
Abstract
Predation can drive variation in prey phenotypes by selecting for both constitutive and inducible defensive traits, which can act in concert to protect prey from predators. To understand the adaptive evolution of prey traits it is important to determine which traits are true targets of natural selection. Selection gradient analysis is a useful tool for doing this. We combined field surveys of the extensive intraspecific variation in defensive shell traits of the intertidal whelk Nucella lamellosa with laboratory experiments to determine which traits were associated with risk of crab predation, and which traits were under selection by the whelks’ major crab predator, Cancer productus. Field surveys showed strong associations between risk of crab predation and shell shape, shell thickness, and apertural area, but not apertural teeth. Laboratory choice experiments in which crabs were offered a variety of whelk shell morphs revealed that thinner–shelled whelks with wider apertures were consumed significantly more often than thicker–shelled whelks with narrower apertures, confirming previous findings, and resulting in positive selection differentials for these traits. However, multiple logistic regression revealed that only shell thickness, and none of the other traits measured, had a significant effect on whelk survival, indicating only weak directional selection for this trait. The mechanism for crabs selecting for shell thickness in whelks is likely through increased handling time and decreased profitability. Given that shell thickness is a phenotypically plastic trait induced by C. productus, future work should examine the degree to which selection by crabs acts on both constitutive levels and inducibility of shell thickness in N. lamellosa.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Examining how intraspecific variation in prey traits translates into performance and fitness can provide important insight into the adaptive evolution of those traits, as well as the dynamics of predator–prey interactions (Yamamichi et al. 2011). Most prey adaptively evolve multiple defensive traits in response to selective pressure from predators (Bourdeau 2009; Langerhans 2007; Dalziel and Boulding 2005; Sepúlveda et al. 2012) and these traits, which are often correlated, can act separately or in combination to defend prey against these predators (Schlichting an Wund 2014; Stoks et al. 2016). To understand how multiple, correlated defensive traits evolve in prey, it is necessary to determine on which traits, or trait combinations, natural selection is acting.
One approach for doing this is selection gradient analysis (e.g., DeWitt and Langerhans 2003), where a focal prey organism is exposed to the predator, or predators, thought to be exerting strong selection, and determining which traits are most associated with prey survival. Selection gradient analysis can be a powerful tool for identifying true targets of selection among multiple, correlated defensive traits in prey (Lande and Arnold 1983; DeWitt and Langerhans 2003). Furthermore, selection gradient analysis generates a standardized metric that quantifies both the magnitude and direction of selection acting on prey traits that is comparable across studies (Lande and Arnold 1983). Therefore, selection gradient analyses, when carried out on natural prey populations in the field, or in laboratory mesocosm experiments in combination with field surveys of intraspecific trait variation, can generate strong inferences about selection acting on natural populations (e.g., DeWitt and Langerhans 2003).
Crab–marine gastropod predator–prey pairs are excellent systems with which to study selection on multiple, correlated defensive traits, due to the long co–evolutionary history and escalation of offensive and defensive traits in these two groups (Vermeij 1977). In several species of rocky shore gastropods, individuals from populations on wave–protected shores, where shell–breaking crabs are abundant and pose a high risk of predation, develop more well–defended shells than individuals from populations on wave–exposed sites, where crabs tend to be scarce (Crothers 1983). These well–defended shells tend to be comprised of multiple, correlated and presumably well–integrated traits, including: thickening of the apertural lip of the shell, which strengthens it against apertural lip–peeling attacks by crabs (Palmer 1985a); narrowing of the shell aperture, which makes the soft tissue less vulnerable to shell entry attacks like winkling and peeling (DeWitt et al. 2000); reduction of the shell spire, which prevents spire clipping attacks (DeWitt et al. 2000; Edgell and Neufeld 2008), and a wider shell shape that hinders grip on the shell–body whorl and spreads crushing forces more evenly over the surface of the shell (DeWitt et al. 2000; Bourdeau 2012). These traits can be an integrated combination of genetically–controlled development and a phenotypically plastic response induced by chemical cues associated with risk of crab predation (Palmer 1985b; Appleton and Palmer 1988; Bourdeau 2012). Whelks from protected shores with high crab abundance will have both constitutive traits and exhibit greater plastic responses to crab presence than those from wave-exposed shores (Bourdeau 2012). Crabs induce a plastic change in shell shape either directly (Appleton and Palmer 1988; Palmer 1990; Rochette et al. 2007), or indirectly through starvation resulting from crab-induced foraging suppression (Bourdeau 2010a).
Rocky shore whelks in the genus Nucella in particular, are well-known for intraspecific variation in defensive shell morphology (Crothers 1982; Currey and Hughes 1982; Kitching 1976; Vermeij and Currey 1980). Studies that have experimentally examined the adaptive value of morphological variation within species in the genus have consistently demonstrated that thicker-shelled individuals are more resistant to attack by shell-breaking crabs than thinner shelled ones (Hughes and Elner 1979; Kitching and Lockwood 1974; Kitching et al. 1966; Wellington and Kuris 1983). Shells of N. lamellosa are among the most variable of the genus, with variation in shell shape, sculpture, and apertural lip thickness the most dramatic and often correlated with habitat and changing drastically over short distances (Crothers 1984; Appleton and Palmer 1988; Bourdeau 2012). Increased shell thickness in N. lamellosa is associated with habitats in which crabs, particularly the red rock crab, Cancer productus, are abundant (Crothers 1983) and N. lamellosa from shores with high risk of predation by C. productus exhibit greater plasticity for shell thickness in the presence of C. productus chemical cues (Bourdeau 2009, 2010a,b, 2012, 2013). The adaptive value of thicker shells in N. lamellosa has been examined in laboratory experiments with C. productus, where thinner-shelled individuals of N. lamellosa were more likely to be eaten than thicker-shelled individuals (Palmer 1985a). Thus, thicker shells would appear to be one target of selection in N. lamellosa by C. productus.
