
Abstract
A multifunctional Pseudomonas putida X3 strain was successfully engineered by introducing methyl parathion (MP)-degrading gene and enhanced green fluorescent protein (EGFP) gene in P. putida X4 (CCTCC: 209319). In liquid cultures, the engineered X3 strain utilized MP as sole carbon source for growth and degraded 100 mg L−1 of MP within 24 h; however, this strain did not further metabolize p-nitrophenol (PNP), an intermediate metabolite of MP. No discrepancy in minimum inhibitory concentrations (MICs) to cadmium (Cd), copper (Cu), zinc (Zn), and cobalt (Co) was observed between the engineered X3 strain and its host strain. The inoculated X3 strain accelerated MP degradation in different polluted soil microcosms with 100 mg MP kg−1 dry soil and/or 5 mg Cd kg−1 dry soil; MP was completely eliminated within 40 h. However, the presence of Cd in the early stage of remediation slightly delayed MP degradation. The application of X3 strain in Cd-contaminated soil strongly affected the distribution of Cd fractions and immobilized Cd by reducing bioavailable Cd concentrations with lower soluble/exchangeable Cd and organic-bound Cd. The inoculated X3 strain also colonized and proliferated in various contaminated microcosms. Our results suggested that the engineered X3 strain is a potential bioremediation agent showing competitive advantage in complex contaminated environments.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
With advancements in industry, agriculture, and economy, soil contamination with hazardous organic and/or inorganic chemical compounds has gradually exacerbated, resulting in a scarcity of fertile agricultural lands, especially in developing countries (Tsatsakis et al. 2002). Among soil pollutants, organophosphate pesticides, such as methyl parathion (O,O-dimethyl O-4-nitrophenyl phosphorothioate, MP), and heavy metals, such as cadmium (Cd), have been extensively investigated because of potential risks to public health (Zafar et al. 2007; Rani et al. 2009). Therefore, a highly efficient technique should be developed to remediate these contaminated soils (Xu et al. 2012).
Microbes are decomposers necessary to maintain soil fertility. Beyond that, some microorganisms detoxify pesticides or immobilize heavy metals in the environment (Wu et al. 2008). The ubiquity of pesticide degradation and metal immobilization in the environment suggests that bioremediation plays an important role in the removal of organic or inorganic pollutants (Singh et al. 2008; Tiwari and Guha 2013). For instance, MP-degrading microorganisms and enzymes have been used as bioremediation agents; metal-resistant microorganisms have also been applied as immobilizing agents to remediate metal-contaminated environments (Hornel et al. 2002). Furthermore, MP-degrading microbes, including Pseudomonas sp. A3 (Ramanathan and Lalithakumari 1999), Plesiomonas sp. M6 (Cui et al. 2001), and Pseudomonas sp. WBC-3 (Zhang et al. 2008), have been identified. Shen et al. (2010) revealed that Stenotrophomonas sp. SMSP-1 isolated from the sludge of a wastewater treatment system of a pesticide manufacturer can hydrolyze MP to p-nitrophenol (PNP) and dimethyl phosphorothioate but cannot degrade PNP any further. An organophosphorus hydrolase–encoding gene (ophc2) has also been cloned successfully from this bacterium. Other genes for organophosphate degradation (OPD) and methyl parathion degradation (MPD) are found among various soil microbes, including Flavobacterium sp. (Siddavattam et al. 2003), Pseudaminobacter sp., Achromobacter sp., Brucella sp., and Ochrobactrum sp. (Zhang et al. 2006). Bacteria, such as Pseudomonas aeruginosa KUCd1 (Sinha and Mukherjee 2008) and Pseudomonas sp. strain RB (Ayano et al. 2014), which can immobilize Cd, have been isolated and used for microbial remediation. Nevertheless, functional microbes are necessary to eliminate various types of pollutants co-existing in the environment, but a versatile remediation agent is difficult to develop because of competition among functional microbes. With improved knowledge of microbial genomes and proteomes, we can manipulate microbes for biotechnological applications, such as environmental remediation (Wu et al. 2008). Thus, recombinant and multifunctional microorganisms should be constructed and used as potential agents to remediate polluted environments (Timmis et al. 1994).
Sequestration and/or immobilization is an efficient remediation strategy for heavy metals; as such, surface expression of metal-binding proteins in host strain is induced to enhance metal resistance of bacterial cultures for bioremediation. For instance, mouse metallothionein has been introduced to the surface of Ralstonia eutropha CH34 and inoculated in Cd-polluted soil; this strain can immobilize approximately 70 % of Cd accessible to plants, thereby enhancing biomass production of tobacco (Nicotiana bentaminana) (Valls et al. 2000). Degradation is also a primary mechanism to eliminate organic pollutants. Pseudomonas putida and Moraxella sp., host strains commonly found in soil, have been used to recombine organophosphorus hydrolase (OPH) genes; the engineered strain exhibits 70-fold higher activity than Escherichia coli::oph (Shimazu et al. 2001; Lei et al. 2005; Yang et al. 2008), suggesting that the use of a common soil bacterial strain as a remediation agent is a good alternative to alleviate soil contamination. However, studies have mainly focused on the enhancement of bacterial resistance to single type of pollutants. Limited information is available regarding the development of multifunctional engineered strains and their applications in complex polluted environments.
