
Abstract
Where to place marine protected areas (MPAs) and how much area they should cover are some of the most basic questions when designing MPAs. Based on the theory of island biogeography, larger reserves are likely to protect more species and individuals but smaller reserves have been shown to positively influence populations. In this study, we assess a localised population of the ecologically and economically important southern rock lobster (Jasus edwardsii) inside and outside a small reserve. We used standardised fishery assessment trapping methods to sample J. edwardsii populations inside a reserve and an adjacent area outside the reserve. The population characteristics of the captured individuals were compared inside and outside the reserve using t tests (male size, female size, number of reproductive females, number of individuals and biomass), and we found that there were significantly greater numbers and larger individuals and biomass inside the reserve. However, many assessments of MPA effectiveness are confounded by differences in habitat. To account for possible differences in habitat, we collected multibeam bathymetry data to allow us to characterise seafloor structure and video data to assign each sampling location to a biotope class based on macroalgae assemblages. Then, using generalised linear models (GLMs), we assessed differences in populations while accounting for habitat. The GLMs revealed that there was still a significant difference in populations inside the reserve despite habitat differences inside and outside the reserve. We demonstrate a methodological approach to provide a baseline data set to assess MPA effectiveness through time and measure how habitat may respond to indirect consequences of fishing or other human impacts at the species or ecosystem level. We also highlight some of the limitations in sampling design and data availability common in MPA studies and resulting implications for assessment.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Marine protected areas (MPAs) continue to grow in popularity as anthropogenic impact on marine ecosystems persists (e.g. over-fishing, destructive fishing methods, pollution, coastal development; Halpern 2003). Although MPAs are popular tools for managing fisheries and/or preserving representative biodiversity (Allison et al. 1998; Lauck et al. 1998), decisions on their placement are usually part of social and political processes where biological effects are not always considered or known at inception (Jones et al. 1992; Agardy 1994). Regardless, where to place MPAs and how much area they should cover are some of the most basic questions when designing MPAs. Edgar et al. (2014) found that a combination of five key features (no take, well-enforced, old (>10 years), large (>100 km2), and isolated by sand or deep water) contributed to MPA success and positive trajectories of population recovery. Additionally, based on the theory of island biogeography, larger reserves are likely to protect more species (MacArthur and Wilson 1967) but there are still mixed results on how the size of an MPA impacts the recovery of heavily fished species. A review by Halpern (2003) showed that the impact of reserves was positive irrespective of size, and some MPA studies have found that there are minimal or insignificant correlations between size and effectiveness, depending on the aspect considered (Côté et al. 2001; Guidetti and Sala 2007). However, for an MPA of any size to be effective, it requires the incorporation of essential habitat to the species targeted for protection (Lee et al. 2015).
One of the potential benefits of MPAs is the ability to restore stocks of commercially fished species within their boundaries with the expectation of spillover to areas outside the protected area through movement of adults or as a source for larval dispersal (Harmelin-Vivien et al. 2008; Lester et al. 2009; Harrison et al. 2012). The effectiveness that MPAs provide to meet conservation goals depends on the level of protection legislated and enforced (Fitzsimons 2011). For example, globally it is estimated that only 6 % of MPAs are classified as strict ‘no-take’ nature reserves (Costello and Ballantine 2015). Considering that 60 % of these are less than 1 km2 (Costello and Ballantine 2015), local scale information is required to assess the conservation effectiveness and resource protection afforded by these small reserves (McLaren et al. 2015). There are examples of small MPAs having positive effects on the recovery of fishery-targeted species. A small marine reserve in China had a positive effect on recovery of sea urchin populations (Lau et al. 2011). In addition, Afonso et al. (2011) showed that groupers—an endangered species—had high site fidelity within a small marine reserve in the Azores, mid-north Atlantic, suggesting that small reserves can promote recovery of endangered fish species as long as the species has a high site fidelity and do not emigrate outside reserve boundaries. Therefore, as long as an MPA captures the home range or activity area of a species targeted for protection, biomass can increase inside the boundaries (Kramer and Chapman 1999) and result in a ‘reserve effect’ (Halpern and Warner 2003).
In 1998, Australia committed to establishing a system of MPAs within its marine jurisdiction after signing onto the Convention on Biological Diversity. These actions resulted in the creation of large MPAs offshore and a network of MPAs in coastal regions. The coastal MPAs were often restricted in size due to conflicting interests of stakeholders (Wescott 2006; Agardy et al. 2011; Kearney et al. 2012). Each state in Australia is responsible for developing MPAs in their jurisdiction, which is out to three nautical miles from the coastline (Wescott 2006). In 2002, the state of Victoria greatly increased its coverage of MPAs and now 5.3 % of the 8000 km2 state waters in Victoria, Australia, is currently fully protected in no-take reserves (Wescott 2006). These no-take areas were set up to maintain examples of Victoria’s biodiversity and associated ecological processes (Parks Victoria 2007). Although management of fisheries species is not an explicit goal of Victorian MPAs, there is potential that these protected areas are supplementing other fishery management actions by not allowing take within their boundaries. They also provide an opportunity to serve as benchmarks against which non-protected marine areas may be compared.
One fishery species potentially benefitting from MPAs is the southern rock lobster, Jasus edwardsii. Jasus edwardsii is a commercially, recreationally and traditionally important species throughout its range in Southern Australia and New Zealand (Booth 1997; Gardner et al. 2003). The southern rock lobster (J. edwardsii) fishery in Victoria, Australia is the second largest fishery and makes up 27 % of wild-caught value within the state. This fishery is managed through both effort controls on number of licences and pots and also through total allowable commercial catch, which has recently declined due to poor stock status (Punt et al. 2013). Although exploitation has been linked to declining stocks of J. edwardsii, environmental change, such as change in ocean temperatures or currents, can also affect larval size, growth rate (Bermudes and Ritar 2008) and recruitment (Ridgway 2007). With declining stocks of J. edwardsii, MPAs have the ability to maintain a residual biomass as biomass is reduced through fishing.
