
Abstract
Vascular calcification is an important pathological condition associated with increased risk of cardiovascular mortality. Hydroxyapatite (HA) found in such deposits is the same polymorph of calcium (Ca) found in bone, indicating calcification may involve mechanisms akin to bone formation. Vascular smooth muscle cells (Vsmcs) have been shown to undergo phenotypic change to osteoblast-like cells. However, the mechanisms underlying this phenotypic change are unclear, and whether the stimulus to become osteogenic is a result of loss of mineralization inhibitors or early mineral deposits is not known. Our aim in this study is to identify mechanisms and signal transduction pathways that cause differentiation of Vsmcs into osteoblast-like cells in the presence of HA. We first characterized vascular origin of Vsmcs by studying the expression of smooth muscle cell markers: myosin heavy chain and smooth muscle actin along with SM22α at both mRNA and protein levels. Vsmcs grown on HA exhibited progressive change in cellular morphology at 3-, 7-, and 14-day time points. Culturing of Vsmcs on HA disc resulted in decrease in media Ca levels and increased expression of Ca-sensing receptor (CaSR) on Vsmcs resulting in upregulation of intracellular CaSR signaling leading to increased BMP-2 secretion. BMP-2 pathway mediated differentiation of Vsmcs to osteoblast-like cells shown by expression of osteogenic markers like runt-related transcription factor 2, osteocalcin, and alkaline phosphatase at mRNA and protein levels. Blocking CaSR by NPS-2143 reduced BMP-2 secretion and blocking the BMP-2 pathway by LDN-193189, a BMP inhibitor, modulated expression of osteogenic markers confirming their role in osteogenesis of Vsmcs.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Vascular calcification is a leading cause of increased morbidity and mortality in patients [1–3]. Vascular calcification can be divided into two distinct categories based on location. Atherosclerotic intimal layer calcification is located in the intimal layer and is associated with atherosclerosis. Monckeberg’s arteriosclerosis (MAC) occurs in the arterial medial layer and is associated with elastic lamellae [3, 4] in patients with chronic kidney disease and diabetes.
MAC is thought to occur because of an increase in serum Calcium (Ca) and Phosphate (P) levels [5] or loss of calcification inhibitors in the vasculature, such as matrix-gla protein (MGP), fetuin, osteoprotegerin, and osteopontin (OPN) [6–8]. Studies have also indicated that the transformation of Vsmcs into osteoblast-like cells is a key mechanism in progression of vascular calcification [9]. There is evidence of the existence of a subpopulation of smooth muscle cells in the vascular wall, calcifying vascular cells (CVCs), which form nodules and calcify under long-term culture. These structures have been shown to share properties with bone, including increased ALP activity, and OCN and OPN expressions [10–12]. Bone morphogenetic proteins (BMPs) belong to the transforming growth factor beta (TGF-β) superfamily, form ectopic bone, and play a role in vascular calcification [13]. Several studies have demonstrated that BMP-2, 3, 4, and 7 can upregulate differentiation markers of mature osteoblasts like ALP and OCN expression [14]. In addition, experiments have demonstrated that BMP-2 acts in differentiation of mesenchymal stem cells into osteoblast-like cells, which suggests that it may play a role in regulating specific differentiation pathways [15]. However, it is unclear if passive spontaneous early mineralization occurs prior to osteogenesis of cells, especially for elastin-specific calcification in Monckeberg’s arteriosclerosis. Using an in vitro model, several studies have focused on understanding the process of vascular calcification. The in vitro calcification of primary mouse [16], human [17], and bovine [18] Vsmcs are commonly utilized models of vascular calcification.
We have recently shown that osteogenic trans-differentiation of Vsmcs occurs during culture on HA surface and calcified vascular elastin [19]. The aim of the current study is to understand cellular signaling pathways involved in the differentiation of Vsmcs into osteoblast-like cells upon culture on HA surface. We report that culturing of Vsmcs on HA surface results in upregulation of calcium-sensing receptor (CaSR), which stimulates expression of BMP-2. The BMP-2 pathway regulates the differentiation of Vsmcs into osteoblast-like cells through the SMAD-5 signaling pathway.