In the current study, an enhanced green fluorescent protein (EGFP)-expressing X3 bacterium that can degrade MP and immobilize Cd was engineered. The remediation efficiency of co-existing MP and Cd in contaminated soils was investigated by monitoring MP residues, Cd distribution, and bacterial population in polluted soil microcosms along with bioremediation process.
Materials and methods
Bacterial strains and culture conditions
The bacterial strains used in this study are listed in Table 1. MP-degrading strain P. putida Hn (CCTCC: AB2015045, preserved in China Center for Type Culture Collection, Wuhan, China) was isolated in our laboratory. This strain can mineralize 100 mg L−1 MP completely within 6 h and can endure the maximum tolerance concentration of 1 g L−1 MP. Heavy metal–resistant strain P. putida X4 (CCTCC: 209319, preserved in China Center for Type Culture Collection, Wuhan, China) was also isolated in our previous study (He et al. 2012; Xu et al. 2012). These bacteria were cultured and maintained in Luria–Bertani medium (LB) at 28 °C. To investigate whether the engineered strain utilizes MP as sole carbon source, we used a mineral salt medium (MSM) supplemented with 100 mg L−1 MP. The MSM contained 3.0 g of K2HPO4, 0.01 g of NaCl, 0.001 g of FeSO4·7H2O, 1.5 g of KH2PO4, 0.1 g of MgSO4, and 1 g of (NH4)2SO4 in 1 L of deionized water at pH 7.0.
Bacterial inoculum was prepared as follows: Test bacterial strains were grown in LB containing 50 μg mL−1 kanamycin or 100 μg mL−1 ampicillin for selection purposes. Bacterial cells were washed with sterile 0.85 % NaCl, centrifuged at 5000×g for 10 min, resuspended in an equal volume of sterile 0.85 % NaCl, and quantified using the dilution plate count technique. In all of the experiments in the current study, the cell density of the bacterial inoculum was adjusted to 107 colony forming units (CFU) mL−1, and bacterial cultures were incubated at 28 °C in a rotary shaker at 180 rpm unless otherwise stated.
Construction of plasmid WB6
The primers used to construct the engineered strain are listed in Table 2. These primers were synthesized by Beijing Sunbiotech Co., Ltd. (Beijing, China). The genomic DNA of the bacteria was obtained according to a previously described method (He et al. 2012). egfp gene was obtained from pEGFP-N3 (GenBank accession no. U55763) with the primers egfp-f and egfp-r; XbaI was incorporated in the primers to facilitate directional cloning of the PCR product into the XbaI site of the pUC19 vector to generate pUC19-EGFP. Kanamycin-resistant gene (Km r) was amplified from the genome of the pET-28a vector and then ligated in pUC19-EGFP to yield pUC19-EGFP-Km via the EcoRI site. PrpsL, a promoter of 30S ribosome subunit, was amplified from P. putida X4 (CCTCC: 209319, Fu et al. 2008). The amplified DNA fragment of PrpsL was cloned in the PstI-SalI sites of the vector to yield pUC19-EGFP-Km-PrpsL. The MP-degrading gene mpd (GenBank accession no. KP677551) was amplified from the genome of P. putida Hn (CCTCC: AB2015045) by using the primers mpdF7BamHI and mpdR13SacI. The PCR product was digested with BamHI and SacI and then ligated in a similarly digested pUC19-EGFP-Km-PrpsL to generate pUC19-EGFP-Km-PrpsL-MPD. The egfp-Km r-PrpsL-mpd fusion gene was released from pUC19-EGFP-Km-PrpsL-MPD with PstI and subcloned in the Broad host vector pBHR1 (Szpirer et al. 2001) to generate plasmid pWB6. The ligation product was transformed into competent E. coli DH5α cells by using standard procedures (Sambrook and Russell 2001). Positive clones containing the pWB6 plasmid were screened on LB agar plates containing 50 μg mL−1 kanamycin and 0.1 g L−1 MP.
Engineered P. putida X3 strain
pWB6 was transformed into P. putida X4 (CCTCC: 209319) host strain by using a tri-parent conjugation method. Transformants were cultivated on an LB agar plate supplemented with 50 μg mL−1 kanamycin, 100 mg L−1 MP, and 2 mM Cd2+ and then incubated at 28 °C for 48 h. Positive clone was selected by monitoring the yellow transparent halo generated by the bacteria. The selected EGFP-tagged multifunctional engineered strain was named P. putida X3.
Discrepancy of growth curve between engineered X3 strain and X4 host strain
The prepared bacterial inoculum (1 %) of X3 and X4 strains was inoculated in a flask containing 100 mL of LB in triplicate, incubated at 28 °C, and 180 rpm. Bacterial cultures were sampled at the following intervals: 0, 1, 2, 3, 4, 5, 7, 8, 9, 10, 12, 16, and 20 days; the optical density of the number of bacterial cells was determined at 600 nm (OD600) by using a UV-visible spectrophotometer.