Shell thickness in N. lamellosa, however, is only one of several correlated shell traits that may be selected for by C. productus. For example, individuals of N. lamellosa develop larger apertural teeth in the chemical presence of C. productus than in its absence (Appleton and Palmer 1988). The apertural teeth are hypothesized to act as an inducible defense by reducing the effective apertural area of the shell, thus preventing the crab from inserting its chelae into the aperture for a peeling attack (Appleton and Palmer 1988). N. lamellosa from shores with higher risk of predation by C. productus also develop wider, short-spired, narrow-apertured shells, while those from low-risk habitats develop elongate, tall-spired, wider aperture shells, suggesting among-habitat divergence in shell shape driven by selection from C. productus (Bourdeau 2012). Wider, short-spired shells with narrower apertures are hypothesized to be more resistant to crushing, spire-clipping, and peeling attacks than elongate, high-spired shells with wider apertures (Appleton and Palmer 1988; Lowell et al. 1994; Bourdeau 2009). Both shell shape and thickness also contribute to overall shell strength in N. lamellosa, suggesting that shell shape and inducible shell thickening have evolved jointly to form an effective defense in habitats where crab predation risk is high (Bourdeau 2012).
The C. productus–N. lamellosa predator–prey system has been used for studies of co-evolution (Vermeij and Currey 1980), prey selection (Palmer 1985a), and adaptive phenotypic plasticity (Appleton and Palmer 1988; Edgell and Neufeld 2008; Bourdeau 2009, 2012). However, no previous studies have attempted to quantify multivariate selection for this system. Although there are no studies showing simple Mendelian inheritance of shell traits in Nucella lamellosa per se., the shell sculpture of laboratory-raised individuals of a closely related congener (N. emarginata–now N. ostrina) have been shown to be at least partly heritable in a Mendelian fashion (Palmer 1985b). Furthermore, documented interpopulation differences in phenotypic plasticity and constitutive development of defensive shell traits in N. lamellosa (Appleton and Palmer 1988; Bourdeau 2012) imply that natural selection has favored the retention of these traits in habitats where predatory crabs are more common (e.g., wave-sheltered habitats). Although these differences alone do not constitute evidence of past natural selection, (as genetic drift (Masel 2011) could have also played a role in the differentiation between the two morphs) given sufficient genetic variation for constitutive shell development and shell phenotypic plasticity, N. lamellosa provides a tractable system for studying how natural selection acts on these traits. To examine how multiple, correlated defensive shell traits evolved in N. lamellosa, we combined: (1) field surveys to examine associations between shell trait variation and intensity of crab predation in natural populations of N. lamellosa; (2) experiments to examine how intraspecific variation in N. lamellosa shell traits translates into performance and fitness; and (3) selection gradient analyses to identify true targets of selection imposed on N. lamellosa shell traits by C. productus.
Methods
Associations between shell shape variation and crab predation risk in the field
We sampled whelks from eight populations over multiple years from two distinct geographic regions: northern California (2015, 2016) and San Juan Island, Washington (2005, 2016; Fig. 1). At each sampling location, we haphazardly collected 38–64 whelks from the mid-to-low-intertidal zone, along 20–75 m transects placed parallel to the shore (electronic supplementary Table 1S). The length of the transects and the number of whelks collected at each site depended on available sampling area and whelk density, respectively; with more whelks collected from sites with higher densities so as to collect a large enough sample size, but not over-collect from a site. Whelks were returned to either Friday Harbor Marine Laboratories on San Juan Island, Washington (FHL) or the Telonicher Marine Lab (TML) in Trinidad, California and frozen for later morphometric analysis. Before processing, whelks were individually cataloged with a site code and assigned a number. Numbers were written with industrial strength marker in two places on the main body whorl of the shell and covered with a thin application of clear coat nail polish for protection.

Map of collection locations in northern California a: Point St. George (PSG; 41°46′27.41"N, 124°14′56.69"W), Baker Beach (BB; 41° 2′57.37"N, 124° 7′40.54"W), Luffenhotlz Beach (LHB; 41° 2′25.58"N, 124° 7′15.81"W)), and Cape Mendocino (CM; 40°23′55.50"N, 124°22′53.72"W) and San Juan Island, Washington b: County Park (COP; 48°34′39.49"N, 123°10′25.94"W), Land Bank Trust (LBT; 48°31′38.90"N, 123° 9′20.40"W), False Bay (FB; 48°28′49.84"N, 123° 3′55.37"W), and Cattle Point (CAP; 48°27′15.07"N, 122°58′30.39"W)
Risk of crab predation was assessed for each sample population by quantifying the number of crab scars, indicative of unsuccessful crab attacks, on individual whelks (Stafford et al. 2015). To account for the effect of snail growth rate on scar occurrence, only scars on the first body whorl of the shell were counted (Molinaro et al. 2014). Although differences in repair scar frequency can result from either variation in the number of attacks, or variation in attack success rate (Vermeij 1982, 1978), repair scars record actual attacks and can be used to produce quantitative estimates of crab predation among localities. Furthermore, whereas this method did not allow us to calculate the number of crabs at each site, there is strong relationship between the presence of repair scars in intertidal gastropods and rock crab abundance (Stafford et al. 2015), indicating that repair frequency is a valid tool for comparing crab predation intensity between gastropod populations (Stafford et al. 2015; Tyler et al. 2015).
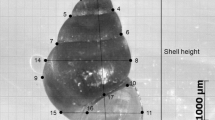
For each whelk, six linear measurements were taken with digital calipers (± 0.01 mm): shell length and width, two apertural lip thicknesses (one at the shoulder of the apertural lip and one at the base, which were averaged; Fig. 2a), and aperture opening length and width (Fig. 2b). Variation in shell thickness is difficult to accurately quantify. We chose to measure lip thickness directly with precision digital calipers (± 0.01 mm). Lip thickness was chosen for two reasons: 1) it is highly correlated with shell mass (r = 0.85, df = 399, P < 0.0001), which is a good estimate of resistance to crushing attacks, 2) it also provides a more localized estimate of shell thickness that is relevant for peeling attacks. Furthermore, measuring apertural lip thickness for this relatively large and thick-shelled species is generally more repeatable than measuring smaller, thinner shelled species. In the case of N. lamellosa, we could measure shell thickness with a repeatability of within 0.1 mm. However, because there is variation in thickness along the apertural lip of individual snails, we chose to use the average of two values (at the shoulder and at the base) as our response variable. After measuring shell dimensions, whelks were lightly boiled to ease the removal of soft tissue from the shell. Once the soft tissue was removed, the shell was dried in a drying oven for 36 h at 30℃ and weighed on a digital balance (± 0.01 g).

