
Abstract
Many prey species mob predators to drive them away, thereby reducing their immediate and future predation risk. Given that mobbing is risky, it may also serve as an opportunity for males to advertise their phenotypic quality to females; however, this idea remains untested. We tested this hypothesis with a field experiment in south-eastern Brazil that assessed the response of sexually dimorphic bird species to models of two diurnal owls: a ferruginous pygmy owl (Glaucidium brasilianum), which mainly eats small birds, and a burrowing owl (Athene cunicularia), which mainly eats invertebrates and thus poses a low risk to birds. Across 19 bird species, the mobbing intensity was higher when facing the less-dangerous owl, and more males engaged in predator mobbing than females. The mobbing intensity of males was higher with a larger number of conspecific females present. This finding indicates that males may use mobbing to display their phenotypic quality to females, suggesting that predator mobbing may be influenced by sexual selection.
Significance statement
Predation is an important evolutionary force, often leading to an evolutionary arms race between predators and their prey. A puzzling form of prey-predator interactions is predator mobbing. In a wide range of species, prey individuals approach predators and show characteristic visual and acoustic displays. The primary function of mobbing is to drive the predator away; however, it may also serve as an opportunity to advertise phenotypic quality to conspecifics. Field experiments showed that the mobbing intensity of males increased with the number of conspecific females in the audience, suggesting that female choice may influence the evolution of mobbing behavior.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Predation is a key cause of mortality in many species, and accordingly, prey species have evolved a number of anti-predatory defenses, such as mimicry, camouflage, predator mobbing, or feigning death (Caro 2005). Prey should benefit from adjusting their responses according to the risk posed by the predator. Indeed, field studies have shown that birds behave differently in response to different predators depending on the risk they pose (Griesser 2009; Motta-Junior and Santos-Filho 2012).
Mobbing is a widespread anti-predatory behavior, which consists of a prey approaching a potential predator to harass it by giving calls, swooping over it, or even physically attacking it. While mobbing is costly (Dugatkin and Godin 1992; Krama and Krams 2005) and potentially lethal (Sordahl 1990), it has been suggested to be adaptive (Curio et al. 1978b). Predators often move away when being mobbed, reducing the immediate and future risk of an attack (Pavey and Smyth 1998). In addition, predator mobbing may serve as an opportunity to receive support during future predator-mobbing events (Krams et al. 2008) or as a social learning opportunity for juveniles (Griesser and Suzuki 2016) to learn to recognize predators (Griesser and Suzuki 2017).
Moreover, mobbing may serve as a costly signal (Maklakov 2002) of phenotypic quality to conspecifics (Arnold 2000), especially potential mates (Curio et al. 1983; Regelmann and Curio 1986). In many species, males are generally more aggressive than females (Francis et al. 1989; Bard et al. 2002) and mob predators more vigorously than females (Curio et al. 1983; Maklakov 2002; Griesser and Ekman 2005). However, it remains unclear why males and females differ in their mobbing behavior, and whether this behavior may be influenced by sexual selection.
Here, we investigate whether the presence of females influences the mobbing intensity of males in 19 bird species. We exposed birds to two sympatric owl species that vary in their risk to birds and measured the size of the mobbing assembly and the mobbing intensity of all individuals that joined the mobbing assembly. We tested the following two non-exclusive hypotheses: (i) if birds recognize the risk posed by a predator and mobbing aims at reducing the immediate risk, the birds will mob a dangerous predator more intensely than a low-risk predator and (ii) if mobbing is a sexually selected behavior, males will mob more intensely when more conspecific females are present, using it as an opportunity to advertise their phenotypic quality.
Material and methods
Study site
This study was conducted on Cauaia Ranch, Minas Gerais State, south-eastern Brazil (19° 28′ S 44° 01′ W) between February 2011 and February 2012, encompassing both the breeding season (October to March) and the non-breeding season (April to September). The landscape at the study site is dominated by a mosaic of semi-deciduous forests, Brazilian savannah areas, and agricultural fields. All the experiments were conducted in the edge of forest patches with sparse tree and bushes, providing perches to birds during mobbing but allowing good visibility of birds in all directions for at least 20 m.
Experimental procedure
We selected 18 locations that were at least 250 m apart to minimize the risk of resampling the same individuals on the same day (Bibby et al. 2012). Within each location, we placed the model on a 1.5-m pole, 2 m away from an approximately 3-m high tree. We used two models of owl species that are similar in their coloration and size but differ in their risk to birds: a ferruginous pygmy owl model (Glaucidium brasilianum, body length = 16.8 cm; 43% of its diet consists of birds (Carrera et al. 2008)), and a burrowing owl model (Athene cunicularia, body length = 22.4 cm; 95% of its diet consists of arthropods (Zilio 2006) but it occasionally eats birds (Motta-Junior 2006)). An earlier study showed that the size of predators may affect the mobbing intensity of birds (Templeton et al. 2005). Thus, using similarly sized species controls for the influence of body size on mobbing intensity.
