
Abstract
The mating systems of symbiotic crustaceans vary broadly, ranging from monogamy to polygamy to promiscuity. This disparity in mating systems coupled with wide differences in host ecology provides opportunities to study the effect of environmental conditions on the social behavior of marine organisms. In this study, we tested the prediction that symbiotic crustaceans inhabiting relatively large and abundant host species are promiscuous rather than monogamous. The host traits above constraint host monopolization and favor host-switching behaviors by males individuals and, thus, ultimately drive promiscuity as a mating system in symbiotic crustaceans. As a model system we used Dissodactylus crinitichelis, a small ‘pea-crab’ that inhabits the relatively large and abundant sand dollar Encope emarginata. We described the population distribution of D. crinitichelis at two intertidal localities in Sergipe, Brazil, and conducted laboratory experiments to infer the mating system of this species. Dissodactylus crinitichelis inhabit sand dollars as adult heterosexual pairs more frequently than expected by chance alone, but these heterosexual pairs did not exhibit size-assortative pairing. Sexual dimorphism was reversed in D. crinitichelis, with males attaining smaller average body sizes than females but exhibiting more elaborated weaponry (male > female claws). These findings disagree with expectations for monogamous symbiotic crustaceans. Laboratory experiments demonstrated that male and female crabs switch among host individuals rather frequently and heterosexual pairs retrieved from the field did not remain together for long times. Our results argue against social monogamy and in favor of promiscuity in D. crinitichelis. Our study supports the notion that host abundance and large body size (relative to that of symbiotic guests) favor promiscuity in symbiotic crustaceans.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The adoption of a symbiotic lifestyle is a major environmental adaptation in marine organisms (Baeza 2015). Symbiotic partnerships are typically composed of small organisms (hereafter termed symbiotic guests or symbionts) that reside in or on much larger invertebrates and vertebrates that act as hosts, including sea anemones, ascidians, corals, sponges, sea urchins, oysters, fishes, reptiles (sea turtles), and whales, among others (Carranza et al. 2003; Nogata and Matsumura 2005; Campos et al. 2009; Wirtz et al. 2009). Host species vary considerably in their biology and ecology, and a wide diversity of host-use patterns and mating systems have been described for symbiotic guest species (Baeza and Thiel 2007). For instance, among crustaceans, a species-rich group of marine invertebrates (Martin and Davis 2006), some symbiotic guest species dwell in or on their hosts as solitary individuals (Baeza and Stotz 2003) or as monogamous pairs (Baeza 2008). Other symbiotic guest crustaceans form large aggregations with no evident demographic structure (Baeza and Piantoni 2010), and a few establish well-structured groups within their hosts (Duffy 1996, 2007). This disparity in mating systems and host ecology explains why symbiotic crustaceans are currently being used as a model system to study the effects of environmental conditions on the social behavior of marine organisms (De Bruyn et al. 2009; Ocampo et al. 2012; Ory and Dudgeon 2013; Jossart et al. 2014; Ambrosio and Baeza 2016; Baeza et al. 2015, 2016).
Baeza and Thiel (2007) proposed a model that predicts the mating system of symbiotic guest crustaceans. This model considers that off-host predation risk together with host size (relative to that of the symbiotic guest), abundance, and morphological complexity drive host guarding and host switching in symbiotic crustaceans. These two behaviors (guarding and switching of hosts) determine the rate and number of interactions among potential competitors and mates, ultimately driving the mating system of symbiotic crustaceans.
Specifically, Baeza and Thiel’s (2007) model predicts that monogamy is adaptive in symbiotic crustaceans inhabiting morphologically simple hosts that attain small body sizes (relative to that of the symbiotic guest) in tropical environments where predation risk away from hosts is high. Under the conditions above, movement among hosts is constrained and monopolization of hosts is favored in males and females due to host scarcity and its value in offering protection. Thus, males and females optimize their reproductive success by sharing ‘their’ host individual with a member of the opposite sex for long time periods. Under monogamy, minimal sexual dimorphism in body size and weaponry should exist in symbiotic crustaceans due to the low intensity of sexual selection in this mating system. In turn, promiscuity is adaptive in symbiotic crustaceans inhabiting relatively large hosts that are naturally abundant, morphologically complex and living in environments where off-host predation risk is low. Under these circumstances, both males and females are expected to move frequently among host individuals. Host morphological complexity and large body size render host monopolization ineffective by symbiotic crustaceans (Baeza and Thiel 2003, 2007). Under such conditions, promiscuity is favored, as male host-switching behavior is not constrained; as soon as receptive females are found, males should mate and abandon them immediately to continue searching for other receptive females (Baeza and Thiel 2007). Baeza and Thiel (2007) also suggested that sexual size dimorphism should be reverse (males < females) in promiscuous symbiotic crustaceans. Small body size in males is suggested to increase male agility and, thus, encounter rate with potential mating partners (Baeza and Thiel 2007 and references therein).
In this study, we test the hypothesis stating that symbiotic organisms living in association with relatively large and abundant host species in environments in which off-host predation risk is low are not monogamous but rather promiscuous (Baeza and Thiel 2007). We used the pea-crab Dissodactylus crinitichelis and its sand dollar host Encope emarginata as a model system. The sand dollar E. emarginata, one of the echinoids inhabited by D. crinitichelis, is found in soft-bottom estuarine regions, from the intertidal zone to a depth of ~50 m (Gray et al. 1968; Pérez et al. 2012). Dissodactylus crinitichelis obtains food and protection against predators from this sand dollar (Reeves and Brooks 2001, and references therein). Encope emarginata is one of the most abundant species in soft-bottom benthic communities (Pires-Vanin 2001), exhibits a clumped distribution, and can reach densities as high as 5 individuals m−2 (Guilherme et al. 2015). Although no studies have quantified predation intensity in shallow soft-bottom environments inhabited by sand dollars and D. crinitichelis, it is likely that off-host predation risk for individuals is low for this pea crab. Indeed, D. crinitichelis is an agile and fast-burrowing species that quickly hides among sand grains and/or mud when off-host individuals (DF, personal observation), a behavior known to decrease predation rate, as reported before for burrowing shrimps (Kathiresan and Bingham 2001). Overall, the information above suggests that the environment does not constrain, at least not intensively, movements of this symbiotic crab among different host individuals (Baeza and Thiel 2003, 2007, see also De Bruyn et al. 2016). Additionally, the sand dollar E. emarginata attains 140 mm or more in test length (Pérez et al. 2012). In contrast, the symbiotic crab D. crinitichelis does not reach body sizes larger than 9.0 mm carapace width (Williams 1984). Thus, the sand dollar host of D. crinitichelis further represents a large refuge that most likely is very difficult (i.e., expensive in terms of time and energy) to monopolize by symbiotic crabs (Baeza and Thiel 2007). In general, the pea-crab D. crinitichelis should not be monogamous but rather promiscuous, with males (and maybe females) switching among host individuals in search of reproductive partners. In agreement with the notion above, limited information indicates that crabs are often found living in groups on the surface of sand dollars and heterosexual pairs are uncommon (Martinelli-Filho et al. 2014; Guilherme et al. 2015). Nonetheless, detailed studies on the sociobiology of this symbiotic species are still lacking.
