
Abstract
Remarkably little is known regarding the spawning area, spawning ecology or life history after recruitment of tropical eels to coastal areas across the Indo-Pacific region. In contrast, eel species in temperate areas, including the American, Australasian, European and Japanese eels, are known to have well-defined spawning and recruitment seasons, and spawning migrations generally take place during autumn months. Using monthly collections and analyses of both gonadosomatic index and gonad histology, we determined that the final stage of maturation for seaward spawning migration occurred throughout the year and year-round spawning and recruitment were evident in the tropical coastal areas from the open ocean. This non-seasonal spawning ecology is notably different from that of temperate catadromous eels. The unique year-round spawning ecology of Anguilla bicolor bicolor may facilitate mating between adult eels from different year classes (cohorts) or from different habitats.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Catadromous eels of the genus Anguilla Schrank, 1798, are semelparous fish with long spawning migrations and extended larval durations (Tesch 2003). These eels are classified into 19 species (Ege 1939; Castle and Williamson 1974; Arai et al. 1999; Watanabe et al. 2013). Among these species, 13 are tropical and six are temperate species. In general, temperate eels are distributed mainly in subtropical/temperate areas, whereas tropical eels are distributed mainly in tropical/subtropical regions. Despite the different biogeographical distributions of these two types of eels, all anguillids spawn in the tropical/subtropical ocean and have leptocephalus larvae that disperse via ocean currents from their oceanic spawning area to continental habitats, where they metamorphose into glass eels (Tesch 2003).
After growing for years in rivers, estuaries and coastal waters, eels return to their origin to spawn and subsequently die. Temperate eels mostly have well-defined spawning and recruitment seasons, long larval durations and panmictic populations. The recruitment period in tropical eels differs depending on species and/or site. A. marmorata, A. bicolor bicolor and A. mossambica were present 5 months in the year in Réunion Island, Mascarene Islands, western Indian Ocean (Robinet et al. 2003). In contrast, tropical eels of A. celebesensis, A. marmorata and A. bicolor pacifica had year-round recruitment to their habitats in North Sulawesi, Indonesia, western Pacific Ocean (Arai et al. 1999, 2001; Sugeha et al. 2001). The spawning period of tropical eels is still unknown. Additionally, several tropical eel species are composed of multiple populations, defined by their use of different spawning areas (Arai et al. 2003, 2014; Minegishi et al. 2008). These findings suggest that characteristics of both life history and ecology differ between temperate and tropical eels.
Ecological studies examining the migration and early life history of anguillid eels have been conducted mainly on temperate eels due to the high demand for these species, particularly European and Japanese eels, as a fisheries resource. The spawning location and season have been described for temperate eels Anguilla anguilla and A. rostrata (Schmidt 1922) and A. japonica (Tsukamoto 1992), and there have been several studies investigating the early life history and recruitment of these species. The spawning migration of temperate eels generally takes place during autumn months, between August and December (Kotake et al. 2007; Acou et al. 2008; Verreault et al. 2012) in the Northern Hemisphere and between February and May in the Southern Hemisphere (Todd 1981). Further, favorable environmental conditions to start their migration are also well studied. The exact timing depends on both internal (morphological, endocrine and metabolic) and environmental factors operating across a wide range of time scales, from day (e.g., precipitation, wind, atmospheric depression and sunshine hours) to month (e.g., lunar phase) and even season (e.g., water flow and level, water temperature, turbidity and photoperiod) (Acou et al. 2008; Breukelaar et al. 2009). However, no study to date has confirmed the timing and season of seaward migration for spawning in tropical eel species.
In this study, we aimed to understand the spawning timing and migration in a tropical catadromous eel Anguilla bicolor bicolor using monthly captured specimens. The timing of maturation and seaward migration for spawning was examined by investigating fluctuations in both the gonadosomatic index and gonadal histology. This study provides the first evidence of year-round spawning in a catadromous eel species.