EGFP expression in P. putida X3
EGFP expressed by P. putida X3 was monitored through fluorescence microscopy (Cassidy et al. 2000) to investigate whether fluorescent intensity produced by EGFP can estimate bacterial population during incubation. X3 strain was inoculated in 100 mL of LB and incubated at 28 °C and 180 rpm. At each sampling time, 1 mL of bacterial culture was collected, centrifuged at 5000×g for 10 min, washed three times with equal volume of sterile 0.85 % NaCl, and detected using a multifunctional fluorescence detector (ModulusTM Fluorometer, Turner BioSystems, CA, USA).
Stability of the plasmid in the engineered X3 strain
To investigate the stability of pWB6 in the engineered P. putida X3 strain, we conducted two experiments: (1) a subculture of X3 strain was continuously cultured on LB media without any selective pressure until 50 generations were produced. The parental engineered X3 strain and its subculture of 50 generations were then cultivated in LB media until OD600 reached 2.0. The fluorescent intensity of these samples was detected using a multifunctional fluorescence detector (ModulusTM Fluorometer, Turner BioSystems, CA, USA). The loss of pWB6 likely resulted in a decrease in fluorescent intensity compared with the host engineered strain; and (2) two hundred colonies of engineered X3 strain were inoculated on an LB media plate without any selective pressure and on LB media with 50 mg L−1 kanamycin. All of the plates were incubated at 28 °C for 24 h. The strain that lost pWB6 did not grow on LB media amended with kanamycin. The rate of the loss of plasmid was determined by counting the number of colonies.
MP degradation with X3 in pure culture
The bacterial inoculum of X3 strain was prepared as mentioned in the preceding section to detect MP-degrading ability of X3 strain. The prepared culture was inoculated in 100 mL of MSM supplemented with 100 mg L−1 MP as sole carbon source. The cultures were then incubated at 28 °C and 180 rpm for 40 h. At desired intervals, culture samples were removed and extracted using equal volume of n-hexane to determine the amount of residual MP and the amount of transformed PNP. The experiments were performed in triplicate. The samples were analyzed through high-performance liquid chromatography (HPLC) in 2487 dual absorbance detector (Waters Co., Milford, MA, USA) (Li et al. 2007). The mobile phase was methanol/water (80:20, v/v) and the flow rate was 1.0 mL min−1. The injection volume of sample was 10 μL at each time. MP and PNP were detected at wavelengths of 275 and 290 nm by using a hypersil ODS2 C18 column (5 μm, 4.6 × 25 mm) of Dalian Elite, respectively. The retention times of MP and PNP were 4.5 and 3 min, respectively. Medium without inoculated bacteria was also sampled and evaluated in the same manner for the control setup.
Heavy metal resistance of engineered X3 strain
The bacterial growth of X3 strain was monitored in LB supplemented with Cd2+ concentrations ranging from 0 to 10 mM with an interval of 0.5 mM to evaluate Cd2+ tolerance of genetic X3 strain. Bacterial inoculum and incubation conditions were the same as those indicated in preceding sections. The host X4 strain was used as positive control. Bacterial growth was routinely determined at OD of 600 nm. Minimum inhibitory concentration (MIC) of X3 strain with other heavy metals, such as Co2+, Cu2+, and Zn2+, was also investigated using the same detection method.
Bioremediation of composite contaminated soils by engineered X3 strain
Surface soil (0–10 cm) was collected from a cornfield free from organophosphate pesticide and heavy metal contamination at Huazhong Agricultural University, Wuhan, Hubei Province, China. The soil was air-dried and sifted through a 2-mm sieve. In this experiment, four types of soil were prepared as follows: (1) natural soil, (2) Cd-contaminated soil, (3) MP-contaminated soil, and (4) Cd-MP-contaminated soil. Cd-contaminated soil was prepared by adding the appropriate Cd2+ solutions to approximately 3 kg of the sieved soil (with final concentration of 5 mg kg−1 dry soil) and maintained at room temperature for at least 7 days to adequately equilibrate Cd in soil. In this period, soil moisture was maintained at 0.2 (Elvang et al. 2001). A portion of Cd pre-treated soil (1500 g in dry weight) and an equal amount of natural soil were supplemented with MP (with a final concentration of 100 mg kg−1 dry soil) under aseptic conditions to prepare Cd-MP-contaminated soil and MP-contaminated soil, respectively. Each type of prepared soil (200 g) was individually transferred to a 300-mL flask. Portions of flasks were inoculated with bacterial X3 strain with a final concentration of 107 CFU g−1 dry soil; the remaining flasks without any bacterial inoculation were used as controls. Three replicates were prepared for each of the eight different treatments. All of the flasks were incubated at 28 °C in the dark.
