
Abstract
Many wood physical and mechanical traits are important functional attributes for tree species, but variation in these traits among taxonomic categories such as between gymnosperms and angiosperms is still poorly documented. Here, the systematic differences in 12 traits and their allometric relationships between the two tree categories and the potential effects of phylogeny are explored based on a database for major gymnosperm and angiosperm tree species across China. The results are summarized below: (1) means of wood traits were all significantly lower in gymnosperms than in angiosperms. (2) Air-dried density (ADD) and tangential shrinkage coefficient (TSC) are key traits that summarize the correlations among wood traits for gymnosperms, while ADD and radial shrinkage coefficient (RSC) represent those for angiosperms. The allometric slopes of other traits, except for hardness of transverse section (HES), against ADD for gymnosperms were significantly steeper than or similar to the corresponding slopes for angiosperms. On the contrary, the slopes of other traits (except TSC) against RSC for gymnosperms were shallower than or similar to their counterparts for angiosperms. Generally, wood traits were positively related with each other, except that TSC was negatively related to density-related (ADD, BD) and hardness-related traits (HES, HRS and HTS) in gymnosperms. (3) Phylogeny had significant effects on some wood traits of gymnosperms, but had no effects on traits of angiosperms. The present analyses demonstrated a systematic difference in wood traits between two major plant categories, which suggests the evolutionary divergence (TSC, RSC) and convergence (ADD) in key functional traits among woody plants.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Wood physical and mechanical properties are not only related to the mechanical support of stems and limbs (Niklas 1992), and the hydraulic function (low density with high hydraulic conductivity) (Hacke et al. 2001; Preston et al. 2006), but also affect the morphological structure of individual plants (Sterck et al. 2001; Poorter et al. 2006). Some wood traits (e.g., density) are closely linked with the survival and mortality rate, reproductive time (Swenson and Enquist 2007; Wright et al. 2003) and life span (Sterck et al. 2001) of trees, as well as the growth rate (Sotelo et al. 2007b) of stem diameter and canopy (King et al. 2005). Wood traits are also connected with resource competition among species (Baker et al. 2004), community dynamics and ecosystem functions (Chave et al. 2006; Zhang et al. 2011). Therefore, studying of wood physical and mechanical properties can benefit our understanding of ecological processes from individual plants to ecosystem scales.
Current studies mainly focused on wood density (Swenson and Enquist 2007; Chave et al. 2009; Zanne et al. 2010; Zheng and Martínez-Cabrera 2013). A number of studies reported other traits, such as correlations between wood shrinkage and tree growth (Sotelo et al. 2007b); bending strength (MOR) and bending modulus of elasticity (MOE) of wood in a tropical rain forest (Van et al. 2006); crushing strength, stem-growth traits and branch-wood traits of Calycophyllum spruceanum (Sotelo et al. 2007a, 2003), wood color variables among trees and shrub species (Sotelo et al. 2013). Nevertheless, few studies have ever addressed the variations in wood traits and their interrelations between gymnosperms and angiosperms (Zhu et al. 2015). Therefore, a comprehensive comparison of wood traits between gymnosperms and angiosperms is of great importance.
Gymnosperm and angiosperm trees differ markedly not only in evolutionary history and distribution, but also in wood structure. Gymnosperms are the original seed plants, mainly distributed in areas with nutrient-poor soils (Aerts 1995). The structure of gymnosperms wood is relatively simple, consisting of tracheid and rays, with tracheids mainly responsible for the function of transport and mechanical support. On the other hand, angiosperm wood is mostly composed of vessels, rays, fibers and parenchyma cells. These differences are expected to cause quite different wood physical and mechanical attributes between gymnosperms and angiosperms. However, the differences, if any, in wood traits and their interrelations were seldom documented between these two plant categories, and it is also unclear whether these traits are affected by phylogeny.
