
Abstract
The trophic fate of various food sources is of central interest for ecologists, yet not well understood in coastal lagoon food webs. In this field study, fatty acids (FA), stable isotopes (SI), and compound-specific isotopic analysis (CSIA) on FA were used to investigate how diets from oceanic and local sources are retained in a bivalve species (Spondylus crassisquama; Lamarck 1819) along a transect in the Ojo de Liebre lagoon (BCS, Mexico). Results from SI and FA indicated the contribution of oceanic diatoms at the entrance of the lagoon, through 15N enrichment, and higher proportions of 16:1n-7 and 20:5n-3 in digestive glands. In the inner bay, higher abundance of 18-carbon FA (18:1n-9, 18:3n-3, 18:4n-3) suggested a higher contribution of microheterotrophs, including (dino)flagellates and ciliates, to the diet of this bivalve derived from local production. Significant spatial differences for the δ13C of FA highlighted changes in the origin of food sources. Indeed, a 13C depletion was observed in the δ13C of heterotrophic flagellates biomarkers in individuals from the innermost station, revealing that their origin in the diet of bivalves differs within the lagoon, highlighting the importance of local processes (sediment resuspension, remineralization) in the trophic functioning of the lagoon. The δ13C values of FA considered as diatoms biomarkers (16:1n-7 and 20:5n-3) were consistent, which suggests that diatoms assimilated have very similar origins throughout the lagoon. The complementary of the tracers used here allowed for a better understanding of the trophic functioning of this coastal lagoon submitted to oceanic influences.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Coastal lagoons are complex and productive ecosystems, and their primary production is supported by a highly diverse origin of nutrients (Nixon 1982). At the interface between land and ocean, lagoons are characterized by shallow water and a large diversity of potential food sources, including continental inputs, salt marsh, seagrass, macroalgae, and phytoplankton, and resulting in marked spatial patterns in food web interactions (Carlier et al. 2008; Clavier et al. 2014). In coastal lagoons, benthic assemblages are largely dominated by suspension-feeding bivalves (Bachelet et al. 2000). Because these organisms are sedentary primary consumers, they are good indicators of processes occurring at the base of the food web (Marchais et al. 2013). Therefore, studying the nature and the origin of food sources involved in their diet provides a spatially and temporally integrated overview of organic matter transfer between benthic and pelagic systems, and across ecosystem boundaries (i.e., between lagoons and the nearby coastal ocean).
The trophic ecology of suspension-feeding invertebrates inhabiting coastal environments has been extensively investigated over the last 20 years using indirect biomarker-based approaches. Among the most common, the analysis of bulk carbon and nitrogen stable isotopes (SI) has provided valuable insights into the origin and nature of bivalve food sources (Riera 2007; Marchais et al. 2013). Because primary producers typically display contrasted SI composition according to their benthic vs pelagic (France 1995) or freshwater vs marine (Riera and Richard 1996) origin, and because the SI composition of consumers are indicative of their source, this approach is currently widely used to address questions in trophic ecology (Fry 1988; Layman et al. 2012). However, to be efficient, this method requires primary producers displaying contrasted SI composition. Although different species of planktonic microalgae can vary in their SI values (Falkowski 1991; Vuorio et al. 2006), it is generally difficult to differentiate between potential diet sources that share the same habitat (Kharlamenko et al. 2001). Therefore, it remains a challenge for bulk SI ecology to assess the dietary contribution of different primary producers to consumers.
The analysis of fatty acids (FA) is a powerful and complementary approach, which is based on the assumption that most animals have limited ability to biosynthesize polyunsaturated FA (PUFA) or FA of bacterial origin de novo (Budge et al. 2006), and thus, the FA composition of a consumer resembles that of its food source (Dalsgaard et al. 2003). Among the different FA synthesized by primary producers, some of them are considered as trophic biomarkers because they characterize specific food sources; for example, palmitoleic acid (16:1n-7) and eicosapentaenoic acid (20:5n-3) are used as biomarkers of diatoms, and stearidonic acid (18:4n-3) and docosahexaenoic acid (22:6n-3) are abundant in (dino)flagellates (Parrish et al. 2000; Kharlamenko et al. 2001; Alfaro et al. 2006). Therefore, it is possible to assess the contribution of specific taxonomic groups (e.g., diatoms, dinoflagellates, marine plants) in the diet of primary consumers (Kelly and Scheibling 2012); by paying attention when analyzing bivalve tissues presenting different turnover rates resulting in diverse integrating dietary information over time (e.g., the digestive gland (DG), has faster turnover rates than muscle tissues) (Lorrain et al. 2002; Dalsgaard et al. 2003; Nerot et al. 2015). Because SI can be useful in assessing food sources, in particular when using stable C and N isotopes in combination (Lorrain et al. 2002; Schaal et al. 2016) and FA can differentiate between algal, bacterial or even terrestrial sources (Kainz et al. 2002), these two approaches have been successfully used in combination to unravel trophodynamic trajectories in a variety of coastal environments (Alfaro et al. 2006; Jaschinski et al. 2008; Allan et al. 2010). The combined use of SI and FA is based on the assumption that specific SI values or FA molecules are attributed to distinct sources, which is unlikely in complex ecosystems (Kharlamenko et al. 2001). For instance, it would be impossible to characterize the precise origin (i.e., oceanic or coastal) of the diet of a suspension feeder assimilating planktonic dinoflagellates and benthic diatoms using only bulk SI and FA approaches.
Compound-specific isotope analysis (CSIA) represents an ecologically complementary approach, through which the identification of the SI composition of source-specific compounds can characterize their origin. For example, CSIA has been successfully used in trophic ecology to highlight the contribution of ice algae to the diet of bivalves (Gaillard et al. 2017), or dietary shifts in benthic invertebrates at the vicinity of finfish farming (Colombo et al. 2016).
Lagoon, such as the Ojo de Liebre lagoon (Baja California Sur, Mexico), are of particular interest for trophic ecology as they rely on a wide diversity of both local sources (autochthonous) such as local phytoplankton or resuspended benthic sources, including eelgrass detritus (Zostera marina; Linnaeus, 1753), and allochthonous sources, such as oceanic phytoplankton supplied to the lagoon by tide current. Oceanic phytoplanktonic input could be more intense during important oceanic events occurring from April to June in this region (Du et al. 2015).