If D. crinitichelis is not socially monogamous but rather promiscuous, then we should expect that (1) the population distribution of this crab on Encope emarginata will be random, (2) the sex distribution of crabs in pairs will be random with male–female pairs occurring less often than expected by chance alone or not occurring at all, (3) heterosexual pairs (if any) will not exhibit size-assortative pairing, (4) sexual dimorphism in body size will be observed, and (5) crabs will exhibit frequent movements between host individuals and heterosexual pairing will not be stable over time. If these predictions (1–5) are confirmed, this would strongly indicate that D. crinitichelis crabs do not share host individuals with a member of the opposite sex for long time periods (as would be expected to occur in monogamous species). Instead, this pattern (e.g., random distribution of heterosexual pairs. no size-assortative pairing) would imply that the species is promiscuous, with, e.g., males actively abandoning a host individual shortly after reproducing with a female (Adams et al. 1985; Baeza 1999; Baeza and Thiel 2007). Here, we describe the host-use pattern of D. crinitichelis, including population distribution, male–female association pattern, and sexual dimorphism. Laboratory experiments were conducted to reveal the mating system adopted by this pea-crab.
Materials and methods
Study area and collection of sand dollars and crabs
Sampling was conducted during September 2014 at Vaza-Barris, a large estuarine system in the State of Sergipe, northeastern Brazil. During low tide periods, sandbanks are exposed, revealing large intertidal areas (Fig. 1a). Specimens of Dissodactylus crinitichelis Moreira, 1901 (Fig. 1b) were retrieved from the surface of the sand dollar Encope emarginata (Leske, 1778), a common inhabitant of sandbanks. Encope emarginata lives from the lower intertidal zone up to the shallow sublittoral (<0.5 m depth) (Fig. 1c). Crabs and sand dollars were sampled on two sand banks, Croa do Goré (11°05′47″S–37°09′30″W) (CG), located approximately 5 km from the mouth of the estuary, and Ilha dos Namorados (11°09′07″S–37°09′36″W) (IN), located at the mouth of the estuary. We recorded water temperature with a mercury thermometer (INCOTERM, accuracy = 0.5 °C) and salinity with a hand-held refractometer (BIOBRIX, precision = 0.5) at each study site. Temperature (T) and salinity (S) were slightly greater in IN (T = 23 °C, S = 40) than in CG (T = 21 °C, S = 35).

a The sandbanks at Vaza-Barris estuary, Brazil, during low tide periods; b dorsal view of Dissodactylus crinitichelis; c Encope emarginata buried in sediment in the upper region of midlittoral; d Dissodactylus crinitichelis in symbiosis with Encope emarginata, a crab in the oral region and a crab in the lunule
Host abundance and colonization intensity by symbiotic crabs were determined, at each study site, by haphazardly distributing ten 4 m2 quadrats (2 × 2 m) in the lower intertidal zone and shallow sublittoral (<0.3 m depth) parallel to the shoreline. Approximately, five meters separated each quadrat from another. We estimated host density (number of sand dollars m−2) by counting in each quadrat the totality of sand dollars observed. In addition, a total of 100 sand dollars was gently collected with a small shovel at each study site, placed in a sieve (500 µm mesh size), and immediately rinsed with seawater. After sand dollars were rinsed, the total number of specimens of D. crinitichelis present on each sand dollar was recorded (Fig. 1d). All crabs collected from each sand dollar were placed individually and together with their sand dollar into plastic bags filled with seawater (50 ml). Crabs and ‘their’ sand dollars were transported live in thermal boxes to the laboratory for further analysis.
Sizing and sexing in Dissodactylus crinitichelis
In the laboratory, crabs were preserved in 70% ethanol. The maximum diameter (MD) of each sand dollar was measured with a digital caliper to the nearest 0.01 mm. The carapace width (CW) of each crab was measured under the stereomicroscope (Leica M205 C) using the imaging software Leica Application Suite (LAS) version 4.4. Crabs were sexed according to the gross shape of the abdomen (narrow in males and wide in females) and the number of pleopods (two pairs in males and four pairs in females). Also, the developmental stage (juvenile or adult) of each crab was determined depending on a particular combination of abdominal and pleopod shape. Crabs with a wide abdomen, nearly covering the entire ventral surface of the cephalothorax, and with setose pleopods were classified as adult females. Crabs with a narrow abdomen and two pairs of short gonopods were classified as adult males. Crabs with a triangular abdominal shape and with two and four pairs of non-setose pleopods were classified as juvenile males and females, respectively (see Haefner 1990). For a few of the smallest individuals in the population, sex was not determined and these are treated here as individuals of undetermined sex. Lastly, the presence or absence of eggs carried underneath the abdomen in mature females was recorded.
Population distribution in Dissodactylus crinitichelis
We examined the host use pattern of D. crinitichelis, which includes a description of its population distribution, male–female association pattern, and host–crab body size relationships. First, we examined whether or not D. crinitichelis lives solitarily, in pairs, or gregariously on the surface of the sand dollar E. emarginata. For this purpose, we examined whether or not the distribution of D. crinitichelis on the sand dollar host (i.e., the frequency of occurrence of hosts without crabs and with different numbers of crabs) differed from a random distribution. We compared the observed distribution (i.e., frequency of occurrence of hosts with zero, one, two, three or more crabs) with the Poisson distribution (Elliot 1983). The Poisson distribution describes random occurrences when the probability of occurrence of an event (i.e., presence of a symbiotic guest individual on a host individual) is low (as in this study) (see Zar 2010 for details). The departure of an observed distribution from an expected random Poisson distribution indicates a uniform distribution of symbiotic guests on host individuals, i.e., the number of symbiotic guest per host individual is the same [often one] indicating territorial behavior in symbiotic species (Baeza and Thiel 2003; Ambrosio and Baeza 2016); or a gregarious distribution of symbiotic guests on host individuals, i.e., the number of two or more symbiotic guests per host individual is larger than expected by chance alone denoting gregarious behavior in symbiotic guest species (Baeza and Thiel 2003).
The analysis above was conducted for the entire crab population and only for adult individuals. If significant differences between expected Poisson and observed distribution were observed, specific frequencies between the observed and expected distributions were compared by subdivision of the Chi square test. Also, we used the sequential Bonferroni correction during a posteriori comparisons to correct for type I error given that we are conducting multiple statistical tests with the same dataset (Zar 2010).
A number of sand dollar hosts were found to harbor crab pairs (see results). To determine whether the sexes are randomly distributed among crab pairs inhabiting the same host individual, the observed distribution was compared with the binomial distribution. The expected random frequencies of distribution of the different sexes were calculated based on the proportion of males and females recorded in the population. A Chi square test of goodness of fit was used to inspect for significant differences between observed and expected distributions. This analysis was carried out for two datasets: (1) when pairs of adult crabs were the only guests, and (2) when pairs of adult crabs cohabited the same host, with and without non-reproductive crabs (juveniles and/or crabs of unknown sex). A Chi square test of goodness of fit was used to inspect for significant differences between observed and expected (binomial) distributions (Sokal and Rohlf 1981).
Sexual dimorphism in Dissodactylus crinitichelis
We tested whether or not D. crinitichelis exhibited sexual dimorphism in terms of body size. For this purpose, we compared the average CW of male and female crabs using a t test. Assumptions of normality and homogeneity of variances were evaluated using the Kolmogorov–Smirnov and Levene tests, respectively, and found to be satisfactory before conducting the t test.
