
Abstract
Biological control of cyanobacteria is a well-researched area with a central focus on laboratory-scale studies. Numerous reports have been made on algicidal isolates, with bacteria as a major component of the antagonists. The research in this review draws a brief summary of what is currently known in the area of freshwater cyanobacteria being inhibited by bacterial isolates. Proteobacteria, Bacteroidetes and Firmicutes are among the most commonly reported phyla of bacteria associated with or employed in this research area. However, there are limited reports of upscaling these control measures beyond the laboratory scale. Lytic control agents are the most commonly reported in the literature with subsequent cyanotoxin release. From a water quality perspective, this is not feasible. Based on the available literature, temperature, pH and nutrient changes have been explored in this short review as possible contributors to less optimal bacterial performance. Moreover, the investigation into optimising some of these parameters may lead to increased bacterial performance and, therefore, viability for upscaling this biological control. Through the compilation of current research, this review offers insight to live predator-prey cell interactions between cyanobacteria and algicidal bacteria.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The current research will assess the feasibility of bacteria as cyanobacterial control agents beyond the laboratory. This will be based on consolidation of the existing information on biological control of cyanobacteria, based on the reviews of Sigee et al. (1999) and Gumbo et al. (2008) as well as numerous other publications on this topic. The focus will be more specifically in the area of bacteria as a biological control agent to living cyanobacterial cells in freshwater environments (Sigee et al. 1999; Gumbo et al. 2008).
With the rise in cyanobacterial bloom occurrences and toxicity, their mitigation is a crucial area of research. Lakes from across the continents in the first world and developing countries alike have been plagued with these blooms. Not only do they pose a threat to human and animal health, but they also present a challenge in terms of water treatment facilities (Ndlela et al. 2016). A review by Paerl et al. (2016) has indicated the significance in developing mitigation measures for cyanobacterial blooms, especially since they will be more difficult to curb due to increased temperatures caused by climate change in the future (Paerl et al. 2016).
The concept of biological control is the use of natural enemies to control a target organism, and among the measures previously mentioned in a review by Sigee et al. (1999), is the use of bacteria as a means to control cyanobacterial cells. In the broader focus of biological control, a review by Verschuere et al. (2000) thoroughly investigated the use of probiotic bacteria as possible biological control agents. The key modes of action listed by bacteria as methods of biological control indicate antagonism as the more common mode of action in aquaculture (Verschuere et al. 2000).
In the study of bacteria as control agents in aquaculture, a variety of inhibitory compounds such as antibiotics, bacteriocins, siderophores and lysosomes is produced by probiotic bacteria. Whilst controlling other organisms, these compounds can have a positive growth effect as well on unicellular algae and this needs to be clearly understood prior to implementation of certain species as biological control agents (Verschuere et al. 2000).
Previous studies employing the use of bacteria in curbing cyanobacteria have been conducted primarily at laboratory scale, with focus on a dominant species within a bloom or on axenic cultures of a given cyanobacterial isolate (Nakamura et al. 2003; Choi et al. 2005; Jung et al. 2008; Shao et al. 2014; Su et al. 2016a). These studies seldom account for the mixed cultures of phytoplankton or the possible variations in temperature during the exposures under lab conditions or the fluctuations within the natural environment. A recent study has found some of the interventions in curbing cyanobacterial blooms were not as effective as reported, with a strong caution to critically evaluate these control measures among which bacteria, artificial mixing and algae were mentioned (Lürling et al. 2016).
Recent research by Demeke (2016) describes the use of metabolites from the bacteria Flexibacterium in the control of the filamentous cyanobacteria Oscillatoria and the lytic activity of the bacteria Bacillus against the cyanobacteria Aphanizomenon flos-aquae (Demeke 2016). These are just a few of many examples and show the interest and potential in the area of biological control. Over the past decade, advancements in the use of bacteria as cyanobacterial control agents are not as extensive.
Aim of research
Although not exhaustive, this research aims to establish a timeline of developments in this mode of cyanobacterial control and what is currently known, with the intention of collating pitfalls and possible future research challenges. The study is focused on the viability of this biological control and which factors could be optimised for further progress, based on the available literature.
Data collection approach
The literature used within this mini-review was collected from the following databases: Google Scholar, Scopus and EBSCOhost with the following key words: “algicidal bacteria”, “Microcystis/Oscillatoria/Cylindrospermopsis/Anabaena/Aphanizomenon inhibiting bacteria”, “biological control of cyanobacteria”, “algicidal bacteria” and “bacteria lysing cyanobacteria”, with the time frame delineated to 2000–2017. Further delineation was done based on focusing on freshwater-related studies and the lysis/suppression/control of living cyanobacterial cells by living bacterial agents, with minimal focus on microcystin degraders and bacterial metabolites employed to control whole cyanobacterial cells.
Control agents associated with cyanobacteria and the susceptibility of the target microorganisms
An extensive review (Van Wichelen et al. 2016) describes the susceptibility of Microcystis sp. to a variety of control agent ranging from viruses to fungi and bacteria. The lack of application of this form of biological control outside laboratory conditions is also mentioned. This is of concern as a number of possible control measures with living organisms have been explored with minimal upscaling opportunities, with recommendations made against upscaling beyond the lab in some studies (Kim et al. 2008).
The first challenge comes from the lack of information on the type of bacteria and interactions among species that may hinder how effective a given control agent may be under field conditions as opposed to laboratory-based findings. The diversity and intercellular interactions are numerous to be adequately accounted for. To quantify or individually explore these factors is complex and requires further intensive research to paint a more conclusive picture. Therefore, although there are numerous studies and reports of effective microorganisms applicable in laboratory studies (Choi et al. 2005; Jung et al. 2008; Kim et al. 2008; Zhang et al. 2016), none of these have been conclusively upscaled or pursued beyond the lab due to the myriad of uncertainties that present themselves within a mixed population. Another question that arises is how well do these control agents regulate their algicidal characteristics in the natural environment within a mixed microbial population? Taking a closer look at Microcystis specifically which has the most available literature and is the causative agent of the most common toxic blooms, if cyanobacteria, particularly Microcystis, present such a wide susceptibility to environmental isolates, how they are able to continue proliferating and creating toxic aquatic conditions? Table 1 presents a synopsis of the known types of control agents and the type of cyanobacterium they have a predatory or lytic impact on, exclusive of bacterial isolates from various studies (Zhang et al. 2008; Wang et al. 2011; Gao et al. 2012a; Mialet et al. 2013; Mohamed et al. 2014; Leitão et al. 2018). The use of bacteria as control agents will be discussed in more depth in this review.
Another area of interest is the form of species, such as Microcystis, as laboratory strains and their natural form. Generally, under natural conditions, these cells present as large colonial isolates with mucilage layers, surrounded by various microorganisms, which have been believed to have symbiotic relations with the cyanobacteria. This colonial form is not often observed in laboratory strains that present as unicellular, and a recent study (Geng et al. 2013) found that laboratory-grown strains are not able to revert back to colonial form after subculturing within the laboratory and they tend to form two-celled or unicellular cells, which offer reduced resilience as opposed to colonies. This differential response is also observed in lab studies when exposing both forms of cells. Therefore, one may consider the study of biological control of unicellular cells to not be fully indicative of the possible response in a natural ecosystem. An earlier study by Yang et al. (2006) found that colonial Microcystis were more successful at warding off grazing by cladocerans as opposed to the unicellular isolates, which indicates a possible mechanisms of defence that is not often noted with tested unicellular isolates employed in the laboratory. In the findings of a recent study, these cells can morph into various morphotypes of Microcystis within colonies, due to the changing environmental conditions within the waterbody. This indicates a steady shift of Microcystis species, and these are not often observed in targeted biological control efforts under controlled conditions. The study further indicates the value of this defence mechanism to a given species, hence the value in a control agent that is effective within a mixed population and also has enough host specificity to remain effective against the targeted cluster of organisms (Man et al. 2018). Moreover, more evidence of the natural prevalence of the cells as colonies has been supplied in a more recent literature review on the colony formation characteristics of Microcystis (Man et al. 2018).