Materials and Methods (Detailed Materials and methods available in online Supplementary Resource 1)
Vascular Smooth Muscle Cell Culture
Rat aortic smooth muscle cells were isolated according to Orlandi et al. [20]. Cells were grown and maintained in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10 % fetal bovine serum incubated at 37 °C with 5 % CO2. Passage numbers 2–6 were used and cells were cultured on one single HA disc in a 12-well plate at 1 × 106 cells/well. For each experiment, three HA discs were used and the experiments were repeated three times. The Ca level in the medium was not altered except for experiments studying the effect of Ca in the medium on Vsmcs.
HA Disc preparation
The HA powder (Sigma-Aldrich, MO) used to prepare discs was uniaxially pressed at 40 MPa into pellets without a binder using a 1/2-inch cylindrical die lubricated with a thin layer of stearic acid. The HA pellets were sintered in a 1,100 °C with ramp rate set at 2 °C/min and soaking time of 2 h.
Protein Isolation
Cells were briefly rinsed with 10 mL ice-cold phosphate buffer saline (PBS) and then trypsinized with 0.25 % trypsin for 5 min. Cell layers were harvested and centrifuged at 25 °C for 5 min at 1,000 rpm and lysed in 200 µl of RIPA buffer (50 mM Tris–HCl; pH 7.5; 135 mM NaCl; 1 % Triton X-100; 1 % sodium deoxycholate; 2 mM EDTA; 50 mM NaF; 2 mM sodium orthovanadate; 10 mg/ml aprotinin; 10 mg/ml leupeptin; and 1 mM PMSF). Samples were centrifuged at 10,000 rpm for 5 min at 4 °C, and total protein concentration was measured using bicinchoninic acid (BCA) protein assay according to the manufacturer’s protocol (Pierce, Rockford, IL).
Statistical Analysis
Quantitative analysis of message levels are presented as mean ± Standard Error (SE) for all experiments (n = 3). Significance of differences between control and treatment experiments was evaluated by Student's unpaired t test; ‘‘*” indicates p < 0.05 were considered to be significant.
Results
Characterization of Smooth Muscle Cells and an HA Surface
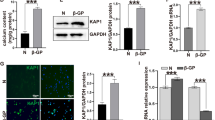
We analyzed gene expression of two typical smooth muscle cell (SMC) lineage markers like SMA and MHC by PCR and immunofluorescence. Rat aortic Vsmcs express SMA and MHC at mRNA (Fig. 1a) and protein level (Fig. 1b). Vsmcs also expressed SMC lineage markers, which were examined using Western blot analysis. A 150 kDa band for MHC, 42 kDa for SMA, and 23 kDa for SM22α were observed (Fig. 1c). GAPDH (36 kDa) was used as a loading control. Based on these results, Vsmcs were confirmed to be of smooth muscle lineage.

Characterization of Vsmc markers and HA surface. a Gene expression of Vsmc markers MHC and SMA by RT-PCR, where GAPDH was used as an internal control. b Immunofluorescence confocal laser scanning microscopy for expression of Vsmc markers, MHC (green fluorescence) and SMA (red fluorescence). Nuclei were stained blue by DAPI. Scale bar 100 μm. HA hydroxyapatite. c Protein expression of MHC, SMA, and SM212α measured by Western blot. d Structure of HA with a typical tiled appearance and Vsmcs exhibiting a typical smooth muscle cell morphology when cultured under noncalcifying conditions. e–g Vsmcs cultured on HA surface for 3–14 days shows morphological change, loss of typical smooth muscle structure, and progressive rounding (Color figure online)
Vsmcs were grown on the HA surface and were characterized using field emission-scanning electron microscopy. Vsmcs grown on coverslips were used as control; they showed a typical vascular phenotype and morphology (Fig. 1d). The HA surface had a characteristic tiled appearance (Fig. 1d). At 3 days, Vsmcs cultured on HA showed morphological changes and loss of typical vascular phenotype evidenced by the rounding of cells as compared to the control Vsmcs (Fig. 1e). At 7 and 14 days Vsmcs exhibited progressive changes in morphology and a further departure from the characteristic Vsmc phenotype (Fig. 1f–g).
