
Abstract
Having a sufficient supply of energy, usually in the form of ATP, is essential for all living organisms. In this study, however, we demonstrate that it can be beneficial to reduce ATP availability when the objective is microbial production. By introducing the ATP hydrolyzing F1-ATPase into a Lactococcus lactis strain engineered into producing acetoin, we show that production titer and yield both can be increased. At high F1-ATPase expression level, the acetoin production yield could be increased by 10 %; however, because of the negative effect that the F1-ATPase had on biomass yield and growth, this increase was at the cost of volumetric productivity. By lowering the expression level of the F1-ATPase, both the volumetric productivity and the final yield could be increased by 5 % compared to the reference strain not overexpressing the F1-ATPase, and in batch fermentation, it was possible to convert 176 mM (32 g/L) of glucose into 146.5 mM (12.9 g/L) acetoin with a yield of 83 % of the theoretical maximum. To further demonstrate the potential of the cell factory developed, we complemented it with the lactose plasmid pLP712, which allowed for growth and acetoin production from a dairy waste stream, deproteinized whey. Using this cheap and renewable feedstock, efficient acetoin production with a titer of 157 mM (14 g/L) acetoin was accomplished.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
ATP is one of the most important energy carriers in living organisms, and its availability has an impact on most aspects of cellular metabolism (Zhou et al. 2009). Besides the many reactions that directly depend on ATP, ATP also has various regulatory roles, and a classic example is the mammalian phosphofructokinase 1 (PFK1), which catalyzes the committed step in glycolysis. For this enzyme, ATP is both a substrate and an allosteric inhibitor, and ATP therefore is an important player in terms of regulating the glycolytic activity (Kemp and Foe 1983). Because of the central role of ATP, it is highly important to ensure that the supply of ATP is appropriate when designing cell factories for production of various compounds (Lee and Kim 2015). There are several reported cases where enhancing the ATP supply was beneficial for microbial production, e.g., of succinate (Singh et al. 2011; Liang et al. 2013; Liu et al. 2012), L-valine (Park et al. 2007), or recombinant protein (Kim et al. 2012), but examples of the opposite have also been reported. Patnaik et al. was able to stimulate the glycolytic flux in Escherichia coli under aerobic conditions, by overexpressing phosphoenolpyruvate synthase, which generated an ATP consuming futile cycle (Patnaik et al. 1992). Hädicke et al. used the same futile cycle for improving lactate production in E. coli under anaerobic conditions (Hädicke et al. 2015). Koebmann et al. took a different approach and expressed the F1-ATPase in the cytoplasm of E. coli and achieved the same effect, i.e., stimulation of glycolytic flux (Koebmann et al. 2002b). Causey et al.reported an engineered E. coli strain that efficiently produced acetate, and that had a very high glycolytic flux, which was reported to be partly due to ATP hydrolysis by the F1-ATPase. To achieve cytoplasmic F1-ATPase activity, these researchers deleted the genes encoding the membrane-associated part of the (F1F0)-ATP synthase, which effectively liberated the F1 part of the ATP synthase (Causey et al. 2003).
Introducing an additional ATP consumption (AAC) is thus one possible way to increase the flux in a pathway, which generates a surplus of ATP, but there can be other reasons for introducing ATP-consuming reactions. Lan and Liao introduced an ATP-consuming non-natural pathway into engineered cyanobacteria, to bypass the thermodynamically unfavorable condensation of two molecules of acetyl-CoA to acetoacetyl-CoA, and achieved production of 1-butanol biosynthesis from CO2 (Lan and Liao 2012). Although introducing AAC can result in superior performance, there can be drawbacks associated with this, since an AAC often is accompanied by a reduction in growth rate and less biomass is formed, which ultimately affects the productivity. Having a high productivity is frequently of utmost priority in order for microbial-based processes to be economically feasible (Venayak et al. 2015), and there have been no clear demonstrations of AAC improving titer, yield, and productivity simultaneously.