Table 1 summarises the commonly reported antagonists from different trophic levels against cyanobacteria and how these predators commonly control cyanobacterial isolates.
Heterotrophic bacteria associated with cyanobacterial blooms
Apart from the known characteristics of most bacteria and their robust growth, being non-fastidious and ubiquity, they form an integral part of the diversity in algal blooms (Eiler and Bertilsson 2004). This is significant in assessing whether the commonly reported control agents fall into the phyla most commonly indicated in cyanobacterial control. More importantly, this may establish whether there is any link between these specific control agents and how closely they are observed in the cyanobacterial phycosphere. An assessment of bacterial communities found that the phyla Proteobacteria and Bacteroidetes were among the most common phyla in blooms of cyanobacteria Microcystis and Anabaena sp. These phyla were also found in various blooms in Swedish lakes (Eiler and Bertilsson 2004). Various studies have been conducted in understanding cyanobacterial blooms and, therefore, the bacterial communities associated with them. Naturally, cyanobacteria exist as co-cultures with a variety of bacterial phyla. When assessed, Shi et al. (2011) and Cai et al. (2014) found that there are aggregates of free-living bacteria as well as attached bacterial species which are within the mucilage of colonial Microcystis cells. The bacterial diversity has been found to change with the different stages of a bloom (Shi et al. 2011) and with various species (Bagatini et al. 2014). Some phyla are in higher abundance at the degradation phase of a bloom, whilst some are more represented at the start or peak of blooms. The fact that these microorganisms freely exist within bloom conditions makes them ideal control agents, although it is understood that certain classes aid the proliferation of cyanobacteria (Tian et al. 2007; Shi et al. 2009).
From the above-mentioned literature studies, Alphaproteobacteria, Betaproteobacteria and Bacteroidetes are the more closely associated and reported species associated with non-axenic cyanobacterial aggregates. This applies to studies conducted on Anabaena, Aphanizomenon, Cylindrospermopsis and Microcystis cultures. A study by Cai et al. (2014) found similar phyla associated with Microcystis, with Alphaproteobacteria most abundant in the small cell aggregates and Firmicutes most abundant in the larger cyanobacterial cell aggregates. Louati et al. (2015) reported that bacterial communities differed based on the cyanobacterial blooming at a given time and that there is a beneficial relationship between cyanobacterial attached bacteria, seen in the accelerated growth of non-axenic cyanobacteria as compared to axenic strains. Firmicutes was also reported to be in lower abundance in this study as compared to Proteobacteria and Actinobacteria. With Bacillus sp. isolates being commonly reported as algicidal bacteria, it is interesting to note that they are not abundant in the smaller cyanobacterial cell aggregates, which are in closer proximity to the cyanobacteria.
Free-living and cyanobacterial attached bacteria have been differentiated based on the aggregates they form after filtration and separation from each other. The cells found closest to the cyanobacterial cells’ mucilage are thought to have a mutualistic beneficial relation or considerable tolerance to the given species (Bagatini et al. 2014). This is supported by an earlier study conducted by Bouvy et al. (2001), where the cyanobacteria Cylindrospermopsis raciborskii did not impose a major change in biodiversity or specific species suppression in the bacterial or zooplankton community within a reservoir study in Brazil. The indication of the bacterial diversity in cyanobacterial aggregates in nature is that the more beneficial bacteria are found in closer proximity to the cyanobacteria. This may explain the difficulty in natural bloom control, as predatory isolates would be lower in abundance, with a reduced contact time to the targeted cyanobacterium.
Factors associated with cyanobacterial control that may be optimised
If bacteria are to be applied on a larger scale as control agents, some factors would require optimisation. From previous literature, this review has identified temperature, nutrients and pH as parameters that could be amended for improved bacterial performance that may be viable on a larger scale, beyond the laboratory. These parameters were easily identified as they were the most often recorded in the studies included here. Although these are not the only key factors to be considered, they are the most frequently documented.
Temperature differences and their possible implications
Numerous studies on algicidal bacteria have been conducted under laboratory conditions. The algicidal bacteria are often cultured under different nutritional and temperature conditions, slightly higher than those of the targeted cyanobacteria. This might be one of the factors to consider for increased bacterial efficiency in controlling blooms on a larger scale. There is considerable evidence that growth temperatures significantly impact the protein activity (Patke and Dey 1998) in microorganisms, with a proven increase in growth rate at temperatures beyond 30 °C for Bacillus, Escherichia coli, Salmonella and Listeria sp. strains (Membré et al. 2005). Studies on the effects of temperature on bacterial isolates have been modelled and better understood from as far back as the 1980s (Ratkowsky et al. 1982; White et al. 1991). A 2010 report assessing clinical isolates of Staphylococcus sp. also found increased biofilm populations and antimicrobial properties at higher temperature, supporting the findings that most isolated clinical bacteria are mesophilic in nature (Hajdu et al. 2010). Although a variety of factors influence the growth rate of bacteria in conjunction with temperature, a similar phenomenon of temperature influencing growth rates in a number of studies (Pietikäinen et al. 2005).
Rampelotto (2013) considered cyanobacteria as being the most resilient to temperature changes among the prokaryotes, with their discovery and viability occurring in a wide expanse of environmental conditions. Almost two decades ago, research on the effect of lowered temperatures on mesophiles and psychrophilic bacteria indicated that the affinity for sequestering or binding given substrates was lowered at decreased temperatures. There was a noteworthy difference in the cell affinity (Nedwell 1999), with respiratory rate inhibition even in psychrophiles, when temperatures were lowered beyond the optimum (Pomeroy et al. 1991). Temperature changes of 10 °C showed a distinct decrease in cell affinity for mesophiles (Nedwell 1999). Movahedi and Waites (2002) further showed that cold shock resulted in a protein expression change in sporulating Bacillus cells, compared to an earlier study showing a growth lag in vegetative cells after temperature drops of about 20 °C (Graumann and Marahiel 1999). From this research, although resilience may be increased in further generations, it is clear that drastic temperature changes are not ideal in the optimal performance of a mesophilic microorganism. To provide a bit of contrast, a study on the heat shock response of E. coli cells showed that a 12 °C temperature increase to 42 °C subsequently increased the growth rate of these cells, whilst higher temperatures of 48 °C to 55 °C indicated an initial increase and, thereafter, cell growth inhibition (Soini et al. 2005). However, it appears that temperature changes beyond 10 °C were induced in most experimental cases where bacteria are isolated to control cyanobacteria. Although the studies here include clinical isolates, the trend of lowered growth rate at lower temperatures for mesophiles is clear.