Diagram showing linear measurements of the shells of Nucella lamellosa: a shell length and width, and two apertural lip thicknesses (one at the shoulder of the apertural lip and one at the base), b aperture opening length and width, and c positions of the 11 landmarks used in the geometric morphometric analysis
For shell shape analysis each shell was photographed with a digital camera mounted on a lighted copy stand. We partially embedded shells in modeling clay on the copy stand with the aperture facing up and parallel to the camera lens. Digital photos were uploaded into tpsDig software (tpsRelw version 1.44; Rohlf 2006), which we used to place 11 landmarks on the digital image of each shell (Fig. 2c). We analyzed the shell shape of field-collected whelks with landmark-based geometric morphometrics (Bookstein 1989; Rohlf and Marcus 1993), a method that retains the geometry of shape and facilitates visualization and statistical analysis of shape differences among groups. These landmarks were put through a generalized Procrustes analysis, allowing us to remove the effects of specimen size, position, and orientation in the digital images. The subsequent aligned landmark configurations were used to generate shape variables as partial warp scores (tpsRelw version 1.44; Rohlf 2006). Warp scores are principal components of the distribution of shapes and summarize the variation in local shape deformations among the specimens. To summarize the variation in shape among the specimens, we performed a relative warp analysis on the partial warp scores to quantify the variation in shell shape between individuals (RW; Rohlf and Marcus 1993). We also quantified the proportion of whelks with apertural teeth in each population.
To quantify shell strength, we determined the maximum force (N) required to catastrophically break the main body whorl of individual shells. While this method does not allow us to determine the whelks’ resistance to apertural peeling or spire clipping, it did allow us to test how well the whelks could combat the crushing attacks employed by C. productus (Zipser and Vermeiji 1978). This was accomplished by placing the shell aperture down in the center of a stationary platen on a single column series 3340 Instron Universal Testing System and crushing it between the stationary platen and a compression platen at a rate of 1 mm s−1 (Blundon and Vermeij 1983).
Laboratory experiment 1: Influence of apertural teeth on crab predation
For experiment 1, we collected 140 whelks from three of the four Northern California field sites that had differing crab predation pressure (Table 1; PSG, LHB, and BB), selecting whelks of similar size (26–44 mm) and shape while only keeping individuals with toothed apertural lips. For logistical reasons, we were only able to do this experiment in northern California. Whelks were returned to TML and labeled with a site code and individual number on the main body whorl with nail polish pens, followed by a layer of acrylic clear coat to protect the label. We used box traps to collect C. productus from Trinidad Pier in Trinidad, California (41.031830, − 124.084929) and selected crabs of similar size (carapace width: 97–127 mm), with both claws intact, and an equal ratio of males to females.
To manipulate the presence of apertural teeth we used a rotary sanding tool to sand down the apertural teeth of half the experimental whelks (without altering the thickness of the apertural lip basal to the teeth). Apertural teeth were left intact in the other half of the experimental whelks, but to control for any effects of sanding, we sanded (for the same duration and intensity as the experimental whelks) a randomly chosen location on their main body whorls (this procedural control had no effect on the whelks’ susceptibility to shell breakage when compared to completely intact shells). Seven C. productus were each given 20 whelks; 10 with apertural teeth removed, and 10 controls. Enclosures had no shelter for whelks, so crabs could always reach them during the experiment. Crabs were starved for 6d before the experiment. We stopped the experiment after 36 h after the crabs had consumed only 30% of the whelks to ensure that we were capturing their preference and not allowing the crabs to simply eat the whelks they were enclosed with, at which point we removed the whelks from the enclosures, and categorized individual snails as “consumed” or “alive”.
Laboratory experiment 2: Crab selection on whelk shell traits
To quantify the effect of crab selection on whelk shell morphology, we did a laboratory experiment in which we offered crabs a range of N. lamellosa shell morphologies. We hand-collected 325 whelks from the four Northern California field sites that had differing crab predation pressure (Table 1; PSG, LHB, CM, and BB), selecting individuals that were similar in size (26–45 mm) and maximizing the range of shell shape variation. Whelks were returned to TML and labeled and measured the same as in Experiment 1 (see above). Additionally, whelks were photographed and measured for linear and geometric morphometrics as in the field survey above.
We collected crabs (C. productus) in baited box traps off of Del Norte Dock in Eureka, California (40.472665, -124.112047) and Trinidad Pier, again selecting crabs with both claws intact and an equal ratio of males to females. Crab carapace widths (mean = 114.9 mm, SD = ± 11.3 mm) were measured with calipers.
In the first experimental trial, we placed 5 C. productus in flow-through tanks (304.8 × 190.5 × 203.2 mm L × W × H) covered in black plastic to block out the unpredictable light schedule in the lab at night, since rock crabs are primarily nocturnal (Rebach 1985). As with Laboratory experiment 1, there was no shelter provided for the whelks, so they were always accessible to the crabs during the experiment. Each crab was starved for 6d prior to the experiment, then offered 22 live whelks that represented a range of shell shapes as determined by relative warp scores. C. productus feeding was stopped after 60 h (~ 30% consumption). We did a second trial of this same experiment with another 5 C. productus. We terminated this trial after 48 h (~ 30% consumption). At the end of each trial we removed live whelks from holding tanks and categorized individuals as “consumed” or “alive”. Surviving snails were collected and saved.