Previous studies on the effect of MPAs on J. edwardsii have shown variable responses to spatial protection indicating that supply-side dynamics may have a significant influence on recovery patterns (Freeman et al. 2012). Some studies showed increased size and density inside MPAs (Edgar and Barrett 1997; Kelly and MacDiarmid 2003; Barrett et al. 2009a), but other studies showed that unmanaged fishing effort displaced by closed areas can have negative effects on populations in locations left open to fishing (Gardner et al. 2000). A modelling study completed in Tasmania, Australia showed that the placement of large MPAs may have negative impacts on J. edwardsii populations unless allowable catch was reduced by at least as much as the amount displaced by MPA implementation (Haddon et al. 2002). Additionally, foraging ranges may impact the ability of a small MPA to protect populations if foraging ranges cross MPA boundaries. Jasus edwardsii also have an extensive larval stage (~2 years) metamorphosing to the post larval stage (puerulus), which settle on coastal shelf rocky reefs. (Morgan et al. 2013). Therefore, J. edwardsii would likely benefit from a network of MPAs that protects several local populations across their dispersal range so that there are a larger number of healthy source populations to replenish numbers along the geographic range of a network through larval dispersal and adult movement.
To determine whether there is a reserve effect, protection status needs to be decoupled from habitat availability when comparing areas inside and outside MPAs. However, one challenge in MPA assessment is often lack of habitat information at a scale relevant to MPAs (Young and Carr 2015) and their biodiversity. The development of seafloor mapping technologies such as multibeam sonar allows us to quantify variation in seascape characteristics in unprecedented detail (Brown et al. 2011). In addition, remote video techniques allow for the rapid assessment of benthic algal communities (Young et al. 2015). Combined, these physical and biotic data can provide an assessment of environment and habitat conditions that may affect the distribution of keystone species. They can also provide a useful baseline to track indirect consequences of habitat response to protection. A method of testing for reserve effects without confounding the results with differences in available habitat is to integrate protection status with mapping and monitoring of reef ecosystems into models such as generalised linear models (GLMs) or generalised additive models (GAMs). By incorporating habitat, these models enable testing of reserve effects while accounting for differences in habitat inside and outside reserves.
In this study, we conduct a case study to determine the effect of a small MPA on a localised population of J. edwardsii with the following objectives: (1) determine whether a small MPA is associated with increased J. edwardsii biomass or size of individuals; and (2) determine the effect of seafloor structure, biological habitat and distance from MPA on J. edwardsii count, size and reproductive condition. To meet these objectives, we used standardised fishery stock assessment methods to sample J. edwardsii populations inside and outside a small MPA. We also collected multibeam data to provide information on the structure of the seafloor and video data to classify biological benthic habitats. We then compared the populations inside and outside the MPA to see whether there were significant differences and used GLMs to incorporate benthic habitats into the inside/outside comparison.
Materials and methods
Study site
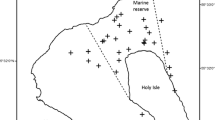
The study site for this project is a small MPA, the Merri Marine Sanctuary (MMS) and its adjacent waters. The MMS is located on the coast at Warrnambool, Southwest Victoria, Australia (Fig. 1). Established in November 2002, it covers 29 ha and has a depth range from 0 to 15 m (Barton et al. 2012). The seafloor within the MMS is dominated by shallow subtidal rocky reefs interspersed with sand patches (Monk et al. 2008). The MMS meets three of the five criteria outlined by Edgar et al. (2014) for successful MPAs [no take, well-enforced and old (>10 years)].

Study site for the project located off Warrnambool, Victoria, in south-eastern Australia. The light grey circles represent sample locations inside and outside the Merri Marine Sanctuary (outlined in black)
Jasus edwardsii survey
In February 2013, we used standardised fishery assessment trapping methods to sample J. edwardsii populations. Lobster pots were baited with 1 kg of locally available bait and escape gaps were wired shut (Woods and Edmunds 2013) as an extension to the Parks Victoria Subtidal Reef Monitoring Program. We deployed 40 spatially referenced pots within the MMS and 100 pots in areas surrounding the sanctuary over three nights in the MMS (n = 15, 15, 10) and two nights outside the MMS (n = 50, 50; Fig. 1) to limit the potential of temporal variability associated with catch. The sampling outside the MMS was part of the annual Victorian Department of Environment and Primary Industries fishery independent fixed site survey (Department of Primary Industries, 2009). The sampling inside the MMS was an adjunct to the Parks Victoria Victorian Subtidal Reef Monitoring Program.
Captured J. edwardsii were counted and sexed, females were assessed for reproductive condition, and all lobsters were measured for carapace length (CL). To calculate J. edwardsii biomass, we used the length–weight relationship provided in Punt (2003) and used by Woods and Edmunds (2013): W = aCL b, where W is the weight in kilograms, CL is carapace length and a and b are coefficients related to sex and size class (Females: a = 0.000271, b = 3.135; Males: a = 0.000285, b = 3.114).