We performed 96 experiments with each model throughout the year (five to six experiments in each location per model). The order of the experiments in each location followed a Latin square design, and we randomly chose different experimental sites within each location. To attract birds to the model and simulate the presence of a live predator, we placed a speaker below the model and played back vocalizations of the respective predator species (calling bouts of 30 s followed by 15 s silence). The observer was positioned 10–15 m from the model wearing camouflage clothes. We registered the behaviors of all present individuals and their distance to the predator during 10 min with a voice recorder. It was not possible to record data blind because our study involved observation of wild animals in the field.
We assessed the maximum mobbing intensity for each bird that mobbed the models during the experiment using a scale of mobbing intensity from 1 to 7 (following Chandler and Rose 1988; Motta-Junior and Santos-Filho 2012): (1) an individual was >10 m away from the model making visual displays, emitting warning calls, or being silent, (2) an individual was ≤10 m and >5 m away making visual displays, emitting warning calls, or being silent, (3) an individual was ≤5 m and >2 m away and being silent, (4) an individual was ≤5 m and >2 m away making visual displays and/or giving warning calls, (5) an individual was ≤2 m away and being silent, (6) an individual was ≤2 m away making visual displays and/or giving warning calls but not attacking the model, and (7) an individual was physically attacking the model. Distances between the birds and the model were assessed with the help of distance marks in trees placed in the four main cardinal directions (north, south, east, and west) before the experiment.
In total, 79 different bird species mobbed the models, but we included only 19 species that have conspicuous sexual color dimorphism to assess the response variable in our analyses (Table 1), allowing unambiguous discrimination of females and males in the field. These species belong to six different taxonomic groups which are mostly poorly studied but display a large variety of mating, parental care, and social systems (del Hoyo et al. 2015). Trochilidae (hummingbirds) are typically polygynyous and females alone care for the brood. Galbulidae (jacamars) usually have biparental brood care and males engage in courtship feeding. Picidae (woodpeckers) have biparental brood care and live solitarily, in couples or in small family groups. Thamnophilidae (antbirds) usually have biparental brood care and are family-group living. Tyrannidae (tyrant flycatchers) are presumably monogamous and pair-bonded and have biparental brood care. Thraupidae (tanagers) usually have biparental brood care and extra-pair paternity is common, and are frequently part of mixed flocks or large groups. The specific breeding and social systems have not been studied in the study populations and thus were not considered here. However, given that female choice is predominant in birds (Searcy 1979; Majerus 1986), it is expected that males use any opportunity to advertise their phenotypic qualities to their current mate, future mates, or potential extra-pair mating partners.
Statistical analyses
We used the software R for our analyses (R Core Team 2015). To test the influence of the presence of female and male conspecifics on the maximum mobbing intensity of each individual in the mobbing assemblage, we used a generalized linear mixed model using Markov chain Monte Carlo techniques in the package MCMCglmm (Hadfield 2010). We used the maximum mobbing intensity as our response variable and included the following predictors: predator model (high-risk or low-risk), interaction between the sex of the individual and the total number of conspecific females in the mobbing assemblage, interaction between the sex of the individual and the number of conspecific males in the mobbing assemblage, interaction between the sex of the individual and the number of total individuals in the mobbing assemblage (conspecifics and heterospecifics), and season (breeding or non-breeding). Given that we tested specific hypothesis based on a priori predictions, all non-significant terms were retained in the models. We included a consensus tree of a recent phylum-wide phylogeny (Jetz et al. 2012) and the location of each experiment as random factors to control for species-specific differences in their abundance and mobbing behavior and to control for potential non-independency of the data, since individuals were not marked. The MCMCglmm model resulted in balanced trace plots and an effective sample size of around 1000 for all variables. We checked the model convergence by analyzing the trace plots, the residuals, and the predicted values of the model. We checked for collinearity among the main effects through the variance inflation factor using the package “car” (Fox and Weisberg 2011). All values were <2.2, showing that collinearity among the main effects was negligible (supplementary material).
An ordinal regression model with phylogenetic control was not practicable, due to quasi-separation of the data (i.e., some clades were only represented by a single rank). Therefore, maximum mobbing intensity was analyzed using a linear mixed model. An earlier simulation study showed that ordinal variables with more than five levels can be reasonably assessed using approaches designed for continuous data (Rhemtulla et al. 2012). We note that an ordinal regression in a generalized linear mixed model without phylogenetic control yields qualitatively similar results (supplementary material).