Viewing this point in the context of African waters, Ndlela et al. (2016) found water temperatures during blooms which can range from above 30 °C to as low as 10 °C. A South African study by Gumbo et al. (2010) on the biological control of Microcystis reported the isolation of algicidal Bacillus mycoides using the plaque assay, with subsequent exposure using this Bacillus as a control organism. Cultures were grown separately, with bacterial isolates at 37 °C, whilst the target cyanobacteria were kept at ambient temperature in a different medium; thereafter, the control agent was co-cultured with the cyanobacteria at a 1:1 volume ratio and observed over 6 days (Gumbo et al. 2010).
Research from Oberholster and Botha (2010) analysing the hyper scum and toxicity of Microcystis sp. in Lake Hartbeespoort showed that over a period of 6 months at the peak of the bloom, temperatures ranged between 23 and 25 °C. This is further supported by the findings of the research in Theewaterskloof, where a bloom occurred at temperatures as low as 14 °C (Oberholster et al. 2015). Another study by Oberholster et al. (2009), assessing animal mortalities to microcystins, also indicated that the average water temperature between 2005 to 2006 was 25.1 °C in Lake Hartbeespoort, over that period, with a clear indication that temperature increases of around 0.84 °C over the years due to climate change have influenced the toxicity and prevalence of cyanobacterial blooms (Oberholster et al. 2009).
In light of this information among other reports, why is the temperature of control agents seldom uniform to blooms considered in the lab-scale implementation of biological control? Listed in Table 2 are the different temperatures that bacteria and target cyanobacteria are grown under, based on previous research (Manage et al. 2000; Choi et al. 2005; Mu et al. 2007; Ren et al. 2010; Yang et al. 2012; Lin et al. 2014; Zhu et al. 2014; Su et al. 2016a, b; Zhou et al. 2016). Only research over the last few years indicates a consideration of temperature similarity and, in some cases, nutrient acclimatisation as in the case of Shao et al. (2014) in lab-scale studies. Marked in red are temperature differences that indicate a difference greater than 10 °C which may induce the previously mentioned cold shock in bacterial isolates.
This is seen in many of the lab-scale studies listed in this article, with the general approach being to enhance the predator through rich media and higher temperatures. This begs the question whether the conditions employed would result in effective biological control in the natural environment. Furthermore, how may one feasibly upscale this technology in an aquatic system without major hampering of the biodiversity? Another question that arises is whether the response to climate changes and therefore increased nuisance algal blooms is eradication, suppression or toxin removal. Although there are numerous papers reporting the use of microcystin-degrading bacteria, exclusive of live cell-to-cell interaction, which indicate the metabolites and proteins that lead to cyanobacterial death and inhibition (Ji et al. 2009; Luo et al. 2013; Yang et al. 2014), the focus of this review is on live cell interaction between the predator and the targeted cyanobacteria.
Alkalinity and acidity: the other factors in biological control
Numerous studies indicate a preference for cyanobacterial proliferation at more alkaline conditions (Gumbo et al. 2008), with a pH of as high as 10 being optimal for the proliferation of blooms (Gao et al. 2012b). A study on the mass culturing of Synechocystis for its multiple benefits in industrial applications found that higher pH conditions were primarily useful in warding off potential predators, as these maintain an advantage to grow at more alkaline conditions in comparison to other freshwater species that graze on it. Optimal growth did not differ much from neutral pH up to a pH of 10.5, which greatly limited the growth capacity of the grazer. Given that alkalinity is higher under natural bloom conditions and that over continuous culture, there is often an increased pH level created by the presence of cyanobacterial biomass (Touloupakis et al. 2016), and it stands to a reason that control agents need to be alkalotolerant or maintain circum-neutral pH conditions when exposed to the target cyanobacteria. Another earlier study found optimal growth of filamentous Schizothrix calcicola at pH levels between 6 and 8. More acidic conditions appeared to limit the pigment expression of this cyanobacterium (West and Louda 2011). Based on the literature referred to earlier in this study on the varied temperatures of the control agent and prey, the culture media referred to in most studies are neutral, with these conditions favourable for most mesophiles. Therefore, although the value of alkalinity is crucial under algal bloom proliferation, there is limited information on the changes brought about by the control agent from this perspective under laboratory test conditions reviewed for the purposes of this report. Do heterotrophic bacteria lower the pH in the cyanobacterial culture medium? Is this also one of the changes that occurs in the growth medium under lab conditions to limit cyanobacterial growth?
Nutrient competition: some thoughts and ideas
Nutrient competition under algal bloom conditions is vital in enabling the domination of the cyanobacterial cells. The gas vesicle of isolates of Microcystis, for example, enables regulation through the water column as well as light shading of other phytoplankton through the formation of surface scum colony aggregates. As Paerl et al. (2016) indicate, the survival and competition strategies of this group of prokaryotes are quite impressive. Therefore, the issue of nutrient competition in biological control using heterotrophs is a crucial consideration. Being mostly heterotrophic, the response to reduced nutrients in the presence of cyanobacterial dominance may be a potential trigger to the bacterial predator response to the cyanobacterial cell. This may mean that at low nutrients, which are under the peak of bloom conditions or when the bloom is dominating the diversity in the aquatic system, the nutrient limitation gives way to microcystins or the cyanobacterial cell being a plausible nutrient source for heterotrophic bacteria. The competition for phosphorous, as indicated in a study by Vadstein et al. (2003) in a microcosm, was the limiting nutrient in the co-existence of algae and heterotrophic bacteria in an aquatic system, whilst the availability of organic carbon was greatly contributed by algal predators (rotifers) than the algae themselves. Moreover, if we consider the research findings of Ji et al. (2017) when assessing nutrient competition between eukaryotic algae and the cyanobacterium Microcystis, we find that at the typical high pH and low CO2 conditions, eukaryotic algae are capable of competing at these conditions. Since the depletion of carbon dioxide by cyanobacteria under bloom conditions has been a hampering impact to the photosynthetic activities of other phytoplankton, this is an interesting finding. However, the nutritional requirements of the heterotrophs differ in the need of organic carbon, noted also in their uptake of NH4+ from the nitrogen fixer Aphanizomenon (Adam et al. 2016), which makes use of N2 fixation under limited nitrogen conditions. Heisler et al. (2008) also describe the ability of some species of cyanobacteria to shift to organic sources of nutrients, which means that in some blooms, this is a direct competition for nutrients between the cyanobacteria and the bacteria. Perhaps under bloom conditions, the limited nutrients to all microorganisms shift the heterotroph towards predation as a means to survive. The (exopolysaccharide) EPS layer of Microcystis and the cells of filamentous cyanobacteria are viable nutrient sources for heterotrophs with a limited food source. Therefore, it may be that the bacterial predation of cyanobacteria is affected by numerous factors including the limitation of nutrients under bloom conditions.