To determine sexual dimorphism in terms of claw size, in a total of 50 males and 50 females of D. crinitichelis, we examined whether the claw on the first pair of pereopods increases linearly with body size. For this purpose, the relationship between the length of the claw and body size of crabs (CW) was examined using the allometric model y = ax b (Hartnoll 1978, 1982). The slope b of the log–log least-squares linear regression represents the rate of exponential increase (b > 1) or decrease (b < 1) of the claw with a unit of increase in body size of crabs. To determine if the relationship deviates from linearity, a t test was used to test if the estimated slope b deviates from the expected slope of unity (Sokal and Rohlf 1981). If the cheliped grows more or less than proportionally with a unit increase in body size of crabs, then the slope should be greater or smaller than the unity, respectively (Hartnoll 1978). Lastly, a covariance analysis (ANCOVA) was performed, to test the linear and angular coefficients between the sexes for the dependent variable.
Experimental test of host fidelity in Dissodactylus crinitichelis
Collection and acclimation of sand dollars and crabs
We conducted two experiments to examine crab host fidelity: crab movement among host individuals, and fidelity of paired crabs collected from the field. These experiments were designed to improve our knowledge about social interactions and the mating system of Dissodactylus crinitichelis.
For this purpose, E. emarginata sand dollars harboring one or more adult crabs were collected at depths ranging from 0.5 to 1.5 m from Croa do Goré during October, 2015. Crabs were carefully removed from ‘their’ sand dollar hosts and placed individually into plastic bags filled with seawater (50 ml). Sand dollars were placed separately into plastic bags filled with seawater (500 ml) and sand from the same sampling site. Crabs inhabiting sand dollars solitarily or in groups of three or more were used in experiment 1 while heterosexual pairs (n = 60) found sharing a single host individual were used in experiment 2. In the laboratory, females of these heterosexual pairs were examined under the stereomicroscope (Leica M205 C), in order to evaluate: (1) the proportion of ovigerous females, and (2) the developmental stage of embryos. Embryos carried by brooding females were classified according to three different categories (Wehrtmann 1990); stage I: rounded embryos with yolk uniformly distributed, no eye pigments visible, stage II: ovoid embryos, embryos with eye pigments elongated, stage III: ovoid embryos, with eyes well developed and the pleon free.
In the laboratory, crabs and sand dollars were measured to the nearest millimeter with a caliper (precision = 0.01 mm) and acclimated for 24 h in a plastic tank (103 × 84 × 57 cm, 250 L) with sand and seawater retrieved from the sampling site above. Crabs and sand dollars were maintained under constant temperature (26 ± 1 °C), salinity (35 ± 1) and photoperiod (12L:12D) similar to that found at the sampling site and seawater was constantly aerated during the experiments below using air pumps. The two experiments below were conducted under controlled laboratory conditions in a single circular tank (diameter = 45 cm, 10 L). The tanks contained a sediment layer 1 cm thick collected from the field and the water temperature, salinity and air supply were the same as those used before while maintaining crabs in a rectangular tank before the start of the experiment. All replicates in each experiment were conducted with healthy, active crabs and sand dollars after 24 h of initial acclimation. No crab was used more than once during each experiment.
Experiment 1: movements of crabs between sand dollar individuals
This experiment was conducted to test if male and female crabs are sedentary or if they move rather frequently among different host individuals. In this first experiment, either four male crabs (treatment 1, n = 30 replicates) or four female crabs (treatment 2, n = 30 replicates) were maintained together with two sand dollars in a single circular tank (see above). A total of 240 crabs were used in this experiment; 120 males with average (X) ± standard deviation (SD) = 5.92 ± 0.59 mm CW and 120 females with X ± SD = 6.38 ± 0.56 mm CW). However, due to logistic constraints, only 20 sand dollars (maximum diameter: X ± SD = 11.24 ± 0.56 mm MD) were used (10 per treatment, with sand dollar pairs haphazardly assigned to each replicate). Before the start of each replicate, either the four male or female crabs in treatment 1 or 2, respectively, were placed together on the oral surface of one sand dollar that was tagged by gently tying a thin colorless fishing line to one of its lunulae. The second sand dollar in the same replicate was not tagged. During the experiment, the two sand dollars could roam around freely in the circular tank. All experimental crabs were individually marked with a non-toxic permanent pen on the dorsal carapace. The position of each crab in each replicate was recorded twice, once at 12 h, and then again at 24 h after the start of each replicate. Each sand dollar was carefully checked for the presence/absence of crabs. Crabs not observed on the surface of sand dollars were recorded as dwelling among the sand grains. Crabs were either recorded on the tagged host, on the untagged host, or among the sand grains without any contact to sand dollars. To compare the number of crabs (males and females) that had left the initial host after 12 and 24 h, we used a linear mixed effects (LME) model due to the time-dependency of observations (12 and 24 h) (Crawley 2013).
We are aware that the experimental crabs might have moved away from and back to their original hosts between observation periods and, thus, our observations represent conservative minimum estimates of crab movement from an original host individual (see Thiel et al. 2003).
Experiment 2: fidelity of paired heterosexual crabs
This experiment was conducted to test for fidelity between male and female crabs found in heterosexual pairs in the field. In the laboratory, one ‘intruder’ crab (male or female, depending upon treatment) was placed on the surface of the same host individual already harboring a heterosexual pair retrieved from the field.
In this experiment, one heterosexual pair plus an extra ‘intruder’ male crab (treatment 1, n = 30 replicates) or one heterosexual pair plus an extra ‘intruder’ female crab (treatment 2, n = 30 replicates) were maintained with two sand dollars in the same circular tank. A total of 180 crabs were used; 90 males with X ± SD = 6.26 ± 0.55 mm CW and 90 females with X ± SD = 6.39 ± 0.64 mm CW). However, as in the first experiment, only 20 sand dollars (maximum diameter: X ± SD = 10.64 ± 0.61 mm MD) were used (10 per treatment, with sand dollar pairs haphazardly assigned to each replicate). Before the start of each replicate, either the heterosexual pair plus the ‘intruder’ male or female crab in treatments 1 and 2, respectively, were placed together on the oral surface of one sand dollar that was tagged by gently tying a thin colorless fishing line to one of its lunulae. The second sand dollar in the same replicate was not tagged. During the experiment, the two sand dollars could roam around freely in the circular tank. In the two treatments, only intruder crabs were marked with a non-toxic permanent pen on the dorsal carapace. The position of each crab in each replicate was recorded as described in the previous experiment. We tested whether or not the sex of the intruder affected the population distribution of crabs (i.e., frequency of occurrence of solitary crabs, pairs, and trios) in this experiment using a contingency table (Zar 2010). Also, we tested whether or not the sex of the intruder affected the proportion of crabs found in heterosexual pairs in the field. For this purpose, we specifically compared the frequency of occurrence of field-collected heterosexual pairs between the two treatments using a second contingency table (Zar 2010). These analyses above were conducted separately for data recorded after 12 and 24 h, considering the dependence of the data between observation times. If D. crinitichelis does not exhibit long-term pairing (i.e., is not socially monogamous), we expected that the field pairs would quickly separate in this experiment.
As in the first experiment, we are aware that the experimental crabs might have moved away from and back to their original hosts between observation periods and, thus, our observations represent conservative minimum estimates of crab movements from an original host individual (see Thiel et al. 2003).
Results
Host-use pattern of Dissodactylus crinitichelis
The density of the sand dollar E. emarginata ranged from 1.5 to 3.25 individuals m−2 (X ± SD = 2.35 ± 0.67) at Croa do Goré (CG) and from 0.75 to 6.25 individuals m−2 (X ± SD = 2.17 ± 1.58) at Ilha dos Namorados (IN). Host density did not differ statistically between the two sites (Mann–Whitney U test, U = 33, n 1 = 20, n 2 = 20, P = 0.195). Sand dollar body size ranged from 57.0 to 132.0 mm MD (X ± SD = 110.9 ± 11.6) at CG and from 66.0 to 150.0 mm MD (X ± SD = 113.1 ± 12.7) at IN. Sand dollar body size did not differ statistically between the two sites (Mann–Whitney U test, U = 4548, n 1 = 200, n 2 = 200, P = 0.268).