Three approaches, specifically plate counting, fluorescence detection, and real-time PCR (RTm-PCR) analysis, were applied to monitor the growth of X3 strain in different types of contaminated soils. Culturable inoculated strain was counted according to a previously described method (Xu et al. 2012) with slight modifications. In brief, culturable bacterial cells of X3 were measured by transferring 1 g of soil from bacterium-inoculated soil into a 50-mL sterile centrifuge tube containing 9 mL of sterile 0.85 % NaCl. The tube was shaken at 180 rpm and at 28 °C for 10 min. Tenfold serial dilutions were prepared in 0.85 % NaCl and plated in triplicate on LB agar plates supplemented with 100 mg L−1 MP, 1 mM Cd2+, and 50 mg mL−1 kanamycin. X3 strain could be identified on the basis of a yellow transparent halo generated by PNP, an intermediate metabolite of MP. The total DNA of strains was extracted from the soil sample (Dong et al. 2006). RTm-PCR primers for egfp gene were designed: F 5′-CAAGCAGAAGAACGGCATCAAG-3′ and R 5′-GTGGTTGTCGGGCAGCAG-3′ (Wang et al. 2004). RTm-PCR was performed initially with pre-denaturation at 95 °C for 2 min, followed by 40 cycles of denaturation at 95 °C for 10 s, annealing at 57 °C for 15 s, and extension at 68 °C for 15 s; temperature was increased from 72 to 95 °C at a rate of 1 °C per step. PCR was performed in an ABI Prism© 7000 Sequence Detection System (Applied Biosystems Inc., Foster City, CA, USA). The cycle threshold values obtained from triplicate samples were converted to CFU of P. putida X3 g−1 dry soil by using the standard curves generated from different dilutions of P. putida X3 (Xu et al. 2012). Bacterial population was estimated using the method for detecting fluorescence produced by engineered X3 strain; in brief, 0.3 g of soil was transferred into a 2-mL sterile centrifuge tube with 1 mL of phosphate buffered saline (PBS) containing 1.81 g of Na2HPO4·2H2O, 0.24 g of K2HPO4, 0.08 g of NaCl, and 0.20 g of KCl in 1 L of distilled water. After the mixture was vortexed for 10 min and centrifuged at 4000×g for 1 min, the supernatant containing inoculated bacterial cells was detected using a multifunctional fluorescence spectrometer (ModulusTM Fluorometer, Turner BioSystems, CA, USA). X3 strain cultures were grown to a cell density of 8.6 × 108 CFU mL−1 (log phase) in LB broth at 28 °C and 180 rpm, diluted by tenfold, and transferred to soil (0.3 g dry weight) with a volume of 50 μL to determine the standard curves of fluorescence detection. Soil supernatants were prepared as mentioned above in this section.
Soil samples were extracted with methanol to analyze residual MP (Hassan et al. 2010). Soil samples were filtered using 0.22-μm filter membranes and analyzed through HPLC by using 2487 dual absorbance detector (Waters Co., Milford, MA, USA). A simple sequential extraction procedure (Sposito et al. 1982) was used to fractionate soil Cd into different operationally defined forms with soluble/exchangeable fraction, organic-bound fraction, inorganic-bound fraction, and residual fraction. Reagent blanks and analytical duplicates were prepared whenever appropriate to ensure accuracy of our analysis. Cd was determined using a graphite-furnace atomic adsorption spectrometer (AAS, F-240, VARIAN, Palo Alto, CA, USA).
Sterilization treatment
In order to avoid contamination by genetically modified organism (GMO), appropriate sterilization treatments were carried out in our study. In brief, for the cultures of pure engineered bacteria or corresponding intermediate strains, all the cultures were sterilized in autoclaves at 121 °C for 30 min after each experiment. The soils inoculated with engineered bacteria were sterilized in autoclaves three times with the following procedure after each microcosm experiment: 121 °C for 60 min, 60 °C for 60 min, and 28 °C for 24 h. The sterilized sample was inoculated into LB medium and incubated at 28 °C for 24 h to ensure no survival of engineered bacteria. No-inoculation treatment was set up as the control.
Results
Properties of engineered X3 strain
pWB6 was constructed successfully with a highly efficient expression of egfp and mpd genes controlled by constitutive promoter PrpsL (Fig. 1a). After the recombinant vector was transformed into P. putida X4, an EGFP-tagged multifunctional genetically engineered P. putida X3 strain was obtained. Supplementary Fig. S1 shows the growth of the engineered X3 strain was almost similar to that of X4 strain. This result suggested that plasmid transformation did not affect metabolic activities and growth of the host strain. P. putida X3 with a constitutive EGFP expression was observed under a fluorescence microscope (Fig.1b). Our results also showed that pWB6 was persistent in the subcultures (Supplementary Fig. S2). The degree of fluorescence produced by EGFP of X3 cells corresponds to the growth of X3 strain (Fig. 1c). This property could be used to efficiently monitor the survival or abundance of X3 in the environment.

Constructed plasmid pWB6 (a). Fluorescence of X3 strain was observed using a fluorescence microscope at a magnification of ×640 (b). Fluorescence curve and growth curve of X3 strain (c). Bar represents standard deviations (SD) of the mean (n = 3)
Function of the engineered X3 strain
Figure 2a shows that abiotic factors slightly contributed to MP degradation. X3 strain with mpd gene could utilize MP as a sole carbon source and could efficiently degrade MP to PNP. X3 strain could degrade 100 mg L−1 of MP completely within 24 h. However, the intermediate metabolite PNP could not be further utilized by the engineered strain; thus, this metabolite was retained in the cultures. The engineered P. putida X3 strain and P. putida X4 host strain could tolerate the same concentration of each heavy metal in the liquid media (7 mM Cd2+, 5 mM Co2+, 6 mM Cu2+, and 7.5 mM Zn2+), suggesting that the introduction of pWB6 to the host strain did not elicit any adverse effect on heavy metal resistance.