Based on a database of wood traits across major tree species in China, the difference in 12 wood physical and mechanical properties between gymnosperms and angiosperms, and the phylogenic effects on these traits were examined. In addition, the allometric relationships (Niklas 1994) between wood traits of gymnosperms versus angiosperms were explored. Specifically, the following hypotheses were tested: (1) gymnosperms have lower values of wood physical and mechanical traits than angiosperms. (2) Allometric relationships between wood traits of gymnosperms are different from those of angiosperms. (3) Phylogeny may have effects on wood traits of both gymnosperms and angiosperms. Thus, based on the above hypotheses, the similarity and differences of the 12 wood traits between gymnosperms and angiosperms across China were explored and it was attempted to afford more information about the criteria to determine wood usage.
Materials and methods
Wood physical and mechanical properties
A comprehensive database of wood traits for major gymnosperm and angiosperm tree species across China, collected from the studies, was established (Anatomy and Properties of Chinese Woods, Wood Physical and Mechanical Properties of Main Tree Species in China, Atlas of Gymnosperms Woods of China, Wood Properties of Main Tree Species from Plantation in China; see Reference S1 for more details). Description and definition of the 12 wood traits are as below.
BD, basic density: the oven-dry mass of a wood sample divided by its green volume. ADD, air-dried density: the air-dry mass of a wood sample divided by its air-dried volume. RSC/TSC/VSC, radial/tangential/volumetric shrinkage coefficient: the relative change of dimension with change of wood moisture content across the radial/tangential/volumetric plane. MOR, bending strength: the stress in a material just before it yields in a flexure test. MOE, bending modulus of elasticity: an intensive property that is computed as the ratio of stress to strain in flexural deformation, or the tendency for a material to bend. CSG, compression strength parallel to grain: maximum stress sustained by a compression parallel-to-grain specimen. RES, resilience: the capacity of wood to absorb energy when it is deformed elastically and then upon unloading to have this energy recovered. HES/HRS/HTS, hardness of transverse/radial/tangential section: resistance to indentation using a modified Janka hardness test, measured by the load required to embed an 11.28-mm ball to one-half its diameter in transverse/radial/tangential plane (Green et al. 1999). More detailed description of the mathematical formulas for the 12 wood physical and mechanical traits is shown as supporting information (Appendix S1).
These traits can well characterize the main wood properties in density (ADD, BD), dry shrinkage coefficient (RSC, TSC and VCS), resilience (MOE, RES), strength and hardness (MOR, CSG, HES, HRS and HTS).
The gymnosperms in this study involve 98 species belonging to 27 genera and six families: Cupressaceae, Taxaceae, Podocarpaceae, Cephalotaxaceae, Pinaceae and Taxodiaceae. The woody angiosperms consist of 372 species belonging to 66 genera and 76 families (Fig. 1).

Wood sampling sites for all the gymnosperm and angiosperm tree species in this study
Data analysis
Shapiro–Wilk test was used for data normality (Shapiro and Wilk 1965). For both gymnosperms and angiosperms, the 12 wood physical and mechanical traits did not follow a normal distribution (Fig. 2). Consequently, MOE, RES, HRS and HTS were log-transformed, and other traits were square-root transformed, to improve normality before data analyses. Then, t test was used to compare the mean values of the 12 individual wood traits between gymnosperms and angiosperms. Principal component analysis (PCA) was conducted for the 12 wood traits, for gymnosperms and angiosperms separately. Traits with the greatest contribution to the first two PCA axes were identified as the key traits that summarized the correlations among wood traits in this study. Then, it was examined how the allometric relationships between other traits and these key traits differ among gymnosperms and angiosperms (Warton et al. 2012). Allometric analyses were conducted with type-II regressions (standard major axis regression). The type-II regression slopes were compared with the standard F tests (Warton et al. 2006) between gymnosperms and angiosperms.