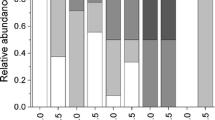
Frequency of occurrence of crabs on sand dollars was 81% at CG and 96% at IN (Table 1). The difference in the frequency of occurrence of crabs on sand dollars between the two study localities was significant (Chi square test of independence, χ 21 = 11.05, P < 0.001). Most often, crabs were not present on sand dollars smaller than 79 mm in test length. Furthermore, three or more crabs were recorded only on the sand dollars in the size classes larger than 103–111 and 95–103 mm at CG and IN, respectively (Fig. 2a, b).

The effect of host body size in host occupancy by Dissodactylus crinitichelis. Relative abundance of D. crinitichelis pea crabs per sand dollar host individual in different host body size classes at a Croa do Goré and b Ilha dos Namorados, Brazil
A total of 412 crabs were collected. Of these, 154 (37.37%) crabs were collected at CG and 258 (62.62%) at IN (Table 1). Of the crabs collected at CG and IN, 51 (33.11%) and 174 (67.44%), respectively, were juveniles or crabs whose sex was not possible to determine (Table 1). Considering only sexed crabs, the sex ratio of D. crinitichelis did not differ significantly from a 1:1 ratio at CG (sex ratio = 0.543; binomial test, P = 0.351) and IN (sex ratio = 0.476; binomial test, P = 0.704).
Considering only those sand dollars harboring crabs, the density of D. crinitichelis ranged from 1 to 4 individuals (X ± SD = 1.9 ± 0.94) at CG and from 1 to 12 individuals (X ± SD = 2.68 ± 1.52) at IN. A strong positive correlation was observed between host size and the number of crabs per sand dollar at CG (Linear regression, F 1,98 = 10.37, r 2 = 0.095, P = 0.001). On the other hand, no correlation between host size and the number of crabs per sand dollar was observed at IN (F 1,98 = 1.21, r 2 = 0.012, P = 0.273). Also, a positive correlation was observed between host size and the number of female crabs found on a single sand dollar host (F 1,56 = 5.03, r 2 = 0.066, P = 0.028) and between host size and the number of male crabs (F 1,98 = 4.72, r 2 = 0.036, P = 0.032) at CG. No correlation was found between host size and the number of female crabs found in a single sand dollar host (F 1,98 = 2.01, r 2 = 0.010, P = 0.159) and between host size and the number of male crabs (F 1,98 = 0.32, r 2 = −0.006, P = 0.570) at IN.
Population distribution in Dissodactylus crinitichelis
For the analysis of population distribution and male–female association pattern in D. crinitichelis, the data from the two localities were pooled together because of the absence of differences among sampling sites in the body size of hosts and the minimal differences in occupancy pattern of hosts by crabs (see above). The total number of D. crinitichelis crabs per sand dollar host ranged from 0 to 12 (X ± SD = 2.07 ± 1.47). The sex ratio of D. crinitichelis was not statistically significantly from an expected unbiased (1:1) sex ratio in all different group sizes (binomial test, P > 0.05). A two-way ANOVA did not demonstrate an effect of group size (F 6,285 = 1.18, P = 0.31) or sex (F 1,285 = 1.15, P = 0.28) on crab CW. The interaction term in this analysis was not statistically significant (F 6,285 = 0.71, P = 0.64).
Considering the totality of our samples (including both mature and immature individuals), a total of 41 (20.5%), 21 (10.5%), 5 (2.5%), 1 (0.5%), and 1 (0.5%) sand dollars harbored three, four, five, six, and twelve crabs, respectively (Fig. 3a). These frequencies are smaller when only adult crabs are considered in our calculations, of which a total of 4 (2.0%) and 2 (1.0%) sand dollars harbored, respectively, three and four crabs (Fig. 3b).

Population distribution of the pea crab Dissodactylus crinitichelis, symbiotic with the sand dollar Encope emarginata at Vaza-Barris estuary, Brazil. a Observed population distribution of all crabs found on hosts does not differ significantly from the expected Poisson random distribution. b Observed frequency only for adult crabs found on hosts differs significantly from the expected Poisson random distribution
The distribution of Dissodactylus crinitichelis on the sand dollar host did not differ significantly from a Poisson random distribution (observed vs. expected Poisson distribution, Chi square test of goodness-of-fit, χ 24 = 2.70, P = 0.608) (Fig. 3a). However, when we analyzed the distribution only of adult crabs, significant differences between observed vs. expected Poisson distributions were found (observed vs. expected Poisson distribution only for adult crabs, Chi square test of goodness-of-fit, χ 22 = 12.66, P = 0.001) (Fig. 3b). The discrepancy between observed and expected distribution was mostly explained by the observed number of hosts harboring two adult crabs that was higher than expected by chance alone (χ 21 = 7.61, P = 0.005, see below).
A total of 53 sand dollars harbored a single symbiotic crab over the entire study period (26.50% of all sand dollars collected and 29.94% of the total sand dollars infected). A total of 25 (47.17% of 53 sand dollars harboring a single crab) sand dollars were infected with a single female crab (7 juveniles, 6 non-ovigerous, and 12 ovigerous); 21 (39.62% of 53 sand dollars harboring a single crab) sand dollars harbored a single male (8 juveniles and 13 adults); and 7 (13.21% of 53 sand dollars harbored a single crab) sand dollars harbored a crab of unknown sex.
A total of 50 sand dollars harbored two adults crabs, of which 20 (40%) sand dollars harbored only two adults crabs, while the other 30 (60%) sand dollars harbored a pair of adults crabs and one or more non-reproductive crabs (juveniles and/or crabs of unknown sex). In both cases, the number of sand dollars inhabited by a heterosexual pair (n = 17 and 43, respectively) was significantly higher than the number predicted by a random binomial distribution (Chi square test of independence, χ 21 = 21.99, P < 0.001 and χ 21 = 58.50, P < 0.001, respectively) (Fig. 4a, c). Moreover, both in sand dollars inhabited by only two adult crabs, as in those inhabited by two adult crabs and one or more non-reproductive crabs, no correlation was observed between the body size of D. crinitichelis males and females comprising adult heterosexual pairs (Spearman rank correlation, r s = 0.040, n = 17, P = 0.877 and r s = −0.040, n = 43, P = 0.745, respectively) (Fig. 4b, d).

Male–female association pattern of Dissodactylus crinitichelis found as pairs on sand dollar host individuals. a Observed frequency of pair of adults crabs (n = 20) differs significantly from the expected binomial random distribution. b Relationship between carapace width (CW) of males and females of D. crinitichelis found as adults heterosexual pairs (n = 17) on sand dollar host individuals. c Observed frequency of pair of adults crab without (n = 20) or with (n = 30) no reproductive crabs differs significantly from the expected binomial random distribution. d Relationship between carapace width (CW) of males and females of D. crinitichelis found as adults heterosexual pairs (n = 43) on sand dollar host individuals
Sexual dimorphism in Dissodactylus crinitichelis
The carapace width (CW) of male and female crabs varied between 2.22 and 7.58 mm (X ± SD = 5.16 ± 1.43) and between 2.71 and 8.46 mm (X ± SD = 5.60 ± 1.36), respectively (Fig. 5a). Females attain larger final and average body sizes than males in Dissodactylus crinitichelis (t test, t 297 = 2.72, P = 0.006).

