) House finch (Carpodacus mexicanus). The Birds of North America Online, Cornell Lab of Ornithology, http://bna.birds.cornell.edu/bna/species/046
Baker PJ, Molony SE, Stone E, Cuthill IC, Harris S (2008) Cats about town: is predation by free-ranging pet cats Felis catus likely to affect urban bird populations? Ibis 150:86–99
Beissinger SR, Osborne DR (1982) Effects of urbanization on avian community organization. Condor 84:75–83
Biondi LM, Bo MS, Vassallo AI (2010) Inter-individual and age differences in exploration, neophobia and problem-solving ability in a Neotropical raptor (Milvago chimango). Anim Cogn 13:701–710
Bókony V, Kulcsar A, Toth Z, Liker A (2012) Personality traits and behavioral syndromes in differently urbanized populations of house sparrows (Passer domesticus). PLoS One 7:e36639
Bókony V, Lendvai AZ, Vagasi CI, Patras L, Pap PL, Nemeth J, Vincze E, Papp S, Preiszner B, Seress G, Liker A (2014) Necessity or capacity? Physiological state predicts problem-solving performance in house sparrows. Behav Ecol 25:124–135
Breviglieri CPB, Romero GQ (2016) Snakes and forbidden fruits: non-consumptive effects of snakes on the behaviors of frugivorous birds. Behav Ecol Sociobiol 70:777–783
Cole EF, Cram DL, Quinn JL (2010) Individual variation in spontaneous problem-solving performance among wild great tits. Anim Behav 81:491–498
Echeverria AI, Vassallo AI (2008) Novelty responses in a bird assemblage inhabiting an urban area. Ethology 114:616–624
Emery NJ (2006) Cognitive ornithology: the evolution of avian intelligence. Philos T Roy Soc B 361:23–43
Giraudeau M, Mousel M, Earl S, McGraw KJ (2014) Parasites in the city: degree of urbanization predicts poxvirus and coccidian infections in house finches (Haemorhous mexicanus). PLoS One 9:e86747
Griffin AS, Diquelou M, Perea M (2014) Innovative problem solving in birds: a key role of motor diversity. Anim Behav 92:221–227
Hänninen L, Pastell M (2009) CowLog: open source software for coding behaviors from digital video. Behav Res Methods 41:472–476
Harvey PH, Krebs JT (1990) Comparing brains. Science 249:140–146
Hill GE (1993) Geographic variation in the carotenoid plumage pigmentation of male house finches (Carpodacus mexicanus). Biol J Linn Soc 49:63–86
Hopper LM, Price SA, Freeman HD, Lambeth SP, Schapiro SJ, Kendal RL (2014) Influence of personality, age, sex, and estrous state on chimpanzee problem-solving success. Anim Cogn 17:835–847
Jackson DA (1993) Stopping rules in principle component analysis: a comparison of heuristical and statistical approaches. Ecology 74:2204–2214
Jimenez G, Lemus JA, Melendez L, Blanco G, Laiolo P (2011) Dampened behavioral and physiological responses mediate birds’ association with humans. Biol Conserv 144:1702–1711
Klem D Jr (1990) Collisions between birds and windows: mortality and prevention. J Field Ornithol 61:120–128
Klett-Mingo JI, Pavon I, Gil D (2016) Great tits, Parus major, increase vigilance time and reduce feeding effort during peaks of aircraft noise. Anim Behav 115:29–34
Krebs JR, Sherry DF, Healy SD, Perry VH, Vaccarino AL (1989) Hippocampal specialization of food-storing birds. P Natl Acad Sci USA 86:1388–1392
Lefebvre L (1995) The opening of milk bottles by birds—evidence for accelerating learning rates, but against the wave-of-advance model of cultural transmission. Behav Process 34:43–53
Marler P (1970) A comparative approach to vocal learning: song development in white-crowned sparrows. J Comp Physiol Psychol 71:1–25
Marzluff JM (2001) Worldwide urbanization and its effects on birds. In: Marzluff JM, Bowman R, Donnelly R (eds) Avian ecology and conservation in an urbanizing world. Springer, New York, pp 19–47
Mikami OK, Mikami K (2014) Structure of the Japanese avian community from city centers to natural habitats exhibits a globally observed pattern. Landscape Ecol Eng 10:355–360