Histograms showing the distribution of 12 wood physical and mechanical traits of gymnosperms and angiosperms. BD basic density; ADD air-dried density; RSC radial shrinkage coefficient; TSC tangential shrinkage coefficient; VSC volumetric shrinkage coefficient; MOR bending strength; MOE bending modulus of elasticity; CSG compression strength parallel to grain; RES resilience; HES hardness of transverse section; HRS hardness of radial section; HTS hardness of tangential section
Based on the APG III system (Bremer et al. 2009), a phylogenetic tree for the species was generated in this study. The tree topology was obtained with the online Phylomatic [storedtree = Phylomatic tree R20120829 (plants)] (Webb and Donoghue 2005). The branch lengths were estimated with the bladj function of Phylocom (Wikstrom et al. 2001; Webb et al. 2008), based on the node ages (Wikstrom et al. 2001). Then, the phylogenetic Euclidean distances (Greenacre and Primicerio 2008) among gymnosperms (or angiosperms) species were calculated at the family level. Euclidean distance is the physical distance in two- or three-dimensional space to multidimensional space, often referred to as the “Pythagorean distance” as well (Greenacre and Primicerio 2008). The equation: \(d_{x,y} = \sqrt {\sum\nolimits_{n = 1}^{n} {\left( {x_{n} - y_{n} } \right)^{2} } }\). Euclidean distance among species is based on the distance in phylogeny, which relies on a measure of “genetic distance” between the sequences being classified. Then, a set of coordinates marking the positions of the species will be generated based on node age within the phylogenetic trees. The Euclidean distance, i.e., the evolutionary distances among species, can be obtained with the above equation. For each wood trait, the Euclidean distances among gymnosperms (or angiosperms) were also calculated. Mantel test (Mantel 1967) was used to relate Euclidean distance matrix of species to Euclidean distance matrix of wood traits. This analysis was conducted separately for gymnosperms and angiosperms to examine whether they differed in the phylogenetic control of wood traits.
All statistical analyses were performed using R package 3.0.2 software. PCA was performed with FactoMineR package (Josse et al. 2008), while allometric analysis was conducted with the smart package (Warton et al. 2012). The phylogenetic and trait Euclidean distances between species pairs were calculated with ape package (Paradis et al. 2004) and simba package, respectively.
Results
Statistics of wood traits for gymnosperms and angiosperms
Wood traits varied greatly but with different coefficients of variance (CV): HRS and HTS had larger CV than the other traits (Table 1). Consistent with the current hypothesis, means of all wood traits were lower (P < 0.05) in gymnosperms than angiosperms (e.g., 411 vs. 552 kg m−3 for ADD, 0.15 vs. 0.18% for RSC, 0.29 vs. 0.30% for TSC; all P < 0.05) (Table 1; Fig. 2). Gymnosperms had smaller CV of wood traits than angiosperms, except for RSC.
Key wood traits for gymnosperms and angiosperms
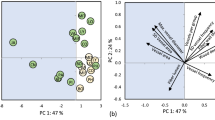
For gymnosperms, the PCA analysis on 12 wood traits showed that the first two axes of PCA explained most (85.9%) of the variation (Table 2, Table S2, S3), which is enough to reflect the majority of information on wood physical and mechanical traits of gymnosperms. For the first axis of PCA, the traits of density (ADD, BD), resilience (MOE, RES), strength and hardness (MOR, CSG, HES, HRS and HTS) had high contributions, with ADD the most important (12%). For the second axis, only RSC, TSC and VSC had high explanatory powers, and TSC was the most important (33.5%). Thus, ADD and TSC, with the highest relative contribution to the first and second axis, respectively, can serve as “key traits” to characterize the two major axes of wood trait variations for gymnosperms.
PCA analysis on wood traits of angiosperms also showed that the first two PCA axes together explained 88.4% of the variation. ADD was the most important contributor (9.7%) to the first PCA axis. However, RSC was the most important (30%) for the second axis, which is different from gymnosperms. Thus, ADD and RSC can be regarded as the key wood traits for angiosperms tree species.
Allometric relationships of wood traits against ADD, TSC and RSC
Most wood physical and mechanical traits had positive effect on the key traits (ADD, RSC and TSC) based on the allometric analysis, except for the traits reflecting density (ADD and BD) and hardness (HES, HRS and HTS) against TSC for gymnosperms (Fig. 3).