In this context, the aim of this study was to apply three trophic biomarker-based approaches to examine spatial patterns of organic matter sources and their biochemical composition assimilated by S. crassisquama. It was hypothesized that lower oceanic phytoplankton availability in the inner part of the lagoon, which is less connected to the ocean, would result in higher reliance on local food sources. Stable isotope, FA, and CSIA values in consumers located close to the ocean were, thus, expected to resemble those of the ocean, testing the effect of spatial diet proximity to the ocean and how far its impact is perceived inside the lagoon. Biomarker values of consumers furthest away from the ocean were expected to resemble autochthonous diet sources, highlighting the effect of local diet incorporation.
Materials and methods
Sampling
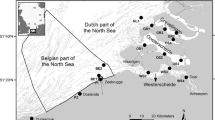
The Ojo de Liebre lagoon is a 446-km2 lagoon situated on the Pacific coast of Baja California hosting one of the most important bivalve fisheries (in particular Spondylus crassiquama; Lamarck 1819) of western Mexico. Three stations were sampled in June 2017 along a transect (Fig. 1); Station 1: located 3.5 km outside the bay; Station 2: at 6.1 km from the mouth, inside the lagoon; and Station 3: 15.5 km inside the lagoon. Sampling depths ranged from 7 to 10 m. Stations 1 and 3 corresponded to the most outer and inner sites, respectively, where the target species, S. crassisquama, could be found. At each station, 10 individuals of S. crassisquama of similar size (~ 12 cm of length), and potential food sources (particulate organic matter; POM, and, sediment–water interface organic matter; SOM) were collected by scuba divers. The Zostera marina fragments, at two different degradation stages (later called fresh and detrital), were also collected from the shore at the eastern part of the sampling area. Bivalves were placed in cooler boxes with water from sampling sites at ambient temperature, while collected surface water and sediment cores and eelgrass detritus were protected from light using black plastic bags until transportation (within 3 h) to the lab facilities at the Northwestern Center for Biological Research (CIBNOR, Guerrero Negro, BCS). Once in the laboratory, bivalve DG and adductor muscles were dissected, packed and stored in liquid nitrogen.

(Map modified from Google earth)
Location of the 3 sampling stations in the Ojo de Liebre lagoon (Baja California Sur, Mexico). Station 1 is located at 3.5 km outside the bay. Stations 2 and 3 are located inside the lagoon, respectively, at 6.1 km and 15.5 km from the mouth of the lagoon.
Food sources (SOM, POM, and eelgrass fragments) were prepared for analysis by the following methods; (a) sediment–water interface were soaked in filtered sea water (GF/F 0.7 µm), and allowed to settle. The supernatant containing fine organic particles (SOM), potentially affected by resuspension events and, therefore, available to benthic suspension feeders, was filtered on pre-combusted (6 h, 450 °C) GF/F filters. Four subsamples of SOM (two for SI, and two for FA analyses) were prepared for each sampling station, packed and frozen in liquid nitrogen; (b) particles from surface water were filtered on pre-combusted (6 h, 450 °C) GF/F filters until clogging. Immediately after filtration, filters subsamples of POM (n = 2 for SI, and n = 2 for FA analyses) were packed and frozen in liquid nitrogen immediately after filtration, and; (c) large epiphytes were first removed from eelgrass fragments, which were then packed and stored in liquid nitrogen. Once at the main laboratory (CIBNOR, La Paz), all samples were stored at − 80 °C until further analysis.
Stable isotopes analysis
Muscles and eelgrass samples were freeze-dried (24 h) and homogenized using a ball mill. Because bivalve muscles are generally poor in lipids, lipids were not removed from muscle tissues prior to SI analysis (Lorrain et al. 2002). Indeed, the measured C:N ratios did not exceed 3.5, rendering lipid removal unnecessary for SI analysis (Post et al. 2007). Muscle tissues (~ 25 mg dry weight) and particles of SOM and POM (scraped off from the freeze-dried filters; ~ 1.5 mg dry weight) were packed in tin capsules (8 × 5 mm) for subsequent bulk SI analysis.
All samples were analyzed by continuous flow on a Flash EA 2000 IRMS elemental analyzer coupled to a isotope ratio mass spectrometer (IRMS; Thermo Fisher Scientific Delta V Plus). Isotopic calibration was done using international standards (IAEA-600 Caffeine, IAEA-CH-6 Sucrose and IAEA-N-2 Ammonium Sulphate). The standard deviation calculated with the acetanilide working standard (Thermo Scientific), repeatedly measured after seven samples, was ± 0.1‰ (n = 19) for both δ13C and δ15N. Results were expressed in delta (δ) units with respect to international standards (Vienna Pee Dee Belemnite for δ13C and atmospheric nitrogen for δ15N), following the equation: δ13C or δ15N = [(Rsample/Rstandard) − 1] × 103 (expressed in ‰), where R is 13C/12C or 15N/14N.
Fatty acid analysis
Lipid extraction
Total lipids of freeze-dried and homogenized bivalve DG and eelgrass fragments (between 0.2 and 0.4 g), and POM and SOM filter samples were extracted using chloroform/methanol (6 mL; 2:1; v/v) applying a modified method of Folch et al. (1957). Samples were then sonicated at 4 °C during 5 min, to ensure complete extraction of lipids. Until further analysis, all lipid extracts were stored at − 20 °C under nitrogen atmosphere to avoid oxidation.
Separation of neutral lipids
Digestive gland lipids were separated into neutral and polar lipids (the latter was not analyzed in the present study) following the method described elsewhere (Le Grand et al. 2014). In brief, an aliquot (1/6) of the total lipid extract was evaporated under nitrogen, recovered with 3 washes using chloroform/methanol (0.5 mL; 98:2; v/v) and spotted at the top of a silica gel column (40 mm × 4 mm, silica gel 60A 63–200 µm rehydrated with 6% H2O; 70–230 mesh). Neutral lipids (NL) were eluted using chloroform/methanol (10 mL; 98:2; v/v) and collected in glass vials containing an internal standard (2.3 µg of tricosanoic acid; C23:0). Lipid fractions were then stored at − 20 °C until further analyses.