Statistical and selection gradient analyses
For the field survey data, we used Akaike Information Criterion (AIC; Burnham and Anderson 2004) to select among several candidate linear mixed models describing the relationship between shell strength (the response variable) and shell characteristics. Shell morphological data collected from whelks from Luffenholtz Beach (Trinidad, CA) appeared to be outliers, and so were removed from the analysis. We think this is justified due to the highly dynamic physical and biological nature of this site. Luffenholtz Beach is a west-facing sandy beach flanked to the north and the south by two rocky reefs. The mouth of a freshwater creek bisects the beach, but the relative size and position of the creek and its mouth can change drastically from season-to-season and year-to-year. We have noted that seasonal and annual changes in the creek flow affect the amount of freshwater input and sedimentation to each of the rocky reefs on the beach, leading to large changes in crab abundance through time. We, therefore, do not expect whelk shell traits from 2 years of sampling to be highly correlated with the highly variable crab abundances at this site. We tested for effects of region on crab predation pressure using an ANOVA with site as a random factor (Upton 1992). For laboratory experiment 1, we used a Fisher’s Exact test (Upton 1992) to test for the independence of apertural tooth presence and whelk survival.
For laboratory experiment 2, we estimated the intensity of directional selection using a logistic regression model (Janzen and Stern 1998), which is more suitable for estimating selection gradients when outcomes are dichotomous (e.g., absolute fitness recorded as survival or death), and sample sizes are relatively small (e.g., n = 50–100) than the traditionally-used linear regression model (Arnold 1983; Lande and Arnold 1983). The logistic regression model relates the survival probability for an individual to that individual’s trait values:
where W is the selection outcome for an individual (1 = survival; 0 = death); W(z) is the survival probability for a set of traits (z1…zn); α = (α1, α2, …, αk)T are the logistic regression coefficients for traits (z1…zn); T signifies matrix transposition; and α0 is an intercept (Janzen and Stern 1998). Multiple logistic regression was done with absolute fitness (survival = 1; death = 0) as the response variable (Janzen and Stern 1998) and whelk shell traits as the predictor variables. Logistic regression coefficients were converted into linear regression coefficients (i.e., selection gradients as in Lande and Arnold 1983) using the methods described in Janzen and Stern (1998), which gives the average gradient vector (βavggrad) and requires calculating W(z) for each individual (Janzen and Stern 1998).
In addition to calculating βavggrad, we also calculated selection differentials, which are the univariate equivalent to a selection gradient (Janzen and Stern 1998). Standardized selection differentials and selection gradients indicate the direction and strength of selection: the magnitudes of these quantities correspond to the strengths, whereas the signs of these quantities correspond to the directions (Arnold 1983; Lande and Arnold 1983). For example, a positive selection differential would show, e.g., that whelks with thicker shells are more likely to survive crab predation than those with thinner shells. The selection differential is the difference between trait means before and after selection. We calculated selection differentials for each whelk using:
S = μx—μz/σz.
where μx represents the average focal trait of the surviving whelks after selection, μz is the average focal trait of all the whelks before selection, and σz is the standard deviation of the focal trait before selection (Janzen and Stern 1998).
Results
Relationship between shell shape variation and crab predation risk in the field
Shell traits were highly correlated in N. lamellosa (electronic supplementary Fig. 1S) and amplification of defensive shell traits was associated with risk of crab predation across collection sites. Whelks from sites with higher crab predation risk (Table 1) had thicker apertural lips (OLS regression; R2 = 0.70, df = 6, P = 0.02; Fig. 3a), narrower aperture openings (OLS regression, R2 = 0.85, df = 6, P < 0.001; Fig. 3b), and wider shells with shorter spires (OLS regression, RW1; R2 = 0.71, df = 6, P = 0.017; Fig. 3c; electronic supplementary Fig. 2S). The proportion of individuals with toothed shells was not related to crab predation pressure, although this relationship approached significance (linear regression, R2 = 0.50, df = 6, P = 0.07; Fig. 3d). There was no effect of region on crab predation pressure (ANOVA: F1,3 = 0.73; P = 0.43). Relative warp 1 (RW1; shell shape) was characterized mainly by the length to width ratio of the shell (OLS regression, R2 = 0.31, df = 702, P < 0.001; AIC = -143.41). Increased shell strength (force-to-fracture, N) was also associated with increased crab predation risk (OLS regression; R2 = 0.83, df = 346, P = 0.004; Fig. 4). Force to fracture was best explained by the additive effect of shell thickness and RW1 (multiple linear regression; R2 = 0.33, df = 345, P < 0.001; AIC = 414.5).

Relationships between crab predation risk and a apertural lip thickness, b apertural aspect ratio, c RW1, which accounted for 36% of the total shape variation. Thin–plate spline deformation grids representing the average observed range in RW1 are shown to facilitate visualization of shell shapes corresponding to highest and lowest observed values. Negative values of RW1 are associated with wider shells with shorter spires and positive values are associated with elongate shells with taller spires, and d proportion of individuals with apertural teeth in Nucella lamellosa. Dashed lines indicate ordinary least squares fit with 95% confidence intervals

Relationship between crab predation risk and the force (N) required to fracture shells of Nucella lamellosa. Dashed lines indicate ordinary least squares fit with 95% confidence intervals
Experiment 1: Influence of apertural teeth on crab predation
In Experiment 1, 41 N. lamellosa were consumed by C. productus. Crabs consumed a total of 20 whelks with apertural teeth removed, leaving 32 individuals intact with no sign of attack. Crabs attacked but were unsuccessful (as noted by ‘peeled’ or chipped apertural lips in surviving whelks) at consuming 17 individuals with apertural teeth removed. Crabs consumed a total of 21 whelks with intact apertural teeth, 27 were left intact, and 22 were unsuccessfully attacked. Crabs consumed the same number of whelks with apertural teeth intact and with apertural teeth removed (chi-square test, χ2 = 1.08, df = 2, P = 0.58; electronic supplementary Fig. 3S).