Results
We observed at least one individual of a sexually dimorphic species in 54 experiments. Overall, 165 individuals of 19 sexually dimorphic species approached the predator models (mean number of total individuals − sexually dimorphic and non-sexually dimorphic − per experiment = 13.09, ranging from 1 to 21). The majority of the individuals that mobbed were males (n = 108), independent of the predator species. While 126 individuals mobbed the high-risk model, only 39 individuals mobbed the low-risk model. More males mobbed when exposed to both the high-risk predator model (82 males, 44 females) and the low-risk predator model (26 males, 13 females). In most experiments, no conspecific individuals were present in the mobbing assemblage, independent of the sex of the mobber (Table 2). The number of conspecific females varied between 0 and 3 (mean = 1.53), while the number of conspecific males varied between 0 and 7 (mean = 2.27).
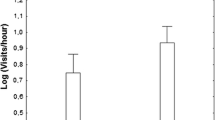
Overall, birds mobbed a low-risk predator model more intensively than a high-risk predator model (Fig. 1, Table 3), but males and females did not differ in their mobbing intensity (Table 3). We did not find a difference in mobbing intensity between the breeding and non-breeding season (Table 3). Total mobbing assemblage size (counting conspecific and heterospecific individuals) did not influence the mobbing behavior of either sex (Table 3). However, the composition of the audience influenced the mobbing behavior of males. They mobbed more intensely when more conspecific females were present (Fig. 2; Table 3), but the number of males had no influence (Table 3). The mobbing behavior of females did not depend on the audience size or composition (Table 3).

Birds mobbing intensity according to the predator stimuli. Boxplots showing the median, quartiles of, and maximum and minimum mobbing intensity per treatment

Mobbing intensity of females and males in relation to the number of conspecific females in the mobbing assemblage based on the predicted values from the model. Gray areas represent the 95% credible interval
Discussion
Mobbing is an anti-predatory behavior that primarily aims at moving the predator away, reducing both the immediate and future predation risk (Pavey and Smyth 1998). Since mobbing is risky (Sordahl 1990), it may also serve as an honest signal of phenotypic quality (Zahavi and Zahavi 1997), giving males the opportunity to display their quality to potential mates. Our results support this hypothesis, by showing that male birds mob more intensively in the presence of more conspecific females but not in the presence of more conspecific males.
This pattern could reflect two different processes: males may increase their mobbing intensity after more females join a mobbing assemblage or more conspecific females may join the assemblage due to high male-mobbing intensity. However, it is difficult to assess when males perceive the presence of females in the field, which would be critical to understand which of the two processes occurs. Regardless of the order of events, our data show that the mobbing intensity was not related to assemblage size but only to the number of conspecific females, which suggests that mobbing is influenced by sexual selection. This novel finding highlights the general importance of sexual selection and may help to better understand the evolution of anti-predatory behaviors.
It is well known that birds recognize the specific risks predators pose (Curio et al. 1978a; Griesser 2008). In contrast to our prediction, our experiments showed that birds mob a low-risk predator more intensively than a high-risk one, supporting findings from previous studies (Forsman and Mönkkönen 2001; Griesser and Ekman 2005; Griesser 2009). Thus, despite that driving away a high-risk predator is more beneficial, mobbing it is riskier, suggesting that immediate costs influence mobbing intensity.
In a wide range of species, males show more aggression than females (Bard et al. 2002). Our experiment confirms that males were more likely to mob predators (Maklakov 2002; Griesser and Ekman 2005), yet males were sensitive to the number of conspecific females in the mobbing assemblage. Earlier studies showed that males can be sensitive to their audience, adjusting sexual displays depending on the number and/or the quality of (potential) partners (Matos and McGregor 2002; Dubois and Belzile 2012). Clearly, it would be interesting to assess in future experiments whether an increased mobbing effort actually results in a higher mating success for males. If mobbing is costly signaling and therefore sexually selected, we would expect that the audience effect would be higher in polygamous species than in monogamous species (Kirkpatrick et al. 1990). Since the mating and parental care systems for the birds in our study population are not studied, we could not assess their influence on the mobbing intensity of males.
Experiments in chaffinches Fringilla coelebs showed that males increased their predator-mobbing effort during the first weeks of the breeding season (Krams and Krama 2002). This result was suggested to reflect interspecific reciprocity, but it may also reflect a benefit of advertising quality to neighbor females (to gain access to extra-pair mating opportunities) or to neighbor males (as a consequence of male-male competition). Similarly, male great tits Parus major mob predators more intensely than females (Curio et al. 1983), which may reflect that males are using mobbing to advertise phenotypic qualities. Thus, mobbing could be a sexually selected signal in other species but more experiments are needed to validate the generality of this hypothesis.