Expression of Osteogenic/Chondrogenic Markers Using qPCR
Expression of three typical osteogenic markers, Runt-related transcription factor (RUNX-2), alkaline phosphatase (ALP), and OCN (Osteocalcin), and two chondrogenic markers, aggregan and Col 10a1, in Vsmcs grown on an HA surface for 3, 7, and 14 days was examined by RT-PCR. At Day 3, a progressive increase in expression of early osteogenic markers was observed. RUNX-2 expression increased almost six-fold and continued to increase with time (Fig. 2a). At 3 days, no significant increase in ALP levels was observed. However, at 7- and 14-day time points, a three-to-eight-fold increase in levels of ALP was observed (Fig. 2b). At Day 3, a two-fold increase in expression of OCN was observed; at Day 7 and Day 14, expression increased five-to-nine-fold (Fig. 2c). No significant change in expression levels as compared to control was observed for chondrogenic markers like aggregan and Col 10a1 (Fig. 2d, e), suggesting lack of chondrogenesis.

Expression of osteogenic/chondrogenic markers in Vsmcs cultured on Hydroxyapatite at Day 3, Day 7, and Day 14. D1 Day 1, D7 Day 7, D14 Day 14. a–c Gene expression of RUNX2, ALP, and OCN, respectively, as measured by RT-PCR. A progressive increase in mRNA levels of RUN-X2, ALP, and OCN was observed. GAPDH was used as an internal control, and data were normalized to GAPDH levels. Quantitative analysis of message levels by RT-PCR are presented as mean ± SE ‘‘*” (n = 3), p < 0.05 (Student’s unpaired t test). d, e Gene expression of chondrogenic markers Col 10a1 and Aggregan. At Day 3, Day 7, and Day 14, Col 10a1 and Aggregan expressions were reduced as compared to control. GAPDH was used as an internal control and data were normalized to GAPDH levels. f Immunofluorescence staining for osteogenic marker Collagen type I (green fluorescence). Nuclei were stained with DAPI. g Immunofluorescence staining for smooth muscle cell marker MHC (red fluorescence); nuclei were stained with DAPI (Color figure online)
Expression of Collagen I and MHC was examined by immunofluorescence staining (Fig. 2f). As compared to control, Collagen I expression increased as a result of culture on HA, and expression of the SMC marker MHC decreased (Fig. 2g), suggesting that Vsmcs were becoming more synthetic type.
Effect of Ca Levels in Medium on Trans-Differentiation of Vsmcs
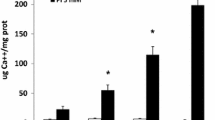
To elucidate the mechanism of this phenotypic change in Vsmcs, Ca levels in culture media were monitored over 14 days. Vsmcs grown without HA showed no significant change in media Ca levels (Fig. 3a). Vsmc cultured on HA surface (Vsmcs + HA + Media) showed a significant decrease in the Ca levels with time. HA discs alone when incubated in the medium (HA + Media) showed similar trend (Fig. 3a). We also tested the Phosphate (P) levels in media. Vsmcs grown without HA showed no significant change in media P levels (Fig. 3a). When HA discs alone were incubated in cell culture medium, the P levels showed a similar decreasing trend with time but the reduction in P levels was much smaller.

Effect of HA on trans-differentiation of Vsmcs. a DMEM media Ca and P levels of Vsmcs grown on HA surface at different time points were examined. A significant decrease in Ca levels in medium for HA alone or Vsmcs cultured on HA was observed at all three time points. b Protein expression level of CaSR and BMP-2 measured by Western blot shows progressive upregulation of both CaSR and BMP-2 for 3, 7, and 14 days. c Gene expression of BMP-2 measured by RT-PCR shows progressive increase. Quantitative analysis of message levels is presented as mean ± SE ‘‘*” (n = 3), p < 0.05 (Student’s unpaired t test). d Immunofluorescence staining for BMP-2 protein (red fluorescence) shows increase in expression of BMP-2 upon culturing on HA surface as compared to control. Nuclei were stained by DAPI. e Gene expression of MGP measured by RT-PCR shows decrease in expression of MGP in HA cultured Vsmcs. Quantitative analysis of message levels is presented as mean ± SE ‘‘*” (n = 3), p < 0.05 (Student’s unpaired t test). f Protein expression levels of CaSR and BMP-2 in Vsmcs treated with CaSR antagonist NPS 2143 for 72 h. Both CaSR and BMP-2 expressions were reduced in antagonist-treated cells. g Schematic representation of CaSR mediated stimulation of BMP-2 expression