Allometric relationships between the three key traits (ADD, TSC and RSC) and the other wood traits. The fitted lines show the standardized major axis regression results (P < 0.05). Equation above is for gymnosperms. Below is for angiosperms. BD basic density; ADD air-dried density; RSC radial shrinkage coefficient; TSC tangential shrinkage coefficient; VSC volumetric shrinkage coefficient; MOR bending strength; MOE bending modulus of elasticity; CSG compression strength parallel to grain; RES resilience; HES hardness of transverse section; HRS hardness of radial section; HTS hardness of tangential section
The allometry slopes of RSC, TSC, MOE against ADD for gymnosperms were 1.2–1.3 times greater than the corresponding slopes for angiosperms (P < 0.05). The slope of HES against ADD for gymnosperms was 1.2 times smaller than that of angiosperms (P < 0.05). There was no significant difference in the allometric slopes of the other six traits against ADD between gymnosperms and angiosperms (P > 0.05; Fig. 3, Table S1).
The allometric analysis on TSC versus other traits showed that the slopes of most traits against TSC for angiosperms were 1.2–1.6 times greater than those for gymnosperms (P < 0.05; Fig. 3), except that the slopes for RSC, VSC and MOE were not significantly different between gymnosperms and angiosperms.
The slopes of most traits (except TSC) against RSC for angiosperms were 1.2–2.0 times greater than the corresponding slopes for gymnosperms (P < 0.05; Fig. 3, Table S1). All slopes were greater than 1.0, except that the slope of TSC against RSC was not significantly different from 1.0 (P > 0.05), for both gymnosperms and angiosperms.
Influence of phylogeny on wood mechanical and mechanical traits
There were significant positive correlations between the Euclidean distance matrix of the traits (BD, TSC, VSC and CSG) and the phylogenetic Euclidean distance matrix for gymnosperms (r > 0.1, P < 0.05; Table 3, Fig S1), suggesting that between-species differences in these traits were significantly associated with phylogenetic distance between species. However, no significant correlation between trait distance and phylogenetic distance was found in angiosperms (Table 3).
Discussion
Difference in wood traits between gymnosperm and angiosperm tree species
Comparison of 12 wood physical and mechanical traits between major gymnosperm and angiosperm tree species showed that the values of all wood traits of angiosperms were higher than those of gymnosperms. The high trait values might suggest a generally better wood functioning or some competitive advantages for angiosperms, which may contribute to the currently wider spread of angiosperm versus gymnosperm trees over the Earth.
Functions (which can be indicated by traits) are consistent with structures. Wood physical and mechanical traits are important functional attributes for tree species. So, the difference in these wood traits may be explained by the different wood anatomical features of gymnosperms versus angiosperms. Gymnosperm wood consists of tracheids, rays and parenchyma. The relative proportions of the three tissues are generally similar among gymnosperms (Cheng 1985). On the other hand, angiosperm wood is structurally more complex, composed of vessels, fibers, rays and parenchyma. The relative proportions of these wood tissues, however, are markedly variable across angiosperm tree species. Consistently, the current results showed that gymnosperm wood traits demonstrated smaller CVs than angiosperms (Table 1). Previous studies also found that wood traits of angiosperms varied more dramatically with climate than those of angiosperms (Cheng 1985; Carlquist 2001). Tracheid and wood fiber cells are mainly responsible for mechanical support in gymnosperms and angiosperms, respectively. However, the tracheid walls are thinner than the fibers walls, which explained why gymnosperms had lower values of strength-related (CSG and MOR) and hardness-related (HES, HRS and HTS) traits than angiosperms (Table 1).
The difference in wood traits between gymnosperms and angiosperms is further reflected in their “key traits.” Previous studies have determined the central role of wood density in woody plant functioning (Swenson and Enquist 2007), as shown by the current PCA analysis that ADD was the common “key traits” of the two taxonomic categories. Furthermore, TSC was another key trait for gymnosperms, while RSC was another major trait for angiosperms. For gymnosperms, cross-field pitting (i.e., the pores between the ray parenchyma cells and the adjacent vertical tracheids) usually occurs on radial walls, leading to much more pits on the tracheid radial walls than on tangential walls (Cheng 1985). Therefore, radial shrinkage is more limited than tangential shrinkage in wood of gymnosperms, which may be the possible reason that TSC was the key trait of gymnosperms. No difference was found in the number and size of pits between the radial and tangential vessel walls of angiosperm woods (Bao et al. 1984). However, the number, size and cell wall thickness of vessels and fibers are much more variable radially than tangentially. This structural feature of wood might partly explain why RSC was a key wood trait besides ADD for angiosperms.