Fatty acid analysis by gas chromatography
The NL fraction of DG as well as aliquots of POM, SOM and eelgrass total lipid extracts was evaporated to dryness under nitrogen. Fatty acids methyl esters (FAME) were obtained by an acidic transesterification following a modified method by (Le Grand et al. 2014). In brief, transesterification was performed by the addition of 0.8 mL of a H2SO4/methanol mixture (3.4%; v/v) to the dried extracts containing NL and heated at 100 °C for 10 min. After cooling, hexane (0.8 mL) and distilled water saturated in hexane (1.5 mL) were added. Vials were then homogenized and centrifuged (at 738g for 1 min at 20 °C). Only the organic phase-containing FAME was kept and washed two more times with distilled water saturated in hexane (1.5 mL). At each step, the aqueous phase was discarded.
Analyses of FAME were conducted on a Varian CP8400 gas chromatograph (GC), by simultaneous separation on two columns with two different phase polarities: one polar (ZBWAX: 30 m × 0.25 mm ID × 0.2 µm, Phenomenex) and another apolar column (ZB5HT: 30 m × 0.25 mm ID × 0.2 µm, Phenomenex). The GC runs followed these temperature programs: the GC oven temperature was raised to 150 °C at 50 °C min−1, then to 170 °C at 3.5 °C min−1, to 185 °C at 1.5 °C min−1, to 225 °C at 2.4 °C min−1, and finally to 250 °C at 5.5 °C min−1 and maintained for 15 min. The GC was equipped of two splitless injectors regulated at 220 °C, and two flame-ionization detectors (280 °C), using hydrogen as vector gas. Identification of FAME was realized by comparison of their retention times with those of commercial standards from Sigma (Supelco 37 Component FAME Mix, the PUFA no. 1 and no. 3, and the Bacterial Acid Methyl Ester Mix) and an in-house standard mix from marine bivalves and microalgae. The FA content (%) was expressed as the mass percentage of the total FA content.
Compound-specific isotopic analysis on fatty acids
Five bivalve samples previously analyzed for FA composition were randomly selected for compound-specific isotopic analysis. The utilization of GC (Thermo Fisher Scientific GC ISOLINK TRACE ULTRA) allowed the separation of FAME using the same apolar phase (Agilent DB5) and the same analytical program as for GC-FID analysis (see above). Each of the FA was converted into CO2 by combustion in the ISOLINK furnace and transferred to the CONFLO IV interface and then introduced to the IRMS (Delta V Plus; Thermo Fisher Scientific). As above, FAME were identified by comparison of their retention time with those of commercial standards and in-house standard mixtures as described above. Both FA 18:1n-9 and 18:3n-3 did coelute on the apolar column used in GC-C-IRMS analysis, and were, therefore, analyzed simultaneously. Fatty acids kept for δ13C analyses were selected based on their abundance and detection in CSIA (i.e., with amplitudes > 800 mV).
Calibration of δ13C values
The δ13C values were calibrated using the F8-3 standard mixture of eight acid ethyl and methyl esters (14:0, 16:0, 18:0, and 20:0, with δ13C values ranging from − 26.98 ± 0.02‰ to − 30.38 ± 0.02‰) supplied by Indiana University Stable Isotope Reference Materials, inserted before and after every 3 analytical replicates of each sample.
Correction of the δ13C contribution of the carbon added by trans-esterification
Because of the addition of a methyl group derived from the methanol used in the transesterification reaction, δ13C of FAME analyzed here differed from the original δ13C of FA present in source material. To correct the contribution of the carbon from this additional methyl group, several free FA (14:0, 16:0, 16:1n-7, 17:0, 18:0, 18:1n-9, 18:2n-6, 18:3n-3, 18:3n-6, 20:0, 20:4n-6, 20:5n-3, 22:0, 23:0, and 24:0; Sigma), were trans-esterified with the same methanol as the one used for samples. Prior to trans-esterification, the δ13C values of each of those free FA were determined using continuous flow on a Flash EA2000 elemental analyzer coupled to a Delta V Plus mass spectrometer (Thermo Fisher scientific). After trans-esterification of free FA, the δ13C values of the resulting FAME were measured using the GC-C-IRMS system. Then, an average correction was calculated using the following equation:
where n is the number of carbon atoms in the free FA (Abrajano et al. 1994). The average δ13C value (− 39.6‰) was calculated for the methyl-derived carbon based on the difference between the δ13C values of the corresponding free FA and their respective FAME and this value was used to correct the FAME δ13C by rearranging the above equation. Further studies found no kinetic isotope effect associated with trans-esterification (Budge et al. 2008; Graham et al. 2014); thus, this aspect was not considered for δ13C analysis.
Analytical error
The analytical error (consisting of the SD of 10 analyses of the C16:0 FA laboratory standard interspersed during the samples run) was 0.2‰. Therefore, the analytical precision was estimated to be ≤ 0.2‰.
Statistical analysis
Normality (Shapiro–Wilks test) and homoscedasticity (Bartlett test) were tested prior to parametric or non-parametric tests. Parametric tests were applied when conditions of normally distributed data and homoscedasticity were satisfied; otherwise, non-parametric tests with Bonferroni’s adjustment method were performed. The isotopic variation in bivalve muscles was compared among stations by Kruskal–Wallis (KW) tests followed by Conover–Iman multiple comparisons. Only the FA > 0.5% of total FA were considered to compare the dissimilarity among stations, by a permutational variance analyses (PERMANOVA). Subsequently, a test of similarity percentages analyses (SIMPER) was carried out to assess the most discriminant FA responsible of the difference among stations (> 80%). Each FA accounting for > 80% of dissimilarity among stations, and presenting an abundance permitting its detection in CSIA in all DG samples, was accounted. For that, one-way analysis of variance (ANOVA) followed by Tukey’s HSD, or KW tests followed by Conover–Iman multiple comparison were carried out to compare the difference of their percentage of neutral lipids, and their respective δ13C values, between the individuals from the three stations. All statistical analyses and graphics were performed with the free software R (Core Team 2017), with R Version 3.4.1 (2017 06 30).