Sexual dimorphism in Dissodactylus crinitichelis. a Size frequency distribution of body size (CW) in males and females. (b) Relative growth of the left propodus length as a function of carapace width. Scale bars = 5 mm. Linear regression equations obtained after log–log transformation of the data are shown for each sex in Table 1
A positive correlation was detected between CW and the length of the propodus of the claw in crabs of the two sexes (Table 2; Fig. 5b). In males, the growth of the length of the claw was positively allometric with respect to CW; the slope of the relationship between male CW and claw length differed significantly from unity (b = 1.50, P < 0.01). In females, the growth of the length of the claw was also positively allometric with respect to CW (b = 1.24, P < 0.01). An analysis of covariance (ANCOVA) indicated a significant effect of sex (F 1,107 = 16.41, P < 0.01) and CW in claw length (F 1,107 = 3066.44, P < 0.01) and the interaction term of this analysis was significant (F 1,107 = 26.57, P < 0.01) (Table 2). Although the length of the claw and the growth rate of this structure were greater in males than in females of D. crinitichelis, sexual dimorphism in terms of claw size was not considerable in the species (Fig. 5b).
Experiment 1: movements of crabs between sand dollar individuals
In this experiment, to determine if male and/or female crabs switch between host individuals, nearly 35 and 60% (after 12 and 24 h, respectively) of the male crabs were observed to leave sand dollars on which they were originally placed at the start of the experiment. In turn, nearly 40 and 55% of the female crabs departed from the original host after 12 and 24 h, respectively (Fig. 6). The difference between male and female crabs in movement rate was not statistically significant (Linear mixed effects model, F 1,58 < 0.0001, P = 1.00, Fig. 6) and the number of crabs that left the initial host individual increased over time (after 12 and 24 h) for both sexes (F 1,29 = 27.040, P < 0.001, Fig. 6).

Movemement pattern of Dissodactylus crinitichelis under experimental conditions. Average percentage (±SD) of male and female crabs Dissodactylus crinitichelis that left the initial host after 12 and 24 h. For each treatment (male or female), 30 replicas were conducted. A linear mixed effects (LME) model test was conducted due to time dependency of the experimental units
Experiment 2: fidelity of paired heterosexual crabs
Of the 60 females of D. crinitichelis collected and used in this experiment, 53.33% (n = 32) carried embryos, of which 37.50% (n = 12) carried embryos in stage I, 31.25% (n = 10) carried embryos in stage II, and 31.25% (n = 10) carried embryos in stage III of development.
At the start of this second experiment, one of the sand dollars did not harbor any crab while the second sand dollar harbored a crab trio (i.e., heterosexual pair retrieved from the field plus an intruder) (Fig. 7a). After 12 h of initiating the experiment, when the intruder was a female, the crab population distribution was the following: in 16 (53.33%) replicates we observed a pair of crabs cohabiting on the same sand dollar (14 of these were heterosexual pairs) while a third crab was found either in the sediment or on the second sand dollar living solitarily, in 9 (30%) replicates we observed crab trios (all crabs inhabiting the same host), and in 5 (16.67%) replicates we observed two solitary crabs, one per available host, while the third crab was found in the sediment (Fig. 7b). After 12 h of initiating the experiment, when the intruder was a male, the crab population distribution was the following: in 20 (66.67%) replicates we observed a pair of crabs cohabiting on the same sand dollar (19 of these were heterosexual pairs) while a third crab was found either in the sediment or on the second sand dollar living solitarily, in 4 (13.33%) replicates we observed crab trios (all crabs inhabiting the same host), and in 6 (20%) replicates we observed two solitary crabs, one per available host, while the third crab was found in the sediment (Fig. 7b).

Fidelity of paired heterosexual crabs in Dissodactylus crinitichelis under experimental conditions. Percentage of replicates in which crabs were found on sand dollars solitarily, in pairs, or in trios at the start of the second experiment (a), after 12 (b), and after 24 (c) hours of initiating the experiment, when a pair plus a male or a female intruder crab of Dissodactylus crinitichelis were introduced in the same container. Percentage of original pairs and new pairs formed during the experimental period after 12 (d) and after 24 (e) hours of initiating the experiment. For each treatment (male or female intruder crab), 30 replicas were conducted
After 24 h of initiating the experiment, when the intruder was a female, the crab population distribution was the following: in 19 (63.33%) replicates we observed a pair of crabs cohabiting on the same sand dollar (17 of these were heterosexual pairs) while a third crab was found either in the sediment or on the second sand dollar living solitarily, in 6 (20%) replicates we observed crab trios (all crabs inhabiting the same host), and in 5 (16.67%) replicates we observed two solitary crabs, one per available host, while the third crab was found in the sediment (Fig. 7c). After 24 h of initiating the experiment, when the intruder was a male, the crab population distribution was the following: in 18 (60%) replicates we observed a pair of crabs cohabiting on the same sand dollar (17 of these were heterosexual pairs) while a third crab was found either in the sediment or on the second sand dollar living solitarily, in 4 (13.33%) replicates we observed crab trios (all crabs inhabiting the same host), and in 8 (26.67%) replicates we observed we observed two solitary crabs, one per available host, while the third crab was found in the sediment (Fig. 7c). The observed population distribution (frequency of occurrence of crabs inhabiting the host solitarily, in pairs or trios) did not differ statistically between treatments after 12 h (Contingency table, χ 22 = 2.46, P = 0.292, Fig. 7b) and 24 h (χ 22 = 1.12, P = 0.571, Fig. 7c) of initiating the experiment.
After 12 h of initiating the experiment, when the intruder was a female, the frequency of heterosexual pairs (i.e., retrieved from the field or newly formed pairs) was the following: in 11 (78.57%) of the 14 replicates with a heterosexual pair we observed the original couple of crabs (i.e., heterosexual pair retrieved from the field) cohabiting on the same sand dollar and in 3 (21.43%) replicates we observed an heterosexual pair comprising the intruder female and a male (Fig. 7d). When the intruder was a male, the frequency of heterosexual pairs was the following: in 10 (52.63%) of the 19 replicates with a heterosexual pair we observed the original couple of crabs cohabiting on the same sand dollar, in 9 (47.37%) we observed a couple formed with the intruder male (Fig. 7d). After 24 h of initiating the experiment, when the intruder was a female, the frequency of heterosexual pairs was the following: in 9 (52.94%) of the 17 replicates with a heterosexual pair we observed the original couple of crabs cohabiting on the same sand dollar, in 8 (47.06%) we observed a couple formed with the intruder female (Fig. 7e). When the intruder was a male, the frequency of heterosexual pairs was the following: in 8 (47.06%) of the 17 replicates with a heterosexual pair we observed the original couple of crabs cohabiting on the same sand dollar, in 9 (52.94%) we observed a couple formed with the intruder male (Fig. 7e). The frequency of occurrence of heterosexual pairs retrieved from the field and newly formed pairs did not differ statistically between treatments after 12 h (Contingency table, χ 21 = 2.34, P = 0.126, Fig. 7d) and 24 h (χ 21 = 0.12, P = 0.732, Fig. 7e).
