
Abstract
A novel uric-acid-responsive regulatory system was developed in Escherichia coli by adapting the HucR-related regulatory elements from Deinococcus radiodurans into E. coli. The induction performance of this system was compared to the performance of both the pBAD and pET systems. Our novel regulatory system was induced in a dose-dependent manner in the presence of uric acid and exhibited low basal expression in its absence. The system was characterized by a wide dynamic range of induction, being compatible with various E. coli strains and not requiring genomic modifications of the bacterial host. E. coli DH5α and DH10B were the most suitable host strains for optimal performance of this system. In conclusion, we developed a regulatory system with potential for applications in both recombinant protein expression and metabolic optimization.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Regulatory systems that respond to different inducers in Escherichia coli have multiple applications, such as in recombinant protein expression. E. coli is a highly attractive host strain for recombinant protein expression due to its rapid growth and the high-cell-density cultures that can be obtained on inexpensive substrates (Amann et al. 1988; Baneyx 1999; Hannig and Makrides 1998). As a consequence, various inducible expression systems, such as the commonly used pBAD and pET systems, have been developed in E. coli. Similarly, synthetic biology studies and processes of metabolic optimization require tightly controlled genetic regulatory systems (Guido et al. 2006; Keasling 1999). Therefore, development of novel regulatory systems is the subject of intense research.
The E. coli genome is well understood and easy to manipulate; thus, it is a common host in molecular biological studies and the host of choice for the development of several regulatory systems. Although the LacI regulated promoter P lac and its derivatives P tac and P trc are widely used expression system, they exhibit comparatively weak activity and are susceptible to high basal expression in the absence of inducers (Amann et al. 1983; Khlebnikov and Keasling 2002; Stark 1987). The pET expression system is an alternative to P lac -based promoters which is widely used for recombinant protein expression due to a robust response to inducers and high-level expression. However, the pET system exhibits pronounced basal expression, and its use is restricted to strains able to provide T7 RNA polymerase which is required for pET expression (Samuelson 2011; Tabor 2001; Wagner et al. 2008). The l-arabinose-inducible pBAD system is characterized by low basal expression, although it exhibits modest level of activity in the presence of l-arabinose (Guzman et al. 1995). Since l-arabinose is a natural substrate for bacteria, the pBAD system requires host strains carrying genetic modifications to inactivate genes involved in l-arabinose metabolism and harboring constitutive expression of the arabinose transporter. In addition, large-scale application of these regulatory systems is hampered by the high prices of their inducers, isopropyl β-d-1-thiogalactopyranoside (IPTG) or l-arabinose (Peti and Page 2007). Therefore, development of novel regulatory systems with the potential to overcome these limitations is in great demand. In recent years, the evolved tightly regulated TetR promoter PLtetO-1 (Lutz and Bujard 1997), the p-isopropylbenzoate (cumate)-inducible expression system (Choi et al. 2010), fatty acid/acyl-CoA biosensors (Zhang et al. 2012), the cell-growth-induced pLAI system (Nocadello and Swennen 2012), and the Fur-dependent system for toxic protein expression (Guan et al. 2013) were developed either for heterologous protein expression or for dynamic sensor-regulator systems to optimize metabolic pathways.
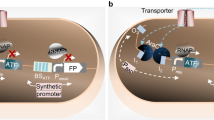
The heterotrophic mesophilic bacterium Deinococcus radiodurans is distinguished for its resistance to harsh environments, such as those characterized by high levels of ionizing or UV radiation, and the presence of bulky chemical adducts or other agents that damage DNA (Battista 1997; Cox and Battista 2005). Hypothetical uricase regulator (HucR) from D. radiodurans belongs to the MarR family of transcriptional regulators. This transcription factor acts as a homo-dimer that represses its self-expression as well as a neighboring uricase locus by binding with high affinity to a single binding site within a promoter/operator region shared by both genes (Fig. 1). Association of HucR to DNA is antagonized by binding of uric acid to HucR, an event that releases the promoter and as a consequence leads to transcription of both genes (Wilkinson and Grove 2004, 2005). Since uric acid is a scavenger of reactive oxygen species, the HucR system might contribute to the intrinsic resistance of the host strain to oxidative stress (Hooper et al. 1998).