Results
Stable isotopes analyses
No significant differences in bulk δ13C values were observed in bivalve muscles between the different stations (KW test, H25 = 2.12, P = 0.35). In contrast, a significant δ15N difference was observed (KW test, H25 = 17.71, P < 0.001) (Fig. 2). Individuals from stations 1 and 2 displayed similar δ15N (8.3‰ and 8.5‰, respectively) (Fig. 2), while station 3 was 15N depleted (7.2‰). For logistical reasons, only two replicates of POM and SOM could be sampled per site, which could not allow to statistically compare the results for these potential food sources. The pattern observed in POM was the same as that observed for bivalve muscles with stations 1 and 2 being 15N enriched compared to station 3 (from 6.5 and 6.9‰ for stations 1 and 2, respectively, to 4‰ at station 3) (Fig. 2). The POM δ13C value sampled outside the lagoon (− 22.8‰) was slightly lower than at stations 2 and 3 (− 21.1‰ and − 20.9‰, respectively). SOM displayed quite a different spatial pattern than the one observed for POM, with station 1 being 15N enriched and 13C depleted (δ15N = 7.3‰, δ13C = − 14.5‰) compared to the two inner stations 2 and 3 (δ15N = 5.2‰ and 4.6‰, δ13C = − 10.2‰ and − 13‰, respectively). Both samples of Zostera marina were 13C enriched (− 10.0‰ for fresh eelgrass and − 11.2‰ for detrital eelgrass) compared to other samples. Fresh eelgrass displayed higher δ15N than detrital eelgrass (5.6‰ and 1.8‰, respectively) (Fig. 2).

Stable Isotope (SI) bi-plot (δ13C and δ15N, mean values ± SD when n > 2) of the adductor muscles of Spondylus crassisquama and of its potential food sources, in June 2017, at the 3 sampled stations. Samples from station 1 are shown in dark blue, 2 in light blue, and 3 in green. Bivalves are represented by squares, POM by circles, SOM by triangles, and eelgrass by crosses (* fresh; x detrital)
Fatty acids composition
The FA composition of neutral lipids of DG varied significantly among stations (PERMANOVA, F = 48.76, P < 0.001). Individuals from stations 1 and 2, the closest to the mouth of the bay, were characterized by higher contents of 14:0, 20:4n-6, and the diatom biomarkers 16:1n-7 and 20:5n-3 than individuals from station 3 (ANOVA or KW tests followed by post hoc; Table 1). The PUFA 22:6n-3 was slightly more abundant in individuals from station 1 (12.1%) than in those from station 2 (11.4%) and station 3 (10.5%) (Table 1). The DG of individuals from the most inner station in the lagoon (station 3) had significantly higher contents of C18-FA (18:0, 18:1n-9, 18:3n-3, and 18:4n-3) and C16-FA (ANOVA or KW tests followed by post hoc; Table 1).
Some FA in POM samples presented the same spatial variations than in bivalve DG, in particular 16:1n-7 that presented higher proportions at the external station (Table 2), and 18:4n-3 that showed higher values at the inner station (Table 2). When compared to bivalve DG, the PUFA 18:4n-3 and 20:5n-3 in POM were similar along the sampled transect (Table 2), and the 18:1n-9 content was lower at stations 2 and 3 (3% and 2.8%, respectively) compared to station 1 (5.2%) (Table 2).
Compound-specific isotopic analyses
The δ13C values of almost all selected FA of bivalve DG differed significantly among stations (ANOVA or KW tests followed by post hoc; Table 1), with station 3 being different from stations 1 and 2, except for 20:4n-6, 20:5n-3 and 18:1n-7 (Table 1, Fig. 3). The differences among stations were significant for most FA; however, the range of variation did not exceed 1‰ for some FA, such as 16:1n-7 (from − 27‰ at the external station, to − 25.9‰ at the inner station; Table 1), and 20:5n-3 at station 2 (− 26.9‰) differed significantly from station 3 (− 27.6‰; Table 1) (Fig. 3). The variability of δ13C values of 18:1n-9 + 18:3n-3 was 1.9‰ (Fig. 3), ranging from − 29.7‰ (station 1) to − 29.1‰ (station 2), and finally to − 31.0‰ (station 3; Table 1). Because these FA coeluted on the apolar column used for GC-C-IRMS analysis, they were reported jointly. The δ13C values of 18:4n-3 displayed the highest range of variability (2.3‰) (Fig. 3), ranging from − 29.2‰ at stations 1 and 2 to − 31.5‰ at station 3 (Table 1). Finally, 22:6n-3 varied in its δ13C values among the stations (1.8‰; Fig. 3) with higher δ13C values at the external stations 1 and 2 (− 26.1‰ and − 26.9‰, respectively) compared to lower δ13C values at the inner station 3 (− 27.5‰) (Table 1). For C18 FA (18:1n-9, 18:3n-3, 18:4n-3) and 22:6n-3 of bivalve DG, samples from station 3 were always more depleted in 13C compared to other stations (Table 1 and Fig. 3).

Box plot of δ13C values (‰; n = 5) of some selected neutral lipid fatty acid (FA) of Spondylus crassisquama digestive gland in June 2017 at each stations (1, 2, and 3). Only the FA accounting for more than 80% of dissimilarity between stations, presented the greatest abundance permitting their good detection in CSIA, and present in all samples are shown. Different letters indicate significant difference between the stations (ANOVA or KW tests followed by multiple comparison tests, at the significant level α < 0.05). Samples from station 1 are shown in dark blue, 2 in light blue, and 3 in green
Similar trends in isotopic variability of FA were found in POM samples than in bivalve DG, with 18C and 22:6n-3 always more depleted in 13C with increasing distance from the mouth of the lagoon (Table 2). For example, the δ13C values of 18:4n-3 were similar at station 1 (− 31.6‰) and 2 (− 31.4‰) and more depleted at station 3 (− 32.3‰; Table 2), and the 22:6n-3 showed δ13C depleted values along the transect (from − 25.6 and − 27.5‰ at stations 1 and 2 to − 34.9‰ at station 3; Table 2). Like in DG samples, both FA, 16:1n-7 and 20:5n-3, were 13C enriched at the internal station (Table 2) in POM samples. Indeed, 16:1n-7 and 20:5n-3 showed, respectively, δ13C from − 26.7‰ and − 29.5‰ at the external station, to − 25.8‰ and − 28.7‰ at the station near to the mouth, and to − 25.7‰ and − 27.5‰ at the inner station (Table 2).