Seafloor structure data
We collected multibeam sonar data in strips over each pot location to characterise the localised seafloor structure using a Kongsberg Maritime EM2040C in March–May 2014 (Fig. 2). In total, we collected 164 ha of data with 8 ha within the MMS. Once collected, these data were manually cleaned in CARIS HIPS and SIPS 8.1 using standard hydrographic data cleaning procedures, exported at 0.50-m resolution and brought them into IVS Fledermaus for interpolation. Once interpolated to fill small gaps in the 0.5-m resolution data, we converted the data to a digital elevation model (DEM) and brought the DEM into ArcGIS 10.1 for analysis.

Bathymetry data and derivatives used to characterise seafloor structure: a coverage of the multibeam sonar data (dark grey) in relation to the pot sampling locations (white) inside and outside the Merri Marine Sanctuary with extent of area used to show derived variables outlined in a rectangle, b depth, c slope, d rugosity, e bathymetric position index at 25 m, f curvature, g standard deviation of depth, h cosine of aspect (northing) and i sine of aspect (easting)
We used the DEM to derive a number of structure variables (using the ArcGIS software plugin Benthic Terrain Modeller 3.0; Wright et al. 2012): depth, slope, rugosity, bathymetric position, curvature, variation in depth, cosine of aspect and sine of aspect (Table 1; Fig. 2). We selected these indices for their known influence on distributions of biological assemblages (Ierodiaconou et al. 2007; Rattray et al. 2009) and their expected influence on J. edwardsii distribution. We then calculated the mean value of each index within a 40-m circular radius around each pot location using the Focal Statistics tool. Characterising seafloor structure at this range allowed to account for variation of seafloor structure characteristics around each pot location that could impact J. edwardsii movement.
Biotope characterisation
In addition to the physical structure of the seafloor, benthic algal communities can also be used as an index of environment and habitat conditions that may affect the distribution of J. edwardsii. To assess biogenic habitat, we used a delta vision HD underwater video camera at a 45° angle to survey the benthic habitat around each trap position. The video footage was classified into one or more biotopes in accordance with a modified JNCC scheme (Connor et al. 2004), established for classifying subtidal reef communities across the state of Victoria. The scheme is based on the floristic (seaweed) composition of the reef community and is hierarchical. Following the modified JNCC classification scheme, biotope complexes were determined according to major structural features; biotopes were identified according to suites of conspicuous species; and sub-biotopes were defined by less obvious differences in species composition but typically reflect more subtle geographic variations. There were 40 distinct biotopes and sub-biotopes recognised from the video ground truthing. The hierarchical scheme was used to pool these into 11 biotope complex classes for quantitative analysis (Table 2).
Comparison of Jasus edwardsii populations inside and outside reserve
Statistical tests for male and female size (average weight), female reproductive condition, number of individuals and total catch (biomass) per unit effort (CPUE, average catch per pot lift) were conducted. Separate t tests were run to compare male size, female size, female reproductive condition, count and CPUE inside and outside the MMS. To offset the problem of multiple comparisons, a Bonferroni correction was applied to the alpha level of 0.05 to reduce the occurrence of a Type I error (α = 0.05/5 = 0.01).
To determine whether seafloor structure and biogenic habitat contributes to J. edwardsii distribution inside and outside the MMS, GLMs were created using the glm function in R statistical software (R Development Core Team 2015). GLMs are flexible and appropriate for analysing ecological relationships (Austin 1987) because they do not force data into unnatural scales and allow for nonlinearity and non-constant variance structures in the data (Hastie and Tibshirani 1990, Guisan et al. 2002). Prior to running the GLMs, we tested for variable distribution using Cleveland dotplots and correlation between variables using Pearson’s correlation and variance inflation factors (VIFs). We tested for assumptions of the GLMs, including independence of observations (spatial autocorrelation). No variables required transformation, and only those variables with Pearson’s correlation coefficients less than 0.50 and VIF values less than five were included in the same model. Best fit GLMs were developed for each response variable (male size, female size, total biomass, count and female reproductive condition) using an iterative approach where variables were removed and/or replaced until the best fit model was developed. Model fit was determined using deviance explained, variable significance and AIC when available (e.g. when response variable was made up of integer values).
Results
Comparison of Jasus edwardsii populations inside and outside reserve
A total of 715 J. edwardsii were captured with 328 captured inside the MMS (40 pot lifts) and 387 in surrounding waters (100 pot lifts; Table 3). Overall, more females were captured in comparison with males and males had a greater range in size. Much of the female population was below the legal minimum length (LML), with a higher number of lobsters just above the LML inside the MMS when compared to outside (Fig. 3). The male population did not have a marked difference in abundance above and below the LML. The mode of the size frequency structure was above the LML inside the sanctuary and below the LML outside the sanctuary. The largest observed individuals were males inside the sanctuary. Outside the MMS, only 52 individuals of legal size were captured compared to the 158 captured inside the MMS despite less than half the effort. Legal sized individuals were more prevalent inside the MMS where 48 % were of legal size while only 13 % outside the MMS were of legal size.

Jasus edwardsii male and female size distributions (carapace length) inside and outside the Merri Marine Sanctuary (MMS) with the length of legal size for males and females displayed as a dashed black line in each distribution plot: a males inside the MMS, b females inside the MMS, c males outside the MMS and d females outside the MMS
The t tests used to test for differences inside and outside of the MMS on female and male size (average weight), female reproductive condition, number of individuals and total catch per unit effort (CPUE) measured as biomass showed that there were significant differences in each of these characteristics of the J. edwardsii population inside and outside the MMS. There was a significant difference in both female and male size with sizes larger inside the MMS compared to outside (female weight: t = 3.574, P value = 0.001; male weight: t = 3.988, P value < 0.001). There were significantly more reproductive females per pot lift inside the MMS compared to outside (t = 5.887, P value < 0.001). There was significantly higher abundance inside compared with outside the MMS, both in terms of counts of individuals per pot lift (t = 5.338, P value < 0.001) and biomass per pot lift (t = 6.283, P value < 0.001).