Scheme of genetic organizations of the HucR regulatory system in original strain D. radiodurans. The relative orientations of the open reading frames (ORFs) are represented by arrows. HucR binding site (hucO, TCAGTAGGTAGACATCTAAGTATC) is indicated by a dashed box. In the absence of uric acid (or xanthine), HucR protein binds to hucO and prevents the binding of RNA polymerase. When uric acid (or xanthine) antagonizes the HucR-hucO binding, RNA polymerase initiates transcriptional process of uricase and HucR. Adapted from Wilkinson and Grove (2004)
We adapted the HucR system for use in E. coli, in order to develop a novel uric-acid-responsive regulatory system. A series of E. coli strains were used to test the system and for comparisons with the established pBAD and pET expression systems. We have demonstrated that this novel system is tightly regulated, highly induced, and fine-tuned by the cheap and non-toxic inducer uric acid.
Materials and methods
General
The bacterial strains and plasmids used in this study are listed in Table 1. E. coli strains DH5α, DH10B, BL21(DE3), and BW25113 (Datsenko and Wanner 2000) were used for the recombinant protein expression. E. coli strains were cultivated in Luria-Bertani broth (LB) at 37 °C. Antibiotics were used at the following concentrations: ampicillin, 100 μg/ml; kanamycin, 50 μg/ml. Restriction enzymes, DNA polymerase, T4 polynucleotide kinase, and T4 DNA ligase were purchased from Takara Bio, Inc. (Dalian, China). Oligonucleotides were purchased from Life Technologies (Shanghai, China). The sequences of codon-optimized hucR gene (GeneBank Accession number KM596850) sequence, promoter Pcp6 (Karin and Ruhdal 1998) sequence, and a multicloning site sequence were synthesized by Life Technologies (Shanghai, China, Fig. S1 and S2).
Plasmid construction
The backbone of a pSHYa plasmid is based on commercial plasmid DsRed, containing P lac controlled encoding gene of red fluorescent protein rfp. The structural features of the uric-acid-induced promoters and the pSHYa plasmid are summarized in Fig. 2. Sequences of primers used in this study are listed in Table 2.

Features of vectors. a Plasmid map of pSHYa. b Sequences of promoters PhucR, PhucR2, and PhucR3. The −35 and −10 site in the promoter are indicated by underline. The regulatory protein HucR binding sequences are indicated by shaded areas. c The MCS region and 6× histidine tag encoding sequences in plasmid pSHYb
-
1.
pHY. Promoter Pcp6 was amplified using primer Pcp6-ClaI-fwd and Pcp6-rev, resulting in PCR product F1; the hucR gene was amplified with primers hucR-fwd and hucR-rev, resulting in PCR product F2; the ygfU gene (GeneBank Accession number AP009048) (Papakostas and Frillingos 2012) was amplified from E. coli genomic DNA using primers ygfU-fwd and ygfU-NheI-rev, resulting in PCR product F3; the DsRed vector was amplified with primers DsRed-linearize-ClaI and DsRed-linearize-NheI, resulting in PCR product F4. All PCR products were gel-purified, and equimolar aliquots (1.5 nmol each) of fragments F1, F2, and F3 were PCR-assembled without primers. Then, the outer primers hucR-ClaI-fwd and ygfU-NheI-rev were added to the assembly reaction, and the PCR product F5 was amplified. Product F5 was ligated with product F4 after digestion with ClaI and NheI, resulting in plasmid pHY.
-
2.
pHYa. PCR was performed using pHY as template with primers promoter-lacO-deletion-fwd and promoter-lacO-deletion-rev to apply site-directed mutagenesis as described (Zheng et al. 2004), resulting in plasmid pHYa which contained a mutated lacO downstream promoter P lac .