The decrease in Ca levels in media when Vsmcs were cultured on HA surface resulted in upregulation of CaSR in cells at 3, 7, and 14 days. As CaSR affects BMP-2 expression, we tested BMP-2 levels. A similar pattern of increase in BMP-2 expression at both mRNA and protein levels was observed for Vsmcs cultured on HA surface (Fig. 3b). A two-fold increase in BMP-2 mRNA levels at Day 3, a four-fold increase at Day 7, and a 12-fold increase at Day 14 were observed (Fig. 3c). Immunofluorescence-labeling studies for BMP-2 further corroborated mRNA and protein data, showing increased BMP-2 expression at Days 3, 7, and 14 when compared to control (Fig. 3d). Thus, these data suggest that CaSR stimulated expression of BMP-2 by mechanism shown in schematic representation (Fig. 3g). To further confirm role of CaSR in stimulation and secretion of BMP-2, we blocked CaSR by NPS 2143, a known CaSR antagonist. We show that CaSR levels were significantly reduced as compared to untreated control (Fig. 3f). BMP-2 levels were also reduced in Vsmcs treated with NPS 2143, suggesting that CaSR activation is involved in secretion of BMP-2. Thus, these results indicate that trans-differentiation of Vsmcs into osteoblast-like cells under calcifying conditions is mediated via the BMP-2 signaling pathway stimulated by CaSR. MGP is known BMP-2 inhibitor. Thus, MGP expression levels were also examined by RT-PCR. At Day 3, MGP expression was slightly lower as compared to control. At Day 7 and Day 14, MGP expression was significantly lower as compared to control (Fig. 3e).
BMP-2–SMAD-5-Mediated Trans-Differentiation of Osteoblast-Like Cells
Increase in BMP-2 expression levels regulates osteogenesis through the SMAD signaling pathway. BMP-2 increases SMAD-5 expression, which translocates to the nucleus and is phosphorylated (Fig. 4a). We therefore evaluated SMAD-5 expression levels in Vsmcs grown on HA by Western blot. SMAD-5 (60 kDa) expression increased progressively at 3, 7, and 14 days as compared to control (Fig. 4b). Expression of phosphorylated SMAD (pSMAD, 61 kDa) increased in a manner similar to SMAD (60 kDa) (Fig. 4b). GAPDH was used as a loading control. The effect of media Ca levels on cellular BMP-2 expression in cells cultured on HA surface was examined. Vsmcs not grown on an HA surface either in the presence or absence of Ca did not express BMP-2 (Fig. 4c lane 1 and 2), and thus, media Ca did not play a significant role. The cells grown on HA in Ca-deficient media expressed lower levels of BMP-2 (Fig. 4b lane 4) than the cells cultured on HA in media containing Ca (Fig. 4c lane 3). A similar expression pattern for SMAD-5 levels was observed. Densitometry analysis of BMP-2 protein expression levels show reduced expression of BMP-2–SMAD-5 (Fig. 4d).

BMP-2–SMAD-5-mediated trans-differentiation of osteoblast-like cells. a Schematic representation of the signaling pathway activated by BMP-2 receptor, which ultimately causes Vsmc differentiation into osteoblast-like cells. b BMP-2, SMAD-5, and pSMAD-5 protein levels measured by Western blot show upregulation in the protein levels for 3, 7, and 14 days. GAPDH was used as loading control. c Protein expression levels of BMP-2 and SMAD-5 in Vsmc in the absence of Ca (lane 3) and the presence of Ca (lane 4) in culture media. Results show a significantly lower expression (43 kDa BMP-2) and (60 kDa SMAD-5) in Vsmcs on HA surface upon culturing in Ca-deficient medium. Vsmcs not grown on HA but in the presence of Ca (lane 1) and the absence of Ca (lane 2) did not express BMP-2. d Densitometry analysis of BMP-2 protein expression levels shows reduced expression of BMP-2 after culture for 3 days
Downregulation of BMP-2–SMAD-5 Pathway by Small Molecule Inhibitor
To further understand the underlying mechanism of BMP signaling in Vsmcs, cells were treated with a small molecule inhibitor LDN-193189, a selective inhibitor for BMP type I receptor kinases that is known to inhibit BMP signaling through pSMAD-5 expression [21]. At both 3 and 7 days, cells grown on HA surface and treated with LDN 193189 showed reduced BMP-2 and pSMAD-5 levels (Fig. 5a, b). To further understand downstream effects of BMP inhibition, activity of key osteogenic marker ALP was studied. Using an ALP colorimetric assay, a dose-dependent decrease in specific activity of ALP was examined. ALP activity in control (cells grown on HA in the absence of an inhibitor) was defined as 100 %. After 3 days of treatment with an LDN inhibitor, the specific activity reduced to 67 and 41 % for samples treated with 50 and 100 nM, respectively (Fig. 5c). At 7 days, a similar dose-dependent decrease in ALP was observed (66 and 56 % for samples treated with 50 and 100 nM, respectively) (Fig. 5d). These findings suggest that blocking the BMP-2 pathway reduces expression of key osteogenic markers like ALP.