Several other factors may influence differences in mobbing behavior in general and between sexes. Temperate birds have been shown to mob more intensely during the breeding season (Shedd 1983), especially when a predator is close to their nest (Kryštofková et al. 2011). Accordingly, the breeding status can influence the mobbing intensity of males and females across species. However, we did not find a significant difference in mobbing intensity between the breeding and non-breeding season. This result may reflect that tropical and temperate birds differ in their response to nest predators or that the species included in this study breed in different micro-habitats than the ones we chose for our experiment. Furthermore, the mating status of males could influence their mobbing effort, which would be interesting to address in future studies.
To conclude, our results suggest that males across 19 species can use mobbing as an opportunity to display their phenotypic quality to females, highlighting the ubiquitous importance of sexual selection (Andersson 1994). Female choice can be influenced by the motor skills of males, which provides clues about their ability to defend a nest or forage successfully (Barske et al. 2011). Since mobbing a predator is risky, only males with good motor skills that are capable of swiftly escaping can afford to approach a predator, making predator mobbing a potentially honest signal of phenotypic quality (Zahavi and Zahavi 1997).
References
Andersson MB (1994) Sexual selection. Princeton University Press, Princeton
Arnold KE (2000) Group mobbing behaviour and nest defence in a cooperatively breeding Australian bird. Ethology 106:385–393
Bard SC, Hau M, Wikelski M, Wingfield JC (2002) Vocal distinctiveness and response to conspecific playback in the spotted antbird, a neotropical suboscine. Condor 104:387–394
Barske J, Schlinger BA, Wikelski M, Fusani L (2011) Female choice for male motor skills. Proc R Soc Lond B 278:3523–3528
Bibby CJ, Burgess ND, Hill DA (2012) Bird census techniques. Academic Press, London
Caro T (2005) Antipredator defenses in birds and mammals. University of Chicago Press, Chicago
Carrera JD, Fernandez FJ, Kacoliris FP, Pagano L, Berkunsky I (2008) Field notes on the breeding biology and diet of ferruginous pygmy-owl (Glaucidium brasilianum) in the dry Chaco of Argentina. Ornitol Neotrop 19:315–319
Chandler CR, Rose RK (1988) Comparative analysis of the effects of visual and auditory stimuli on avian mobbing behavior. J Field Ornithol 269–277
Core Team R (2015) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna https://www.R-project.org/
Curio E, Ernst U, Vieth W (1978a) Cultural transmission of enemy recognition: one function of mobbing. Science 202:899–901
Curio E, Ernst U, Vieth W (1978b) The adaptive significance of avian mobbing. Z Tierpsychol 48:184–202
Curio E, Klump G, Regelmann K (1983) An anti-predator response in the great tit (Parus major): is it tuned to predator risk? Oecologia 60:83–88
del Hoyo J, Elliot A, Sargatal J, Christie DA, de Juana E (2015) Handbook of the birds of the world alive. Lynx Edicions, Barcelona
Dubois F, Belzile A (2012) Audience effect alters male mating preferences in zebra finches (Taeniopygia guttata). PLoS One 7:e43697
Dugatkin LA, Godin J-GJ (1992) Reversal of female mate choice by copying in the guppy (Poecilia reticulata). Proc R Soc Lond B 249:179–184
Forsman JT, Mönkkönen M (2001) Responses by breeding birds to heterospecific song and mobbing call playbacks under varying predation risk. Anim Behav 62:1067–1073
Fox J, Weisberg S (2011) An {R} companion to applied regression, Second edn. Sage, Thousand Oaks CA
Francis AM, Hailman JP, Woolfenden GE (1989) Mobbing by Florida scrub jays: behaviour, sexual asymmetry, role of helpers and ontogeny. Anim Behav 38:795–816
Griesser M (2008) Referential calls signal predator behavior in a group-living bird species. Curr Biol 18:69–73
Griesser M (2009) Mobbing calls signal predator category in a kin group-living bird species. Proc R Soc Lond B 276:2887–2892
Griesser M, Ekman J (2005) Nepotistic mobbing behaviour in the Siberian jay, Perisoreus infaustus. Anim Behav 69:345–352
Griesser M, Suzuki TN (2016) Kinship modulates the attention of naive individuals to the mobbing behaviour of role models. Anim Behav 112:83–91