-
3.
pSHYa. PCR was performed using plasmid pHYa as template with the primer pairs PhucR-fwd/PhucR-rev, PhucR2-fwd/PhucR2-rev, and PhucR3-fwd/PhucR3-rev, respectively, resulting in PCR products 1, 2, and 3. Products 1, 2, and 3 were treated with T4 polynucleotide kinase and then self-ligated, resulting in plasmid pSHYa, pSHYa2, and pSHYa3, respectively, which had the 18-bp HucR binding sequence located between −35 and −10 motif, downstream of the −10 motif, and at both of the above locations, respectively.
-
4.
pSHYb. The fragment including a multiple cloning site (MCS) sequence and encoding sequences of 6× histidine tags were amplified with primers (MCS-KpnI-fwd and MCS-BglII-rev) and inserted into the pSHYa plasmid after digestion by KpnI and BglII, resulting in plasmid pSHYb in which the MCS was placed downstream of the PhucR promoter.
-
5.
pSHY-lacZ. The lacZ gene was amplified from E. coli genomic DNA with primers lacZ-KpnI-fwd and lacZ-BglII-rev. The PCR product was ligated into plasmid pSHY after digestion of KpnI and BglII, resulting in plasmid pSHY-lacZ.
-
6.
pET-rfp. Gene rfp was amplified using pSHYa as template with the primers pET28a-rfp-NheI-fwd and pET28a-rfp-XhoI-rev. The PCR product was ligated into a pET28a vector after digestion of NheI and XhoI, resulting in plasmid pET-rfp.
-
7.
pBAD-rfp. Gene rfp was amplified using pSHYa as template with the primers pBAD-rfp-NcoI-fwd and pBAD-rfp-KpnI-rev. The PCR product was ligated into pBAD/Myc-HisC after digestion of NcoI and KpnI, resulting in plasmid pBAD-rfp.
Fluorescence assays
A single colony of the indicated host strain harboring plasmid pSHYa, pBAD-rfp, or pET-rfp was grown overnight in LB medium at 37 °C and then diluted 1:1000 (unless otherwise indicated) in the same medium and grown till optical density (OD600) reached 0.6 (early exponential phase). Inducers of indicated concentrations were added. MOPS buffer (16 mM) was used to maintain the cultural broth at around pH 7.0. The culture was allowed to grow under inducing conditions for 7 h (unless otherwise indicated), and an aliquot of 500 μl was removed and centrifuged, and the cells were resuspended in 500 μl of 100-mM potassium phosphate buffer (pH 6.0). OD600 and RFP fluorescence emission were measured with a SynergyMx Multi-Mode Microplate Reader (BioTek, Vermont, USA) (556-nm excitation filter and 586/20-nm emission filter). The fluorescence data were normalized with respect to OD600. All reported data represent the mean of three independent data points in Figs. 3, 4a, b, and 6 and Table 3.

Comparison of induction performances of promoter PhucR (circle), PhucR2 (square), and PhucR3 (triangle) by measuring RFP fluorescence at the indicated concentrations of the inducer uric acid under the modified standard induction conditions (with 1:100 dilution of culture inoculation and an induction time of 10 h)

The dose-response curves of plasmid pSHYa in DH5α toward the inducer uric acid (a) and xanthine (b), as well as that of plasmid pSHY-lacZ in DH5α toward the inducer uric acid (c). The inducers were added when cell density (OD600) reached 0.6, and a 7-h induction course was allowed. Xanthine concentrations exceeding ~7 μM significantly reduce growth, making the dose response of the pSHYa plasmid to exogenous xanthine difficult to quantify at elevated concentrations
Flow cytometry analyses of the cells harboring plasmid pSHYa induced under various concentrations of uric acid were performed as described previously (Lee and Keasling 2005) with a BD FACS AriaIII flow cytometer (BD Biosciences, San Jose, USA) and excitation at 561 nm, and the fluorescence emission was detected using a 610/20-nm band-pass filter.