Lactococcus lactis is a well-characterized lactic acid bacterium, which for several reasons has great potential as a cell factory for production of various chemical compounds. It has a high glycolytic flux, is able to metabolize a broad range of carbohydrates, has a well-characterized metabolic network, and is easy to manipulate genetically (Teusink and Smid 2006). Its long record of safe use in the dairy industry is also an important asset, especially for production of food ingredients (Hugenholtz 2008), where safety has high priority. One compound that potentially could be produced using L. lactis is acetoin, which naturally is made by this and other lactic acid bacteria (LAB) in small amounts. This compound is commonly found in various fermented dairy products, where it contributes to the buttery aroma. Besides its important role in these products, there are, however, a broad range of other uses for acetoin, e.g., within the cosmetic and pharmaceutical industries (Xiao and Lu 2014), and it, among the top 30 most important biomass derived value-added chemicals, because of its many current and potential applications (Werpy and Petersen 2004). Acetoin is derived from pyruvate, from which it can be formed through the action of the two enzymes α-acetolactate synthase (ALS; EC 2.2.1.6) and α-acetolactate decarboxylase (ALD; EC 4.1.1.5; Fig. 1).
Here, we explore the possibility for efficiently producing acetoin using a genetically engineered L. lactis, and we investigate whether an AAC could have a beneficial effect on acetoin yield and productivity. The AAC is achieved via plasmid-based expression of the F1-ATPase. We find that the yield of acetoin can be increased by increasing expression of the F1-ATPase and describe the effect that the F1-ATPase has on biomass formation and growth. By fine-tuning F1-ATPase expression, we show that it is possible to optimize yield, titer, and productivity simultaneously. We furthermore demonstrate that it is possible to convert a waste stream from the dairy industry into acetoin using our developed cell factory.
Materials and methods
Strains and plasmids
The bacterial strains and plasmids used are listed in Table 1. The plasmid pCS1966 (Solem et al. 2008) was used for deleting genes in L. lactis. The recombinant plasmids used for expressing the F1-ATPase were extracted from strains BK1502, BK1503, and BK1536 (Koebmann et al. 2002a).
Growth conditions
For the characterization of different L. lactis strains, including the specific growth rate, glycolytic flux, biomass, and product yield, cells were grown in synthetic amino-acid (SA) medium (Jensen and Hammer 1993) with the following modification: 40 mM MOPS buffer was replaced with 30 mM β-glycerophosphate disodium, supplemented with 0.2 % (w/v) yeast extract (SA + yeast extract (SAY) medium) and 9.5 mM glucose. The cultivation was carried out at 30 °C and 200 rpm. Erythromycin was added to the cultures at a concentration of 5 μg/mL.
Statistical analysis
Student’s t test analysis was performed in order to compare the means of two independent data sets, e.g., acetoin production levels in different recombinant strains overexpressing F1-ATPase with that of the reference strain Apk80. The null hypothesis, assuming no difference in the production levels between different strains, was tested. If the calculated t value exceeded the tabulated value at the level of significance (p = 0.05), we conclude that the samples do differ from one another and that the difference is significant. We also calculated the probability (p value) of getting this t value by chance alone.
Assays involving non-growing cells
L. lactis was grown in 30-mL SAY medium with 10 mM glucose to an OD600 of 1.0. The cultures were cooled and centrifuged at 5000×g for 10 min. Finally, the cells were resuspended in 30 mM β-glycerophosphate disodium buffer to an OD600 of 1.0 and around 9 mM glucose. Samples were withdrawn for glucose measurement.
Measurements of the intracellular ATP and ADP concentrations
Samples were taken from cultures during the exponential growth phase at cell densities of OD600 between 0.4 and 0.5. Then, 0.6 mL of cell culture was mixed with 0.6 mL of 80 °C phenol (equilibrated with 10 mM Tris-1 mM EDTA [pH 8]) supplemented with 0.6-g glass beads (106 μm in diameter; Sigma) and immediately vortexed vigorously for 20 s. After 40 min at room temperature, the samples were vortexed again for 20 s. The two phases were separated by centrifugation at 14,000 rpm for 15 min. The water phase was subsequently treated two times with 1 volume of chloroform to extract residual phenol. The ATP and ADP concentrations were then measured by using a luciferin-luciferase ATP monitoring kit as recommended by the manufacturer. The concentration of ATP was measured first. Subsequently, the ADP content in the same sample was converted to ATP by adding 1 U of pyruvate kinase and 1 mM phosphoenolpyruvate (PEP), and then, the concomitant increase in luminescence was recorded.
DNA techniques
Plasmid DNA from L. lactis was isolated as described by Koebmann et al. (Koebmann et al. 2002a). Cells of L. lactis were made electrocompetent as described by Liu et al. (Liu et al. 2016b). The gene-encoding butanediol dehydrogenase ButBA was deleted based on the method described by Solem et al. (Solem et al. 2008).