Effect of LDN (BMP-2 inhibitor) on Vsmcs. a, b Protein expression levels of BMP-2 and SMAD-5 in Vsmcs cultured on HA disc after LDN treatment for 3 days and 7 days, respectively, shows a dose-dependent decrease in protein levels quantified by densitometry analysis. BMP-2 and SMAD-5 expression levels in cells cultured on HA but not treated with LDN were defined as 100 %. c, d Relative expression of ALP activity in Vsmcs after LDN treatment for 3 days and 7 days, respectively, showed a dose-dependent decrease in ALP activity in LDN-treated samples as compared to control (no inhibitor). No inhibitor control was defined as 100 %. Quantitative analysis of message levels is presented as mean ± SE ‘‘*” (n = 3), p < 0.05 (Student’s unpaired t test)
Discussion
Our data shows that Vsmcs undergo morphological changes and lose their vascular phenotypic markers, start expressing BMP-2, SMAD-5, and other osteogenic markers in response to culturing on HA surface.
Vascular calcification in chronic kidney disease patients is mediated by an increase in circulating Ca and P levels that causes oversaturated plasma [21]. In fact, an increase in Ca and P levels in cell culture medium has been shown to transform Vsmcs to osteoblast-like cells [12]. Studies have revealed a link between vascular calcification and osteogenesis [22–26]. There is evidence showing expression of several key bone formation proteins as well as bone structural proteins in atherosclerotic plaques [10, 24]. Bostrom et al. demonstrated that an increase in MGP expression in the artery wall inhibits calcification by binding and blocking BMP-2 interaction with its receptors [27]. MGP-deficient mice have been shown to die within 4–6 weeks due to extensive medial arterial calcification as a result of uninterrupted BMP-2 activity [27, 28]. Chen et al. found higher BMP-2 levels associated with uremic serum as compared to normal serum, which influenced BMP-2-mediated calcification in bovine SMCs treated with phosphate [29].
It is unclear at this time if mineralization of elastin in the arteries occurs first in the absence of Vsmc phenotypic change to osteoblast-like cells or such transformation is a pre-requisite. To test if mineral deposits can change Vsmc phenotype, we cultured Vsmcs on HA discs. Culturing Vsmcs on HA led to an increase in BMP-2 activity. It also led to the decrease in MGP expression, a key gene known to inhibit BMP-2 interaction with BMP receptor [27]. In a BMP-2-mediated signaling pathway, SMAD gets phosphorylated, translocates to the nucleus, and is known to regulate expression of transcription factors like RUNX-2 and CBFA, which are responsible for osteoblast differentiation [30]. We show that Vsmcs undergo morphological changes as observed by SEM, immunofluorescence, qPCR, and Western blot. A decrease in the expression of MHC, a smooth muscle cell marker and an increase in osteogenic markers like ALP, RUNX-2, and OCN were observed. Furthermore, inhibitor blocking of the BMP-2 pathway caused a decrease in BMP-2 expression and pSMAD-5 levels, which caused a reduction in expression of osteogenic markers like ALP, suggesting that BMP-2 regulates SMAD-5 signaling and differentiation of Vsmcs into osteoblast-like cells. These data strongly suggest that mineral deposits in the arteries can cause Vsmc phenotypic change.