SDS-PAGE
The expressions of RFP under the control of different regulatory systems were assessed with sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). Cultures (1.5 ml) prepared in the “Fluorescence assays” section in the presence or absence of inducers were centrifuged, and the cells were resuspended in 0.5 ml of sodium phosphate buffer (pH 7.0, 50 mM). Then, the cells were lysed by ultrasonication, and the crude cell extracts were obtained by centrifugation. The crude cell extracts were applied to SDS-PAGE (15 % separating gel).
Measurement of β-galactosidase activities
β-Galactosidase activity was measured using o-nitrophenyl-β-d-galactopyranoside (ONPG) as substrate (Noh et al. 2009). The hydrolysis reaction was performed by incubating 10-μl crude enzyme extracts prepared by ultrasonication method with 140 μl of 1 mM ONPG in McIlvaine buffer (200 mM Na2HPO4, 100 mM citric acid, pH 6.0) at 37 °C for 10 min, and the reaction was stopped by adding 150 μl of 200 mM Na2CO3. The increase in absorbance at 420 nm due to the release of o-nitrophenol (ONP) was measured. One unit (U) of enzyme activity was defined as the amount of enzyme required to liberate the equivalent of 1 μmol of ONP per minute under the assay conditions. All reported data represent the mean of three independent data points in Fig. 4c.
Results
Development of the uric-acid-responsive regulatory system in E. coli
A regulatory system in E. coli is composed of a regulatory protein and the promoter under its control. The HucR protein was constitutively expressed from the pSHYa plasmid (Fig. 2a). The structure of the consensus regulatory promoter in E. coli consists of an operator and −35 and −10 motifs that facilitate recognition and binding by the RNA polymerase. Although the sequences of the −35 and −10 motifs are highly conserved, the 16–18-bp-long intervening region sequence between them is randomly organized (Harley and Reynolds 1987). The HucR-controlled promoters PhucR, PhucR2, and PhucR3 were constructed by placing an 18-bp sequence, composed of a pseudopalindromic sequence with two 8-bp half-sites separated by 2 bp (TAGGTAGACATCTAAGTA) from the 24-bp D. radiodurans HucR binding site (TCAG TAGGTAGACATCTAAGTA TC) (Wilkinson and Grove 2004) at different locations in the P lac promoter (Fig. 2b). PhucR exhibited the greatest inducing potential, compared with the other two promoters (PhucR2 and PhucR3, Fig. 3). The results suggested that the intensity of induction of gene expression is related to the location of the HucR binding sequence. Therefore, PhucR was chosen for further studies. To avoid induction effect of the P lac promoter by IPTG, the lacO operator was mutated to prevent binding by the LacI repressor. The mutated promoter precluded induction of PhucR by IPTG (data not show). YgfU, the functional uric acid transporter responsible for importing uric acid into cytoplasm in E. coli (Papakostas and Frillingos 2012), was overexpressed from the pSHYa plasmid. Furthermore, an MCS region and encoding sequences of 6× histidine tags were subcloned downstream of PhucR (Fig. 2c), facilitating subcloning of functional genes under the regulation of PhucR.
Induction of the pSHYa regulatory system
RFP expressed under the control of the PhucR promoter was used to characterize the properties of the pSHYa system. The half-maximal and maximal induction occurs at 23 and 50 μM uric acid, respectively (Fig. 4a). RFP expression was induced over 500-fold between 0 and 50 μM uric acid. RFP expression was almost linear when uric acid concentration was increased from 2 to ~30 μM, indicating that the system could be used to quantify environmental uric acid at concentrations that fall within this range. This system could also be effectively induced but with decreased induction fold when the inducer uric acid was supplemented at the stationary phase of growth (data not shown). Since regulatory protein HucR in the original strain was also reported to be induced by xanthine (Wilkinson and Grove 2004, 2005), the induction behavior of this regulatory system in response to xanthine was also tested in E. coli (Fig. 4b). RFP expression was induced about 10-fold in the presence of 7 μM of xanthine. Cell growth was inhibited in the presence of xanthine at a concentration higher than 7 μM, probably due to the NaOH solution used to dissolve xanthine.