Previously reported bacterial control agents
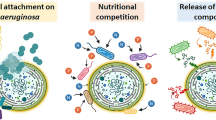
Among the literature, a number of bacterial control agents have been discovered. Figure 1 summarises a brief timeline of bacteria reported to control cyanobacteria and/or algae. Although the list is not exhaustive, it is clear that there are common species reported against the cyanobacteria Microcystis specifically. In correlation with the findings on bacterial communities during a bloom, Pseudomonas, Xanthobacter, Xanthomonas and Alcaligenes as shown in Fig. 1 are isolates under the Proteobacteria phylum, which is abundant in cyanobacterial bloom communities, as previously discussed. Streptomyces (Phankhajon et al. 2016) is within the phylum Actinobacteria, which has also been reported among the abundant phyla. Of interest, however, are the reports on Bacillus, which is within the Firmicutes phylum. This has been less frequently reported to be in abundance as opposed to the other phyla during a cyanobacterial bloom, although Shi et al. (2009) have mentioned this phylum among those within a bloom. Moreover, in the study by Cai et al. (2014), Firmicutes was found in the larger cyanobacterial cell aggregates. The algicidal effect of Aquimarina isolates in a previous study also confirms the presence of Bacteroidetes phyla within the water column where the cyanobacterial bloom occurred (Chen et al. 2011). Although these species exist within the bloom diversities, other studies have implied that the species attached to the cyanobacterial cells have a symbiotic relationship with each other (Shi et al. 2011). In the same study, pathogens Aeromonas and Shewanella sp. were also reported to be positively associated with Microcystis, serving as a potential protection, whilst the other organisms were detected during the vigorous period of the Microcystis bloom.

Research timeline of findings related to algicidal bacteria from the year 2000. Findings from each author are separated by a semicolon, with a summary of the bacterial strain and the cyanobacteria found to be lysed or inhibited. Most of the current research is against Microcystis sp., with fewer publications against other species
Mayali and Azam (2004) reviewed marine algicidal bacteria, which interestingly indicated similar phyla such as those discussed in freshwater studies mentioned here. Repeatedly, earlier studies found that contact time is a significant factor in the effective control or lysis of cyanobacteria by predatory bacteria. Understanding why these control measures are not always effective on their own to curb blooms leads to a conundrum of factors. Among the recent literatures, it appears that the phylum Firmicutes has several effective predatory isolates, especially from the Bacillus genus. This is interesting to note as this phylum is generally found in lower abundance under bloom conditions.
In addition, most of the biological control implementations have been under small-scale laboratory conditions (Choi et al. 2005), with most culture conditions favouring the proliferation of the predator, usually at mesophilic conditions (30 °C), whilst the cyanobacteria are cultured at ambient temperatures. This is often under different nutrient and temperature conditions, which are not always representative of cyanobacterial bloom conditions in nature.
Table 3 summarises a few examples of the control organism used as well as the effective ratios of the predator to prey in causing lysis or inhibition of specific cyanobacteria based on the available literature.
From Table 3, it is evident that most of the research has been conducted at laboratory scale (Fraleigh and Burnham 1988; Shunyu et al. 2006; Chen et al. 2011; Zhang et al. 2011; Tian et al. 2012; Luo et al. 2013; Jia et al. 2014). The effective numbers range from as little as a million bacterial cells up to a 100 million, with effective ratios as low as 1:100 all the way up to 100:1 of the control agent. The laboratory-scale studies show that the Proteobacteria phylum is one of the more commonly reported bacterial control agents, and this supports the findings of Manage et al. (2009), who first reported on external microcystin degraders which were outside this phylum, isolated within the UK. The suppression was based on cell lysis or growth limitation higher than 80% of the starting populations (Manage et al. 2009).
The earlier findings of Proteobacteria as the most common control agents may also be supported by the microbial diversity and abundance during a bloom, as it has been established that, at the peak of the bloom, Proteobacteria and Bacteroidetes are abundant. Algicidal microcystin degraders, however, were also commonly reported from Firmicutes, particularly from Bacillus (Mayali and Azam 2004; Shunyu et al. 2006; Pei et al. 2007; Zhang et al. 2011), which may be logical if Firmicutes increases towards the decline of a bloom.
In the case of liquid cultures, which is the most commonly reported experimental procedure, the predator bacterium is either isolated from within the bloom waters or seldom externally sourced. Screening of primary inhibition is conducted, and based on this, further research is done to analyse algicidal activity and the effects. The predator and prey are grown under separate conditions, with the cyanobacteria grown as axenic or non-axenic strains at ambient temperature, on BG-11, Z8 or similar culture media (Nakamura et al. 2003). The predator is most often grown at higher temperatures, in a different medium. Cells of the predator are then introduced to the cyanobacterial population, and cell death or lysis is measured microscopically or using similar methods as listed above. This shortfall usually is not representative of the natural conditions within the environment as most cultures are axenic cyanobacteria. Table 3 shows the different experiments that have been applied in the control of freshwater cyanobacteria.
Different conditions of biological control implementation using bacteria
Using Fig. 1 as a timeline guide of recent studies conducted in this area, there are a few general conditions that are applied in previous studies to study algicidal or inhibitory effects of bacterial isolates on targeted cyanobacteria. On a laboratory scale, the studies are conducted primarily through
-
Plaque assays on solid agar medium (Rashidan and Bird 2001; Gumbo et al. 2010)
-
Liquid medium growth assays (Nakamura et al. 2003; Choi et al. 2005; Gumbo et al. 2010)
-
Flow cytometric assessments of cell viability and death through live and dead stains, microscopy and cell surface monitoring (Cellamare et al. 2010; Zhou et al. 2012; Gumbo et al. 2014)
The methods listed above are among the most commonly used, with chlorophyll measurements being the primary measurement of cell death. With Microcystis specifically, this has proven to be difficult as it is not an accurate estimation of cell abundance or death. Flow cytometry has been recommended by Gumbo et al. (2014) as well as Cellamare et al. (2010). Most recent is the study conducted by Chapman (2016), where flow cytometry is employed as a predictive measure for blooms, among the known technologies.
The typical experimental set-up involves the culturing of mostly axenic or non-axenic cyanobacterial cultures, which are grown to a certain point, and thereafter, the predatory isolate is added at the optimised working ratios to achieve cell suppression. Most of these effective agents are of the Proteobacteria or Firmicutes genus, often tested against Microcystis aeruginosa. The findings indicate cell lysis, which is often associated to increased toxicity of the test waters. As a result, most of the studies are not suitable for upscaling purposes or further development as is. The other findings indicate predator-to-prey ratios of at least 1:1, for reasonable cyanobacterial suppression and/or lysis.
Research by Su et al. (2016a, b) as referred to in Table 2 indicated overall toxin suppression, which is not commonly reported, although this is against an axenic culture of Microcystis.
Based on the available literature, adaptation of this particular control method (i.e. bacterial biological control of cyanobacteria) is approached with caution, simply due to the numerously reported effects of lysis as well as the unknown further biological impacts. In-depth reviews of the contact time and lysis mechanisms have indicated that no prescribed method may be effective against varied biodiversity (Gumbo et al. 2008). A more recent synopsis of biological control in marine environments equally stresses the point of uncertainty in the possible outcomes of implementation as opposed to terrestrial biological control experiments. Moreover, augmentative biological control appeared to be a more feasible projection compared to the introduction of non-indigenous species (Atalah et al. 2015).
A very interesting 2017 report (Osman et al. 2017) assessed the interactions of axenic cyanobacterial cultures in the presence of known cyanolytic heterotrophs. The analysis of gene upregulation and downregulation indicated stress from both the heterotrophs and the cyanobacteria from a nutrient competition perspective as well as from a growth limitation aspect for Microcystis sp. However, the Aphanizomenon flos-aquae isolate tested under similar conditions indicated cell damage over a 96-h period but there were no clear indications of growth limitation. The study led to the findings that the cell stress response was elicited by the attachment and aggregation of the predatory heterotroph to the cyanobacteria as well as extracellular compounds such as antibiotics, which affect photosynthesis in the cyanobacterial cells. Of further interest among the many findings in this research is the expression of peroxiredoxin, a protection from toxic peroxides and upregulation of cold shock proteins by some of the bacterial isolates. The study also found an energy metabolic pathway shift in the bacterial isolates, indicating nutrient-related stress. These findings corroborate the theories in this review and are a key contribution to understanding the complex cell-to-cell interactions between the cyanobacterial isolates and their heterotrophic bacterial prey. This study was conducted on laboratory axenic strains of cyanobacteria.