Discussion
Results from this study suggest that organic matter from the ocean is transported into the lagoon, contributing to the primary production and to the diet of benthic filter feeders. The CSIA on FA data indicate that diatoms (indicated by 16:1n-7 and 20:5n-3) assimilated by bivalves have a very similar marine origin throughout the lagoon, according to their comparable δ13C values among stations. Moreover, results also demonstrate that a diversity of heterotrophic flagellates, characterized by higher contents of 18 carbon FA, and 22:6n-3 may also enter the diet of bivalves. The variable δ13C values of these FA among stations suggest that their origin varies according to the location within the lagoon, and depleted values at the inner station indicate an influence of local processes in the diet of these consumers.
The most striking result from bulk SI analysis of bivalve muscles and POM was the significant 15N enrichment observed near the entrance of the lagoon, with individuals from stations 1 and 2 displaying similar δ15N values that were higher than in the inner bay. This reveals an oceanic influence, and particularly the contribution of 15N oceanic inputs to the diet of bivalves near the entrance of the lagoon, but also that such dietary influence of the ocean did not extend to the innermost part of the lagoon. This oceanic influence may be explained by active wind-driven seasonal upwelling occurring outside the lagoon from April to June (Ibarra-Obando et al. 2001; Du et al. 2015). It has been suggested that water masses containing upwelled biological material could be transported into coastal lagoons of the Pacific coast of Baja California (Zaytsev 2003), promoting primary production and potentially affecting their trophic functioning. The hypothesis that upwelled waters affect the diet of bivalves inside the lagoon is based on the fact that during upwelling conditions, the biomass of large phytoplankton increases (Taylor and Landry 2018), and as upwelled nitrate is rapidly consumed by phytoplankton (O’Reilly et al. 2002), its δ15N increases. Although coastal upwelling influence on the studied lagoon was not supported by chlorophyll a or temperature data, such high δ15N with no changes in δ13C have been reported in various systems under upwelling influence worldwide (O’Reilly et al. 2002; Hill and McQuaid 2008; Reddin et al. 2015).
The spatial differences found in the FA composition of neutral lipids in the DG of bivalves indicate a change in the proportion and possibly in the nature of food sources with increasing distance from the mouth of the lagoon. The DG at the entrance of the lagoon (stations 1 and 2) had higher 16:1n-7 and 20:5n-3 contents when compared to individuals from the inner station (station 3), indicating a higher contribution of diatoms in their diet (Parrish et al. 2000; Alfaro et al. 2006). A previous study revealed that during the late spring, diatoms are the dominant phytoplankton taxa found in the Northern California current (Du et al. 2015). The higher importance of diatoms in the diet of bivalves near the entrance of the lagoon, therefore, supports the influence of oceanic primary production. Individuals from the inner station had lower contents of these two FA, suggesting a dilution of oceanic inputs further into the lagoon. The importance of 22:6n-3 in DG of individuals at the external station suggests that dinoflagellates also represent an important dietary contribution of Spondylus crassisquama (Parrish et al. 2000). This contribution decreases slightly with increasing distance from the mouth of the lagoon, being the lowest at the internal station, suggesting a decreasing contribution of dinoflagellates to this station. However, the high abundance of 18:4n-3, another dinoflagellate marker (Kharlamenko et al. 2001; Bachok et al. 2003; Nerot et al. 2015), at the inner station (station 3) suggests the opposite. Such conflicting information by these two dinoflagellate markers may indicate different assemblages being assimilated by bivalves in the lagoon, but such change in dietary biomarkers does not necessarily suggest any shift in contribution of dinoflagellates to the diets of these consumers. Bivalves from the inner bay were also characterized by higher C18-FA contents (18:1n-9, 18:3n-3). These FA can be characteristic of different dietary sources, such as seagrass (Kharlamenko et al. 2001) or heterotrophic flagellates constituting microbial food webs (Zhukova 1991). Although seagrass beds are known to be a source of dissolved organic matter (DOM; Fenton and Ritz 1988), bivalve muscles were too 13C depleted as compared to eelgrass δ13C to be in accordance with a significant trophic contribution of Zostera marina. Consequently, the abundance of these FA likely reflects the contribution of heterotrophic microorganisms (i.e. bacteria, ciliates, flagellates, dinoflagellates), relying on locally produced DOM, to the diet of bivalves. This highlights a higher diversity of food sources for bivalves at the internal station than for those submitted to oceanic influences and shows that diet source variation within this and possibly other lagoons is based on allochthonous and autochthonous contribution. Such various diet sources could also present differences in terms of quality and quantity, which consequently impact bivalve stocks and consequently fisheries. More detailed understanding of spatio-temporal dynamics of these food resources would lead to a better management of bivalve stocks in the Ojo de Liebre lagoon and elsewhere.