Acetoin fermentation
In order to compare the titer, yield, and productivity for acetoin production, the recombinant strains with different F1-ATPase activities were cultivated in SAY medium with around 93 and 180 mM glucose, respectively. The cultivation was performed at 30 °C and 200 rpm. Samples were collected for determining cell density (OD600), glucose, and acetoin concentrations by HPLC as described by Liu et al. (Liu et al. 2016a).
Waste stream conversion
The waste stream (residual whey permeate) from dairy industries contains large amounts of lactose and other nutrients (Liu et al. 2016b). The lactose plasmid pLP712 was transformed into the acetoin-producing strain Abk1503 to get strain AL001, and AL001 was grown in the diluted waste stream containing 99 mM lactose, supplemented with 2 % (w/v) yeast extract. The cultivation was carried out at 30 °C and 200 rpm. Samples were collected for measuring cell density (OD600), lactose, and acetoin concentrations.
Results
Redirecting the metabolic flux from homo-lactic to homo-acetoin
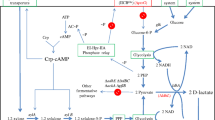
The fermentative and homo-lactic bacterium L. lactis MG1363 normally converts around 90 % of the glucose consumed into lactate (Liu et al. 2016b). We have previously described a derivative of this strain, CS4363, where the genes encoding three lactate dehydrogenases (LDH), phosphotransacetylase (PTA), and alcohol dehydrogenase (ADHE) are deleted (Solem et al. 2013), and this strain is unable to grow anaerobically because it is unable to regenerate NAD+. In the presence of oxygen, however, where the NADH oxidase (NoxE) is active, CS4363 is able to grow with the formation of acetoin as the main fermentation product (Table 2). We have further developed this strain and inactivated the native butanediol dehydrogenases (encoded by butBA) in order to avoid the conversion of acetoin formed into 2,3-butanediol, and this resulted in the strain Ace001 (Fig. 1). This strain was able to produce acetoin as the sole fermentation product with a high yield of 0.78 mol/mol glucose (Table 2). It has previously been reported that diacetyl can be formed from α-acetolactate (Liu et al. 2016a), via a non-enzymatic oxidative decarboxylation, but we were unable to detect diacetyl in our experiments.

Redirection of the metabolic flux for acetoin production. The acetoin production pathway is highlighted in the background, while the dotted lines with a cross indicated the inactivated competing pathways. ALS (ILVB) α-acetolactate synthase, ALD α-acetolactate decarboxylase, NoxE NADH oxidase, AhpR alkyl hydroperoxide reductase, F 1 -ATPase the ATP-hydrolyzing F1-ATPase (overexpressed in the recombinant strains), PDHc pyruvate dehydrogenase complex, PFL pyruvate formate lyase, LDH (LDHX, LDHB) lactate dehydrogenases, PTA phophotransacetylase, ADHE alcohol dehydrogenase, ACK acetate kinase, and ButBA butanediol dehydrogenases
Expressing the F1-ATPase in the acetoin-producing strain Ace001
The acetoin production yield for strain Ace001 was quite high, reaching 78 % of the theoretical maximum, and since all the competing product pathways had been blocked, the remaining option for improving product yield seemed to be to reduce biomass formation. Koebmann et al. (Koebmann et al. 2002a) reported that it is possible to uncouple glycolysis from biomass production by expressing the F1-ATPase, and we decided to test whether this approach could be used to increase the acetoin yield further. Existing plasmid constructs carrying the genes encoding the F1-ATPase expressed from various synthetic promoters were available, and we selected three of these with different strength of promoters (Table 1) for our study. These three plasmids were introduced into Ace001, which resulted in Abk1502, Abk1503, and Abk1536. We also introduced the empty vector, pAK80, into Ace001, and the resulting strain Apk80 served as a reference.