MP degradation by X3 strain in mineral salt medium supplemented with 100 mg L−1 MP as sole carbon source. MP: methyl parathion; PNP: p-nitrophenol, an intermediate metabolite of MP; X3: engineered strain. Bar represents standard deviations (SD) of the mean (n = 3)
The MP-degrading and heavy metal–tolerant X3 strain could be applied to remediate soils contaminated with various types of pollutants.
Bioremediation of soils contaminated with Cd and/or MP
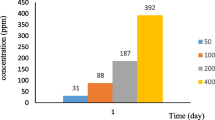
Figure 3 illustrates MP degradation in soil microcosms. MP was degraded more efficiently in MP-contaminated soil microcosms inoculated with X3 strain than in soil microcosms without bacterial inoculation. X3 strain successfully degraded 100 mg kg−1 MP within 40 h in MP-contaminated and MP-Cd-contaminated soils. However, X3 strain degraded higher MP concentrations within 24 h in MP-contaminated soils than in MP-Cd-contaminated soil; this result indicated that the presence of Cd in soil reduced the rate of MP degradation.

MP degradation in various types of contaminated soil microcosms. MP: methyl parathion; Cd: cadmium; X3: engineered strain. Bar represents standard deviations [SD] of the mean (n = 3)
The percentages of various Cd fractions in soils are presented in Fig. 4. In soils without bacterial inoculation (soil-Cd and soil-MP-Cd), soluble/exchangeable Cd decreased from approximately 10 % to approximately 2 % with prolongation of incubation time till 12 days and thereafter increased by more than twofold at the end of the experiment. The percentage of organic-bound fraction also decreased from approximately 15 % to approximately 4 % as incubation time was prolonged, although only slight changes were observed after 12 days. By contrast, inorganic-bound and residual Cd fractions increased for 12 days. No significant changes (p > 0.05) were observed in inorganic-bound and residual Cd fractions in the subsequent incubation time.

Cd distribution in various types of contaminated soil microcosms. Cd-contaminated soil (a), Cd-contaminated soil inoculated with X3 strain (b), MP-Cd-contaminated soil (c), MP-Cd-contaminated soil inoculated with X3 strain (d). MP: methyl parathion; Cd: cadmium; X3: engineered strain
After X3 strain was inoculated, soluble/exchangeable Cd fractions decreased from approximately 10 to <1 % in Cd-contaminated soils during incubation. The amount of organic-bound fractions was lower in soils inoculated with X3 strain in the same incubation period than that in soils without inoculated bacteria. However, inorganic-bound fractions increased from approximately 64 % to approximately 78 % after 60 days of incubation. No significant fluctuations were observed in residual Cd fractions. No evident effect on the distributions of the four Cd forms in soils was observed regardless of the introduction of MP in Cd-contaminated soils.
Plate counting and molecular and fluorescence techniques were employed to investigate the survival of inoculated X3 strain in various types of contaminated soil (Fig. 5). Using plate counting method, we found that the estimated cell density of X3 strain in non-polluted soils decreased for 12 days and then remained at approximately 6.3 in log (CFU dry soil−1) at the end of the experiment (Fig. 5a). By contrast, bacterial cell density of X3 strain increased in other types of polluted soils, increasing trends of inoculated bacterial cell number until 7 days of incubation. The bacterial cell density of X3 strain decreased at 12 days and remained relatively stable thereafter. The population of X3 strain during the entire experiment was slightly higher in Cd-polluted soil than in other soil treatments. After 60 days of incubation, the difference in cell density among the four soil treatments was negligible. Molecular method revealed slight changes in cell density of X3 strain in the four soil treatments, although cell density increased, especially in the first 7 days of incubation in contaminated soils (Fig. 5b). The X3 population in all soil treatments significantly decreased until 20 days. Thereafter, the density of inoculated bacterial cells in various soil treatments exhibited differential changes. The density of inoculated bacterial cells in Cd-polluted soil evidently increased and then remained at approximately 8.0 in log (CFU dry soil−1) at 60 days. Although the cell densities of X3 strain in the three other soil treatments also showed differential increase until 45 d, cell density was approximately 6.5 in log (CFU dry soil−1) at the end of the experiment. Figure 5c shows the changes in cell density of X3 strain in various soil treatments in the experiment. Notably, the density of inoculated bacteria showed a strong fluctuation at the beginning of experiment; thus, changes in bacterial density were initially recorded after 4 days of incubation. The X3 population in non-polluted soil decreased from 8.0 in log (CFU dry soil−1) to 7.0 in log (CFU dry soil−1) during incubation. Changes in the density of inoculated cells in Cd- and/or MP-polluted soils evidently increased initially and then decreased gradually; cell density reached stability at a bacterial count of approximately 8.0 in log (CFU dry soil−1).