Are there stories of success?
Thus far, an assessment of cyanobacterial control methods has found an application of effective microorganisms (EMs), which comprises of a mud ball of microbial consortia to be ineffective in population reduction of lab and wild strains of Microcystis, with effective suppression achieved at very high numbers, possibly due to other factors. It is interesting to note that Lactobacillus, Lactococcus and Saccharomyces comprised the major microbial population in these EM mud balls, which, in conjunction with submerged plants, reduced total nitrogen and phosphates in water. Although mixed data have been collected in the laboratory and field studies in the report, there appear to be various factors influencing the success and failure of the mentioned treatment interventions. This study further questions the proposed effects of these treatments as they appear to be symptomatic treatments as opposed to effective controls of nuisance blooms (Lürling et al. 2016).
More effective results have been observed through the use of metabolites from bacteria or the degradation of microcystins by specific bacteria (Yang et al. 2014), with possibilities for upscaling the treatments on a larger scale (Pei et al. 2007), in a controlled environment. A comprehensive review by Nybom (2013) lists various isolates that have been applied in the bacterial degradation of microcystins and how these microcystins are broken down by the mlr genes, of which mlrA is responsible for the primary breakdown of microcystins. The review continues to indicate the suitability of microbial aggregates in filter systems for microcystin degradation. Interestingly, the author also points towards the use of probiotic bacteria such as Bifidobacterium species for microcystin degradation. The use of other classes of probiotic bacteria in the EM mud balls as previously referred to in the report by Lürling et al. (2016) was, however, not as successful in controlling Microcystis, at a living cell-to-cell interaction approach. It is also interesting to note, however, that microcystins in the environment are degraded within 5–21 days, although most lab-scale studies have periods beyond the minimum 5 days set to observe cyanobacteria or toxin suppression (Welker et al. 2001).
A compilation of some mitigation and prevention strategies at field-scale level has indicated the control of nitrogen and phosphorus ratios in shallow and deep lakes among the possible measures that can be implemented after analysis of the lake system. From a biological intervention perspective, food web management through mussels, which improve light conditions for other macrophytes, and clearing waters through grazing on cyanobacteria are described as a potential intervention. Moreover, the authors state earlier in their report that the food web management has minimal impacts on bloom control (Stroom and Kardinaal 2016). Interestingly, the use of bacteria is not included in these interventions and it may be due to the limited reports on conventional methods for controlling nutrient loading and bloom occurrence in field studies. This really draws attention towards further development of effective predatory bacterial application in bloom mitigation or prevention, which is a key point of this review.
Another publication by Park et al. (2017) indicates how most of the studies in controlling cyanobacterial blooms have been under controlled laboratory conditions. However, the authors propose ultra-sonication, another abiotic measure as a method for bloom control, with compiled reductions of up to 90% algal removal of Aphanizomenon from a field reservoir study by Schneider et al. (2015). Although the study by Park et al. has considerations and a variation of effectiveness in terms of cyanobacterial reduction percentages from different studies, there is a demonstrated effectiveness of this method, with considerations and guidelines for further application in larger-scale water ecosystems.
For the purposes of this review, the focus is mainly on live cell-to-cell interaction between the cyanobacteria and their predatory bacterial isolates, which, apart from the report of Lürling et al. (2016), is rather limited at a larger scale.
Opinion on possible pitfalls
Areas that may require revision is the variation in culture methods and conditions between the predator and prey. The addition of a culture unaccustomed to the bloom conditions may affect the effectiveness of this culture and perhaps the required cell concentrations. This is indicated in the estimation of 6 to 7 days as the required time to present reasonable damage to cyanobacteria when bacterial isolates are concerned.
Moreover, the use of axenic strains in the biological control studies may be cautioned if similar effects cannot be obtained in the natural environment. The fact that these isolates almost never exist as axenic strains in a natural environment outside of the laboratory may indicate that the selected control agent needs to be effective within a mixed diversity. The changes that occur with gold standard isolates such as culture collections of Microcystis isolates may be giving an incorrect representation of the wild strains and possible outcomes with a given control agent. This would require further research of control agent efficiency in different types of blooms, with other co-dominant species and the prospective impact on other aquatic lives.
Control agents need to be examined for their effects on a mixed population of cyanobacteria, which is the common finding in blooms, although certain species may be dominant.
The freshwater studies conducted at present focus majorly on the control of Microcystis-type strains, with other studies focused on marine cyanobacteria (red tides) or diatoms. A handful of these studies look at other cyanobacterial species in detail. At the whole-cell level, it appears that there are few papers indicating suppressed toxin production or toxicity overall through the use of bacterial control agents. This is a crucial aspect to consider if water quality is to be considered.
Another interesting observation is that among the most common control agent phyla, Firmicutes is not often reported as abundant under bloom conditions, indicating that control methods would have to be an augmentative approach, which has not been extensively explored beyond lab-scale studies. Two-step type interventions have also been proposed, where the control agent, specifically, is added in addition to other conventional treatments.
Of importance are the critical assessments of dose-responses/effective ratios and the streamlining of which phytoplankton diversities the proposed agent can be effective against. Cell numbers, chlorophyll and biomass need to be effectively assessed as reliable dosing methods, without blindly applying control agents into waters that have little prior assessment apart from assessing the dominant cyanobacteria.
In closing, the current collection of literature to our knowledge indicates slowed progress in the further application of this research beyond laboratory conditions and further effects beyond the typical toxin release upon lysis, which is counterproductive to the biological control purpose of cyanobacteria. Different temperatures, nutrient conditions and turbulence have repeatedly been indicated as important factors in determining how effective a bacterium may be at effectively reproducing the estimated inhibitory effects on cyanobacteria.
The reasons listed above among the complex interactions within the bloom diversities may be among the reasons there is slow advancement in this area of research, with the remaining major question: why do these lytic agents fail to regulate bloom occurrences in nature?