To confirm the findings pertaining to RFP expression, we tested the ability of the pSHYa system to induce LacZ expression. The half-maximal induction occurred in the presence of 10.5 μM uric acid, and a maximum of ~200-fold induction in LacZ activity was observed (Fig. 4c). These results indicate that the pSHYa regulatory system can be used to regulate expression of various genes.
Flow cytometry
The capacity to fine-tune gene expression is an important feature of regulatory systems employed in metabolic optimization processes. Flow cytometry was used to measure the extent of induction of the pSHYa system under different concentrations of uric acid. The analysis revealed that cells expressing the pSHYa system showed a homogeneously induced population over the uric acid concentration ranges tested. Furthermore, the average expression level in the cell population could be subtly adjusted by modifying uric acid concentration (Fig. 5), which was also indicated in a response curve of pSHYa (Fig. 4a). The results demonstrated that induction by uric acid was dose dependent and that the system is amenable to fine-tuning of gene expression.

Flow cytometry analysis of RFP expressions from plasmid pSHYa induced by 0 μM (long dashes), 10 μM (gray line), 20 μM (dots), and 40 μM (black line) uric acid
Time course induction analysis
Expression of the pSHYa system over time was measured under a uric acid concentration range. In the absence of uric acid, expression was low, indicating that leaky expression did not occur within the test hours (Fig. 6). At a concentration of uric acid of 5 and 30 μM, expression of RFP increased over time, reaching saturation at about 6 h after induction.

Time courses of RFP expression from plasmid pSHYa induced by 0 μM (square), 5 μM (circle), or 30 μM (triangle) of uric acid in DH5α
Comparison of the regulatory performance of pSHYa with other regulatory systems in E. coli
To compare the regulatory performance of the uric-acid-responsive regulatory system with the pBAD or pET systems in E. coli, the reporter gene rfp was introduced into pBAD/Myc-HisC and pET28a downstream of the PBAD and T7 promoters, respectively. Under the same experimental conditions, the pET system showed the highest induction and basal expression, whereas the pBAD system was the least prone to leaky expression in the absence of an inducer but only showed modest induction. In the absence of inducer, the pSHYa system exhibited much lower levels of basal expression than that of the pET system, but somewhat higher than that of the pBAD system, indicating that gene expression was fairly tightly regulated in the pSHYa system. The induction fold illustrates the dynamic range of the response to an inducer and was calculated as the ratio of fluorescence in cells exhibiting maximal induction levels over fluorescence in cells in the absence of an inducer. The pET system was induced to 40-fold with a high level of basal expression, whereas the pSHYa (in its most suitable host strain DH5α) and pBAD systems were induced over 500 and 200-fold, respectively (Table 3). Maximal RFP expression from the pSHYa system was as high as ~50 % of that from the pET system, while its basal expression in the absence of an inducer was only ~3 % of that from the pET system (Table 3). The induction capabilities of regulatory systems were confirmed by SDS-PAGE (Fig. 7).

Comparative analysis of RFP expressions. RFP expressions from pSHYa, pET, and pBAD regulatory systems in the indicated host strains in the presence and absence of inducers were assessed with SDS-PAGE. Plus sign, in the presence of the inducer (1 mM IPTG, 1 mM arabinose, or 50 μM uric acid); negative sign, in the absence of inducer; M, molecular weight markers
Performance of the pSHYa system in various host strains
Since the pSHYa system could function in multiple strains without requiring any genetic modification in the host, the performance of the pSHYa system was studied in the E. coli strains BW25113, DH10B, DH5α, and BL21(DE3) (Table 3). The attributes of the pSHYa system varied among different strains. The system showed low basal expression in the absence of an inducer in all the strains under investigation. The highest maximal induction and the highest induction fold were obtained in DH5α and DH10B strains, indicating that these are the strains where the pSHYa system achieves optimal performance. It is noteworthy that in the host strain BL21(DE3), the induction fold of the pSHYa system was comparable to the pET system, but displayed much less basal expression.