Results from the GLMs, applied to determine the effect of seafloor structure and biogenic habitat on the distribution of J. edwardsii, provided differing results for each of the population characteristics (Table 4). To ensure adequate sample sizes per group, we used biotope complex, rather than biotope as an explanatory variable (i.e. the reduced number of classes allowed for more observations per class). Each population characteristic was included in a model with and without biotope complex as a variable because the sample sizes were skewed by biotope complex. By running models with and without biotope complex, we were able to determine the effect of biotope complex on each population characteristic and whether or not inclusion of biotope complex altered relationships with other variables. The GLMs for male size found depth and distance to MMS to be the habitat variables important to explaining variation when biotope was not included in the model. When biotope complex was included, distance to MMS dropped out at both scales and was replaced by biotope complex. The largest males were found in biotope complex A (high-energy sublittoral rock Phyllospora communities; Fig. 4a). The total deviance explained ranged from 18.7 to 23.2 % with higher deviance explained in those models containing biotope complex as a variable (Table 4). The only variable that came up as a significant factor explaining variation in female size across all GLMs was distance to MMS. The deviance explained was relatively low for these models (pseudo-R 2 = 7.6 %; Table 4). Distance to MMS was the only variable found significant in the GLM for number of reproductive females when biotope was not included (pseudo-R 2 = 21.5 %). When biotope complex was included as a variable, it was also found significant and increased the deviance explained to 25.0 % (Table 4). Biotope Complex A was associated with the largest number of reproductive females (Fig. 4c). Results from the GLMs for the number of individuals per pot lift (count) varied with scale and inclusion of biotope. In all models, depth and distance to MMS were significant and some measure of seafloor complexity (slope or rugosity) was significant in all GLMs except for the GLM with no biotope complex. When biotope complex was included, it was significant and the greatest number of individuals was found in Biotope Complex A (Fig. 4d). The deviance explained for these models ranged from 23.1 to 30.2 % and those models with higher deviance explained contained biotope complex as a variable (Table 4). Finally, distance to MMS, depth and slope were significant in all the GLMs for average biomass per pot lift (CPUE). Also, when biotope was included in the GLM, it was found significant and, again, Biotope Complex A contained the greatest lobster biomass (Fig. 4e). These GLMs for biomass had the highest deviance explained of all the models ranging from 35.6 to 42.6 % (Table 4).

Box plots the Jasus edwardsii male average weight (a), female average weight (b), number of reproductive females (c), number of individuals (d) and average biomass (e) conditional on biotope complex. The width of the box is proportional to the number of observations per class and the circles represent outliers
Discussion
Overall, this study has shown that a small, no-take MPA (25 ha) can support a large population of J. edwardsii with increased size and number of individuals within its boundaries. The information from this study can also be used as a baseline for measuring reserve effects as population trajectories are traced through time. The differences in size and abundance were in agreement with other studies comparing J. edwardsii populations inside and outside MPAs in New Zealand (Kelly et al. 2000; Shears et al. 2006; Freeman et al. 2012) and Tasmania (Barrett et al. 2009a, b). Barrett et al. (2009b) showed an increase in population up to 250 % following MPA protection, with an increased abundance of large J. edwardsii individuals within reserves. These previous studies combined with this study strengthen the evidence that MPAs have the potential to benefit J. edwardsii populations in areas previously targeted by commercial fisheries. In addition, these areas serve to bolster the biomass of broodstock, potentially contributing to the sustainability and catch levels of the fishery.
Our study showed that size and abundance of J. edwardsii outside the MMS increased closer to the sanctuary boundaries, suggesting that the MMS may be supplying individuals to surrounding waters open to the fishery and enhancing catch numbers. Only 13 % of the population outside the MMS were at or above the legal size limit for commercial catch compared to the 48 % inside the MMS and there was a much steeper change in size frequencies at the legal size limit outside compared to inside the MMS. The observed size frequency pattern was consistent with the presence of fishing pressure outside the MMS.
Biotope complex was often the most important variable in the GLMs suggesting that macroalgae communities are important to the distribution and abundance of J. edwardsii. The presence of macroalgae on temperate reefs reduces predation risk, increases structural complexity and provides habitat for prey species (Villegas et al. 2008; Kovalenko et al. 2012). The results from this study suggest that Phyllospora-dominated communities are associated with larger numbers of and larger sized individuals of J. edwardsii. Phyllospora comosa is a large brown macroalgae that forms forests on shallow rocky reefs throughout south-eastern Australia and is commonly called ‘crayweed’ because ‘crayfish’ (spiny lobster; e.g. J. edwardsii) are often found in it. Additionally, Phyllospora-dominated areas were linked to higher abundance of sea urchins and abalone compared to other shallow subtidal habitats (Marzinelli et al. 2014), which are common prey for adult J. edwardsii (Edmunds 1995). It is also possible that the lobster–Phyllospora relationship is not a direct one, with both corresponding to aspects of wave exposure.