References
Adam B, Klawonn I, Sveden JB, Bergkvist J, Nahar N, Walve J, Littmann S, Whitehouse MJ, Lavik G, Kuypers MMM, Ploug H (2016) N2-fixation, ammonium release and N-transfer to the microbial and classical food web within a plankton community. ISME J 10:450–459. https://doi.org/10.1038/ismej.2015.126
Atalah J, Hopkins GA, Fletcher LM, Castinel A, Forrest BM (2015) Concepts for biocontrol in marine environments: is there a way forward? Manag Biol Invasion 6:1–12. https://doi.org/10.3391/mbi.2015.6.1.01
Bagatini IL, Eiler A, Bertilsson S, Klaveness D, Tessarolli LP, Vieira AAH (2014) Host-specificity and dynamics in bacterial communities associated with bloom-forming freshwater phytoplankton. PLoS One 9:e85950. https://doi.org/10.1371/journal.pone.0085950
Bouvy M, Pagano M, Troussellier M (2001) Effects of a cyanobacterial bloom (Cylindrospermopsis raciborskii) on bacteria and zooplankton communities in Ingazeira reservoir (Northeast Brazil). Aquat Microb Ecol 25:215–227. https://doi.org/10.3354/ame025215
Cai H, Jiang H, Krumholz LR, Yang Z (2014) Bacterial community composition of size-fractioned aggregates within the phycosphere of cyanobacterial blooms in a eutrophic freshwater lake. PLoS One 9. https://doi.org/10.1371/journal.pone.0102879
Cellamare M, Rolland A, Jacquet S (2010) Flow cytometry sorting of freshwater phytoplankton. J Appl Phycol 22:87–100. https://doi.org/10.1007/s10811-009-9439-4
Chapman IJ (2016) Developing new approaches for monitoring and controlling the toxic cyanobacterium Microcystis through flow-cytometric analysis. Dissertation, Bornemouth University
Chen WM, Sheu FS, Sheu SY (2011) Novel l-amino acid oxidase with algicidal activity against toxic cyanobacterium Microcystis aeruginosa synthesized by a bacterium Aquimarina sp. Enzym Microb Technol 49:372–379. https://doi.org/10.1016/j.enzmictec.2011.06.016
Choi H-J, Kim B-H, Kim J-D, Han M-S (2005) Streptomyces neyagawaensis as a control for the hazardous biomass of Microcystis aeruginosa (cyanobacteria) in eutrophic freshwaters. Biol Control 33:335–343. https://doi.org/10.1016/j.biocontrol.2005.03.007
Demeke A (2016) Cyanobacteria blooms and biological control methods. Int J Fauna Biol Studies 3(1):32–38
Eiler A, Bertilsson S (2004) Composition of freshwater bacterial communities associated with cyanobacterial blooms in four Swedish lakes. Environ Microbiol 6:1228–1243. https://doi.org/10.1111/j.1462-2920.2004.00657.x
Fraleigh PC, Burnham JC (1988) Myxococcal predation on cyanobacterial populations: nutrient effects. Limnol Oceanogr 33:476–483. https://doi.org/10.2307/2837020
Gao E-B, Gui J-F, Zhang Q-Y (2012a) A novel cyanophage with a cyanobacterial nonbleaching protein a gene in the genome. J Virol 86:236–245. https://doi.org/10.1128/JVI.06282-11
Gao Y, Cornwell JC, Stoecker DK, Owens MS (2012b) Effects of cyanobacterial-driven pH increases on sediment nutrient fluxes and coupled nitrification-denitrification in a shallow fresh water estuary. Biogeosciences 9:2697–2710
Geng L, Qin B, Yang Z (2013) Unicellular Microcystis aeruginosa cannot revert back to colonial form after short-term exposure to natural conditions. Biochem Syst Ecol 51:104–108. https://doi.org/10.1016/j.bse.2013.08.027
Graumann PL, Marahiel MA (1999) Cold shock response in Bacillus subtilis. J Mol Microbiol Biotechnol 1:203–209
Gumbo JR, Ross G, Cloete ET (2008) Biological control of Microcystis dominated harmful algal blooms. Afr J Biotechnol 7:4765–4773
Gumbo JR, Ross G, Cloete TE (2010) The isolation and identification of predatory bacteria from a Microcystis algal bloom. Afr J Biotechnol 9:663–671. https://doi.org/10.4314/ajb.v9i5
Gumbo JR, Cloete TE, van Zyl GJJ, Sommerville JEM (2014) The viability assessment of Microcystis aeruginosa cells after co-culturing with Bacillus mycoides B16 using flow cytometry. Phys Chem Earth 72:24–33. https://doi.org/10.1016/j.pce.2014.09.004
Hajdu S, Holinka J, Reichmann S, Hirschl AM, Graninger W, Presterl E (2010) Increased temperature enhances the antimicrobial effects of daptomycin, vancomycin, tigecycline, fosfomycin, and cefamandole on staphylococcal biofilms. Antimicrob Agents Chemother 54:4078–4084. https://doi.org/10.1128/AAC.00275-10
Heisler J, Glibert PM, Burkholder JM, Anderson DM, Cochlan W, Dennison WC, Dortch Q, Gobler CJ, Heil CA, Humphries E, Lewitus A, Magnien R, Marshall HG, Sellner K, Stockwell DA, Stoecker DK, Suddleson M (2008) Eutrophication and harmful algal blooms: a scientific consensus. Harmful Algae 8:3–13. https://doi.org/10.1016/j.hal.2008.08.006
Ji RP, Lu XW, Li XN, Pu YP (2009) Biological degradation of algae and microcystins by microbial enrichment on artificial media. Ecol Eng 35:1584–1588. https://doi.org/10.1016/j.ecoleng.2008.12.031
Ji X, Verspagen JMH, Stomp M, Huisman J (2017) Competition between cyanobacteria and green algae at low versus elevated CO2: who will win, and why? J Exp Bot 68:3815–3828
Jia W, Huang X, Li C (2014) A preliminary study of the algicidal mechanism of bioactive metabolites of Brevibacillus laterosporus on Oscillatoria in prawn ponds. Sci World J 2014:869149–869111. https://doi.org/10.1155/2014/869149
Jung SW, Kim BH, Katano T, Kong DS, Han MS (2008) Pseudomonas fluorescens HYK0210-SK09 offers species-specific biological control of winter algal blooms caused by freshwater diatom Stephanodiscus hantzschii. J Appl Microbiol 105:186–195. https://doi.org/10.1111/j.1365-2672.2008.03733.x
Kim B-H, Sang M, Hwang S-J, Han M-S (2008) In situ bacterial mitigation of the toxic cyanobacterium Microcystis aeruginosa: implications for biological bloom control. Limnol Oceanogr Methods 6:513–522. https://doi.org/10.4319/lom.2008.6.513
Leitão E, Ger KA, Panosso R (2018) Selective grazing by a tropical copepod (Notodiaptomus iheringi) facilitates Microcystis dominance. Front Microbiol 9:301
Lin S, Pan J, Li Z, Liu X, Tan J, Yang H (2014) Characterization of an algicidal bacterium Brevundimonas J4 and chemical defense of Synechococcus sp. BN60 against bacterium J4. Harmful Algae 37:1–7. https://doi.org/10.1016/j.hal.2014.05.002
Louati I, Pascault N, Debroas D, Bernard C, Humbert JF, Leloup J (2015) Structural diversity of bacterial communities associated with bloom-forming freshwater cyanobacteria differs according to the cyanobacterial genus. PLoS One 10:e0140614. https://doi.org/10.1371/journal.pone.0140614