Discussion
In this study, we hypothesized that the symbiotic crab Dissodactylus crinitichelis was not monogamous but rather promiscuous. We expected that (1) the population distribution of this crab on Encope emarginata would be random, (2) the sex distribution of crabs in pairs would also be random with male–female pairs occurring less often than expected by chance alone or not occurring at all, (3) heterosexual pairs (if any) should not exhibit size-assortative pairing, (4) sexual dimorphism in body size and weaponry should be observed, and (5) crabs should exhibit frequent movements between host individuals and heterosexual pairing should not be stable over time. Our field observations and laboratory experiments agree with the notion that D. crinitichelis is not monogamous and further suggest that this pea-crab exhibits a promiscuous mating system. In the following, we discuss particular characteristics of the association between D. crinitichelis and E. emarginata that argue against monogamy and in favor of promiscuity in the studied crab.
Dissodactylus crinitichelis lives in pairs but pairing is not size-assortative
In D. crinitichelis, the entire (adults + juveniles) population exhibited a random pattern of distribution on sand dollars. However, when only adult crabs were considered, the number of hosts inhabited by paired crabs was greater than that expected by chance alone. Furthermore, adult crabs found in pairs were, almost invariably, heterosexual. The above is often reported for socially monogamous crustaceans (e.g., Pinnixa chaetopterana—Grove and Woodin 1996; Pinnixa transversalis—Baeza 1999; Pontonia margarita—Baeza 2008; Paranchistus pycnodontae—Baeza et al. 2013; Planes major—Pfaller et al. 2014). Nonetheless, if D. crinitichelis was socially monogamous, heterosexual pairs should have exhibited size-assortative pairing, as reported before for symbiotic species that exhibit long-term monogamy (e.g., the coral crab Trapezia ferruginea—Adams et al. 1985; the pea-crab Pinnixa transversalis—Baeza 1999; the shrimp Pontonia margarita—Baeza 2008; the shrimp Paranchistus pycnodontae—Baeza et al. 2013).
Most often, in symbiotic and monogamous crustaceans, in which male and female individuals develop a long-lasting association with one another as well as with their host individuals, a tight correlation between male and female individuals forming pairs is found (Baeza 1999, 2008). Growth limitations imposed by host individuals over their long-term resident crustaceans are invoked to explain such tight male–female symbiont body size as well as host-crustacean body size relationships (see Baeza et al. 2015 and references therein). Our results in D. crinitichelis were in contrast to that reported for symbiotic monogamous species. The weak correlation between the body size of paired male and female adult crabs and between the body size of crabs and the sand dollars hosting them instead suggests that male (and/or female) crabs are shifting among host individuals frequently, in agreement with that reported for putatively promiscuous symbiotic crustaceans (e.g., the porcelain crab Petrolisthes mitra living on sea urchins—Baeza and Thiel 2000).
Dissodactylus crinitichelis exhibits reverse sexual dimorphism
The observed pattern of sexual dimorphism in D. crinitichelis additionally suggests that this crab does not exhibit a socially monogamous mating system and also argues in favor of promiscuity in this species. If D. crinitichelis was socially monogamous, we should have observed this species to display little to no sexual dimorphism in body size and weaponry (e.g., chelipeds used for intra-sexual agonistic interaction) (Baeza and Thiel 2007). The pattern of reverse sexual dimorphism in terms of body size observed in D. crinitichelis (i.e., males < females) is often observed in species in which males compete for females using pure-search mating strategies (Bauer 2004; Baeza and Asorey 2012). In species with a ‘pure-search’ mating system, males constantly search for receptive females, mate quickly, and abandon them shortly after insemination (Correa and Thiel 2003; Bauer 2004). Thus, given that males neither defend females nor resources, there is no selection for large body size in males (Bauer and Thiel 2011). In D. crinitichelis, intra-sexual selection might favor a small male body size because this leads to increases in agility as well as higher encounter rate of males with potential mating partners (Shuster and Wade 2003; Baeza and Thiel 2007). On the other hand, larger body size is likely adaptive in females of D. crinitichelis, because female size correlates well with fecundity in pea crabs (Baeza and Hernáez 2015 and references therein).
Besides body size, the sex-specific difference in resource allocation to the chelipeds also disagrees with a prediction of low sexual dimorphism in terms of weaponry in monogamous species (Baeza and Thiel 2007). Large claws in males compared to females are uncommon in monogamous crustaceans, but common in non-monogamous crustaceans where males frequently compete for females through agonistic interactions (Bauer 2004; Baeza and Thiel 2007). Thus, the slightly larger chelipeds in adult males of D. crinitichelis (compared to females) suggest that males are competing for receptive females via overt aggression. Our results on sexual dimorphism in D. crinitichelis suggest that the body size and allocation to weaponry in males might represent an evolutionary compromise between the optimal body size for switching among hosts while using little energy, and the optimal allocation to weaponry for successful intra-sex competition for females after they are found (Baeza et al. 2016). Additional studies are needed to understand those conditions driving sexual dimorphism with respect to body size and weaponry in symbiotic crustaceans.
Dissodactylus crinitichelis heterosexual pairs do not last long
Lastly, the two laboratory experiments conducted during the present study argue against the notion of long-term monogamy in D. crinitichelis. In the first experiment, in which we examined the movement of crabs between sand dollar individuals, both male and female crabs were observed to abandon host individuals rather soon, within a few hours of starting the experiment. This frequent movement of crabs between host individuals does not fit to what would have been expected for a monogamous species with sedentary males and females. Indeed, our observations fit to that reported for other echinoderm dwelling crabs in which individuals swap among host individuals rather frequently (i.e., Petrolisthes mitra—Thiel et al. 2003). Frequent host-switching behaviors have also been reported before for other promiscuous symbiotic crustaceans that live associated with echinoderms (e.g., the crab Zebrida adamsii—Yanagisawa and Hamaishi 1986; the shrimp Athanas indicus–Gherardi 1991; the shrimp Rhynchocinetes brucei—Ory et al. 2013).
Similarly, in the second experiment we observed that the original heterosexual pair dissolved rather quickly, with males and females not remaining together with their original sexual partners for long times. Furthermore, the formation of new pairs that was often observed during the experiment occurred independently of the sex of the intruder crab. Thus, we interpret the results above as further indication for promiscuity in D. crinitichelis. Although we have manipulated hosts and crabs in both experiments, which may have resulted in higher rates of movement of the crabs, the patterns observed in the experiments are consistent with the notion of pure-search promiscuity in D. crinitichelis.
Dissodactylus crinitichelis is not monogamous
In general, our field and laboratory data argue against monogamy and in favor of promiscuity in D. crinitichelis. Nonetheless, our data do not allow us to definitively conclude that the studied species is promiscuous. Indeed, our study does not reveal additional details regarding the mating behavior of the studied pea crab. Some questions still remain open and additional studies are needed to answer the following: (1) do females copulate during the intermolt period?, (2) do males guard females right before or after copula?, (3) do females exhibit passive or active mate choice?, and (4) in the case males do guard females (even for a short time period), are intruder males actively prevented from occupying the same host individual? The population distribution and, likely, the mating system of D. crinitichelis are similar to that previously reported for Dissodactylus primitivus, an ectoparasite of two burrowing sea urchins (Meoma ventricosa and Plagiobrissus grandis) (Jossart et al. 2014). De Bruyn et al. (2009, 2010) proposed that D. primitivus exhibits a promiscuous mating system, from field data and experiments. Later, Jossart et al. (2014) verified the occurrence of both polyandry and polygyny for this pea crab using microsatellite markers.