Although the Phyllospora biotope complex was mainly sampled within the boundaries of the MMS, male size was the only population characteristic where distance to MMS dropped out of the model when biotope was included. Therefore, the analyses indicated there was a relationship between lobster population parameters and distance from the reserve but that there was also a strong relationship of lobster parameters with habitat type. Both variables are likely to influence J. edwardsii, but without sampling biotopes at varying distances from the sanctuary, we cannot determine the relative importance of each. However, this study shows the importance of assessing multiple aspects of habitat when conducting analyses on the effectiveness of MPAs. Marine protected areas are often established across heterogeneous habitat features, and without accounting for differences in habitat when determining the effectiveness of a reserve, reserve effects cannot be decoupled from natural variability (Claudet and Guidetti 2010).
Jasus edwardsii are obligate crevice dwellers and are found on all rock types and geomorphological structures in the 1- to 200-m depth range, provided there is suitable shelter (Booth 1997; MacDiarmid et al. 1991; Edmunds 1995). They reside in ‘dens’ within crevices or under ledges formed by the reefs and are important reef predators that forage on slow-moving benthic invertebrate prey such as ophiuroids, bivalves, sea urchins and abalone (Jernakoff et al. 1987; Edmunds 1995). Although crevices and shelter are often associated with more complex reef habitat, we recognise that our methods did not measure differences in shelter availability directly. We assume that our substratum complexity indices are related in some way to lobster shelter and foraging habitats and recognise that these analyses could be improved in the future through the inclusion of more direct geomorphological description. In this study, we used different characteristics of the seafloor (e.g. slope, rugosity) as proxies for habitat that may support crevice habitat. We note that lobster crevice habitat can occur on non-complex substratum structures, but the indices used in this study provide a better indication of crevice habitat than no access to seafloor data.
Although the results from this study suggest that this localised population of J. edwardsii are benefitting from this small MPA, there are limitations in the study design. First of all, there is no adequate data on this population before implementation of the MPA and, therefore, we have no conclusive knowledge signifying whether the larger population within the MMS is due to historical distribution or whether the population is responding to a removal of fishing pressure. Additionally, we only evaluated the effect of one MPA in this pilot study. To determine how well MPAs are contributing to the recovery of J. edwardsii populations, more MPAs need to be examined for both lobster populations and habitat characterisations over longer time periods. Then, these data could be used to validate population models already used to assess recovery of J. edwardsii through time (e.g. Hobday et al. 2005). Variation among reserves, both in size and habitat within their boundaries, is known (Halpern 2003) and may result in different effects on J. edwardsii population characteristics. The methods we applied in this study could help to account for variation in habitat across MPAs to find whether there is a reserve effect on J. edwardsii populations. As density-dependent spillover of species biomass into waters adjacent to MPAs has been recorded worldwide (McClanahan and Mangi 2000; Abesamis and Russ 2005; Goñi et al. 2008), the larval export of J. edwardsii from the MMS may contribute to populations outside the study area. Jasus edwardsii have a complex early life cycle and can spend 12–24 months within oceanic waterbodies undergoing 11 larval stages before settling (Thomas et al. 2000; Linnane et al. 2014). Previous studies have suggested that Australian J. edwardsii populations are responsible for some trans-Tasman larval flow that contributes to and possibly maintains New Zealand populations (Chiswell et al. 2003; Morgan et al. 2013).
Another limitation of this study was data availability. Available fisheries monitoring data outside the MMS were limited to the east of the sanctuary. A complete evaluation of the presence or absence of spillover from the MMS to surrounding fished waters, and a better understanding of the confounding of habitat structure, would be assisted by data in all directions away from the MPA. For example, habitat structure to the west of the MMS has high complexity rocky reef within a depth range amenable to Phyllospora comosa and may provide more suitable lobster habitat. Additionally, a balanced design could help to understand whether there is simply a west–east decrease in population and individual size or whether there is more evidence for positive MPA effects. Finally, the sampling only captured a snapshot of J. edwardsii populations over a short period. A better understanding of the effect of the MMS could be gained through repeat, temporal sampling. Tag-recapture data would greatly assist in understanding lobster residency, migration and any net ‘spillover’ effect.
Overall, we found that, the small MPA used in this study, the Merri Marine Sanctuary has a very high J. edwardsii population density relative to the surrounding fished area and that this higher density was not fully explained by habitat differences. However, the lack of data on J. edwardsii populations before MPA implementation requires caution when analysing these results. These findings serve as a baseline data set for use in assessing how the population responds to protection through time. Continued sampling of this MPA into the future, along with other MPAs, will provide us with a more conclusive understanding of how J. edwardsii are responding to complete removal of fishing pressure within MPA boundaries.
Despite its shortcomings, this study shows the potential secondary benefits of Victoria’s MPAs for rock lobster populations. Our study complements findings from McLeod et al. (2008), which suggested that small scale MPAs can be successful in protecting critical habitat of vulnerable species. The evidence for potential enhancement of the fishery directly outside the boundaries of the MMS can also help garner support for protected areas by local communities, as successful implementation of MPAs requires support by fisheries communities. Fishing communities that cannot directly perceive benefits of MPAs are less likely to support them as a management tool (Russ et al. 2004). This is compounded by the fact that there may be a temporal scale of decades before being able to demonstrate measurable benefits of protection (Micheli et al. 2004). The impediment of small MPAs on local fisheries is much less when compared to large MPAs, making them more favourable to fishing communities. However, even small MPAs can cause conflict when there is a loss of traditional fishing grounds or previously favoured locations through protection. Embracing local knowledge through utilisation of local fishers and techniques in MPA assessment may generate a sense of ownership among stakeholders (Voyer et al. 2014), especially if contributing to wider ecosystem-based spatial planning such as accounting for breeding biomass in MPAs in fishery management models. As long as a small MPA is placed in a location beneficial to species’ life cycles (e.g. critical habitat, spawning region), they have the potential to support population recovery, especially when part of a larger network of MPAs.