Materials and methods
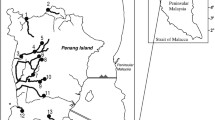
A total of 438 Anguilla bicolor bicolor were collected in Segara Anakan, central Java Island, Indonesia (approximately 7°35′–7°48′S, 108°46–109°03′E), from December 2007 to July 2009 (Fig. 1). All eels were purchased, postmortem, from local fishermen. To examine temporal variability of maturation, eels were collected in December 2007 and year-round from May 2008 to July 2009. The eels were collected using angling and bamboo traps at night. The Segara Anakan consists of a lagoon isolated from the Indian Ocean by Nusakambangan Island (Fig. 1) and comprises a mangrove forest area, a settlement area, and rice field areas. This lagoon is the only major estuarine-mangrove area in the central part of Java. This area experiences two seasons, the rainy or wet season, ranging from November to April, and the dry season, ranging from July to September. This equatorial climate experiences an average high monthly temperature between 28.0 °C (dry season) and 31.3 °C (wet season) (White et al. 1989).

Map showing the location of Segara Anakan, central Java of Indonesia. The approximate locations where eels were collected in Segara Anakan are shown with small stars
After eels were collected, biological parameters, such as total length and body weight, were measured. The sex of each eel was determined by visual and histological observations of the gonads. Whole gonad weight was measured, and gonadosomatic index (GSI; percent relative gonad weight to body weight) was subsequently calculated.
Fragments from the middle region of one gonad were fixed in formalin for histological analysis. Tissue fragments were prepared for resin and routine paraffin embedding. Resin blocks were processed routinely, and paraffin blocks were sectioned at 5 µm and stained with hematoxylin-eosin for histological observation.
Results
Eels were observed to occur throughout the year, all eels were identified as Anguilla bicolor bicolor, and all eels were determined to be female (i.e., no male was collected).
The GSI ranged from 0.0 to 4.5, and the values were highly variable between months (Fig. 2a). Gonadal histology by transverse sections revealed that oocyte characteristics of each maturation stage also varied within months from Stage I (primary stage) to V (final stage for spawning) (Fig. 2b). During Stage I at the early stage of oogenesis, the gonads were composed of primary germ cells, oogonia, lamellae and primary oocytes. During Stage II, immature cells developed with primary oocytes and few oil droplets. The early maturation stage of Stage III exhibited oocytes with cortical alveoli and oil droplets in the cytoplasm (Fig. 3). The early vitellogenic stage of Stage IV exhibited vitellogenic oocytes, nucleus, yolk granules and central yolk platelets, and the oocyte diameter increased drastically and the cytoplasm filled with yolk granules (Fig. 3). During the final stage of maturation, Stage V, midvitellogenic oocytes developed in final preparation for spawning; disintegrated nuclei were observed and whole nuclei were no longer visible (Fig. 3). Eels in more developed stages, such as Stages IV and V, were found every month, although the frequency differed among months (Fig. 2b). These findings suggest that A. bicolor bicolor carries out seaward migration for spawning throughout the year.

Monthly fluctuation of gonadosomatic index (a) and maturation stage (b) in a tropical catadromous eel Anguilla bicolor bicolor collected in Segara Anakan, central Java of Indonesia, between December 2007 and July 2009. ns no study for eel sampling was conducted that month, ne study was conducted, but no eels were collected

Gonadal histology of Anguilla bicolor bicolor collected in Segara Anakan, central Java of Indonesia, between December 2007 and July 2009. Stages correspond to a growth phase (Stages I and II), a pre-migrant phase (III) and two migrating phases (IV and V) in accordance with Durif et al. (2005). Each stage was modified according to Todd (1974) and Lokman et al. (1998). Each scale bar is 100 µm
The mean GSI values of each stage of I, II, III, IV and V were 0.40, 0.55, 1.03, 1.93 and 2.47, respectively (Table 1). Significant differences were found GSI values between the each stage in all combinations (Fig. 4) (Kruskal–Wallis test, p < 0.05–0.0001). A significant positive correlation was found between maturation stage and GSI value (Fig. 4) (Fisher’s Z-Transformation, p < 0.0001).