Wood traits are the main criteria to determine wood usage (Bowyer et al. 2007). Density is currently considered as an important criterion in determining wood strength, elasticity and hardness. The current results supplied useful references for wood applications. In addition to density, the key traits (TSC for gymnosperms and RSC for angiosperms) can also be used as the primary indicators to judge the qualities and usages of the two categories of wood, respectively.
Allometries of wood traits against ADD, TSC and RSC
Although the “key traits” of gymnosperms (ADD and TSC) and angiosperms (ADD and RSC) were generally positively correlated with most other wood traits, TCS of gymnosperms was negatively but nonsignificantly correlated with density traits (ADD and BD) and hardness traits (HES, HRS and HTS) based on the allometric analysis, contrary to the corresponding relations in angiosperms (Fig. 3). The different allometries of TSC versus density and hardness traits between gymnosperms and angiosperms may also reflect the difference in wood structures of the two taxonomic groups. Vessels usually have higher shrinkage than tracheids; the shrinkage of angiosperm wood is mainly controlled by its vessels in both radial and tangential directions. Similarly, the radial shrinkage of gymnosperm wood is controlled by its tracheids. However, tangential tracheid walls are thicker in the ray-contact areas, which is a general characteristic of coniferous wood, called “raythickness” (Ladell 1967; Keith 2007). The thicker tangential tracheid walls (raythickness), together with the concomitant smaller lumina, in the tangential versus radial direction may possibly result in relatively low shrinkage of tracheids tangentially versus radially, thus leading to nonsignificant allometric relationships (P > 0.05; Fig. 3) of TSC against density traits (ADD and BD) and hardness traits (HES, HRS and HTS) of gymnosperm wood. However, the underlying mechanisms need to be further explored.
Influence of phylogeny on wood physical and mechanical traits
Correlation analysis between the Euclidean distance matrixes (Table 3) suggested that wood traits of gymnosperms were significantly affected by phylogeny, but traits of angiosperms did not show phylogenic effects. Phylogeny may interact with the environment in shaping the spatial patterns of wood traits (Zhang et al. 2011). Gymnosperms are ancient seed plants with many epibiotic species (Mutke and Barthlott 2005), which suggests that their phylogenetic tree covers long time span (e.g., the longest Euclidean distance between gymnosperm species is 603, much longer than that (390) between angiosperms in this study). These plants mostly grow in relative similar habitats, for example, with poor soils, dry and cool climate (Aerts 1995). Thus, phylogeny might impose relatively stronger influences on the wood traits of gymnosperms than the habitat environments did. In contrast, angiosperms appeared later on the Earth but have successfully spread to diverse habitats. Consequently, phylogeny may play a less important role than the habitats in shaping the wood traits of angiosperms. Unfortunately, there are no habitat data for these plants to test this hypothesis at the present time. The fact that phylogeny affects only wood traits of gymnosperms may suggest the differential evolutionary controls on plant functional traits of the two categories (gymnosperms vs. angiosperms).
Conclusion
Comparison of wood traits between gymnosperms and angiosperms revealed markedly patterns of evolutionary divergence and convergence in key functional traits of woody plants. Means of wood traits were all significantly lower in gymnosperms than in angiosperms. Air-dried density (ADD) and tangential shrinkage coefficient (TSC) are key traits that summarize the correlations among wood traits (or represent the major axis of wood traits syndrome) for gymnosperms, while ADD and radial shrinkage coefficient (RSC) are key traits for angiosperm. Generally, wood traits for both plant categories were positively related to each other, except that TSC was negatively but nonsignificantly related to density- and hardness-related traits in gymnosperms. The allometry slopes of most traits against ADD for gymnosperms were steeper than those for angiosperms, while the slopes of most traits against RSC were shallower for gymnosperms than those for angiosperms. Phylogeny may have significant effects on some wood traits of gymnosperms, but did not show influences on any wood trait of angiosperms.