Discussion
Regulatory systems play essential roles in both production of recombinant proteins and regulation of metabolic pathways. Although various inducible expression systems are available, more robust regulatory systems are still in demand (Gupta et al. 1999; Studier and Moffatt 1986). The novel uric-acid-responsive regulatory system developed in this study exhibited high maximal expression level and relatively low basal expression. This system exhibits an extremely wide dynamic range of transcriptional regulation, indicating a strong gene expression regulatory capacity.
Since expression of recombinant proteins or variations in metabolic pathways might severely hamper growth of the host strain, low expression levels in the absence of an inducer are favored. The pBAD expression system has been used extensively to control and probe cellular processes due to its relatively low level of basal expression in the absence of arabinose, which might be due to the weak activity of the PBAD promoter (Guzman et al. 1995; Lee and Keasling 2005). However, this expression system is characterized by low maximal induction, and our results confirmed this observation. Although the pSHYa system displayed a much stronger induction capability than the pBAD system in the presence of an inducer, it showed a somewhat higher level of basal expression in the absence of an inducer. Since protein expression levels are dependent on promoter strength and translational efficiency (Hannig and Makrides 1998), the basal expression of the pSHYa system may be reduced by mutating the promoter region (Blazeck and Alper 2013) or by using a weaker ribosome binding site (Barrick et al. 1994). These interventions would probably reduce the fully induced expression level but maintain the wide dynamic range of induction simultaneously.
The advantages of the novel pSHYa system entailed its applicability for use in various E. coli strains, without requiring genomic modifications, under the regulation of uric acid, a cheap, non-metabolized, and non-toxic inducer. Although DH5α and DH10B are strains used for molecular manipulation, mostly, they are not applied for protein overexpression. Due to its performances in the DH5α and DH10B strains, the pSHYa system will facilitate studies of molecular cloning and gene overexpression. Besides, pSHYa could also be applied to host strains like BL21(DE3) and BW25113 with fairly high induction capabilities, opening new opportunities for researchers.
Subtler approaches for metabolic optimization have been hampered by the lack of appropriate systems for fine-tuning gene expression (Keasling 1999; Lee et al. 2011). Our findings indicated that the pSHYa system has the capacity to fine-tune gene expression, providing additional choices of regulatory systems with different effector specificity for synthetic biology studies.
Transcription of YgfU, a specific transporter of uric acid with minimal transporting capacity for xanthine, is dependent on σ54-RNA polymerase in E. coli, suggesting that YgfU is only expressed under nutrient-starvation condition (Maeda et al. 2000). In this study, YgfU was expressed from the constitutive promoter Pcp6 in the pSHYa system for efficiently transporting uric acid from the external environment into the cytoplasm. As a consequence, a minimal concentration of inducer was needed to induce the regulatory system to the greatest extent. In addition, uric acid can be removed from the culture by exposure to uricase under circumstances where inducer contamination of recombinant protein products is undesirable, which is a drawback for inducer IPTG (Figge et al. 1988).
Some promoters in microbes display an all-or-none or autocatalytic induction pattern, partially due to the fact that the expression of the transporter specific for the inducer is under the control of the same promoter (Aaron and Milton 1957; Carrier and Keasling 1999; Siegele and Hu 1997). In this study, YgfU was constitutively expressed to maintain a constant intracellular uric acid concentration and to prevent an all-or-none response of the pSHYa system.
Uric acid is an efficient scavenger of reactive oxygen species, including hydroxyl radicals, superoxide anion, and singlet oxygen, and is considered a critical antioxidant in mammals (Papakostas and Frillingos 2012; Wilkinson and Grove 2005). In the D. radiodurans strain, the uric-acid-responsive HucR regulatory system maintains an optimum level of uric acid, thereby reducing reactive oxygen species concentration and conferring extreme resistance to oxidative stress. Therefore, the regulatory system developed in this study will allow design of microbial sensors in E. coli with sensitive and dynamic responses to extracellular and intracellular levels of uric acid.
In conclusion, we have developed a uric-acid-responsive regulatory system in E. coli that tightly regulates gene expression over a wide dynamic range. This novel regulatory system has potential applications in both recombinant protein expression and metabolic optimization processes.