Inferring diet composition based on CSIA requires FA-specific fractionation coefficients, which are not available for our species, and would require extensive laboratory experiments (Ruess and Chamberlain 2010; Bec et al. 2011). Consequently, our approach did not aim to characterize the accuracy of food sources retention by bivalves, but to scrutinize the trophic spatial variability in an effort to provide evidence of a different spatial origin of the diets for this consumer (Ramos et al. 2003; Van den Meersche et al. 2004). Although no difference in bulk δ13C values was observed in muscles of individuals from the different stations, CSIA detect the isotopic composition of FA that may indicate source-specific differences of FA (Budge et al. 2008; Colombo et al. 2016). Some differences were observed in the range of variation in FA δ13C along the transect depending on biomarkers considered (diatoms vs heterotrophic flagellates). In DG neutral lipids, diatom biomarkers (16:1n-7 and 20:5n-3) displayed the smallest range of variation among the three stations (≤ 1‰), which is of little ecological value. The spatial consistency in the δ13C values of these diatom biomarkers suggests that diatoms assimilated by bivalves have all oceanic origin throughout the bay. Therefore, it seems that phytoplankton of local origin (produced within the lagoon) is of relatively little dietary importance for S. crassisquama. The C18-FA 18:1n-9 and 18:3n-3 were both considered to reveal the contribution of microheterotrophs (flagellates, ciliates, see above). Even if the three stations presented significant differences in their δ13C for 18:1n-9 and 18:3n-3, stations 1 and 2 displayed very similar δ13C values (only differed by 0.6‰), while station 3 was depleted in 13C by 1.9‰ compared to the other stations. This difference suggests that the origin of these microheterotrophs differed within the lagoon. Microheterotrophs are central elements of microbial food webs, feeding on bacteria and other small unicellular organisms, and ultimately relying on DOM (Azam et al. 1983). The 13C depletion observed at the inner station suggests a local source of DOM, which is of primary importance for bivalves in the lagoon. Although the lagoon of Ojo de Liebre hosts one of the largest eelgrass bed of the Northern Pacific (Cabello-Pasini et al. 2003), a seagrass-derived DOM seems unlikely because seagrass is usually 13C enriched compared to other marine primary producers (Hemminga and Mateo 1996), which was also observed in the present study. The possible DOM release from the sediment is also not in agreement with δ13C measured for these FA (around − 24.5‰ in SOM). In fact, such 13C-depleted values in coastal environments are usually associated with red algae, which can be extremely low in δ13C (Raven et al. 2002). Although no extensive population of such algae was observed during sampling, it is possible that such populations exist in the inner bay, supplying a pool of dissolved organic carbon that is subsequently trophically transferred to bivalves.
Dinoflagellate FA biomarkers (18:4n-3 and 22:6n-3) had a very similar spatial distribution pattern as observed for biomarkers of other microheterotrophs (18:1n-9 and 18:3n-3), with a marked 13C depletion at the inner station. Therefore, in contrast to diatoms, dinoflagellates entering the diet of bivalves do not seem to be of marine origin throughout the lagoon. The similarity observed between the CSIA in microheterotrophs and dinoflagellates might suggest that the same factors affect the SI composition of these two possible food sources. In fact, although dinoflagellates are usually described as primary producers, heterotrophy is a widespread pattern in this group (Jeong et al. 2010). Based on these results, it is possible that dinoflagellates assimilated by S. crassisquama in the inner part of the lagoon may rely on heterotrophy, emphasizing a potentially important role of the microbial food web in this enclosed ecosystem.
Conclusion
The use of CSIA provided a new scale of complexity in the diet of Spondylus crassisquama in the Ojo de Liebre lagoon. Although dietary diatoms retained in bivalves appeared to have the same oceanic origin throughout the lagoon, this study indicates that the heterotrophic component of bivalve diet varies spatially in the lagoon, highlighting an influence of local processes in the diet of these consumers. The combination of the three methods used here showed to be complementary, resulting in a more informative analysis of spatial feeding ecology of this and likely also other lagoons. The lower δ13C observed at the most inner station could not be explained in this study, although any potential role of eelgrass in providing DOM for the associated microbial food web could be discarded based on different SI data. Further research is warranted to identify how the origin of basal resources varies and supports food webs in lagoons and to better understand the quantitative importance of dietary sources for bivalves.
References
Abrajano TA, Murphy DE, Fang J, Comet P, Brooks JM (1994) 13C12c ratios in individual fatty acids of marine mytilids with and without bacterial symbionts. Org Geochem 21:611–617. https://doi.org/10.1016/0146-6380(94)90007-8
Alfaro AC, Thomas F, Sergent L, Duxbury M (2006) Identification of trophic interactions within an estuarine food web (northern New Zealand) using fatty acid biomarkers and stable isotopes. Estuar Coast Shelf Sci 70:271–286. https://doi.org/10.1016/j.ecss.2006.06.017
Allan EL, Ambrose ST, Richoux NB, Froneman PW (2010) Determining spatial changes in the diet of nearshore suspension-feeders along the South African coastline: stable isotope and fatty acid signatures. Estuar Coast Shelf Sci 87:463–471. https://doi.org/10.1016/j.ecss.2010.02.004
Azam F, Fenchel T, Field JG, Gray JS, Meyer-Reil LA, Thingstad F (1983) The ecological role of water-column microbes in the sea. Mar Ecol Prog Ser 10:257–263