References
Aerts R (1995) The advantages of being evergreen. Trends Ecol Evol 10:402–407
Baker TR, Phillips OL, Malhi Y, Almeida S, Arroyo L, Di Fiore A, Ewin T, Killeen TJ, Laurance SG, Laurance WF, Lewis SL, Lloyd J, Monteagudo A, Neill DA, Patino S, Patino S, Pitman NCA, Silva JNM, Martinez RV (2004) Variation in wood density determines spatial patterns in Amazonian forest biomass. Glob Chang Biol 10:545–562
Bao FC, Hu R, Tan O, Zhang XP (1984) Fluid permeability in wood and factors affecting on it. Scientia silvae sinicae 20:277–290
Bowyer JL, Shmulsky R, Haygreen JG (2007) Forest products and wood science: an introduction. Blackwell Publishing, Ames
Bremer B, Bremer K, Chase M, Fay MF, Reveal JL, Soltis DE, Soltis PS, Stevens PF, Anderberg AA, Moore MJ, Olmstead RG, Rudall PJ, Sytsma KJ, Tank DC, Wurdack K, Xiang JQY, Zmarzty S (2009) An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG III. Bot J Linn Soc 161:105–121
Carlquist S (2001) Comparative wood anatomy: systematic, ecological, and evolutionary aspects of dicotyledon wood. Springer, Berlin
Chave J, Muller-Landau HC, Baker TR, Easdale TA, Steege HT, Webb CO (2006) Regional and phylogenetic variation of wood density across 2456 neotropical tree species. Ecol Appl 16:2356–2367
Chave J, Coomes D, Jansen S, Lewis SL, Swenson NG, Zanne AE (2009) Towards a worldwide wood economics spectrum. Ecol Lett 12:351–366
Cheng JQ (1985) Wood science. Forestry Publishing House, Beijing
Green DW, Winandy JE, Kretschmann DE (1999) Mechanical properties of wood. Wood handbook: wood as an engineering material. Madison, USDA Forest Service, Forest Products Laboratory. General technical report FPL GTR-113: 4.1–4.45
Greenacre M, Primicerio R (2008) Measures of distance between samples: Euclidean. In: Fundacion BBVA Publication (December 2013), pp 978–84
Hacke UG, Sperry JS, Pockman WT, Davis SD, McCulloh KA (2001) Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 126:457–461
Josse J, Lê S, Husson F (2008) FactoMineR: an R package for multivariate analysis. J Stat Softw 25:1–18
Keith CT (2007) Tangential wall thickenings in conifer tracheids at ray-contact areas. Wood Fiber Sci 7:129–135
King DA, Davies SJ, Supardi MN, Tan S (2005) Tree growth is related to light interception and wood density in two mixed dipterocarp forests of Malaysia. Funct Ecol 19:445–453
Ladell JL (1967) Ray thickenings in the walls of conifer tracheids. Nature 213:470–473
Mantel N (1967) The detection of disease clustering and a generalized regression approach. Can Res 27:209–220
Mutke J, Barthlott W (2005) Patterns of vascular plant diversity at continental to global scales. Biologiske Skrifter 55:521–531
Niklas KJ (1992) Plant biomechanics: an engineering approach to plant form and function. The University of Chicago Press, Chicago
Niklas KJ (1994) Plant allometry: the scaling of form and process. The University of Chicago Press, Chicago
Paradis E, Claude J, Strimmer K (2004) APE: analyses of phylogenetics and evolution in R language. Bioinformatics 20:289–290
Poorter L, Bongers L, Bongers F (2006) Architecture of 54 moist-forest tree species: traits, trade-offs, and functional groups. Ecology 87:1289–1301
Preston KA, Cornwell WK, DeNoyer JL (2006) Wood density and vessel traits as distinct correlates of ecological strategy in 51 California coast range angiosperms. New Phytol 170:807–818
Shapiro SS, Wilk MB (1965) An analysis of variance test for normality (complete samples). Biometrika 52:591–611
Sotelo MC, Vidaurre H, Weber J (2003) Variation in stem-growth and branch-wood traits among provenances of Calycophyllum spruceanum Benth. from the Peruvian Amazon. New For 26:1–16