As MPAs are increasing in application as a conservation tool, methods to determine their effectiveness are necessary. Our methods utilising both physical and biological components of the marine landscape allowed us to potentially decouple any differences in habitat from a reserve effect. Mapping of habitats is an important step in determining whether MPA effects are going to be confounded by differences in suitable habitat. The methods presented in this study allow for variations in habitat when testing for reserve effects and also provide a baseline data set to determine whether habitat changes over time with protection status. With the incorporation of these methods over time, we can begin to more fully understand the effect of MPAs on important fishery species.
References
Abesamis RA, Russ GR (2005) Density-dependent spillover from a marine reserve: long-term evidence. Ecol Appl 15:1798–1812
Afonso P, Fontes J, Santos RS (2011) Small marine reserves can offer long term protection to an endangered fish. Biol Conserv 144:2739–2744
Agardy TE (1994) The science of conservation in the coastal zone: new insight on how to design, implement, and monitor marine protected areas. A marine conservation and development report. IUCN, Gland
Agardy T, Di sciara GN, Christie P (2011) Mind the gap: addressing the shortcomings of marine protected areas through large scale marine spatial planning. Mar Policy 35:226–232
Allison GW, Lubchenco J, Carr MH (1998) Marine reserves are necessary but not sufficient for marine conservation. Ecol Appl 8(1):S79–S92
Austin M (1987) Models for the analysis of species’ response to environmental gradients. Veg Res 69:35–45
Barrett N, Buxton C, Gardner C (2009a) Rock lobster movement patterns and population structure within a Tasmanian marine protected area inform fishery and conservation management. Mar Freshw Res 60:417–425
Barrett NS, Buxton CD, Edgar GJ (2009b) Changes in invertebrate and macroalgal populations in Tasmanian marine reserves in the decade following protection. J Exp Mar Biol Ecol 370:104–119
Barton J, Pope A, Howe S (2012) Marine natural values study Vol 2: Marine Protected Areas of the Central Victoria Bioregion. Parks Victoria Technical Series No. 76. Parks Victoria, Melbourne
Bermudes M, Ritar A (2008) Response of early stage spiny lobster Jasus edwardsii phyllosoma larvae to changes in temperature and photoperiod. Aquaculture 282:63–69
Booth JD (1997) Long-distance movements in Jasus spp. and their role in larval recruitment. Bull Mar Sci 61(1):111–128
Brown CJ, Smith SJ, Lawton P, Anderson JT (2011) Benthic habitat mapping: a review of progress towards improved understanding of the spatial ecology of the seafloor using acoustic techniques. Estuar Coast Shelf Sci 92:502–520
Chiswell SM, Wilkin J, Booth JD, Stanton B (2003) Trans-Tasman Sea larval transport: is Australia a source for New Zealand rock lobsters? Mar Ecol Prog Ser 247:173–182
Claudet J, Guidetti P (2010) Improving assessments of marine protected areas. Aquat Conserv 20:239–242
Connor DW, Allen JH, Golding N, Howell KL, Lieberknecht LM, Northen KO, Reker JB (2004) The marine habitat classification for Britain and Ireland. Version 04.05
Costello MJ, Ballantine B (2015) Biodiversity conservation should focus on no-take Marine Reserves: 94% of Marine Protected Areas allow fishing. Trends Ecol Evol 30(9):507–509
Côté IM, Mosqueira I, Reynolds JD (2001) Effects of marine reserve characteristics of fish populations: a meta-analysis. J Fish Biol 59:178–189
Department of Primary Industries (2009) Victorian rock lobster fishery management plan 2009. Fisheries Victoria management report series no. 70
Development Core Team R (2015) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Edgar GJ, Barrett NS (1997) Short term monitoring of biotic change in Tasmanian marine reserves. J Exp Mar Biol Ecol 213:261–279
Edgar GJ, Stuart-Smith RD, Willis TJ, Kininmonth S, Baker SC, Banks S, Barrett NS, Becerro MA, Bernard ATF, Berkhout J, Buxton CD, Campbell SJ, Cooper AT, Davey M, Edgar SC, Försterra G, Galván DE, Irigoyen AJ, Kushner DJ, Moura R, Parnell PE, Shears NT, Soler G, Strain EMA, Thomson FJ (2014) Global conservation outcomes depend on marine protected areas with five key features. Nature 506:216–220
Edmunds M (1995) The ecology of the juvenile southern rock lobster, Jasus edwardsii (Hutton, 1875) (Palinuridae). PhD dissertation, University of Tasmania, Tasmania, Australia
Fitzsimons JA (2011) Mislabelling marine protected areas and why it matters: a case study of Australia. Conserv Lett 4:340–345
Freeman DJ, MacDiarmid AB, Taylor RB, Davidson RJ, Grace RV, Haggitt TR, Kelly S, Shears NT (2012) Trajectories of spiny lobster Jasus edwardsii recovery in New Zealand marine reserves: is settlement a driver? Environ Conserv 39(3):295–304
Gardner C, Frusher S, Ibbott S (2000) Preliminary modelling of the effect of marine reserves on the catch, egg production, and biomass of rock lobsters in Tasmania. Tasmanian Aquaculture and Fisheries Institute Technical Report 12. p 36
Gardner C, Frusher S, Haddon M, Buxton C (2003) Movements of the southern rock lobster Jasus edwardsii in Tasmania, Australia. Bull Mar Sci 73(3):653–671