Relationship between gonadosomatic index and maturation stage for a tropical eel Anguilla bicolor bicolor that was collected in Segara Anakan, central Java of Indonesia, between December 2007 and July 2009
Discussion
This study is the first description of a spawning period of a tropical eel that extends throughout the year, as revealed by gonadal development and histology. Until recently, considerably less research has examined the spawning migration of tropical eels in comparison with that of temperate eel species. The findings described in this report indicate that tropical eels have life history characteristics that differ markedly from those of temperate eels. Temperate anguillid species make their spawning migration as silver eels during the fall and winter. Silver eels of the Japanese eel Anguilla japonica (Matsui 1957, 1972), the American eel A. rostrata (Verreault et al. 2012), the European eel A. anguilla (Tesch 2003; Vøllestad et al. 1986; Durif et al. 2005, 2009), and the Australian and New Zealand eels A. australis and A. dieffenbachii (Todd 1981; Sloane 1984; Jellyman 1987) all descend freshwater rivers and streams in the fall and enter saltwater, where they begin their marine migration to the open ocean spawning areas. The difference in spawning season duration and timing between tropical and temperate species could be attributed to differences in the seaward migration seasons of maturing adult eels. Analyses of otolith microstructure showed that the age at recruitment of tropical eels was constant throughout the year in A. celebesensis, A. marmorata and A. bicolor pacifica (Arai et al. 2001). The year-round spawning migration of tropical species and constant larval growth extend the period of recruitment to estuarine habitats to year-round in tropical eels. The spawning seasons of tropical eels were found to extend throughout the year by back calculation of otolith daily increment in A. celebesensis, A. marmorata and A. bicolor pacifica in North Sulawesi of Indonesia (Arai et al. 2001) which is close to central Java Island of Indonesia. Additionally, the present results support a year-round spawning period, as evidenced by the monthly occurrence of matured eels in A. bicolor bicolor. However, the recruitment period in tropical eels differs depending on species and/or site. A. marmorata, A. bicolor bicolor and A. mossambica were present 5 months in the year in Réunion Island of western Indian Ocean (Robinet et al. 2003). The periods of spawning in tropical eel recruits to Réunion Island were not year-round (Robinet et al. 2003). Therefore, further study is needed to accumulate more information regarding spawning period in tropical eels.
There remains only limited knowledge of the level of maturation of tropical eels as they begin their spawning migration. Downstream migrating female European eels typically have GSI values of >1.2 (Vøllestad and Jonsson 1986; Durif et al. 2005) but lower than 3.0 (Svedäng and Wickström 1997; Durif et al. 2005). The level of female maturation of Japanese eels as they begin their spawning migration in coastal areas ranged from 1.0 to 4.0 in GSI, and the levels were similar to those collected in the East China Sea and other Japanese coastal areas (Sasai et al. 2001; Kotake et al. 2007). GSI values of Anguilla celebesensis collected in the Poso Lake of Indonesia were greater than 9.0 (11.2 in maximum value) and in the range of GSI values characteristic of spawning conditions in A. japonica and A. marmorata, despite being collected inland in a freshwater lake far from the ocean (Arai 2014). GSI values of A. marmorata collected from same area were also high; all GSI values were greater than 4.0 (6.4 in maximum value). In the present study, GSI value of A. bicolor bicolor increased with advancing of maturation stage (Fig. 4), and it exhibited 2.5 on average with 4.5 at the maximum in Stage V (Table 1) even though the eels were collected from coastal lagoons and not from the open ocean. Thus, these results indicate that tropical eels in the silver phase at the onset of seaward spawning migration have comparatively higher maximum GSI values than those of temperate eels.
Downstream migration of tropical eels has not been properly investigated. Factors affecting downstream migration of temperate silver eels are highly variable both within localities and years and between localities (Vøllestad et al. 1994). However, it is widely recognized that water discharge greatly influences the downstream migration of European (Frost 1950; Lowe 1952; Deelder 1954, 1970; Vøllestad et al. 1986; Jonsson 1991), American (Smith and Saunders 1955) and New Zealand (Todd 1981; Boubée et al. 2001) silver eels. A long-term effect of water temperature on downstream migration based on 10 years of continuous observation in a Nordic river suggested that downstream migration started earlier in years with a lower mean temperature between 9 and 11 °C (Vøllestad et al. 1986). Water temperature might act as a trigger for downstream migration. However, these triggers might not apply to Anguilla bicolor bicolor in central Java Island of Indonesia. Water discharge is quite different between the dry and wet season in central Java Island of Indonesia because the average monthly rainfall ranges from 100 to 180 mm during the dry season and from 180 to 400 mm during the wet season (White et al. 1989). However, downstream migration of A. bicolor bicolor was found to extend throughout the year (Fig. 2). Furthermore, the water temperature does not fluctuate dramatically throughout the year between 27.5 °C (dry season) and 35.0 °C (wet season) (White et al. 1989). This constant climate in central Java Island, Indonesia, might induce non-seasonal seaward migration for spawning in a tropical eel A. bicolor bicolor.