References
Aaron N, Milton W (1957) Enzyme induction as an all-or-none phenomenon. Proc Natl Acad Sci U S A 43:553–566
Amann E, Brosius J, Ptashne M (1983) Vectors bearing a hybrid trp-lac promoter useful for regulated expression of cloned genes in Escherichia coli. Gene 25:167–178
Amann E, Ochs B, Abel K-J (1988) Tightly regulated tac promoter vectors useful for the expression of unfused and fused proteins in Escherichia coli. Gene 69:301–315
Baneyx F (1999) Recombinant protein expression in Escherichia coli. Curr Opin Biotechnol 10:411–421
Barrick D, Villanueba K, Childs J, Kalil R, Schneider TD, Lawrence CE, Gold L, Stormo GD (1994) Quantitative analysis of ribosome binding sites in E. coli. Nucleic Acids Res 22:1287–1295
Battista JR (1997) Against all odds: the survival strategies of Deinococcus radiodurans. Annu Rev Microbiol 51:203–224
Blazeck J, Alper HS (2013) Promoter engineering: recent advances in controlling transcription at the most fundamental level. Biotechnol J 8:46–58
Carrier TA, Keasling JD (1999) Investigating autocatalytic gene expression systems through mechanistic modeling. J Theor Biol 201:25–36
Choi YJ, Morel L, Le François T, Bourque D, Bourget L, Groleau D, Massie B, Míguez CB (2010) Novel, versatile, and tightly regulated expression system for Escherichia coli strains. Appl Environ Microbiol 76:5058–5066
Cox MM, Battista JR (2005) Deinococcus radiodurans - the consummate survivor. Nat Rev Microbiol 3:882–892
Datsenko KA, Wanner BL (2000) One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci U S A 97:6640–6645
Figge J, Wright C, Collins CJ, Roberts TM, Livingston DM (1988) Stringent regulation of stably integrated chloramphenicol acetyl transferase genes by E. coli lac repressor in monkey cells. Cell 52:713–722
Guan L, Liu Q, Li C, Zhang Y (2013) Development of a Fur-dependent and tightly regulated expression system in Escherichia coli for toxic protein synthesis. BMC Biotechnol 13:25–33
Guido NJ, Wang X, Adalsteinsson D, McMillen D, Hasty J, Cantor CR, Elston TC, Collins JJ (2006) A bottom-up approach to gene regulation. Nature 439:856–860
Gupta JC, Jaisani M, Pandey G, Mukherjee KJ (1999) Enhancing recombinant protein yields in Escherichia coli using the T7 system under the control of heat inducible λPL promoter. J Biotechnol 68:125–134
Guzman LM, Belin D, Carson MJ, Beckwith J (1995) Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J Bacteriol 177:4121–4130
Hannig G, Makrides SC (1998) Strategies for optimizing heterologous protein expression in Escherichia coli. Trends Biotechnol 16:54–60
Harley C, Reynolds RP (1987) Analysis of E. coli promoter sequences. Nucleic Acids Res 5:2343–2361
Hooper DC, Spitsin S, Kean RB, Champion JM, Dickson GM, Chaudhry I, Koprowski H (1998) Uric acid, a natural scavenger of peroxynitrite, in experimental allergic encephalomyelitis and multiple sclerosis. Proc Natl Acad Sci U S A 95:675–680
Karin H, Ruhdal JP (1998) Artificial promoter libraries for selected organisms and promoters derived from such libraries. Patent No. WO1998007846
Keasling JD (1999) Gene-expression tools for the metabolic engineering of bacteria. Trends Biotechnol 17:452–460
Khlebnikov A, Keasling JD (2002) Effect of lacY expression on homogeneity of induction from the Ptac and Ptrc promoters by natural and synthetic inducers. Biotechnol Prog 18:672–674
Lee SK, Keasling JD (2005) A propionate-inducible expression system for enteric bacteria. Appl Environ Microbiol 71:6856–6862
Lee T, Krupa R, Zhang F, Hajimorad M, Holtz W, Prasad N, Lee S, Keasling J (2011) BglBrick vectors and datasheets: a synthetic biology platform for gene expression. J Biol Eng 5:1–14