Bachelet G, De Montaudouin X, Auby I, Labourg P-J (2000) Seasonal changes in macrophyte and macrozoobenthos assemblages in three coastal lagoons under varying degrees of eutrophication. ICES J Mar Sci 57:1495–1506
Bachok Z, Mfilinge PL, Tsuchiya M (2003) The diet of the mud clam Geloina coaxans (Mollusca, Bivalvia) as indicated by fatty acid markers in a subtropical mangrove forest of Okinawa, Japan. J Exp Mar Biol Ecol 292:187–197. https://doi.org/10.1016/S0022-0981(03)00160-6
Bec A, Perga M-E, Koussoroplis A, Bardoux G, Desvilettes C, Bourdier G, Mariotti A (2011) Assessing the reliability of fatty acid–specific stable isotope analysis for trophic studies. Methods Ecol Evol 2:651–659. https://doi.org/10.1111/j.2041-210X.2011.00111.x
Budge SM, Iverson SJ, Koopman HN (2006) Studying trophic ecology in marine ecosystems using fatty acids: a primer on analysis and interpretation. Mar Mammal Sci 22:759–801. https://doi.org/10.1111/j.1748-7692.2006.00079.x
Budge SM, Wooller MJ, Springer AM, Iverson SJ, McRoy CP, Divoky GJ (2008) Tracing carbon flow in an arctic marine food web using fatty acid-stable isotope analysis. Oecologia 157:117–129
Cabello-Pasini A, Muñiz-Salazar R, Ward DH (2003) Annual variations of biomass and photosynthesis in Zostera marina at its southern end of distribution in the North Pacific. Aquat Bot 76:31–47. https://doi.org/10.1016/S0304-3770(03)00012-3
Carlier A, Riera P, Amouroux J-M, Bodiou J-Y, Desmalades M, Grémare A (2008) Food web structure of two Mediterranean lagoons under varying degree of eutrophication. J Sea Res 60:264–275. https://doi.org/10.1016/j.seares.2008.10.006
Clavier J, Chauvaud L, Amice E, Lazure P, Van Der Geest M, Labrosse P, Diagne A, Carlier A, Chauvaud S (2014) Benthic metabolism in shallow coastal ecosystems of the Banc d’Arguin, Mauritania. Mar Ecol Prog Ser 501:11–23
Colombo SM, Parrish CC, Whiticar MJ (2016) Fatty acid stable isotope signatures of molluscs exposed to finfish farming outputs. Aquac Environ Interact 8:611–617
Core Team R (2017) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org/
Dalsgaard J, St. John M, Kattner G, Müller-Navarra D, Hagen W (2003) Fatty acid trophic markers in the pelagic marine environment. Adv Mar Biol 46:225–340. https://doi.org/10.1016/S0065-2881(03)46005-7
Du X, Peterson W, O’Higgins L (2015) Interannual variations in phytoplankton community structure in the northern California Current during the upwelling seasons of 2001–2010. Mar Ecol Prog Ser 519:75–87
Falkowski PG (1991) Species variability in the fractionation of 13C and 12C by marine phytoplankton. J Plankton Res 13:21–28. https://doi.org/10.1093/oxfordjournals.plankt.a042367
Fenton GE, Ritz DA (1988) Changes in carbon and hydrogen stable isotope ratios of macroalgae and seagrass during decomposition. Estuar Coast Shelf Sci 26:429–436
Folch J, Lees M, Sloane-Stanley GH (1957) A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem 226:497–509
France RL (1995) Carbon-13 enrichment in benthic compared to planktonic algae: foodweb implications. Mar Ecol Prog Ser 124:307–312
Fry B (1988) Food web structure on Georges Bank from stable C, N, and S isotopic compositions. Limnol Oceanogr 33:1182–1190. https://doi.org/10.4319/lo.1988.33.5.1182
Gaillard B, Meziane T, Tremblay R, Archambault P, Blicher ME, Chauvaud L, Rysgaard S, Olivier F (2017) Food resources of the bivalve Astarte elliptica in a sub-Arctic fjord: a multi-biomarker approach. Mar Ecol Prog Ser 567:139–156
Graham C, Oxtoby L, Wang SW, Budge SM, Wooller MJ (2014) Sourcing fatty acids to juvenile polar cod (Boreogadus saida) in the Beaufort Sea using compound-specific stable carbon isotope analyses. Polar Biol 37:697–705
Hemminga MA, Mateo MA (1996) Stable carbon isotopes in seagrasses: variability in ratios and use in ecological studies. Mar Ecol Prog Ser 140:285–298
Hill JM, McQuaid CD (2008) δ13C and δ15N biogeographic trends in rocky intertidal communities along the coast of South Africa: evidence of strong environmental signatures. Estuar Coast Shelf Sci 80:261–268. https://doi.org/10.1016/j.ecss.2008.08.005
Ibarra-Obando SE, Camacho-Ibar VF, Carriquiry JD, Smith SV (2001) Upwelling and lagoonal ecosystems of the dry Pacific coast of Baja California. In: Seeliger U, Kjerfve B (eds) Coastal marine ecosystems of Latin America. Ecological studies (analysis and synthesis), vol 144. Springer, Berlin, Heidelberg
Jaschinski S, Brepohl DC, Sommer U (2008) Carbon sources and trophic structure in an eelgrass Zostera marina bed, based on stable isotope and fatty acid analyses. Mar Ecol Prog Ser 358:103–114
Jeong HJ, Du Yoo Y, Kim JS, Seong KA, Kang NS, Kim TH (2010) Growth, feeding and ecological roles of the mixotrophic and heterotrophic dinoflagellates in marine planktonic food webs. Ocean Sci J 45:65–91
Kainz M, Lucotte M, Parrish CC (2002) Methyl mercury in zooplankton the role of size, habitat, and food quality. Can J Fish Aquat Sci 59:1606–1615
Kelly JR, Scheibling RE (2012) Fatty acids as dietary tracers in benthic food webs. Mar Ecol Prog Ser 446:1–22
Kharlamenko VI, Kiyashko SI, Imbs AB, Vyshkvartzev DI (2001) Identification of food sources of invertebrates from the seagrass Zostera marina community using carbon and sulfur stable isotope ratio and fatty acid analyses. Mar Ecol Prog Ser 220:103–117
Layman CA, Araujo MS, Boucek R, Hammerschlag-Peyer CM, Harrison E, Jud ZR, Matich P, Rosenblatt AE, Vaudo JJ, Yeager LA (2012) Applying stable isotopes to examine food-web structure: an overview of analytical tools. Biol Rev 87:545–562
Le Grand F, Soudant P, Siah A, Tremblay R, Marty Y, Kraffe E (2014) Disseminated Neoplasia in the soft-shell Clam Mya arenaria: membrane lipid composition and functional parameters of circulating cells. Lipids 49:807–818. https://doi.org/10.1007/s11745-014-3917-4
Lorrain A, Paulet Y-M, Chauvaud L, Savoye N, Donval A, Saout C (2002) Differential δ13C and δ15N signatures among scallop tissues: implications for ecology and physiology. J Exp Mar Biol Ecol 275:47–61. https://doi.org/10.1016/S0022-0981(02)00220-4