Griesser M, Suzuki TN (2017) Naive juveniles are more likely to become breeders after witnessing predator mobbing. Am Nat 189(1):58–66
Hadfield JD (2010) MCMC methods for multi-response generalized linear mixed models: the MCMCglmm R package. J Stat Softw 33(2):1–22
Jetz W, Thomas GH, Joy JB, Hartmann K, Mooers AO (2012) The global diversity of birds in space and time. Nature 491:444–448
Kirkpatrick M, Price T, Arnold SJ (1990) The Darwin-Fisher theory of sexual selection in monogamous birds. Evolution 44:180–193
Krama T, Krams I (2005) Cost of mobbing call to breeding pied flycatcher, Ficedula hypoleuca. Behav Ecol 16:37–40
Krams I, Krama T (2002) Interspecific reciprocity explains mobbing behaviour of the breeding chaffinches, Fringilla coelebs. Proc R Soc Lond B 269:2345–2350
Krams I, Krama T, Igaune K, Mänd R (2008) Experimental evidence of reciprocal altruism in the pied flycatcher. Behav Ecol Sociobiol 62:599–605
Kryštofková M, Haas M, Exnerová A (2011) Nest defense in blackbirds Turdus merula: effect of predator distance and parental sex. Acta Ornithol 46:55–63
Majerus MEN (1986) The genetics and evolution of female choice. Trends Ecol Evol 1:1–7
Maklakov AA (2002) Snake-directed mobbing in a cooperative breeder: anti-predator behaviour or self-advertisement for the formation of dispersal coalitions? Behav Ecol Sociobiol 52:372–378
Matos R, McGregor P (2002) The effect of the sex of an audience on male-male displays of siamese fighting fish (Betta splendens). Behaviour 139:1211–1221
Motta-Junior JC (2006) Relações tróficas entre cinco Strigiformes simpátricas na região central do Estado de São Paulo, Brasil. Rev Bras Ornitol 14:359–377
Motta-Junior JC, Santos-Filho PS (2012) Mobbing on the striped owl (Asio clamator) and barn owl (Tyto alba) by birds in southeast Brazil: do owl diets influence mobbing? Ornitol Neotrop 23:159–168
Pavey CR, Smyth AK (1998) Effects of avian mobbing on roost use and diet of powerful owls, Ninox strenua. Anim Behav 55:313–318
Regelmann K, Curio E (1986) Why do great tit (Parus major) males defend their brood more than females do? Anim Behav 34:1206–1214
Rhemtulla M, Brosseau-Liard PÉ, Savalei V (2012) When can categorical variables be treated as continuous? A comparison of robust continuous and categorical SEM estimation methods under suboptimal conditions. Psychol Methods 17:354–373
Searcy WA (1979) Female choice of mates: a general model for birds and its application to red-winged blackbirds (Agelaius phoeniceus). Am Nat 114:77–100
Shedd DH (1983) Seasonal variation in mobbing intensity in the black-capped chickadee. Wilson Bull 95:343–348
Sordahl TA (1990) The risks of avian mobbing and distraction behavior: an anecdotal review. Wilson Bull 102:349–352
Templeton CN, Greene E, Davis K (2005) Allometry of alarm calls: black-capped chickadees encode information about predator size. Science 308:1934–1937
Zahavi A, Zahavi A (1997) The Handicap Principle: A Missing Piece of Darwin’s Puzzle. Oxford University Press, Oxford
Zilio F (2006) Dieta de Falco sparverius (Aves: Falconidae) e Athene cunicularia (Aves: Strigidae) em uma região de dunas no sul do Brasil. Rev Bras Ornitol 14:379–392
Acknowledgments
We are grateful to José Hein and the employees from the Cauaia Ranch, especially Cida, Peba, and Warley. We thank Gretchen Wagner, Carel van Schaik Erik Willems, and two anonymous reviewers for the advice and comments.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Funding
FCRC received funding from Universidade Federal de Ouro Preto and Science Without Boarders/Capes (BEX 8920133). MG was supported by the Swiss National Science Foundation (PPOOP3_123520, PP00P3_150752).
Ethical statement
The study was conducted under research permits issued by the local authorities (Sistema de Autorização e Informação em Biodiversidade).
Additional information
Communicated by P. A. Bednekoff
Electronic supplementary material
ESM 1
(DOCX 23 kb)
Rights and permissions
About this article

Cite this article
da Cunha, F.C.R., Fontenelle, J.C.R. & Griesser, M. The presence of conspecific females influences male-mobbing behavior. Behav Ecol Sociobiol 71, 52 (2017). https://doi.org/10.1007/s00265-017-2267-7
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-017-2267-7