Goñi R, Adlesrstein S, Alvarez-Berastegui D, Forcada A, Reñones O, Criquet G, Polti S, Cadiou G, Valle C, Lenfant P (2008) Spillover from six western Mediterranean marine protected areas: evidence from artisanal fisheries. Mar Ecol Prog Ser 366:159–174
Guidetti P, Sala E (2007) Community-wide effects of marine reserves in the Mediterranean Sea. Mar Ecol Prog Ser 335:43–56
Guisan A, Edwards TC Jr, Hastie T (2002) Generalized linear and generalized additive models in studies of species distributions: setting the scene. Ecol Model 157:89–100
Halpern BS (2003) The impact of marine reserves: do reserves work and does reserve size matter? Ecol Appl 13(1):S117–S137
Halpern BS, Warner RR (2003) Matching marine reserve design to reserve objectives. Proc R Soc B Biol Sci 270(1527):1871–1878
Haddon M, Buxton CD, Gardner C, Barrett NS (2002) Modelling the effect of introducing MPAs in a commercial fishery: a rock lobster example. Aquatic protected areas - What works best and how do we know? In: Proceedings of the world congress on aquatic protected areas cairns, Australia, August 2002, pp 428–436
Harmelin-Vivien M, Le Diréach L, Bayle-Sempere J, Charbonnel E, García-Charton JA, Ody D, Pérez-Ruzafae A, Reñones O, Sánchez-Jerezc P, Valle C (2008) Gradients of abundance and biomass across reserve boundaries in six Mediterranean marine protected areas: evidence of fish spillover? Biol Conserv 141:1829–1839
Harrison HB, Williamson DH, Evans RD, Almany GR, Thorrold SR, Russ GR, Feldheim KA, van Herwerden L, Planes S, Srinivasan M, Berumen ML, Jones GP (2012) Larval export from marine reserves and the recruitment benefit for fish and fisheries. Curr Biol 22:1023–1028
Hastie TJ, Tibshirani RJ (1990) Generalized additive models. CRC Press, Boca Raton
Hobday D, Punt AE, Smith DC (2005) Modelling the effects of Marine Protected Areas (MPAs) on the southern rock lobster (Jasus edwardsii) fishery of Victoria, Australia. N Z J Mar Freshw 39:675–686
Ierodiaconou D, Laurenson L, Burq S, Reston M (2007) Marine benthic habitat mapping using multibeam data, georeferenced video and image classification techniques in Victoria, Australia. J Spat Sci 52:93–104
Jernakoff P, Phillips B, Maller R (1987) A quantitative study of nocturnal foraging distances of the western rock lobster Panulirus Cygnus George. J Exp Mar Biol Ecol 113:9–21
Jones GP, Cole RC, Battershill CN (1992) Marine reserves: Do they work? In: P second international temperate reef symposium, pp 29–45
Kearney R, Buxton CD, Farebrother G (2012) Australia’s no-take marine protected areas: appropriate conservation or inappropriate management of fishing? Mar Policy 36:1064–1071
Kelly S, MacDiarmid AB (2003) Movement patterns of mature spiny lobsters, Jasus edwardsii, from a marine reserve. N Z J Mar Freshw 37(1):149–158
Kelly S, Scott D, MacDiarmid AB, Babcock RC (2000) Spiny lobster, Jasus edwardsii, recovery in New Zealand marine reserves. Biol Conserv 92:359–369
Kovalenko KE, Thomaz SM, Warfe DM (2012) Habitat complexity: approaches and future directions. Hydrobiologia 685:1–17
Kramer DL, Chapman MR (1999) Implications of fish home range size and relocation for marine reserve function. Enviro Biol Fish 55(1–2):65–79
Lau DCC, Dumont CP, Lui GCS, Qiu J-W (2011) Effectiveness of a small marine reserve in southern China in protecting the harvested sea urchin Anthocidaris crassipina: a mark-and-recapture study. Biol Conserv 144:2674–2683
Lauck T, Clark CW, Mangel M, Munro GR (1998) Implementing the precautionary principle in fisheries management through marine reserves. Ecol Appl 8(1):S72–S78
Lee KA, Huveneers C, MacDonald T, Harcourt RG (2015) Size isn’t everything: movements, home range, and habitat preferences of eastern blue gropers (Achoerodus viridis) demonstrate the efficacy of a small marine reserve. Aqua Conserv 25:174–186
Lester SE, Halpern BS, Grorud-Colvert K, Lubchenco J, Ruttenberg BI, Gaines SD, Airame S, Warner RR (2009) Biological effects with no-take marine reserve: a global synthesis. Mar Ecol Prog Ser 384:33–46
Linnane A, McGarvey R, Gardner C, Walker TI, Matthews J, Green B, Punt AE (2014) Large-scale patterns in puerulus settlement and links to fishery recruitment in the southern rock lobster (Jasus edwardsii), across south-eastern Australia. ICES J Mar Sci 71(3):528–536
MacArthur RH, Wilson EO (1967) The theory of island biogeography. Princeton University Press, Princeton
MacDiarmid AB, Hickey B, Maller RA (1991) Daily movement patterns of the spiny lobster Jasus edwardsii (Hutton) on a shallow reef in northern New Zealand. J Exp Mar Biol Ecol 147:185–205
Marzinelli EM, Campbell AH, Vergés A, Coleman MA, Kelaher BP, Steinberg PD (2014) Restoring seaweeds: does the declining fucoid Phyllospora comosa support different biodiversity than other habitats? J Appl Phycol 26:1089–1096
McClanahan TR, Mangi S (2000) Spillover of exploitable fishes from a marine park and its effect on the adjacent fishery. Ecol Appl 10:1792–1805
McLaren BW, Langlois TJ, Harvey ES, Shortland-Jones H, Stevens R (2015) A small no-take marine sanctuary provides consistent protection for small-bodied by-catch species, but not for large-bodied, high-risk species. J Exp Mar Biol Ecol 471:153–163
McLeod E, Salm R, Green A, Almany J (2008) Designing marine protected area networks to address the impacts of climate change. Front Ecol Environ 7:362–370