Effect of F1-ATPase activity on the intracellular energy state, growth, and metabolic flux
We measured the intracellular ATP and ADP concentrations in the recombinant strains (Fig. 2). As the F1-ATPase activity increased, the intracellular ATP concentration decreased gradually and the ADP level increased, leading to a significant decrease in the [ATP]/[ADP] ratio from 4.9 to 2.3 (Fig. 2). Subsequently, we characterized the specific growth rate, biomass yield, glycolytic flux, and acetoin production yield (Table 2). Expression of the F1-ATPase had a negative effect on the specific growth rate, which gradually decreased as the expression of F1-ATPase increased. At the highest expression level, the biomass yield was reduced by 29 %, while the acetoin production yield increased by 10 %. At all expression levels, the glycolytic flux remained unaffected, close to 20 mmol/h/g DW. To ascertain that the increases in yield of acetoin observed for the F1-ATPase expressing strains in comparison to the reference strain Apk80 were significant, we applied the Student’s t test. All the p values obtained were below 0.05, which strongly indicated that this indeed was the case (Table 2).

Effect of F1-ATPase on the specific growth rate and the intracellular energy state. a Cell density (OD600) is plotted as a function of time for Apk80 (0 MU), Abk1503 (307 MU), Abk1502 (714 MU), and Abk1536 (1082 MU). The number in the parentheses denotes the strength of the promoter responsible for expressing the F1-ATPase encoding genes in terms of Miller units of β-galactosidase expressed from the reporter gene lacZ. b The ATP, ADP concentrations (μmol/g DW), and [ATP]/[ADP] ratio are indicated. All the experiments were conducted two times, and the error bars indicate the standard deviations
The glycolytic flux increases in non-growing cells with the expression of F1-ATPase
We did not observe an increase in the glycolytic flux for exponentially growing cells, which is consistent with the findings of Koebmann et al. (Koebmann et al. 2002a). In non-growing wild-type L. lactis cells, however, Koebmann et al. reported that the glycolytic flux could be increased when F1-ATPase was expressed. This was also what we found for the acetoin-producing strains examined in this study. As the expression of the F1-ATPase was increased, the glucose consumption rate increased gradually from 4.4 mmol/h/g DW in Apk80 to 7.6 mmol/h/g DW in Abk1536, which was a 73 % improvement (Fig. 3). The acetoin flux was increased following the same trend (data not shown).

Effect of F1-ATPase on the glucose consumption rate in non-growing cells. Cells were resuspended in disodium-β-glycerophosphate buffer (30 mM) containing 9 mM glucose to an OD600 of 1.0, and the glucose concentration was monitored over time. The following strains were included: Apk80 (control, filled squares), Abk1503 (307 MU, triangle), Abk1502 (714 MU, circle), and Abk1536 (1082 MU, filled triangles). The error bars indicate the standard deviation which is based on two replicates
Expression of F1-ATPase increases the titer, yield, and productivity for acetoin production
We observed that the acetoin production yield increased by 10 % at the highest expression level of the F1-ATPase (Table 2). To monitor and compare acetoin formation and glucose consumption more closely, we carried out a time course fermentation using two different initial glucose concentrations—93 and 180 mM. It was found that at initial 93 mM glucose, all the glucose was consumed within 8.7 h by Apk80. However, for the strains which expressed F1-ATPase at different levels, it took longer time and after 11.9, 13.1, and 14.4 h, all glucose had been completely consumed by Abk1503, Abk1502, and Abk1536, respectively. This slower consumption rate correlated with the slower biomass formation (Fig. 4a, b), and the final titer of acetoin increased from 68.8 to 77.0 mM (Fig. 4c), which was a 10 % improvement (p value = 0.02). The productivity for acetoin decreased slightly with the expression of the F1-ATPase due to the extended fermentation period (Fig. 4a–c).

Effect of F1-ATPase on acetoin titer, yield, and productivity. a–c Growth, glucose consumption, and acetoin production, respectively, of the recombinant strains expressing the F1-ATPase, in medium containing an initial glucose concentration of 93 mM. d–f Same as in a–c, just using an initial glucose concentration of 180 mM. The data points represent the averages based on two independent growth experiments, and the error range was within 5 %
At the higher glucose concentration (180 mM glucose), Apk80 and Abk1503 were able to consume all the glucose within 42 h and the glucose consumption profiles were quite similar (Fig. 4e), although the biomass density for Abk1503 (OD600 = 5.1) was much lower than that of Apk80 (OD600 = 6.5; Fig. 4d). Since the final acetoin titer was higher for Abk1503 (146.5 mM = 12.9 g/L, 83.0 % of the theoretical maximum) than for the control strain Apk80 (140.4 mM = 12.3 g/L, 78.5 % of the theoretical maximum; Fig. 4f), this demonstrated that both yield and volumetric productivity could be increased by moderate expression of the F1-ATPase, and both were increased by 5 % (p value = 0.04). We have to mention that most of the acetoin was formed within the first 25 h (130 mM for Abk1503 and 125 mM for Apk80) and that the productivity was calculated based on the time where all glucose had been consumed (42 h). At higher F1-ATPase expression levels, i.e., for strains Abk1502 and Abk1536, not all the glucose could be completely consumed within 42 h, and this resulted in lower titers and productivities (Fig. 4d–f).