Number of inoculated X3 strain counted by the following methods: plate counting (a), RTm-PCR (b), and fluorescence detection (c). Natural soil: soil without any contamination; MP: methyl parathion; Cd: cadmium. Bar represents standard deviations (SD); n = 3)
These results suggested that the inoculation of genetically engineered P. putida X3 strain could be applied effectively in the MP biodegradation and in Cd conversion in contaminated soil microcosms. The inoculated bacterial strain could also colonize and survive in various soil treatments during the experiment.
Discussion
In the current study, MP-degrading and Cd-resistant P. putida X3 strain was successfully constructed using a molecular method; a potential bioremediation agent that could efficiently remediate Cd-MP-contaminated microcosm was also developed. The engineered X3 strain could utilize MP as sole carbon source for growth and hydrolyze MP to the metabolic intermediate PNP; however, PNP could not be further degraded by X3 strain (Fig. 2) possibly because of the lack of PNP-degrading genes. Although X3 strain could not mineralize MP completely, this strain could eliminate 100 mg L−1 MP within 24 h (Fig. 2a); thus, this strain could degrade MP more efficiently than that described in a previous study (Cui et al. 2001; Shen et al. 2010). Furthermore, the MIC of the engineered X3 strain in heavy metals was maintained at levels similar to that of the X4 strain. No evident differences in bacterial growth curves were observed between X4 and X3 strains (Fig. 1c). These results suggested that the engineered X3 strain is suitable for the restoration of MP-Cd-contaminated niches.
Microcosm experiment results showed that the introduction of engineered X3 strain efficiently degraded MP and immobilized Cd in MP- and/or Cd-contaminated soils. However, complete MP removal by X3 strain in soil microcosms required approximately 1.7-fold of incubation time compared with that in liquid cultures (Figs. 2a and 3). The MP degradation rate of inoculated X3 strain was also higher in MP-contaminated soil microcosms free from Cd than in microcosms with Cd in the same incubation period, although X3 completely eliminated MP within 40 h in both soil microcosms. This phenomenon can be explained by the following reasons. (1) MP as a hydrophobic compound exhibits high affinity to soil particles, resulting in lower bioavailability and transmission in soils than in liquid cultures. (2) Although the engineered strain can tolerate a relatively high Cd concentration (7 mM), the growth and metabolic activities of this strain are affected by Cd, causing Cd to be considered as a non-essential element in living organisms.
Cd was distributed as incubation time was prolonged (Fig. 4); as a result, the balance of different Cd fractions changed during the experiment, but these changes were delayed after 12 days. X3 strain accelerated the conversion of soluble/exchangeable Cd and organic-bound Cd to inorganic-bound Cd, thereby decreasing bioavailable Cd for living organisms. However, the organic-bound Cd fraction showed an increasing trend at the end of the experiment. Indeed, Cd was effectively immobilized in soils, and this process depends on several factors, such as efficient immobilizing agents (Xu et al. 2012) and environmental parameters (Jiang et al. 2009). Similar to other bacterial strains (El-Helow et al. 2000), X4 strain (the host strain) could be used as a good immobilizing agent of Cd. This metal is absorbed on cellular surfaces or extracellular polymeric substances secreted by X4 (Wei et al. 2011). Conversely, heavy metals can be desorbed from biosorbents (Wang and Chen 2009). Cd can be released from biosorbents into the environment when changes in environmental factors, such as pH, occur (Ardestani and van Gestel 2013).
A competitive microbial agent is an important factor to achieve microbial remediation (Wu et al. 2008). EGFP was selected as a biomarker to quantify the total number of cells and to monitor dynamic changes in the inoculated engineered strain through RTm-PCR and fluorescence detection approaches, in addition to traditional culture method (Fig. 5). Bacterial counts showed that X3 strain could rapidly colonize various soil treatment microcosms; this result suggested that X3 strain is a potential remediation agent for the restoration of a complex contaminated environment. However, different detection methods resulted in a slight difference of the estimated numbers of X3 cells during incubation probably because of the limitations of the techniques employed in this study. The numbers of X3 cells counted through RTm-PCR and fluorescence techniques were slightly higher than those obtained using plate counting method because molecular approach and fluorescence techniques could detect all of the bacterial cells with egfp gene or those expressing EGFP, including living and dead bacteria. Furthermore, gene horizontal transfer may also contribute to the elevated population of target bacteria. Although our results showed that the plasmid pWB6 was stable in engineered X3 strain in pure culture, the stability and mobility of pWB6 might suffer challenges in soil environment. Of course, co-existence of Cd and MP provided a surviving pressure, which was beneficial for the plasmid in engineered X3 strain so as not to lose or transfer. By contrast, the plate counting method monitored living bacteria only. Estimation bias also resulted from the varying efficiency of the techniques employed to extract inoculated bacteria from soil. Based on the results of bacterial counting, our conclusion is that X3 population likely decreased after this strain was inoculated in the non-polluted soil (Fig. 5); this decrease occurred because X3 strain does not exhibit a competitive advantage over indigenous microbes in non-contaminated environment. In artificially contaminated microcosms, changes in the populations of inoculated bacteria indirectly correspond to the toxicity of MP and Cd. After X3 strain was inoculated, bacterial growth was relatively hampered because of low amounts of nutrients and additional concentrations of contaminants in the new environment. As X3 strain adapted to the new conditions in the contaminated microcosms, this strain proliferated, immediately degraded MP, and/or immobilized Cd in the soil. As the bioavailability of soil pollutants decreased, the pressure of MP and/or Cd to other soil microbes was decreased; the competitive advantage of X3 strain over naturally occurring microbes in the contaminated microcosms also decreased. Therefore, X3 population decreased. These results indicated that the engineered X3 strain can proliferate, efficiently degrade MP, and immobilize Cd in a complex polluted soil; indeed, MP- and Cd-contaminated soils can be remediated and restored. However, novel strains that can further degrade intermediate MP metabolites and immobilize persistent of Cd must be developed to improve microbial remediation.