The family Pinnotheridae, to which D. crinitichelis belongs, is a species-rich clade of symbiotic brachyuran crabs with considerable disparity in ecological traits. Members from this family inhabit a diverse array of hosts, including bivalves, polychaetes, burrowing shrimp, gastropods, holothurians, tunicates, and echinoids. These host species also differ widely in terms of biology and ecology (Bell 1984). We have shown that a pea crab that inhabits a relatively large and abundant host species is not monogamous but likely promiscuous, in agreement to that reported for another congeneric species that also lives on a relatively large and abundant host species (i.e., Dissodactylus primitivus—De Bruyn et al. 2009, 2010). In turn, other pea-crab species inhabiting relatively small and scarce host species are socially monogamous (i.e., Pinnixa transversalis—Baeza 1999). Future experimental studies should continue investigating the reproductive behavior of pea crabs concomitantly with host characteristics (i.e., body size, morphological complexity, abundance) as these host-pea-crab associations represent excellent models to improve our understanding about the origin and adaptive value of social and mating systems in symbiotic crustaceans.
Outlook
In this study, we have provided information indicating that D. crinitichelis does not exhibit long-term monogamy as a mating system. Our study supports the notion that environmental conditions, including ecological (i.e., abundance) and morphological (i.e., structural complexity) aspects of the host species, drive the behavior of symbiotic crustaceans. Although we can not definitively conclude that D. crinitichelis is a promiscuous species, the characteristics of and habitat in which E. emarginata lives appears to favor promiscuity in its symbiotic crustacean. Promiscuity has been shown to be advantageous in females from other species. For instance, promiscuity may, among others, ensure sperm supply to females (Sainte-Marie 2007), increase female fecundity or longevity (Jennions 1997) and enhance offspring genetic diversity (Andersson 1994; Jennions and Petrie 2000). Thus, not only environmental features (e.g., host size and abundance) might be favoring promiscuity in D. crinitichelis but also other conditions (increased female fecundity, sperm supply, offspring genetic diversity) affecting female reproductive success. We propose that monophyletic clades of pea crabs (fam. Pinnotheridae) exhibiting both disparity in terms of mating systems (i.e., monogamy, promiscuity, polygyny) and host biology/ecology (i.e., abundance, morphological complexity, relative size) represent a model group to understand the evolution of male and female mating strategies in symbiotic marine invertebrates. Pea crabs inhabiting host species with differing ecology are expected to display dissimilar mating systems.
References
Adams J, Edwards AJ, Emberton H (1985) Sexual size dimorphism and assortative mating in the obligate coral commensal Trapezia ferruginea Latreille (Decapoda, Xanthidae). Crustaceana 48:188–194
Ambrosio LJ, Baeza JA (2016) Territoriality and conflict avoidance explain a sociality (solitariness) of the endosymbiotic pea crab Tunicotheres moseri. PLoS One. doi:10.1371/journal.pone.0148285
Andersson M (1994) Sexual selection. Princeton University Press, Princeton
Baeza JA (1999) Indicators of monogamy in the commensal crab Pinnixa transversalis (Milne Edwards & Lucas) (Decapoda: Brachyura: Pinnoteridae): population distribution, male-female association and sexual dimorphism. Rev Biol Mar Oceanogr 34:303–313
Baeza JA (2008) Social monogamy in the shrimp Pontonia margarita, a symbiont of Pinctada mazatlantica, in the tropical eastern Pacific coast. Mar Biol 153:387–395
Baeza JA (2015) Crustaceans as symbionts: an overview of their diversity, host use and life styles. In: Watling L, Thiel M (eds) The life styles and feeding biology of the Crustacea. Oxford University Press, Oxford, pp 163–189
Baeza JA, Asorey CM (2012) Testing the role of male–male competition in the evolution of sexual dimorphism: a comparison between two species of porcelain crabs. Biol J Linn Soc Lond 105:548–558
Baeza JA, Hernáez P (2015) Population distribution, sexual dimorphism, and reproductive parameters in the crab Pinnixa valdiviensis Rathbun, 1907 (Decapoda: Pinnotheridae). J Crustac Biol 35:68–75
Baeza JA, Piantoni C (2010) Sexual system, sex ratio and group living in the shrimp Thor amboinensis (De Man): relevance to resource-monopolization and sex-allocation theories. Biol Bull 219:151–165
Baeza JA, Stotz W (2003) Host-use and selection of differently colored sea anemones by the symbiotic crab Allopetrolisthes spinifrons. J Exp Mar Biol Ecol 284:25–39
Baeza JA, Thiel M (2000) Host use pattern and life history of Liopetrolisthes mitra, a crab associate of the black sea urchin Tetrapygus niger. J Mar Biol Ass UK 80:639–645
Baeza JA, Thiel M (2003) Predicting territorial behavior in symbiotic crabs using host characteristics: a comparative study and proposal of a model. Mar Biol 142:93–100
Baeza JA, Thiel M (2007) The mating system of symbiotic crustaceans. A conceptual model based on optimality and ecological constraints. In: Duffy JE, Thiel M (eds) Reproductive and social behavior: Crustaceans as model systems. Oxford University Press, Oxford, pp 245–255
Baeza JA, Ritson-Williams R, Fuentes MS (2013) Sexual and mating system in a caridean shrimp symbiotic with the winged pearl oyster in the coral triangle. J Zool 289:172–181
Baeza JA, Hemphill CA, Ritson-Williams R (2015) The sexual and mating system of the shrimp Odontonia katoi (Palaemonidae, Pontoniinae), a symbiotic guest of the ascidian Polycarpa aurata in the coral triangle. PLoS One. doi:10.1371/journal.pone.0121120
Baeza JA, Simpson L, Ambrosio LJ, Guéron R, Mora N (2016) Monogamy in a hyper-symbiotic shrimp. PLoS One. doi:10.1371/journal.pone.0149797
Bauer RT (2004) Remarkable shrimps. Oklahoma University Press, Norman
Bauer RT, Thiel M (2011) First description of a pure-search mating system and protandry in the shrimp Rhynchocinetes uritai (Decapoda: Caridea). J Crustac Biol 31(2):286–295
Bell JL (1984) Changing residence: dynamics of the symbiotic relationship between Dissodactylus mellitae Rathbun (Pinnotheridae) and Mellita quinquiesperforata (Leske) (Echinodermata). J Exp Mar Biol Ecol 82:101–115
Campos E, Campos AR, León-González JA (2009) Diversity and ecological remarks of ectocommensals and ectoparasites (Annelida, Crustacea, Mollusca) of echinoids (Echinoidea: Mellitidae) in the Sea of Cortez, Mexico. Parasitol Res 105:479–487
Carranza A, Domingo A, Verdi A, Forselledo R, Estrades A (2003) First report of an association between Planes cyaneus (Decapoda: Grapsidae) and loggerhead sea turtles in the Southwestern Atlantic Ocean. Mar Turtle Newsl 102:5–7
Correa C, Thiel M (2003) Mating systems in caridean shrimp (Decapoda: Caridea) and their evolutionary consequences for sexual dimorphism and reproductive biology. Rev Chil Hist Nat 76:187–203
Crawley MJ (2013) Statistical computing—an introduction to data analysis using s-plus. Wiley, London