Experiment 2: Selection on whelk shell traits
Multiple logistic regression indicated that shell thickness was the only trait affecting prey survival, but shell length, aperture area, and shell shape (RW1) were not significant factors on prey survival (Table 2). The resulting selection gradient (βavggrad) for shell thickness was small and positive, but the selection gradients for all other traits were not significant (Table 2). Selection differentials were positive and statistically significant for shell thickness (0.39) and apertural area (0.33; Table 2) indicating that crabs selected whelks with thinner shells with wider aperture openings (Fig. 5). Crabs did not select for wider shells with shorter spires (Table 2).

Frequency distribution of apertural lip thickness a and apertural aspect ratio b or surviving (yellow) and non–surviving (blue) Nucella lamellosa. Dashed vertical lines indicate means for each group
Discussion
Our results indicate the defensive shell traits of N. lamellosa positively covary among populations in the field and that this trait variation is positively associated with predation risk from shell-breaking crabs. Even though we only sampled in two regions, we captured a large portion of the variation that N. lamellosa exhibits by sampling in California and Washington. Our findings complement prior observational and experimental work (Crothers 1983; Appleton and Palmer 1988; Dewitt et al. 2000; Bourdeau 2009, 2012) and suggest that shell shape, shell thickness, apertural area, and apertural teeth should be under strong directional selection imposed by crab predation. However, the functionality of these traits has not been previously tested and the selection differentials from our laboratory selection episodes indicate that predation by N. lamellosa’s major crab predator C. productus selects mainly for thicker shells with narrower apertures in this species. Results of the multiple logistic regression suggested that shell length, apertural area, shell shape, and apertural teeth did not factor into whelk survival—only shell thickness had a significant effect. The resulting selection gradient (βavggrad) for shell thickness was weak, but positive, while the selection gradients for all other traits were not significant. These findings confirm previous work that showed that thicker shells provide an adaptive advantage for N. lamellosa against predation from C. productus (Palmer 1985a). Boulding (1984) found a similar result for C. productus—preying on thin- and thick-shelled clams (Protothaca staminea).
Whereas our estimate of βavggrad on N. lamellosa shell thickness (0.10) was significant, it represents relatively weak directional selection (Kingsolver et al. 2001). However, Kingsolver et al. (2001) and Hoekstra et al. (2001) concluded that directional selection on most traits is weak based on their summary of variance-standardized selection gradients; with selection on survival tending to be weaker than selection on fecundity or mating success. Thus, our results are comparable to the median value (0.16) for directional selection found in a variety of natural populations (Kingsolver 2001). We do note that selection, as estimated here and elsewhere, is measured relative to a fitness component and not to true fitness (Hereford et al. 2004). It is interesting that selection on N. lamellosa shell traits by C. productus was either weak or nonsignificant, because the shell traits of whelks and other marine gastropods are generally considered to have been shaped by significant selection pressure by shell-breaking predators like crabs (Vermeij 1977). However, even very small selection coefficients, can have large evolutionary effects over the geologic timescales (Lande 1979) that comprise the co-evolutionary history of crabs and marine gastropods (Vermeij 1977).
Selection differentials are the univariate equivalents of selection gradients and describe the total intensity of selection acting on a trait, including both direct selection on that trait and indirect selection on that trait via direct selection on a correlated trait. In N. lamellosa, lip thickness is strongly correlated with aperture aspect ratio (electronic supplementary Fig. 1S), because a thicker apertural lip makes the distance between the columellar lip and the apertural lip narrower. A thicker apertural lip and a concomitant narrower apertural opening may work together (e.g., trait cospecialization) to prevent crab peeling attacks by making it more difficult for the crab to insert its chelae into the whelk’s apertural opening. Due to trait correlations such as these, the selective effect on one trait in a selection gradient (i.e., multivariate) analysis can be hidden by the effects of a correlated trait. It is likely that the effects of aperture area were not evident in our selection differential analysis because of its correlation with lip thickness. Trait correlations such as these are common in prey defenses and they are expected due to adaptive trait integration.
It is possible that we may have underestimated the strength of directional selection on shell traits in natural populations of N. lamellosa by imposing selection episodes in the laboratory. C. productus generally takes longer to manipulate and break open thick-shelled compared to thin-shelled Nucella (Palmer 1985a; Bourdeau 2013) and under natural conditions it is possible that crabs are more likely to give up on thicker-shelled whelks under the conflicting demands of other environmental stimuli. There may also be correlated traits other than the ones we measured in this study that act as an indicator of whelk shell thickness. For example, thick-shelled N. lamellosa tend to have smooth, rather than frilly shells (Palmer 1985a; Newson et al. 2018). In addition, differences in the hardness via microsctructural differences in thick- and thin-shelled whelks (Avery and Etter 2006; Bourdeau 2010b) may provide a tactile cue to crab predators. Another intriguing possibility is that variation in anti-predator behavior between the thick- and thin-shelled whelks may have affected crab prey choice. For example, N. lamellosa is known to respond to the chemical cues of crabs consuming conspecific snails by crawling under rocks or out of the water (Appleton and Palmer 1988; Bourdeau 2010a). Although we did not provide refugia for whelks in our experiments, we noticed that many individuals began to climb up the sides and to the tops of the experimental aquaria once crabs began feeding. Although all whelks were technically accessible to crabs within the experimental aquaria, if those that responded behaviorally were more difficult to detect or attack, the behavioral response may have affected selection on morphology, particularly if thinner, less well-defended individuals were more likely to respond behaviorally (i.e., trait compensation; DeWitt et al. 1999) as has been observed in other gastropods responding to the chemical scent of shell-breaking predators (Cotton et al. 2004). Finally, it is possible that the variation in shell shape we observed is at least partly a consequence of environmental factors other than crabs, like predatory sea stars (e.g., Bourdeau 2009) or wave action and currents (e.g., Crothers 1983; Pascoal et al. 2012). For example, the predatory sea star Pisaster ochraceus induces elongate, high-spired shells in N. lamellosa and this shell phenotype reduces risk of predation by the sea star more than crab-induced shell shape changes reduce predation by crabs (Bourdeau 2009). Thus, there is an asymmetrical functional trade-off between shell phenotypes that suggests selection by sea stars on shell aspect ratio may be stronger than that of crabs. Furthermore, plastic and heritable variation in shell thickness have shown to be associated with both crab predation risk and wave action in the north Atlantic congener N. lapillus (Pascoal et al. 2012). Future studies should, therefore, incorporate the effects of the abiotic environment with the multiple predator environment that N. lamellosa experiences on shell shape.