The unique year-round spawning ecology and behavior of a tropical eel Anguilla bicolor bicolor may facilitate mating between adult eels from different year classes (cohorts) or from different habitats. Such flexibility in the life history of A. bicolor bicolor might help to maintain the population.
References
Acou A, Laffaille P, Legault A, Feunteun E (2008) Migration pattern of silver eel (Anguilla anguilla, L.) in an obstructed river system. Ecol Freshw Fish 17:432–442
Arai T (2014) Evidence of local short-distance spawning migration of tropical freshwater eels, and implications for the evolution of freshwater eel migration. Ecol Evol 4:3812–3819
Arai T, Aoyama J, Daniel L, Tsukamoto K (1999) Species composition and inshore migration of the tropical eels, Anguilla spp., recruiting to the estuary of the Poigar River, Sulawesi Island. Mar Ecol Prog Ser 188:299–303
Arai T, Limbong D, Otake T, Tsukamoto K (2001) Recruitment mechanisms of tropical eels, Anguilla spp., and implications for the evolution of oceanic migration in the genus Anguilla. Mar Ecol Prog Ser 216:253–264
Arai T, Miller MJ, Tsukamoto K (2003) Larval duration of the tropical eel, Anguilla celebesensis, from the Indonesian and Philippine coasts. Mar Ecol Prog Ser 251:255–261
Boubée JA, Mitchell CP, Chisnall BL, West DW, Bowman E, Haro A (2001) Factors regulating the downstream migration of mature eels (Anguilla spp.) at Aniwhenua Dam, Bay of Plenty, New Zealand. NZ J Mar Freshw Res 35:121–134
Breukelaar AW, Ingendahl D, Vriese FT, De Laak G, Staas S, Klein Breteler JGP (2009) Route choices, migration speeds and daily migration activity of European silver eels Anguilla anguilla in the River Rhine, north-west Europe. J Fish Biol 74:2139–2157
Castle PHJ, Williamson GR (1974) On the validity of the freshwater eel species Anguilla ancestralis Ege from Celebes. Copeia 2:569–570
Deelder CL (1954) Factors affecting the migration of the silver eel in Dutch inland waters. J Cons Perm Int Explor Mer 20:177–185
Deelder CL (1970) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus) 1758. FAO Fisheries Synopsis No. 80 Revision, vol 1, pp 1–73
Durif C, Dufour S, Elie P (2005) The silvering process of Anguilla anguilla: a new classification from the yellow resident to the silver migrating stage. J Fish Biol 66:1025–1043
Durif C, Guibert A, Elie P (2009) Morphological discrimination of the silvering stages of the European eel. Am Fish Soc Symp 58:103–111
Ege V (1939) A revision of the Genus Anguilla Shaw. Dana Rep 16:8–256
Frost WE (1950) The eel fisheries of the River Bann, Northern Ireland, and observations on the age of the silver eels. J Conseil Perm Int Explor Mer 16:358–383
Jellyman DJ (1987) Review of the marine life history of Australasian temperate species of Anguilla. Am Fish Soc Symp 1:276–285
Jonsson N (1991) Influence of water flow, water temperature and light on fish migration in rivers. Nordic J Freshw Res 66:20–35
Kotake A, Arai T, Okamura A, Yamada Y, Utoh T, Oka HP, Miller MJ, Tsukamoto K (2007) Ecological aspects of Japanese eels, Anguilla japonica, collected from coastal areas of Japan. Zool Sci 24:1213–1221
Lokman PM, Vermeulen GJ, Lambert JGD, Young G (1998) Gonad histology and plasma steroid profiles in wild New Zealand freshwater eels (Anguilla dieffenbachii and A. australis) before and at the onset of the natural spawning migration. I. Females. Fish Physiol Biochem 19:325–338
Lowe RH (1952) The influence of light and other factors on the seaward migration of the silver eel (Anguilla anguilla L.). J Anim Ecol 21:275–309
Matsui I (1957) On the record of a leptocephalus and catadromous eels of Anguilla japonica in the waters around Japan with a presumption of their spawning places. J Shimonoseki Coll Fish 7:151–167
Matsui I (1972) Eel biology–biological study. KoseishaKoseikaku, Tokyo