Luo J, Wang Y, Tang S, Liang J, Lin W, Luo L (2013) Isolation and identification of algicidal compound from Streptomyces and algicidal mechanism to Microcystis aeruginosa. PLoS One 8:1–14. https://doi.org/10.1371/journal.pone.0076444
Lürling M, Waajen G, de Senerpont Domis LN (2016) Evaluation of several end-of-pipe measures proposed to control cyanobacteria. Aquat Ecol 50:499–519. https://doi.org/10.1007/s10452-015-9563-y
Man X, Ming L, RC S (2018) Colony formation in the cyanobacterium Microcystis. Biol Rev 0:1399–1420. https://doi.org/10.1111/brv.12401
Manage PM, Kawabata Z, Nakano S (2000) Algicidal effect of the bacterium Alcaligenes denitrificans on Microcystis spp. Aquat Microb Ecol 22:111–117
Manage PM, Edwards C, Singh BK, Lawton LA (2009) Isolation and identification of novel microcystin-degrading bacteria. Appl Environ Microbiol 75:6924–6928. https://doi.org/10.1128/AEM.01928-09
Mayali X, Azam F (2004) Algicidal bacteria in the sea and their impact on algal blooms. J Eukaryot Microbiol 51:139–144. https://doi.org/10.1111/j.1550-7408.2004.tb00538.x
Membré J-M, Leporq B, Vialette M, Mettler E, Perrier L, Thuault D, Zwietering M (2005) Temperature effect on bacterial growth rate: quantitative microbiology approach including cardinal values and variability estimates to perform growth simulations on/in food. Int J Food Microbiol 100:179–186. https://doi.org/10.1016/j.ijfoodmicro.2004.10.015
Mialet B, Majdi N, Tackx M, Azémar F, Buffan-Dubau E (2013) Selective feeding of bdelloid rotifers in river biofilms. PLoS One 8:e75352. https://doi.org/10.1371/journal.pone.0075352
Mohamed ZA, Hashem M, Alamri SA (2014) Growth inhibition of the cyanobacterium Microcystis aeruginosa and degradation of its microcystin toxins by the fungus Trichoderma citrinoviride. Toxicon 86:51–58. https://doi.org/10.1016/j.toxicon.2014.05.008
Movahedi S, Waites W (2002) Cold shock response in sporulating Bacillus subtilis and its effect on spore heat resistance. J Bacteriol 184:5275–5281. https://doi.org/10.1128/JB.184.19.5275-5281.2002
Mu R m, Fan Z q, Pei H y, Yuan X l, Liu S x, Wang X r (2007) Isolation and algae-lysing characteristics of the algicidal bacterium B5. J Environ Sci. https://doi.org/10.1016/S1001-0742(07)60218-6
Nakamura N, Nakano K, Sugiura N, Matsumura M (2003) A novel cyanobacteriolytic bacterium, Bacillus cereus, isolated from a eutrophic lake. J Biosci Bioeng 95:179–184
Ndlela LL, Oberholster PJ, Van Wyk JH, Cheng PH (2016) An overview of cyanobacterial bloom occurrences and research in Africa over the last decade. Harmful Algae 60:11–26. https://doi.org/10.1016/j.hal.2016.10.001
Nedwell DB (1999) Effect of low temperature on microbial growth: lowered affinity for substrates limits growth at low temperature. FEMS Microbiol Ecol 30:101–111. https://doi.org/10.1111/j.1574-6941.1999.tb00639.x
Nybom S (2013) Biodegradation of cyanobacterial toxins. In: Petre M (ed) InTech, Rijeka, p Ch. 7
Oberholster PJ, Botha AM (2010) Use of remote sensing and molecular markers to detect toxic cyanobacterial hyperscum crust: a case study on Lake Hartbeespoort, South Africa. Afr J Biotechnol 9:8791–8799
Oberholster PJ, Botha A, Myburgh JG (2009) Linking climate change and progressive eutrophication to incidents of clustered animal mortalities in different geographical regions of South Africa. J Biotechnol 8:5825–5832
Oberholster PJ, Jappie S, Cheng PH, Botha AM, Matthews MW (2015) First report of an Anabaena bory strain containing microcystin-LR in a freshwater body in Africa. Afr J Aquat Sci 40:21–36. https://doi.org/10.2989/16085914.2014.993583
Osman OA, Beier S, Grabherr M, Bertilsson S (2017) Interactions of freshwater cyanobacteria with bacterial antagonists. Appl Environ Microbiol 83. https://doi.org/10.1128/AEM.02634-16
Paerl HW, Gardner WS, Havens KE, Joyner AR, McCarthy MJ, Newell SE, Qin B, Scott JT (2016) Mitigating cyanobacterial harmful algal blooms in aquatic ecosystems impacted by climate change and anthropogenic nutrients. Harmful Algae 54:213–222. https://doi.org/10.1016/j.hal.2015.09.009
Park J, Church J, Son Y, Kim K-T, Lee WH (2017) Recent advances in ultrasonic treatment: challenges and field applications for controlling harmful algal blooms (HABs). Ultrason Sonochem 38:326–334. https://doi.org/10.1016/j.ultsonch.2017.03.003
Patke, Dey (1998) Proteolytic activity from a thermophilic Streptomyces megasporus strain SDP4. Lett Appl Microbiol 26:171–174. https://doi.org/10.1046/j.1472-765X.1998.00300.x
Pei H y, Hu W r, Mu R m, Li X c (2007) Alga-lysing bioreactor and dominant bacteria strain. J Environ Sci. https://doi.org/10.1016/S1001-0742(07)60091-6
Phankhajon K, Somdee A, Somdee T (2016) Algicidal activity of an actinomycete strain, Streptomyces rameus, against Microcystis aeruginosa. Water Sci Technol 74:1398 LP–1391408
Pietikäinen J, Pettersson M, Bååth E (2005) Comparison of temperature effects on soil respiration and bacterial and fungal growth rates. FEMS Microbiol Ecol 52:49–58
Pomeroy LR, Wiebe WJ, Deibel D, Thompson RJ, Rowe GT, Pakulski JD (1991) Bacterial responses to temperature and substrate concentration during the Newfoundland spring bloom. Mar Ecol Prog Ser 75:143–159
Rampelotto PH (2013) Extremophiles and extreme environments. Life Open Access J 3:482–485. https://doi.org/10.3390/life3030482
Rashidan KK, Bird DF (2001) Role of predatory bacteria in the termination of a cyanobacterial bloom. Microb Ecol 41:97–105. https://doi.org/10.1007/s002480000074
Ratkowsky DA, Olley J, McMeekin TA, Ball A (1982) Relationship between temperature and growth rate of bacterial cultures. J Bacteriol 149:1–5
Ren H, Zhang P, Liu C, Xue Y, Lian B (2010) The potential use of bacterium strain R219 for controlling of the bloom-forming cyanobacteria in freshwater lake. World J Microbiol Biotechnol 26:465–472. https://doi.org/10.1007/s11274-009-0192-2
Schneider OD, Weinrich LA, Brezinski S (2015) Ultrasonic treatment of algae in a New Jersey reservoir. J Am Water Works Assoc 107:E533–E542. https://doi.org/10.5942/jawwa.2015.107.0149
Shao J, Jiang Y, Wang Z, Peng L, Luo S, Gu J, Li R (2014) Interactions between algicidal bacteria and the cyanobacterium Microcystis aeruginosa: lytic characteristics and physiological responses in the cyanobacteria. Int J Environ Sci Technol 11:469–476. https://doi.org/10.1007/s13762-013-0205-4
Shi L, Cai Y, Yang H, Xing P, Li P, Kong L, Kong F (2009) Phylogenetic diversity and specificity of bacteria associated with Microcystis aeruginosa and other cyanobacteria. J Environ Sci 21:1581–1590. https://doi.org/10.1016/S1001-0742(08)62459-6