Development of a sustainable process for acetoin production
As mentioned above, Abk1503 has a superior titer, yield, and productivity for acetoin. Since the L. lactis strain that we work with has been accustomed to growing in milk, and since large amounts of milk-derived waste is generated by the dairy industry, we decided to test if dairy waste (kindly provided by ARLA Foods) could be a suitable feedstock for acetoin production. Our strain is a plasmid-cured dairy derivative, and it is unable to ferment lactose, and for this reason, we first complemented it with the lactose/protease plasmid pLP712 (Wegmann et al. 2012), which resulted in strain AL001. The outcome was that 99 mM (34 g/L) lactose was completely consumed within 55 h with the formation of 157 mM (14 g/L) acetoin (Fig. 5), which constituted a yield of 80 % of the theoretical maximum.

Acetoin production from lactose by strain AL001. The fermentation medium used consisted of deproteinized whey waste (containing 99 mM lactose) to which 2 % (w/v) yeast extract was added. The data points represent the averages based on two independent growth experiments, and the error bars indicate the standard deviation
Discussion
In the present work, we have demonstrated that it indeed can be beneficial for microbial production to introduce an ATP-consuming futile cycle, and we did this by expressing the native F1-ATPase. The effect of the F1-ATPase was apparent as the biomass yield, specific growth rate, and intracellular energy level all were reduced significantly, and this lead to an increased product yield. This finding that it is possible to obtain a reciprocal correlation between biomass and product yield (the less substrates for biomass, the more for products) emphasizes that the ATP pool is an important metabolic target that should be taken into consideration when designing cell factories for high-yield chemical production, especially when these are based on fastidious organisms which require complex and expensive fermentation media.
One drawback of expressing the F1-ATPase was the reduced specific growth rate and biomass yield, and this could affect the volumetric productivity negatively. This trade-off between product yield and volumetric productivity, however, can be avoided if sugar uptake and the pathway leading to the desired product are optimized in terms of capacity. Koebmann et al. (Koebmann et al. 2002b) observed that the introduction of AAC into E. coli could increase the specific sugar uptake rate as well as the glycolytic flux. Under those circumstances, the cells were able to counteract the loss of ATP by increasing the rate at which it was generated, so that growth could be sustained. However, in the current study, we found that the glycolytic flux in exponentially growing cells could not be affected, irrespective of whether the F1-ATPase was expressed or not, which agrees with the previous work, where the effect of F1-ATPase was studied in a wild-type L. lactis background (Koebmann et al. 2002a). In non-growing cells, we found that the glycolytic flux could be increased by 73 % at the highest expression level of the F1-ATPase, and this stimulatory effect was most likely linked to regeneration of ADP; the availability of which is essential for maintaining a high glycolytic flux. The same effect is likely to be responsible for the small increase in productivity that we observed at moderate F1-ATPase expression levels, as F1-ATPase was beneficial for acetoin production in the post-exponential growth phase, where anabolism slowed down and glycolysis was retarded due to the reduced ATP demand for biomass synthesis. We observed this effect at the high glucose concentration (180 mM), where a large fraction of glucose was left after the exponential phase, which could be consumed faster in the presence of the F1-ATPase, whereas at low glucose concentration (93 mM), glucose was consumed mainly in the exponential growth phase, where the negative effect on growth dominated, leading to less biomass formation and reduced productivity. For the strains expressing the highest level of F1-ATPase, strains Abk1502 and Abk1536, overall productivity was severely affected as the negative effect of the F1-ATPase during the exponential phase could not be compensated for in the post-exponential phase, but the final yield was improved significantly. To avoid this negative effect in the exponential phase, one option could be to use customized inducible/tunable promoters (Mijakovic et al. 2005). These can be switched on to the desired level in the post-exponential phase, to drive the expression of the F1-ATPase, and we are currently investigating this option further. However, the potential applications within the area of whole-cell catalysis (De Carvalho 2011), a valuable approach used for producing a variety of chemicals (Goldberg et al. 2007; Matsumoto and Taguchi 2010), seem attractive too.