Micheli F, Halpern B, Botsford LW, Warner RR (2004) Trajectories and correlates of community change in no-take marine reserves. Ecol Appl 14:1709–1723
Monk J, Ierodiaconou D, Bellgrove A, Laurenson L (2008) Using community-based monitoring with GIS to create habitat maps for a marine protected area in Australia. J Mar Biol Assoc UK 88(5):865–872
Morgan EMJ, Green BS, Murphy NP, Strugnell JM (2013) Investigation of genetic structure between deep and shallow populations of the southern rock lobster, Jasus edwardsii in Tasmania, Australia. PLoS One 8(10):e77978. doi:10.1371/journal.pone.0077978
Punt AE (2003) The performance of a size structured stock assessment method in the face of spatial heterogeneity in growth. Fish Res 65:391–409
Punt AE, Trinnie F, Walker TI, McGarvey R, Feenstra J, Linnane A, Hartmann K (2013) The performance of a management procedure for rock lobsters, Jasus edwardsii, off western Victoria, Australia in the face of non-stationary dynamics. Fish Res 137:116–128
Rattray A, Ierodiaconou D, Laurenson L, Burq S, Reston M (2009) Hydro-acoustic remote sensing of benthic biological communities on the shallow south east Australian continental shelf. Estuar Coast Shelf S 84:237–245
Ridgway K (2007) Long-term trend and decadal variability of the southward penetration of the East Australian Current. Geophys Res Lett 34:L13613. doi:10.1029/2007GL030393
Russ GR, Alcala AC, Maypa AP, Calumpong HP, White AT (2004) Marine reserve benefits local fisheries. Ecol Appl 14(2):597–606
Shears NT, Grace RV, Usmar NR, Kerr V, Babcock RC (2006) Long-term trends in lobster populations in a partially protected vs. no-take marine park. Biol Conserv 132:222–231
Thomas CW, Crear BJ, Hart PR (2000) The effect of temperature on survival, growth, feeding and metabolic activity of the southern rock lobster, Jasus edwardsii. Aquaculture 185:73–84
Victoria Parks (2007) Parks Victoria marine research and monitoring strategy 2007–2012. Author: Anthony Boxshall, Parks Victoria
Villegas MJ, Laudien J, Sielfeld W, Arntz WE (2008) Macrocystis integrifolia and Lessonia trabeculata (Laminariales; Phaeophyceae) kelp habitat structures and associated macrobenthic community off northern Chile. Helgoland Mar Res 62:S33–S43
Voyer M, Gladstone W, Goodall H (2014) Understanding marine park opposition: the relationship between social impacts, environmental knowledge and motivation to fish. Aquat Conserv Mar Freshw Ecosyst 24:441–462
Wescott G (2006) The long and winding road: The development of a comprehensive, adequate and representative system of highly protected marine protected areas in Victoria, Australia. Coast Manage 49:905–922
Woods B, Edmunds M (2013) Victorian subtidal reef monitoring program: the reef biota at Merri Marine Sanctuary, February 2013. Parks Victoria Technical Series No. 87. Parks Victoria, Melbourne
Wright D, Pendleton M, Boulware J, Walbridge S, Gerlt B, Eslinger D, Sampson D, Huntly E (2012) ArcGIS Benthic Terrain Modeller (BTM) v. 3.0. Environmental Systems Research Institute, NOAA coastal services centre, Massachusetts office of coastal zone management. http://esriurl.com/5754
Young M, Carr M (2015) Assessment of habitat representation across a network of marine protected areas with implications for the spatial design of monitoring. PLoS One 10(3):e0116200
Young M, Ierodiaconou D, Womersley T (2015) Predicting the abundance of kelp forests on temperate reefs. Remote Sens Environ 170:178–187
Acknowledgments
We thank Parks Victoria for funding the capture of the multibeam sonar and southern rock lobster data used in this study within the Merri Marine Sanctuary. We also acknowledge the Pozible Project- Voyages of discovery and Somers Carroll Productions for funding the multibeam sonar and video data collection. Data analysis was supported by project 2015-025 funded by the Fisheries Research and Development Corporation (FRDC) awarded to DI and MY. We thank members of the crew S. Blake and A. Pope of Deakin University’s research vessel Yolla for assistance in the collection of the Multibeam sonar and video data. We thank the Victorian Department of Environment and Primary Industries (DEPI) fishery-independent fixed site survey programme for access to concurrent sample data for southern rock lobster populations outside the sanctuary. GIS laboratory facilities at Deakin University, Warrnambool, Victoria were used for spatial analyses. Ethics approval for this study was provided by the Department of Primary Industries Wildlife and Small Institutions Animal Ethics Committee (application 02.13). All animals collected in this study were collected using a Victorian Fisheries permit (#10006648).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: S. Connell.
Reviewed by A. Verges and an undisclosed expert.
Rights and permissions
About this article

Cite this article
Young, M.A., Ierodiaconou, D., Edmunds, M. et al. Accounting for habitat and seafloor structure characteristics on southern rock lobster (Jasus edwardsii) assessment in a small marine reserve. Mar Biol 163, 141 (2016). https://doi.org/10.1007/s00227-016-2914-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-016-2914-y