De Bruyn C, Rigaud T, Bruno D, Ridder C (2009) Symbiosis between the pea crab Dissodactylus primitivus and its echinoid host Meoma ventricosa: potential consequences for the crab mating system. Mar Ecol Prog Ser 375:173–183
De Bruyn C, David B, Ridder C, Rigaud T (2010) Asymmetric exploitation of two echinoid host species by a parasitic pea crab and its consequences for the parasitic life cycle. Mar Ecol Prog Ser 398:183–191
De Bruyn C, David B, Motreuil S, Caulier G, Jossart Q, Rigaud T, De Ridder C (2016) Should I stay or should I go? Causes and dynamics of host desertion by a parasitic crab living on echinoids. Mar Ecol Prog Ser 546:163–171
Duffy JE (1996) Eusociality in a coral-reef shrimp. Nature 381:512–514
Duffy JE (2007) Ecology and evolution of eusociality in sponge-dwelling shrimp. In: Duffy JE, Thiel M (eds) Evolutionary ecology of social and sexual systems: crustaceans as model organisms. Oxford University Press, Oxford, pp 387–409
Elliott JM (1983) Some methods for the statistical analysis of samples of benthic invertebrates, 3rd edn. Freshwater Biological Association, Far Sawrey
Gherardi F (1991) Eco-ethological aspects of the symbiosis between the shrimp Athanas indicus (Coutière 1903) and the sea urchin Echinometra mathaei (de Blainville 1825). Trop Zool 4(1):107–128. doi:10.1080/03946975.1991.10539481
Gray IE, McCloskey LR, Weihe SC (1968) The commensal crab Dissodactylus mellitae and its reaction to sand dollar host-factor. J Elisha Mitchell Sci Soc 84:472–481
Grove MW, Woodin SA (1996) Conspecific recognition and host choice in a pea crab, Pinnixa chaetopterana (Brachyura: Pinnotheridae). Biol Bull 190:359–366
Guilherme PDB, Brustolin MC, Bueno ML (2015) Distribution patterns of ectosymbiont crabs and their sand dollar hosts in a subtropical estuarine sandflat. Rev Biol Trop 63(Suppl. 2):209–220
Hartnoll RG (1978) The determination of relative growth in Crustacea. Crustaceana 34(3):281–293
Hartnoll RG (1982) Growth. In: Bliss DE, Abele LG (eds) The biology of Crustacea, 2, embryology, morphology and genetics. Academic Press, New York, pp 111–196
Jennions MD (1997) Female promiscuity and genetic incompatibility. Trends Ecol Evol 12(7):251–253
Jennions MD, Petrie M (2000) Why do females mate multiply? A review of the genetic benefits. Biol Rev 75:21–64
Jossart Q, Wattier RA, Kastally C, Aron S, David B, Ridder CD, Riguad T (2014) Genetic evidence confirms polygamous mating system in a crustacean parasite with multiple hosts. PLoS One. doi:10.1371/journal.pone.0090680
Jr Haefner PA (1990) Morphometry and size at maturity of Callinectes ornatus (Brachyura, Portunidae) in Bermuda. Bull Mar Sci 46:274–286
Kathiresan K, Bingham BL (2001) Biology of mangroves and mangrove ecosystems. Adv Mar Biol 40:81–251
Martin JW, Davis GE (2006) Historical trends in crustacean systematics. Crustaceana 79(11):1347–1368
Martinelli-Filho JE, Santos RB, Ribeiro CC (2014) Host selection, host-use pattern and competition in Dissodactylus crinitichelis and Clypeasterophilus stebbingi (Brachyura: Pinnotheridae). Symbiosis. doi:10.1007/s13199-014-0292-0
Nogata Y, Matsumura K (2005) Larval development and settlement of a whale barnacle. Biol Lett 2:92–93
Ocampo EH, Nuñez JD, Cledón M, Baeza JA (2012) Host-specific reproductive benefits, host selection behavior and host use pattern of the pinnotherid crab Calytraeotheres garthi. J Exp Mar Biol Ecol 429:36–46
Ory NC, Dudgeon Thiel M (2013) Host-use patterns and factors influencing the choice between anemone and urchin hosts by a caridean shrimp. J Exp Mar Biol Ecol 449:85–92
Pérez GHB, Serrato MB, Sanchez CMD (2012) Equinodermos del Caribe colombiano II: Echinoidea Y Holothuroidea. Serie de Publicaciones Especiales de Invemar, Santa Marta
Pfaller JB, Alfaro-Shigueto J, Giffoni B, Ishihara T, Mangel JC, Peckham SH, Bjorndal KA, Baeza JA (2014) Social monogamy in the crab Planes major, a facultative symbiont of loggerhead sea turtles. J Exp Mar Biol Ecol 461:124–132
Pires-Vanin AMS (2001) Identifying the components of ecological variation in a marine benthic megafauna. Rev bras oceanogr 49(1/2):29–38
Reeves MV, Brooks WR (2001) Host selection, chemical detection, and protection of the symbiotic pinnotherid crabs Dissodactylus crinitichelis and Clypeasterophilus rugatus associated with echinoderms. Symbiosis 30:239–256
Sainte-Marie B (2007) Sperm demand and allocation in decapod Crustaceans. In: Duffy JE, Thiel M (eds) Reproductive and social behavior: Crustaceans as model systems. Oxford University Press, Oxford, pp 191–210
Shuster SM, Wade MJ (2003) Mating systems and strategies. Princeton University Press, Princeton
Sokal RR, Rohlf FJ (1981) Biometry, 3rd edn. Freeman, San Francisco
Thiel M, Zander A, Baeza JA (2003) Movements of the symbiotic crab Liopetrolisthes mitra between its host sea urchin Tetrapygus niger. Bull Mar Sci 72:89–101
Wehrtmann IS (1990) Distribution and reproduction of Ambidexter panamense and Palaemonetes schmitti in Pacific Costa Rica (Crustacea, Decapoda). Rev Biol Trop 38:327–329
Williams AB (1984) Shrimps, lobsters, and crabs of the Atlantic coast of the eastern United States, Maine to Florida. Smithsonian Institution Press, Washington
Wirtz P, Melo GAS, Grave S (2009) Symbioses of the decapod crustaceans along the coast of Espírito Santo, Brazil. Mar Biodivers Rec 2:1–9
Yanagisawa Y, Hamaishi A (1986) Mate acquisition by a solitary crab Zebrida adamsii, a symbiont of the sea urchin. J Ethol 4:153–162
Zar JH (2010) Biostatistical analysis, 5th edn. Prentice-Hall, Upper Saddle River
Acknowledgements
The authors are grateful to Dr. Sinara Maria Moreira for her help during fieldwork and experimental observations and to Dr. Leandro de Sousa Souto for help with some of the statistical analyses. The authors are also grateful to the reviewers and Dr. Martin Thiel for providing valuable criticism that helped us to improve earlier versions of the present manuscript. All sampling in this study was conducted according to the applicable state and federal laws. Thanks to Sarah Steedman and John L. Ambrosio for correcting the English language in previous versions of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Funding
This study was not funded by any grant.
Conflict of interest
Author Douglas Fernandes Rodrigues Alves declares that he has no conflict of interest. Author Gustavo Luis Hirose declares that he has no conflict of interest. Author Samara de P. Barros-Alves declares that she has no conflict of interest. Author J. Antonio Baeza declares that he has no conflict of interest.
Ethical approval
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Additional information
Responsible Editor: M. Thiel.
Reviewed by S. M. Shuster and undisclosed experts.
Rights and permissions
About this article

Cite this article
Alves, D.F.R., Hirose, G.L., Barros-Alves, S.d. et al. The mating system of the symbiotic pea-crab Dissodactylus crinitichelis (Brachyura, Pinnotheridae): monogamy or promiscuity?. Mar Biol 164, 200 (2017). https://doi.org/10.1007/s00227-017-3234-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-017-3234-6