Surprisingly, we found no evidence for direct or indirect selection on apertural teeth. We expected to detect directional selection on apertural teeth as this trait has been shown to be induced directly by chemical exudates from C. productus, whether snails are well-fed or starved (Appleton and Palmer 1988). However, despite the finding that apertural teeth are directly induced by crab risk cues in both N. lamellosa (Appleton and Palmer 1988) and its congener N. lapillus (Palmer 1990), the functional significance of this putative inducible defense has not been tested until now. We found that the presence of apertural teeth had no effect on N. lamellosa survival in our manipulative experiment. Furthermore, although we found a weak positive association between apertural teeth and risk of crab predation in the field, this relationship was not significant. Together, these findings cast some doubt on the functional significance of apertural teeth in deterring C. productus attacks on N. lamellosa, at least for the size class of crabs and N. lamellosa used in our experiment. Apertural teeth are thought to partially occlude the apertural opening of the shell, making it more difficult for crabs to insert their chelae into the aperture for a peeling attack (Vermeij 2015). However, toothed shells without concomitant reinforcement of the shell wall would still be vulnerable to crushing attacks by crabs (Vermeij 1978) and C. productus employs both peeling and crushing attacks (Zipser and Vermeij 1978), so apertural teeth may not provide a strong enough fitness advantage against this particular crab predator. Finally, we only used a relatively narrow size range of C. productus from a single location in our laboratory experiments, which may have affected our results. This only allows broad inference about crab selection differentials and gradients within this size range. For example, it is possible that at whelk sizes smaller than those in our experiments, apertural teeth deter entry by crabs smaller than those used in our experiment and testing over a larger range of both whelk and crab sizes could reveal the functional significance of apertural teeth in N. lamellosa.
There is marked interpopulation variation in the shell sculpture and shell shape of N. lamellosa (Spight 1973; Palmer 1985a; Bourdeau 2012) that is correlated with risk from crab predation (Appleton and Palmer 1988; Bourdeau 2012, this study). Whelks from crab-rich habitats have wider shells with shorter spires (all traits associated with RW1 in this study). These traits are thought to spread crushing forces more evenly over the surface of the shell (DeWitt et al. 2000) and make spire-clipping attacks more difficult (Edgell and Neufeld 2008). We found that both RW1 (shell shape) and apertural lip thickness contribute to shell strength, suggesting that shell shape development and lip thickening have evolved jointly in habitats where predation risk from C. productus is high. Thus, it was also surprising that we found no evidence for direct or indirect selection on shell shape. One possibility for the lack of significant selection on shell shape by crab predation in our experiment is that in many habitats, whelks may be exposed to both crabs and other predators (mainly sea stars; Bourdeau 2009). Sea star predation selects for elongate, high-spired shells, which protect the whelks from detection or shell entry attacks by sea stars (Bourdeau 2009). Consequently, diffuse selection on shell shape by both predators may occur, because the combined presence of both predators represents a more common environment than a ‘crab-only’ environment (Strauss et al. 2005). The interacting influences of multiple predator species can generate novel evolutionary forces on defensive traits, resulting in selection regimes that are not often predictable from isolated interactions of prey with a single predator species (DeWitt and Langerhans 2003, Strauss et al. 2005).
Thicker-shelled N. lamellosa individuals increase both the time and energy required by C. productus to successfully break into a shell (Bourdeau 2013), and thicker shelled individuals also provide less food value of snails to crabs, because thicker-shelled individuals have relatively less soft tissue (Bourdeau 2010a). Thicker shells thus appear to influence the probability of being eaten and are selected for, albeit weakly, by C. productus. Because selection episodes occurred over 2–3d, and inducible shell-thickening takes on the order of weeks to months in N. lamellosa (Bourdeau 2010a, b), the shell phenotype of individual whelks remained constant during the episode of selection. Thus, crabs could have selected for individuals with either greater levels of constitutive or inducible shell thickness, or both (Appleton and Palmer 1988; Bourdeau 2012). Future studies should quantify to what degree selection imposed by crab predation acts on both constitutive levels of shell thickness and shell thickness inducibility (i.e., plasticity) in N. lamellosa.
Data accessibility
Data are available upon request from authors.
Code accessibility
Available upon request from authors.