When we characterized the acetoin-forming strains, we used simple batch fermentation, where the sugar metabolized was present from the beginning. To achieve a high acetoin titer, we used a high initial glucose concentration of 180 mM. However, at this concentration, a significant growth retardation was observed, which could be due to either the high osmotic pressure (Kasjet 1987) or an effect of a high acetoin concentration or perhaps a combination. To determine whether acetoin could influence growth, we carried out growth experiments where acetoin was added to the culture medium. We found that the specific growth rate of the wild-type strain was indeed affected at relatively low concentrations, and the specific growth rate was reduced by 20 % in the presence of 2 % (w/v) acetoin (data not shown).
With respect to the economic feasibility of biotechnological processes, titer, yield, and productivity are always key parameters that need to be assessed (Stephanopoulos 2007). As described previously, the AAC strategies are able to increase the specific productivity and yield for chemical production (Hädicke et al. 2015); however, there are no reports showing that the volumetric productivity and yield can be improved simultaneously, and this is something we have achieved in this study by fine-tuning the expression of the F1-ATPase. We selected acetoin production for this investigation as acetoin formation generates ATP (2 mol ATP per mol acetoin formed), and it is essential for the strategy to work that the biosynthetic pathway involved generates surplus ATP. As a proof of concept, we demonstrated that dairy waste, in this case deproteinized whey, could serve as a good feedstock for acetoin production, and the results obtained were promising and indicated that it should be possible to establish a both economical and sustainable production process.
In summary, we obtained a higher titer, yield, and productivity for acetoin production in L. lactis expressing AAC reactions. The potential advantage that lies in uncoupling growth from product formation is apparent, and especially when using fastidious organisms for production, this could be relevant as these require expensive growth media. The effect that we observe could in principle be harnessed for production of a variety of interesting compounds derived from glycolytic intermediates.
References
Causey TB, Zhou S, Shanmugam KT, Ingram LO (2003) Engineering the metabolism of Escherichia coli W3110 for the conversion of sugar to redox-neutral and oxidized products: homoacetate production. Proc Natl Acad Sci USA 100:825–832
De Carvalho CC (2011) Enzymatic and whole cell catalysis: finding new strategies for old processes. Biotechnol Adv 29:75–83
Goldberg K, Schroer K, Lütz S, Liese A (2007) Biocatalytic ketone reduction—a powerful tool for the production of chiral alcohols—part II: whole-cell reductions. Appl Microbiol Biotechnol 76:249–255
Hugenholtz J (2008) The lactic acid bacterium as a cell factory for food ingredient production. Int Dairy J 18:466–475
Hädicke O, Bettenbrock K, Klamt S (2015) Enforced ATP futile cycling increases specific productivity and yield of anaerobic lactate production in Escherichia coli. Biotechnol Bioeng 112:2195–2199
Israelsen H, Madsen SM, Vrang A, EB H, Johansen E (1995) Cloning and partial characterization of regulated promoters from Lactococcus lactis Tn917-lacZ integrants with the new promoter probe vector, pAK80. Appl Environ Microbiol 61:2540–2547
Jensen P, Hammer K (1993) Minimal requirements for exponential growth of Lactococcus lactis. Appl Environ Microbiol 59:4363–4366
Kasjet ER (1987) Bioenergetics of lactic acid bacteria: citoplasmic pH and osmotolerance. FEMS Microbiol Rev 46:233–244
Kemp RG, Foe LG (1983) Allosteric regulatory properties of muscle phosphofructokinase. Mol Cell Biochem 57:147–154
Kim H-J, Kwon YD, Lee SY, Kim P (2012) An engineered Escherichia coli having a high intracellular level of ATP and enhanced recombinant protein production. Appl Microbiol Biotechnol 94:1079–1086
Koebmann BJ, Solem C, Pedersen MB, Nilsson D, Jensen PR (2002a) Expression of genes encoding F1-ATPase results in uncoupling of glycolysis from biomass production in Lactococcus lactis. Appl Environ Microbiol 68:4274–4282