Minegishi Y, Aoyama J, Tsukamoto K (2008) Multiple population structure of the giant mottled eel Anguilla marmorata. Mol Ecol 17:3109–3122
Robinet T, Lecomte-Finiger R, Escoubeyrou K, Feunteun E (2003) Tropical eels Anguilla spp. recruiting to Réunion Island in the Indian Ocean: taxonomy, patterns of recruitment and early life histories. Mar Ecol Prog Ser 259:263–272
Sasai S, Aoyama J, Watanabe S, Kaneko T, Miller MJ, Tsukamoto K (2001) Occurrence of migrating silver eels, Anguilla japonica, in the East China Sea. Mar Ecol Prog Ser 212:305–310
Schmidt J (1922) The breeding places of the eel. Phil Trans R Soc Lond 211:179–208
Sloane RD (1984) Preliminary observations of migrating adult freshwater eels (Anguilla australis australis Richardson) in Tasmania. Aust J Mar Freshw Res 35:471–476
Smith MW, Saunders JW (1955) The American eel in certain fresh waters of the Maritime Provinces of Canada. J Fish Res Board Can 12:238–269
Sugeha HY, Arai T, Miller MJ, Limbong D, Tsukamoto K (2001) Inshore migration of the tropical eels Anguilla spp. recruiting to the Poigar River estuary on north Sulawesi Island. Mar Ecol Prog Ser 221:233–243
Svedäng H, Wickström H (1997) Low fat contents in females silver eels: indications of insufficient energetic stores for migration and gonadal development. J Fish Biol 50:475–486
Tesch FW (2003) The eel. Biology and management of anguillid eels. Chapman and Hall, London
Todd PR (1974) Studies on the reproductive biology of New Zealand freshwater eels. PhD thesis, Victoria University of Wellington, Wellington, New Zealand
Todd PR (1981) Timing and periodicity of migrating New Zealand freshwater eels (Anguilla spp.). NZ J Mar Freshw Res 15:225–235
Tsukamoto K (1992) Discovery of the spawning area for the Japanese eel. Nature 356:789–791
Verreault G, Mingelbier M, Dumont P (2012) Spawning migration of American eel Anguilla rostrata from pristine (1843–1872) to contemporary (1963–1990) periods in the St Lawrence Estuary, Canada. J Fish Biol 81:387–407
Vøllestad LA, Jonsson B (1986) Life-history characteristics of the European eel Anguilla anguilla in the Imsa River, Norway. Trans Am Fish Soc 115:864–871
Vøllestad LA, Jonsson B, Hvidsten NA, Naesje TF, Haraldstad O, Ruud-Hansen J (1986) Environmental factors regulating the seaward migration of European silver eels (Anguilla anguilla). Can J Fish Aquat Sci 43:1909–1916
Vøllestad LA, Jonsson B, Hvidsten NA, Næsje TF (1994) Experimental test of environmental factors influencing the seaward migration of European silver eels. J Fish Biol 45:641–651
Watanabe S, Aoyama J, Hagihara S, Ai B, Azanza RV, Tsukamoto K (2013) Anguilla huangi Teng, Lin, and Tzeng, 2009, is a junior synonym of Anguilla luzonensis Watanabe, Aoyama, and Tsukamoto, 2009. Fish Sci 79:375–383
White AT, Martosubroto P, Sadorra MSM (1989) The coastal environmental profile of Segara Anakan-Cilacap, South Java, Indonesia. ICLARM Technical Reports 25. International Center for Living Aquatic Resources Management, Manila, Philippines
Acknowledgments
We are grateful to Terry Louis Kepel for their kind assistance with the field survey. This work was supported in part by the Ministry of Higher Education Malaysia under the Fundamental Research Grant Scheme (Vot Nos. 59281 and 59406) and the Higher Institution Centre of Excellence (HICoE) Research Grant (Vot No. 66928), under the Institute of Oceanography and Environment (INOS).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: D. Righton.
Reviewed by undisclosed experts.
Rights and permissions
About this article

Cite this article
Arai, T., Abdul Kadir, S.R. & Chino, N. Year-round spawning by a tropical catadromous eel Anguilla bicolor bicolor . Mar Biol 163, 37 (2016). https://doi.org/10.1007/s00227-015-2792-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-015-2792-8