Miranda AC, Schielzeth H, Sonntag T, Partecke J (2013) Urbanization and its effects on personality traits: a result of microevolution or phenotypic plasticity? Glob Change Biol 19:2634–2644
Morand-Ferron J, Cole EF, Rawles JEC, Quinn JL (2011) Who are the innovators? A field experiment with 2 passerine species. Behav Ecol 22:1241–1248
Myers RE, Hyman J (2016) Differences in measures of boldness even when underlying behavioral syndromes are present in two populations of the song sparrow (Melospiza melodia). J Ethol 34:197–206
Papp S, Vincze E, Preiszner B, Liker A, Bókony V (2015) A comparison of problem-solving success between urban and rural house sparrows. Behav Ecol Sociobiol 69:471–480
Preiszner B, Papp S, Pipoly I, Seress G, Vincze E, Liker A, Bókony V (2017) Problem-solving performance and reproductive success of great tits in urban and forest habitats. Anim Cogn 20:53–63
R Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna http://www.R-project.org/
Sol D, Timmermans S, Lefebvre L (2002) Behavioral flexibility and invasion success in birds. Anim Behav 63:495–502
Sol D, Griffin AS, Bartomeus I, Boyce H (2011) Exploring or avoiding novel food resources? The novelty conflict in an invasive bird. PLoS One 6:e19535
Templeton CN, Laland KN, Boogert NJ (2014) Does song complexity correlate with problem-solving performance in flocks of zebra finches? Anim Behav 92:63–71
Therneau TM (2015) A package for survival analysis in S, version 2.38, http://CRAN.R-project.org/package=survival
Therneau TM, Grambsch PM (2000) Modeling survival data: extending the cox model. Springer, New York
Thornton A, Lukas D (2012) Individual variation in cognitive performance: developmental and evolutionary perspectives. Philos T Roy Soc B 367:2773–2783
Valcarcel A, Fernandez-Juricic E (2009) Antipredator strategies of house finches: are urban habitats safe spots from predators even when humans are around? Behav Ecol Sociobiol 63:673–685
Vincze E, Papp S, Preiszner B, Seress G, Bokony V, Liker A (2016) Habituation to human disturbance is faster in urban than rural house sparrows. Behav Ecol 27:1304–1313
Werdenich D, Ludwig H (2006) A case of quick problem solving in birds: string pulling in keas, Nestor notabilis. Anim Behav 71:855–863
Acknowledgments
We would like to thank Ellen Brooks, Kali Fardell, Megan Ipson, Krisjanis Malins, Sarah Shirota, Autumn Tullock, and Jeremiah Wetherby for their help in capture and care of the birds, Arizona State University Department of Animal Care and Technologies for housing the birds, and three anonymous referees for their constructive comments on an earlier version of this paper. This work was supported by Barrett, the Honors College of ASU and the ASU School of Life Sciences Undergraduate Research Grant. This material is based upon work supported by the National Science Foundation under grant number BCS-1026865, Central Arizona-Phoenix Long-Term Ecological Research (CAP LTER).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
All procedures performed in studies involving animals were in accordance with the ethical standards of the institution or practice at which the studies were conducted (Arizona State University). All birds were ethically treated at each step, including capture, housing, treatment, testing, and release. At the end of the study, all surviving birds were released at their original sites of capture. All applicable international, national, and/or institutional guidelines for the care and use of animals were followed during this study.
Additional information
Communicated by N. Clayton
Electronic supplementary material
ESM 1
(DOCX 18.8 kb).
Rights and permissions
About this article

Cite this article
Cook, M.O., Weaver, M.J., Hutton, P. et al. The effects of urbanization and human disturbance on problem solving in juvenile house finches (Haemorhous mexicanus). Behav Ecol Sociobiol 71, 85 (2017). https://doi.org/10.1007/s00265-017-2304-6
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-017-2304-6