Lutz R, Bujard H (1997) Independent and tight regulation of transcriptional units in Escherichia coli via the LacR/O, the TetR/O and AraC/I1-I2 regulatory elements. Nucleic Acids Res 25:1203–1210
Maeda H, Fujita N, Ishihama A (2000) Competition among seven Escherichia coli σ subunits: relative binding affinities to the core RNA polymerase. Nucleic Acids Res 28:3497–3503
Nocadello S, Swennen E (2012) The new pLAI (lux regulon based auto-inducible) expression system for recombinant protein production in Escherichia coli. Microb Cell Fact 11:3–12
Noh K-H, Son J-W, Kim H-J, Oh D-K (2009) Ginsenoside compound K production from ginseng root extract by a thermostable β-glycosidase from Sulfolobus solfataricus. Biosci Biotechnol Biochem 73:316–321
Papakostas K, Frillingos S (2012) Substrate selectivity of YgfU, a uric acid transporter from Escherichia coli. J Biol Chem 287:15684–15695
Peti W, Page R (2007) Strategies to maximize heterologous protein expression in Escherichia coli with minimal cost. Protein Expr Purif 51:1–10
Samuelson J (2011) Recent developments in difficult protein expression: a guide to E. coli strains, promoters, and relevant host mutations. In: Evans JTC, Xu M-Q (eds) Heterologous gene expression in E. coli. Humana Press, New York, pp 195–209
Siegele DA, Hu JC (1997) Gene expression from plasmids containing the araBAD promoter at subsaturating inducer concentrations represents mixed populations. Proc Natl Acad Sci U S A 94:8168–8172
Stark MJR (1987) Multicopy expression vectors carrying the lac represser gene for regulated high-level expression of genes in Escherichia coli. Gene 51:255–267
Studier FW, Moffatt BA (1986) Use of bacteriophage T7 RNA polymerase to direct selective high-level expression of cloned genes. J Mol Biol 189:113–130
Tabor S (2001) Expression using the T7 RNA polymerase/promoter system. In: Ausubel FA, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K (eds) Current protocols in molecular biology. Wiley, New York, pp 16.2.1–16.2.11
Wagner S, Klepsch MM, Schlegel S, Appel A, Draheim R, Tarry M, Högbom M, van Wijk KJ, Slotboom DJ, Persson JO, de Gier J-W (2008) Tuning Escherichia coli for membrane protein overexpression. Proc Natl Acad Sci U S A 105:14371–14376
Wilkinson SP, Grove A (2004) HucR, a novel uric acid-responsive member of the MarR family of transcriptional regulators from Deinococcus radiodurans. J Biol Chem 279:51442–51450
Wilkinson SP, Grove A (2005) Negative cooperativity of uric acid binding to the transcriptional regulator HucR from Deinococcus radiodurans. J Mol Biol 350:617–630
Zhang F, Carothers JM, Keasling JD (2012) Design of a dynamic sensor-regulator system for production of chemicals and fuels derived from fatty acids. Nat Biotechnol 30:354–359
Zheng L, Baumann U, Reymond J-L (2004) An efficient one-step site-directed and site-saturation mutagenesis protocol. Nucleic Acids Res 32:115–122
Acknowledgments
The authors would like to thank Mr. Ye Mao for his technical assistance in this research. This work was supported by the Ministry of Science and Technology of China Grant 2013CB734003, the National Natural Science Foundation of China Grant 21172095, and the Key Research Program of the Chinese Academy of Sciences Grant KSZD-EW-Z-015-2.
Author information
Authors and Affiliations
Corresponding author
Additional information
Chaoning Liang and Dandan Xiong contributed equally to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(PDF 88 kb)
Rights and permissions
About this article

Cite this article
Liang, C., Xiong, D., Zhang, Y. et al. Development of a novel uric-acid-responsive regulatory system in Escherichia coli . Appl Microbiol Biotechnol 99, 2267–2275 (2015). https://doi.org/10.1007/s00253-014-6290-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-014-6290-6