Marchais V, Schaal G, Grall J, Lorrain A, Nerot C, Richard P, Chauvaud L (2013) Spatial variability of stable isotope Ratios in Oysters (Crassostrea gigas) and primary producers along an estuarine gradient (Bay of Brest, France). Estuaries Coasts 36:808–819. https://doi.org/10.1007/s12237-012-9584-x
Nerot C, Meziane T, Schaal G, Grall J, Lorrain A, Paulet Y-M, Kraffe E (2015) Spatial changes in fatty acids signatures of the great scallop Pecten maximus across the Bay of Biscay continental shelf. Cont Shelf Res 109:1–9. https://doi.org/10.1016/j.csr.2015.08.032
Nixon SW (1982) Nutrient dynamics, primary production and fisheries yields of lagoons. In: Proceedings of the international symposium on coastal lagoons SCORI lABO/UNESCO, Bordeaux, 8–14 September 1981, pp 357–371
O’Reilly CM, Hecky RE, Cohen AS, Plisnier P-D (2002) Interpreting stable isotopes in food webs: recognizing the role of time averaging at different trophic levels. Limnol Oceanogr 47:306–309. https://doi.org/10.4319/lo.2002.47.1.0306
Parrish CC, Abrajano TA, Budge SM, Helleur RJ, Hudson ED, Pulchan K, Ramos C (2000) Lipid and phenolic biomarkers in marine ecosystems: analysis and applications. In: Wangersky PJ (eds) Marine chemistry. The handbook of environmental chemistry (Vol. 5 Series: Water pollution), vol 5D. Springer, Berlin, Heidelberg
Post DM, Layman CA, Arrington DA, Takimoto G, Quattrochi J, Montaña CG (2007) Getting to the fat of the matter: models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia 152:179–189. https://doi.org/10.1007/s00442-006-0630-x
Ramos CS, Parrish CC, Quibuyen TAO, Abrajano TA (2003) Molecular and carbon isotopic variations in lipids in rapidly settling particles during a spring phytoplankton bloom. Org Geochem 34:195–207
Raven JA, Johnston AM, Kübler JE, Korb R, McInroy SG, Handley LL, Scrimgeour CM, Walker DI, Beardall J, Vanderklift M, Fredriksen S, Dunton KH (2002) Mechanistic interpretation of carbon isotope discrimination by marine macroalgae and seagrasses. Funct Plant Biol 29:355–378. https://doi.org/10.1071/pp01201
Reddin CJ, Docmac F, O’Connor NE, Bothwell JH, Harrod C (2015) Coastal upwelling drives intertidal assemblage structure and trophic ecology. PLoS One 10:e0130789
Riera P (2007) Trophic subsidies of Crassostrea gigas, Mytilus edulis and Crepidula fornicata in the Bay of Mont Saint Michel (France): a δ13C and δ15N investigation. Estuar Coast Shelf Sci 72:33–41. https://doi.org/10.1016/j.ecss.2006.10.002
Riera P, Richard P (1996) Isotopic determination of food sources of Crassostrea gigas along a trophic gradient in the estuarine bay of Marennes-Oléron. Estuar Coast Shelf Sci 42:347–360
Ruess L, Chamberlain PM (2010) The fat that matters: soil food web analysis using fatty acids and their carbon stable isotope signature. Soil Biol Biochem 42:1898–1910
Schaal G, Nerot C, Grall J, Chouvelon T, Lorrain A, Mortillaro J-M, Savoye N, Brind’Amour A, Paulet Y-M, Le Bris H (2016) Stable isotope ratios in bentho-demersal biota along a depth gradient in the Bay of Biscay: a multitrophic study. Estuar Coast Shelf Sci 179:201–206. https://doi.org/10.1016/j.ecss.2015.10.023
Taylor AG, Landry MR (2018) Phytoplankton biomass and size structure across trophic gradients in the southern California Current and adjacent ocean ecosystems. Mar Ecol Prog Ser 592:1–17
Van den Meersche K, Middelburg JJ, Soetaert K, Van Rijswijk P, Boschker HT, Heip CH (2004) Carbon-nitrogen coupling and algal-bacterial interactions during an experimental bloom: modeling a 13C tracer experiment. Limnol Oceanogr 49:862–878
Vuorio K, Meili M, Sarvala J (2006) Taxon-specific variation in the stable isotopic signatures (δ13C and δ15N) of lake phytoplankton. Freshw Biol 51:807–822
Zaytsev O (2003) Coastal upwelling activity on the Pacific shelf of the Baja California peninsula. J Oceanogr 59:489–502
Zhukova NV (1991) The pathway of the biosynthesis of non-methylene-interrupted dienoic fatty acids in molluscs. Comp Biochem Physiol Part B Comp Biochem 100:801–804
Acknowledgements
Authors would like to thank the contribution of the Guerrero Negro unit of CIBNOR for hosting the first steps of sample preparation after fieldwork, the LIPIDOCEAN analytical facilities for hosting all fatty acid analyses, and the Pôle Spectrométrie Océan (Clément Tanvet) for running bulk SI and CSIA analyses. Authors thank the fishermen crew for the fieldwork, but also Laura Guzman for their precious help during sampling and dissections, and Dr. Elena Palacios and Dr. Kitty Arredondo Vega, for the laboratory facilities provided for the preparation of samples at the CIBNOR La Paz. Authors address their thanks to the Reserva de la biosfera desierto de “el Vizcaino” for their help and support in carrying out sampling for this study. Authors would like to thank Aurelien Boye for his precious help and advices about statistical analyses. Authors also thank Pr. Sandra Shumway and an anonymous reviewer, for their help in improving this paper.
Funding
This study was supported by the ECOS-ANUIES program (PROPHYMUS project), the “Laboratoire d’Excellence” LabexMER (ANR-10-LABX-19) and co-funded by a grant from the French government under the program “Investissements d’Avenir”. MMR’s PhD fellowship was provided by the French Research Ministry and Region Bretagne.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
All applicable international, national and/or institutional guidelines for the care and use of bivalves Spondylus crassisquama were followed. The individuals of S. crassisquama have been sampled under the permit PRMN/DGOPA-011/2017, delivered by the Mexican government.
Additional information
Responsible Editor: S. Shumway.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Reviewed by Undisclosed experts.
Rights and permissions
About this article

Cite this article
Mathieu-Resuge, M., Schaal, G., Kraffe, E. et al. Different particle sources in a bivalve species of a coastal lagoon: evidence from stable isotopes, fatty acids, and compound-specific stable isotopes. Mar Biol 166, 89 (2019). https://doi.org/10.1007/s00227-019-3535-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-019-3535-z