Shi L, Cai Y, Kong F, Yu Y (2011) Changes in abundance and community structure of bacteria associated with buoyant Microcystis colonies during the decline of cyanobacterial bloom (autumn–winter transition). Ann Limnol Int J Limnol 47:355–362. https://doi.org/10.1051/limn/2011047
Shunyu S, Yongding L, Yinwu S, Genbao L, Dunhai L (2006) Lysis of Aphanizomenon flos-aquae (cyanobacterium) by a bacterium Bacillus cereus. Biol Control 39:345–351. https://doi.org/10.1016/j.biocontrol.2006.06.011
Sigee DC, Glenn R, Andrews MJ, Bellinger EG, Butler RD, Epton HAS, Hendry RD (1999) Biological control of cyanobacteria: principles and possibilities. Hyd 395(396):161–172. https://doi.org/10.1051/animres:19950569
Soini J, Falschlehner C, Mayer C, Böhm D, Weinel S, Panula J, Vasala A, Neubauer P (2005) Transient increase of ATP as a response to temperature up-shift in Escherichia coli. Microb Cell Factories 4:9. https://doi.org/10.1186/1475-2859-4-9
Stroom JM, Kardinaal WEA (2016) How to combat cyanobacterial blooms: strategy toward preventive lake restoration and reactive control measures. Aquat Ecol 50:541–576. https://doi.org/10.1007/s10452-016-9593-0
Su JF, Ma M, Wei L, Ma F, Lu JS, Shao SC (2016a) Algicidal and denitrification characterization of Acinetobacter sp. J25 against Microcystis aeruginosa and microbial community in eutrophic landscape water. Mar Pollut Bull 107:233–239. https://doi.org/10.1016/j.marpolbul.2016.03.066
Su JF, Shao SC, Ma F, Lu JS, Zhang K (2016b) Bacteriological control by Raoultella sp. R11 on growth and toxins production of Microcystis aeruginosa. Chem Eng J 293:139–150. https://doi.org/10.1016/j.cej.2016.02.044
Tian B, Yang J, Zhang KQ (2007) Bacteria used in the biological control of plant-parasitic nematodes: populations, mechanisms of action, and future prospects. FEMS Microbiol Ecol 61:197–213. https://doi.org/10.1111/j.1574-6941.2007.00349.x
Tian C, Liu X, Tan J, Lin S, Li D, Yang H (2012) Isolation, identification and characterization of an algicidal bacterium from Lake Taihu and preliminary studies on its algicidal compounds. J Environ Sci 24:1823–1831. https://doi.org/10.1016/S1001-0742(11)60983-2
Touloupakis E, Cicchi B, Benavides AMS, Torzillo G (2016) Effect of high pH on growth of Synechocystis sp. PCC 6803 cultures and their contamination by golden algae (Poterioochromonas sp.). Appl Microbiol Biotechnol 100:1333–1341. https://doi.org/10.1007/s00253-015-7024-0
Vadstein O, Olsen LM, Busch A, Andersen T, Reinertsen HR (2003) Is phosphorus limitation of planktonic heterotrophic bacteria and accumulation of degradable DOC a normal phenomenon in phosphorus-limited systems? A microcosm study. FEMS Microbiol Ecol 46:307–316
Van Wichelen J, Vanormelingen P, Codd GA, Vyverman W (2016) The common bloom-forming cyanobacterium Microcystis is prone to a wide array of microbial antagonists. Harmful Algae 55:97–111. https://doi.org/10.1016/j.hal.2016.02.009
Verschuere L, Rombaut G, Sorgeloos P, Verstraete W (2000) Probiotic bacteria as biological control agents in aquaculture. Microbiol Mol Biol Rev 64:655–671.
Wang K, Wommack KE, Chen F (2011) Abundance and distribution of Synechococcus spp. and cyanophages in the Chesapeake Bay. Appl Environ Microbiol 77:7459–7468. https://doi.org/10.1128/AEM.00267-11
Welker M, Steinberg C, Jones GJ (2001) Release and persistence of microcystins in natural waters. Cyanotoxins Occur Causes, Consequences 85–103
West M, Louda JW (2011) Effect of external pH on cyanobacterial pigment expression. Florida Sci 74:181–186
White PA, Kalff J, Rasmussen JB, Gasol JM (1991) The effect of temperature and algal biomass on bacterial production and specific growth rate in freshwater and marine habitats. Microb Ecol 21:99–118. https://doi.org/10.1007/BF02539147
Yang Z, Kong F, Shi X, Cao H (2006) Morphological response of Microcystis aeruginosa to grazing by different sorts of zooplankton. Hydrobiologia 563:225–230. https://doi.org/10.1007/s10750-005-0008-9
Yang L, Maeda H, Yoshikawa T, Zhou G (2012) Algicidal effect of bacterial isolates of Pedobacter sp. against cyanobacterium Microcystis aeruginosa. Water Sci Eng 5:375–382. https://doi.org/10.3882/j.issn.1674-2370.2012.04.002
Yang F, Zhou Y, Yin L, Zhu G, Liang G, Pu Y (2014) Microcystin-degrading activity of an indigenous bacterial strain Stenotrophomonas acidaminiphila MC-LTH2 isolated from Lake Taihu. PLoS One 9:1–7. https://doi.org/10.1371/journal.pone.0086216
Zhang X, Hu H-Y, Hong Y, Yang J (2008) Isolation of a Poterioochromonas capable of feeding on Microcystis aeruginosa and degrading microcystin-LR. FEMS Microbiol Lett 288:241–246. https://doi.org/10.1111/j.1574-6968.2008.01355.x
Zhang H, Yu Z, Huang Q, Xiao X, Wang X, Zhang F, Wang X, Liu Y, Hu C (2011) Isolation, identification and characterization of phytoplankton-lytic bacterium CH-22 against Microcystis aeruginosa. Limnologica 41:70–77. https://doi.org/10.1016/j.limno.2010.08.001
Zhang B-H, Ding Z-G, Li H-Q, Mou X-Z, Zhang Y-Q, Yang J-Y, Zhou E-M, Li W-J (2016) Algicidal activity of Streptomyces eurocidicus JXJ-0089 metabolites and their effects on Microcystis physiology. Appl Environ Microbiol 82:5132–5143. https://doi.org/10.1128/AEM.01198-16
Zhou Q, Chen W, Zhang H, Peng L, Liu L, Han Z, Wan N, Li L, Song L (2012) A flow cytometer based protocol for quantitative analysis of bloom-forming cyanobacteria (Microcystis) in lake sediments. J Environ Sci (China) 24:1709–1716. https://doi.org/10.1016/S1001-0742(11)60993-5
Zhou S, Yin H, Tang S, Peng H, Yin D, Yang Y, Liu Z, Dang Z (2016) Physiological responses of Microcystis aeruginosa against the algicidal bacterium Pseudomonas aeruginosa. Ecotoxicol Environ Saf 127:214–221. https://doi.org/10.1016/j.ecoenv.2016.02.001
Zhu L, Wu Y, Song L, Gan N (2014) Ecological dynamics of toxic Microcystis spp. and microcystin-degrading bacteria in Dianchi Lake, China. Appl Environ Microbiol 80:1874–1881. https://doi.org/10.1128/AEM.02972-13
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical statement
This article does not contain any studies with animals performed by any of the authors.
Rights and permissions
About this article

Cite this article
Ndlela, L.L., Oberholster, P.J., Van Wyk, J.H. et al. Bacteria as biological control agents of freshwater cyanobacteria: is it feasible beyond the laboratory?. Appl Microbiol Biotechnol 102, 9911–9923 (2018). https://doi.org/10.1007/s00253-018-9391-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-018-9391-9