References
Appleton RD, Palmer AR (1988) Water–borne stimuli released by predatory crabs and damaged prey induce more predator–resistant shells in a marine gastropod. Proc Natl Acad Sci 85:4387–4391. https://doi.org/10.1073/pnas.85.12.4387
Arnold SJ (1983) Morphology, performance and fitness. Am Zool 23:347–361
Avery R, Etter RJ (2006) Microstructural differences in the reinforcement of a gastropod shell against predation. Mar Ecol Prog Ser 323:159–170. https://doi.org/10.3354/meps323159
Blundon JA, Vermeij GJ (1983) Effect of shell repair on shell strength in the gastropod Littorina irrorata. Mar Biol 76:41–45
Bookstein FL (1989) Principal warps: Thin–plate splines and the decomposition of deformations. IEEE Trans Pattern Anal Mach Intell 11:567–585
Boulding EG (1984) Crab–resistant features of shells of burrowing bivalves: decreasing vulnerability by increasing handling time. J of Exp Mar Biol and Ecol 76:201–223
Bourdeau PE (2009) Prioritized phenotypic responses to combined predators in a marine snail. Ecology 90:1659–1669. https://doi.org/10.1890/08-1653.1
Bourdeau PE (2010a) An inducible morphological defense is a passive by–product of behaviour in a marine snail. Proc R Soc B: Biol Sci 277:455–462. https://doi.org/10.1098/rspb.2009.1295
Bourdeau PE (2010b) Cue reliability, risk sensitivity and inducible morphological defense in a marine snail. Oecologia 162:987–994. https://doi.org/10.1007/s00442-009-1488-5
Bourdeau PE (2012) Intraspecific trait cospecialization of constitutive and inducible morphological defenses in a marine snail from habitats with different predation risk. J of An Ecol 81:849–858. https://doi.org/10.1111/j.1365-2656.2012.01965.x
Bourdeau PE (2013) Morphological defense influences absolute, not relative, nonconsumptive effects in marine snails. Behav Ecol 24:505–510. https://doi.org/10.1093/beheco/ars191
Burnham KP, Anderson DR (2004) Multimodel inference: understanding AIC and BIC in model selection. Sociol Methods Res 33:261–304. https://doi.org/10.1177/0049124104268644
Crothers JH (1982) Shell shape variation in dog–whelks (Nucella lapillus (L.)) from the West Coast of Scotland. Biol J Linn Soc 17:319–342. https://doi.org/10.1111/j.1095-8312.1982.tb02025.x
Crothers JH (1983) Variation in dog–whelk shells in relation to wave action and crab predation. Biol J Linn Soc 20:85–102. https://doi.org/10.1111/j.1095-8312.1983.tb01591.x
Crothers JH (1984) Some observations on shell shape variation in Pacific Nucella. Biol J Linn Soc 21:259–281. https://doi.org/10.1111/j.1095-8312.1984.tb00365.x
Currey JD, Hughes RN (1982) Strength of the dogwhelk Nucella lapillus and the winkle Littorina littorea from different habitats. J Anim Ecol. https://doi.org/10.2307/4309
Cotton PA, Rundle SD, Smith KE (2004) Trait compensation in marine gastropods: shell shape, avoidance behavior, and susceptibility to predation. Ecology 85:1581–1584. https://doi.org/10.1890/03-3104
Dalziel B, Boulding EG (2005) Water-borne cues from a shell-crushing predator induce a more massive shell in experimental populations of an intertidal snail. J Exp Mar Biol Ecol 317:25–35. https://doi.org/10.1016/j.jembe.2004.11.015
DeWitt TJ, Langerhans RB (2003) Multiple prey traits, multiple predators: keys to understanding complex community dynamics. J Sea Res 49:143–155. https://doi.org/10.1016/S1385-1101(02)00220-4
DeWitt TJ, Robinson BW, Wilson DS (2000) Functional diversity among predators of a freshwater snail imposes an adaptive trade-off for shell morphology. Evol Ecol Res 2:129–148
Dewitt TJ, Sih A, Hucko JA (1999) Trait compensation and cospecialization in a freshwater snail: size, shape and antipredator behaviour. J Anim Behav 58:397–407. https://doi.org/10.1006/anbe.1999.1158
Edgell TC, Neufeld CJ (2008) Experimental evidence for latent developmental plasticity: intertidal whelks respond to a native but not an introduced predator. Biol Lett 4:385–387. https://doi.org/10.1098/rsbl.2008.0204
Hereford J, Hansen TF, Houle D (2004) Comparing strengths of directional selection: how strong is strong? Evolution 58:2133–2143. https://doi.org/10.1111/j.0014-3820.2004.tb01592.x
Hoekstra HE, Hoekstra JM, Berrigan D, Vignieri SN, Hoang A, Hill CE, Beerli P, Kingsolver JG (2001) Strength and tempo of directional selection in the wild. Proc Natl Acad Sci 98:9157–9160. https://doi.org/10.1073/pnas.161281098
Hughes RN, Elner RW (1979) Tactics of a predator, Carcinus maenas, and morphological responses of the prey Nucella lapillus. J Anim Ecol. https://doi.org/10.2307/4100
Janzen FJ, Stern HS (1998) Logistic regression for empirical studies of multivariate selection. Evolution 52:1564–1571. https://doi.org/10.1111/j.1558-5646.1998.tb02237.x
Kingsolver JG, Gomulkiewicz R, Carter PA (2001) Variation, selection and evolution of function-valued traits in Microevolution rate, pattern, process. Springer, Dordrecht
Kitching JA (1976) Distribution and changes in shell form of Thais spp. (Gastropoda) near Bamfield. BC J Exp Mar Biol Ecol 23:109–126. https://doi.org/10.1016/0022-0981(76)90135-0
Kitching JA, Lockwood J (1974) Observations on shell form and its ecological significance in thaisid gastropods of the genus Lepsiella in New Zealand. Mar Biol 28:131–144
Kitching JA, Muntz L, Ebling FJ (1966) The ecology of Lough Ine. XV. The ecological significance of shell and body forms in Nucella. J Anim Ecol 35(1):113–126. https://doi.org/10.2307/2693
Lande R (1979) Quantitative genetic analysis of multivariate evolution, applied to brain: body size allometry. Evolution. https://doi.org/10.2307/2407630
Lande R, Arnold SJ (1983) The measurement of selection on correlated characters. Evolution 37:1210–1226. https://doi.org/10.2307/2408842
Langerhans RB (2007) Evolutionary consequences of predation: avoidance, escape, reproduction, and diversification. Predation in organisms. Springer, Berlin, Heidelberg
Lowell RB, Fletcher CR, Grahame J, Mill PJ (1994) Ontogeny of shell morphology and shell strength of the marine snails Littorina obtusata and Littorina mariae: different defense strategies in a pair of sympatric, sibling species. J Zool 234:149–164. https://doi.org/10.1111/j.1469-7998.1994.tb06062.x