Sotelo MC, Beaulieu J, Hernández RE (2007a) Genetic variation in wood mechanical properties of Calycophyllum spruceanum at an early age in the Peruvian Amazon. Wood Fiber Sci 39:578–590
Sotelo MC, Hernandez J, Beaulieu RE (2007b) Genetic variation in wood shrinkage and its correlations with tree growth and wood density of Calycophyllum spruceanum at an early age in the Peruvian Amazon. Can J For Res 37:966–976
Sotelo MC, Weber JC, Garcia RA, Silva DA, Muñiz GI (2013) Variation in wood color among natural populations of five tree and shrub species in the Sahelian and Sudanian ecozones of Mali. Can J For Res 43:552–562
Sterck FJ, Bongers F, Newbery DM (2001) Tree architecture in a Bornean lowland rain forest: intraspecific and interspecific patterns. Plant Ecol 153:279–292
Swenson NG, Enquist BJ (2007) Ecological and evolutionary determinants of a key plant functional trait: wood density and its community-wide variation across latitude and elevation. Am J Bot 94:451–459
Van GH, Poorter L, Sterck F (2006) Wood mechanics, allometry, and life-history variation in a tropical rain forest tree community. New Phytol 171:367–378
Warton DI, Wright IJ, Falster DS, Westoby M (2006) Bivariate line-fitting methods for allometry. Biol Rev 81:259–291
Warton DI, Duursma RA, Falster DS, Taskinen S (2012) Smatr 3–an R package for estimation and inference about allometric lines. Method Ecol Evol 3:257–259
Webb CO, Donoghue MJ (2005) Phylomatic: tree assembly for applied phylogenetics. Mol Ecol Notes 5:181–183
Webb CO, Ackerly DD, Kembel SW (2008) Phylocom: software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics 24:2098–2100
Wikstrom N, Savolainen V, Chase MW (2001) Evolution of the angiosperms: calibrating the family tree. Proc R Soc B Biol Sci 268:2211–2220
Wright SJ, Muller-Landau HC, Condit R, Hubbell SP (2003) Gap-dependent recruitment, realized vital rates, and size distributions of tropical trees. Ecology 84:3174–3185
Zanne AE, Westoby M, Falster DS, Ackerly DD, Loarie SR, Arnold SE, Coomes DA (2010) Angiosperm wood structure: global patterns in vessel anatomy and their relation to wood density and potential conductivity. Am J Bot 97:207–215
Zhang SB, Slik JW, Zhang JL, Cao KF (2011) Spatial patterns of wood traits in China are controlled by phylogeny and the environment. Glob Ecol Biogeogr 20:241–250
Zheng JM, Martínez-Cabrera HI (2013) Wood anatomical correlates with theoretical conductivity and wood density across China: evolutionary evidence of the functional differentiation of axial and radial parenchyma. Ann Bot 112:927–935
Zhu JL, Shi Y, Fang LQ, Liu XE, Ji CJ (2015) Patterns and determinants of wood physical properties across major tree species in China. Sci China Life Sci 58:602–612
Acknowledgements
The authors thank the anonymous referees and the editor for their valuable suggestions, which have improved the manuscript. They also thank Dr. Jinlong Zhang from Kadoorie Farm and Botanic Garden, Hong Kong for his great help with phylogenetic analysis. The project was funded by the National Natural Science Foundation of China (#41473068, 31370419, 31370620) and Ministry of Science and Technology of China Special Funding for Basic Works project (#2011FY110300).
Author information
Authors and Affiliations
Corresponding authors
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Zhang, M., Ji, C., Zhu, J. et al. Comparison of wood physical and mechanical traits between major gymnosperm and angiosperm tree species in China. Wood Sci Technol 51, 1405–1419 (2017). https://doi.org/10.1007/s00226-017-0954-1
Received:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00226-017-0954-1