Koebmann BJ, Westerhoff HV, Snoep JL, Nilsson D, Jensen PR (2002b) The glycolytic flux in Escherichia coli is controlled by the demand for ATP. Appl Environ Microbiol 184:3909–3916
Lan EI, Liao JC (2012) ATP drives direct photosynthetic production of 1-butanol in cyanobacteria. Proc Natl Acad Sci USA 109:6018–6023
Lee SY, Kim HU (2015) Systems strategies for developing industrial microbial strains. Nat Biotechnol 33:1061–1072
Liang L, Liu R, Li F, Wu M, Chen K, Ma J, Jiang M, Wei P, Ouyang P (2013) Repetitive succinic acid production from lignocellulose hydrolysates by enhancement of ATP supply in metabolically engineered Escherichia coli. Bioresour Technol 143:405–412
Liu J, Chan SHJ, Brock-Nannestad T, Chen J, Lee SY, Solem C, Jensen PR (2016a) Combining metabolic engineering and biocompatible chemistry for high-yield production of homo-diacetyl and homo-(S,S)-2,3-butanediol. Metab Eng 36:57–67
Liu J, Dantoft SH, Würtz A, Jensen PR, Solem C (2016b) A novel cell factory for efficient production of ethanol from dairy waste. Biotechnol Biofuels 9:33
Liu R, Liang L, Chen K, Ma J, Jiang M, Wei P, Ouyang P (2012) Fermentation of xylose to succinate by enhancement of ATP supply in metabolically engineered Escherichia coli. Appl Microbiol Biotechnol 94:959–968
Matsumoto K, Taguchi S (2010) Enzymatic and whole-cell synthesis of lactate-containing polyesters: toward the complete biological production of polylactate. Appl Microbiol Biotechnol 85:921–932
Mijakovic I, Petranovic D, Jensen PR (2005) Tunable promoters in systems biology. Curr Opin Biotechnol 16:329–335
Park JH, Lee KH, Kim TY, Lee SY (2007) Metabolic engineering of Escherichia coli for the production of L-valine based on transcriptome analysis and in silico gene knockout simulation. Proc Natl Acad Sci U S A 104:7797–7802
Patnaik R, Roof WD, Young RF, Liao JC (1992) Stimulation of glucose catabolism in Escherichia coli by a potential futile cycle. J Bacteriol 174:7527–7532
Singh A, Cher Soh K, Hatzimanikatis V, Gill RT (2011) Manipulating redox and ATP balancing for improved production of succinate in E. coli. Metab Eng 13:76–81
Solem C, Defoor E, Jensen PR, Martinussen J (2008) Plasmid pCS1966, a new selection/counterselection tool for lactic acid bacterium strain construction based on the oroP gene, encoding an orotate transporter from Lactococcus lactis. Appl Environ Microbiol 74:4772–4775
Solem C, Dehli T, Jensen PR (2013) Rewiring Lactococcus lactis for ethanol production. Appl Environ Microbiol 79:2512–2518
Stephanopoulos G (2007) Challenges in engineering microbes for biofuels production. Science 315:801–804
Werpy T, Petersen G (2004) Top value added chemicals from biomass volume I—results of screening for potential candidates from sugars and synthesis gas. National renewable energy laboratory, Oak Ridge, TN
Teusink B, Smid EJ (2006) Modelling strategies for the industrial exploitation of lactic acid bacteria. Nat Rev Microbiol 4:46–56
Venayak N, Anesiadis N, Cluett WR, Mahadevan R (2015) Engineering metabolism through dynamic control. Curr Opin Biotechnol 34:142–152
Wegmann U, Overweg K, Jeanson S, Gasson M, Shearman C (2012) Molecular characterization and structural instability of the industrially important composite metabolic plasmid pLP712. Microbiology 158:2936–2945
Xiao Z, Lu JR (2014) Strategies for enhancing fermentative production of acetoin: a review. Biotechnol Adv 32:492–503
Zhou J, Liu L, Shi Z, Du G, Chen J (2009) ATP in current biotechnology: regulation, applications and perspectives. Biotechnol Adv 27:94–101
Acknowledgments
This work was partly supported by a grant from the Innovation fund Denmark (4106-00037B).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
The article does not contain any studies with animals performed by any of the authors.
Rights and permissions
About this article

Cite this article
Liu, J., Kandasamy, V., Würtz, A. et al. Stimulation of acetoin production in metabolically engineered Lactococcus lactis by increasing ATP demand. Appl Microbiol Biotechnol 100, 9509–9517 (2016). https://doi.org/10.1007/s00253-016-7687-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-016-7687-1