References
Ardestani M, van Gestel CAM (2013) Using a toxicokinetics approach to explain the effect of soil pH on cadmium bioavailability to Folsomia candida. Environ Pollut 180:122–130
Ayano H, Miyake M, Terasawa K, Kuroda M, Soda S, Sakaguchi T, Ike M (2014) Isolation of a selenite-reducing and cadmium-resistant bacterium Pseudomonas sp. strain RB for microbial synthesis of CdSe nanoparticles. J Biosci Bioeng 117(5):576–581
Cassidy MB, Leung KT, Lee H, Trevors JT (2000) A comparison of enumeration methods for culturable Pseudomonas fluorescens cells marked with green fluorescent protein. J Microbiol Meth 40(2):135–145
Cormack BP, Valdivia RH, Falkow S (1996) FACS-optimized mutants of the green fluorescent protein (GFP). Gene 173(1):33–38
Cui Z, Li S, Fu G (2001) Isolation of methyl parathion-degrading strain M6 and cloning of the methyl parathion hydrolase gene. Appl Environ Microbiol 67(10):4922–4925
Dong D, Yan A, Liu H, Zhang X, Xu Y (2006) Removal of humic substances from soil DNA using aluminium sulfate. J Microbiol Meth 66:217–222
El-Helow ER, Sabry SA, Amer RM (2000) Cadmium biosorption by a cadmium resistant strain of Bacillus thuringiensis: regulation and optimization of cell surface affinity for metal cations. Biometals 13:273–280
Elvang AM, Westerberg K, Jernberg C (2001) Use of green fluorencent protein and luciferase biomarkers to monitor survival and activity of Arthrobacter chlorophenolicus A6 cells during degradation of 4-chlorophenol in soil. Environ Microbiol 3(1):32–42
Fu Y, Chen W, Huang Q (2008) Construction of two lux-tagged Hg2 + -specific biosensors and their luminescence performance. Appl Microbiol Biotechnol 79:363–370
Hassan J, Farahani A, Shamsipur M, Damerchili F (2010) Rapid and simple low density miniaturized homogeneous liquid–liquid extraction and gas chromatography/mass spectrometric determination of pesticide residues in sediment. J Hazard Mater 184(1):869–871
He X, Chen W, Huang Q (2012) Surface display of monkey metallothionein α tandem repeats and EGFP fusion protein on Pseudomonas putida X4 for biosorption and detection of cadmium. Appl Microbiol Biotechnol 95:1605–1613
Hornel I, Sutherland TD, Harcourt RL, Russell RJ, Oakeshott JG (2002) Identification of an opd (Organophosphate Degradation) Gene in an Agrobacterium Isolate. Appl Environ Microbiol 68(7):3371–3376
Jiang C, Sun H, Sun T, Zhang Q, Zhang Y (2009) Immobilization of cadmium in soils by UV-mutated Bacillus subtilis 38 bioaugmentation and NovoGro amendment. J Hazard Mater 167:1170–1177
Lei Y, Mulchandani A, Chen W (2005) Improved degradation of organophosphorus nerve agents and p-nitrophenol by Pseudomonas putida JS444 with surface-expressed organophosphorus hydrolase. Biotechnol Prog 21(3):678–681
Li X, He J, Li S (2007) Isolation of a chlorpyrifos-degrading bacterium, sphingomonas sp. strain Dsp-2, and cloning of the mpd gene. Res Microbiol 158:143–149
Ramanathan MP, Lalithakumari D (1999) Complete mineralization of methylparathion by Pseudomonas sp. A3. Appl Biochem Biotechnol 80(1):1–12
Rani A, Souche YS, Goel R (2009) Comparative assessment of in situ bioremediation potential of cadmium resistant acidophilic Pseudomonas putida 62BN and alkalophilic pseudomonas monteilli 97AN strains on soybean. Int Biodeterior Biodegrad 63:62–66
Sambrook J, Russell D (2001) Molecular Cloning: A Laboratory Manual, 3rd ed
Shen YJ, Lu P, Mei H, Yu HJ, Hong Q, Li SP (2010) Isolation of a methyl parathion-degrading strain Stenotrophomonas sp. SMSP-1 and cloning of the ophc2 gene. Biogegradation 21(5):785–792
Shimazu M, Mulchandani A, Chen W (2001) Simultaneous degradation of organophosphorus pesticides and p-nitrophenol by a genetically engineered Moraxella sp. with surface-expressed organophosphorus hydrolase. Biotechnol Bioeng 76(4):318–324
Siddavattam D, Khajamohiddin S, Manavathi B, Pakala SB, Merrick M (2003) Transposon-like organization of the plasmid-borne organophosphate degradation (opd) gene cluster found in Flavobacterium sp. Appl Environ Microbiol 69(5):2533–2539
Singh S, Kang SH, Mulchandani A, Chen W (2008) Bioremediation: environmental clean-up through pathway engineering. Curr Opin Biotechnol 19(5):437–444