Masel J (2011) Genetic drift. Curr Biol 21:R837–R838
Molinaro DJ, Stafford ES, Collins BM, Barclay KM, Tyler CL, Leighton LR (2014) Peeling out predation intensity in the fossil record: a test of repair scar frequency as a suitable proxy for predation pressure along a modern predation gradient. Palaeogeogr Palaeoclimatol Palaeoecol 412:141–147. https://doi.org/10.1016/j.palaeo.2014.07.033
Newson O, Basi R, Palmer AR (2018) Lamellose Axial Shell Sculpture Reduces Gastropod Vulnerability to Sea Star Predation. Biol Bull 235:24–29
Palmer AR (1985a) Adaptive value of shell variation in Thais lamellosa: effect of thick shells on vulnerability to and preference by crabs. Veliger 27:349–356
Palmer AR (1985b) Genetic basis of shell variation in Thais emarginata (Prosobranchia, Muricacea). I. Banding in populations from Vancouver Island. Biol Bull 169:638–651
Palmer AR (1990) Effect of crab effluent and scent of damaged conspecifics on feeding, growth, and shell morphology of the Atlantic dogwhelk Nucella lapillus (L.). In Progress in littorinid and muricid biology. Dordrecht: Springer
Pascoal S, Carvalho G, Creer S, Mendo S, Hughes R (2012) Plastic and heritable variation in shell thickness of the intertidal gastropod Nucella lapillus associated with risks of crab predation and wave action, and sexual maturation. PLoS ONE. https://doi.org/10.1371/journal.pone.0052134
Rebach S (1985) Rhythmicity under constant conditions in the rock crab, Cancer irroratus. Bull Mar Sci 36:454–466
Rochette R, Doyle SP, Edgell TC (2007) Interaction between an invasive decapod and a native gastropod: predator foraging tactics and prey architectural defenses. Mar Ecol Prog Ser 330:179–188. https://doi.org/10.3354/meps330179
Rohlf FJ, Marcus LF (1993) A revolution morphometrics. Trends Ecol Evol 8:129–132. https://doi.org/10.1016/0169-5347(93)90024-J
Rohlf FJ (2006) tpsDig, Digitize landmarks and outlines, version 1.44. Stony Brook, NY: Department of Ecology and Evolution, State University of New York
Schlichting CD, Wund MA (2014) Phenotypic plasticity and epigenetic marking: an assessment of evidence for genetic accommodation. Evolution 68:656–672. https://doi.org/10.1111/evo.12348
epúlveda RD, Jara CG & Gallardo CS, (2012) Morphological analysis of two sympatric ecotypes and predator-induced phenotypic plasticity in Acanthina monodon (Gastropoda: Muricidae). J Molluscan Stud 78:173–178. https://doi.org/10.1093/mollus/eyr058
Spight TM (1973) Ontogeny, environment, and shape of a marine snail Thais lamellosa Gmelin. J Exp Mar Biol Ecol 13:215–228. https://doi.org/10.1016/0022-0981(73)90068-3
Stafford ES, Tyler CL, Leighton LR (2015) Gastropod shell repair tracks predator abundance. Mar Ecol 36:1176–1184. https://doi.org/10.1111/maec.12219
Strauss SY, Sahli H & Conner JK (2005) Toward a more trait-centered approach to diffuse (co) evolution. New Phyto, 81–89
Stoks R, Govaert L, Pauwels K, Jansen B, De Meester L (2016) Resurrecting complexity: the interplay of plasticity and rapid evolution in the multiple trait response to strong changes in predation pressure in the water flea Daphnia magna. Ecol Lett 19:180–190. https://doi.org/10.1111/ele.12551
Tyler CL, Stafford ES, Leighton LR (2015) The utility of wax replicas as a measure of crab attack frequency in the rocky intertidal. Marine Biological Association of the United Kingdom. J Mar Biol Assoc UK 95:361. https://doi.org/10.1017/S0025315414001210
Upton GJ (1992) Fisher’s exact test. J R Stat Soc: A (Statistics in Society) 155:395–402
Vermeij GJ (1977) The Mesozoic marine revolution: evidence from snails, predators and grazers. Paleobiology 3(3):245–258
Vermeij GJ (1978) Biogeography and adaptation: patterns of marine life. Harvard University Press
Vermeij GJ (1982) Gastropod shell form, breakage and repair in relation to predation by the crab Calappa. Malacologia 23:1–12
Vermeij GJ (2015) Gastropod skeletal defences: land, freshwater, and sea compared. Vita Malacologica 13:1–25
Vermeij GJ, Currey JD (1980) Geographical variation in the strength of thaidid snail shells. Biol Bull 158:383–389
Wellington GM, Kuris AM (1983) Growth and shell variation in the tropical eastern Pacific intertidal gastropod genus Purpura: ecological and evolutionary implications. Biol Bull 164:518–535. https://doi.org/10.2307/1541260
Yamamichi M, Yoshida T, Sasaki A (2011) Comparing the effects of rapid evolution and phenotypic plasticity on predator-prey dynamics. Am Nat 178:287–304. https://doi.org/10.1086/661241
Zipser E, Vermeij GJ (1978) Crushing behavior of tropical and temperate crabs. J Exp Mar Biol Ecol 31:155–172. https://doi.org/10.1016/0022-0981(78)90127-2
Acknowledgements
We thank the director and staff of the Telonicher Marine Laboratory and Friday Harbor Laboratories for logistical support, as well as J. Paulsen and volunteers from the PEB Lab who helped with field and lab work. Three anonymous reviewers provided constructive criticism on earlier versions of this manuscript. This is a contribution of the Humboldt State University Telonicher Marine Laboratory and the Humboldt Marine and Coastal Sciences Institute.
Funding
This project received no funding.
Author information
Authors and Affiliations
Contributions
LCM, TIK, TEF, and PEB conceived the project. LCM, TIK, and TEF collected the data. LCM and PEB analyzed the data and wrote the manuscript. All authors have consented to the publication of this manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Ethical approval
Experiments comply with the ethical standards of the country in which they were performed. Organisms were collected under SCP 10571 to PEB from the California Department of Fish and Wildlife.
Additional information
Reviewers: undisclosed experts.
Responsible Editor: P. Ramey-Balci.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
McIntire, L.C., Kotas, T., Flagor, T.E. et al. Weak directional selection on inducible defensive shell traits in a marine gastropod by the inducing shell-breaking predator. Mar Biol 168, 58 (2021). https://doi.org/10.1007/s00227-021-03860-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-021-03860-9