Sinha S, Mukherjee SK (2008) Cadmium-induced siderophore production by a high Cd-resistant bacterial strain relieved Cd toxicity in plants through root colonization. Cur Microbiol 56:55–60
Sposito G, Lund LJ, Chang AC (1982) Trace metal chemistry in arid-zone field soils amended with sewage sludge: I. Fractionation of Ni, Cu, Zn, Cd, and Pb in solid phases. Soil Sci Soci Amer J 46:260–264
Szpirer CY, Faelen M, Couturier M (2001) Mobilization function of the pBHR1 plasmid, a derivative of the broad-host-range plasmid pBBR1. J Bacteriol 183(6):2101–2110
Timmis KN, Steffan RJ, Untermann R (1994) Designing microorganisms for the treatment of toxic wastes. Annu Rev Microbiol 48:525–557
Tiwari MK, Guha S (2013) Kinetics of biotransformation of chlorpyrifos in aqueous and soil slurry environments. Water Res 51:73–85
Tsatsakis AM, Tsakiris IN, Maxaira K, Christakis-Hampsas M, Tzanakakis-Nikitovich DP, Niklis N (2002) Assessment of safe harvesting after methyl parathion application in peaches. Bull Environ Contam Toxicol 68(6):824–830
Valls M, Atrian S, de Lorenzo V, Fernández LA (2000) Engineering a mouse metallothionein on the cell surface of Ralstonia eutropha CH34 for immobilization of heavy metals in soil. Nat Biotechnol 18:661–665
Wang J, Chen C (2009) Biosorbents for heavy metals removal and their future. Biotechnol Adv 27(2):195–226
Wang GJ, Gentry TJ, Grass G, Josephson K, Rensing C, Pepper I (2004) Real-time PCR quantification of a green fluorescent protein-labeled, genetically engineered Pseudomonas putida strain during 2-chlorobenzoate degradation in soil. FEMS Microbiol Lett 233:307–314
Wei X, Fang L, Cai P, Huang Q, Chen H, Liang W, Rong X (2011) Influence of extracellular polymeric substances (EPS) on Cd adsorption by bacteria. Environ Pollut 159:1369–1374
Wu CH, Mulchandani A, Chen W (2008) Versatile microbial surface-display for environmental remediation and biofuels production. Trends Microbiol 16(4):181–188
Xu X, Huang Q, Huang Q, Chen W (2012) Soil microbial augmentation by an EGFP-tagged Pseudomonas putida X4 to reduce phytoavailable cadmium. Int Biodeterior Biodegrad 71:55–60
Yang C, Cai N, Dong M, Jiang H, Li J, Qiao C, Mulchandani A, Chen W (2008) Surface display of MPH on Pseudomonas putida JS 4444 using ice nucleation protein and its application in detoxification of organophosphate. Biotechnol Bioeng 99(1):30–37
Zafar MN, Nadeem R, Hanif MA (2007) Biosorption of nickel from protonated rice bran. J Hazard Mater 143:478–485
Zhang R, Cui Z, Zhang X, Jiang J, Gu JD, Li S (2006) Cloning of the organophosphorus pesticide hydrolase gene clusters of seven degradative bacteria isolated from a methyl parathion contaminated site and evidence of their horizontal gene transfer. Biodegradation 17(5):465–472
Zhang J, Xin Y, Liu H, Wang S, Zhou N (2008) Metabolism-independent chemotaxis of Pseudomonas sp. strain WBC-3 toward aromatic compounds. J Environ Sci 20(10):1238–1242
Acknowledgments
The research was financially supported by the National Natural Science Foundation of China (41230854, 31300102), the National High Technology Research and Development Program of China (“863” Program, 2012AA101402), the Program for Changjiang Scholars and Innovative Research Team in University of China (IRT1247), and the Fundamental Research Funds for the Central Universities (2662015PY016, 2662015PY116).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
This article does not contain any studies with animals performed by any of the authors. All authors have read and approved the final manuscript.
Conflict of interest
The authors declare that they have no competing interests.
Additional information
Rong Zhang and Xingjian Xu contributed equally to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(PDF 341 kb)
Rights and permissions
About this article

Cite this article
Zhang, R., Xu, X., Chen, W. et al. Genetically engineered Pseudomonas putida X3 strain and its potential ability to bioremediate soil microcosms contaminated with methyl parathion and cadmium. Appl Microbiol Biotechnol 100, 1987–1997 (2016). https://doi.org/10.1007/s00253-015-7099-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-015-7099-7