To further understand how culturing of Vsmcs on HA leads to increased BMP-2 expression, the Ca and P levels in the media used for culturing cells were tested. Studies have shown that an increase in Ca/P in the media can cause Vsmcs to exhibit osteoblast-like behavior [31]. Surprisingly, when HA disc was present either alone or with Vsmcs, the media Ca levels dropped significantly. We speculate that Ca from the media could have been adsorbed on the HA surface. This adsorption of Ca may provide higher Ca at the interface of the cell and the HA surface; this would be similar to activity in bone remodeling, in which bone formation by osteoblasts occurs after the initiation of bone resorption by osteoclasts. Bone resorption produces a local increase in the extracellular Ca concentration beneath a resorbing osteoclast and reaches levels as high as 40 mM [32]. This increase in local ionized Ca during remodeling is critical in regulating osteoblast proliferation and/or differentiation via either activation of the CaSR and/or increasing the Ca2+ influx into osteoblasts. The increase in local Ca levels activates the CaSR, which mediates the stimulation and expression of BMP-2. Chattopadhyay et al. have shown that CaSR stimulates proliferation in rat calvarial osteoblasts and can accelerate Vsmc transformation into osteoblast-like cells, which is followed by their proliferation and progressive calcification [33]. Another study has shown that colonic myofibroblasts express the CaSR, and its activation causes synthesis and secretion of BMP-2 [34]. Furthermore, it has been shown that a Ca-induced signaling cascade leads to osteogenic differentiation of human bone marrow-derived mesenchymal stromal cells which is mediated by activation of CaSR triggered by increased Ca levels resulting in increased BMP-2 expression [35, 36]. Our studies have shown that increase in CaSR expression causes increase expression of BMP-2 (Fig. 3) and further activation of BMP–SMAD pathway, leading to osteogenesis in Vsmcs as observed by several-fold over-expression of osteogenic proteins such as RUNX-2, osteocalcin, and ALP. Blocking either CaSR activation by a CaSR antagonist or BMP signaling led to inhibition of osteogenic transformation of Vsmcs. When cells were cultured in Ca- and P-deficient media, we did not see this increase in BMP-2, clearly suggesting that drop in Ca or P levels in the media is not pre-requisite for Vsmc phenotypic change to osteoblast-like cells. Thus, we speculate that ionized media Ca is getting adsorbed on HA surface in our cell culture studies and that might be exerting effect on Vsmcs, leading to trans-differentiation into osteoblast-like cells. However, adsorption of ionized Ca on HA surface needs further testing.
We have previously shown that removal of mineral allows Vsmcs to revert back to their original phenotype [19]. Thus, it is possible that demineralization strategies could reverse vascular pathology in MAC. Furthermore, our findings are relevant to the stents used in vascular surgery which are coated with HA to improve biocompatibility of stents [37]. Our results suggest that deployment of HA-coated stents could alter Vsmc behavior in arteries and lead to vascular calcification. Thus, a closer look at vascular pathology associated with HA-coated stents is necessary.
Conclusion
The present study demonstrates that existing mineral deposits can cause osteogenic transformation of Vsmcs through CaSR- and BMP-2-mediated pathways. Understanding signal transduction pathways that underlie differentiation of Vsmcs into osteoblast-like cells will help us develop novel therapeutic strategies for remediating vascular calcification and associated vascular diseases.
Limitation of the Study
These are strictly in vitro cell culture experiments and direct application of these results for in vivo calcification is cautioned.
Abbreviations
- VSMCs:
-
Vascular smooth muscle cells
- GAPDH:
-
Glyceraldehyde 3-phosphate dehydrogenase
- SMA:
-
Alpha smooth muscle actin
- MHC:
-
Myosin heavy chain
- BMP-2:
-
Bone morphogenetic protein 2
- SMAD-5:
-
Sma- and Mad-related protein 5
- Col2a1:
-
Collagen type II alpha 1
- RUNX2:
-
Runt-related transcription factor 2
- ALP:
-
Alkaline phosphatase
- DAPI:
-
4′,6-Diamidino-2-phenylindole, dihydrochloride
- HA:
-
Hydroxyapatite
- MGP:
-
Matrix-gla protein
References
Roberto Wayhs, Allan Zelinger, Paolo Raggi (2002) High coronary artery calcium scores pose an extremely elevated risk for hard events. J Am Coll Cardiol 39(2):225–230
Yadon Arad, Louise Spadaro, Ken Goodman, David Newstein, Guerci AD (2000) Prediction of coronary events with electron beam computed tomography. J Am Coll Cardiol 36(4):1253–1260
London GM, Guerin AP, Marchais SJ, Métivier F, Pannier B, Adda H (2003) Arterial media calcification in end-stage renal disease: impact on all-cause and cardiovascular mortality. Nephrol Dial Transplant 18(9):1731–1740
Goodman WG, London G, Amann K, Block GA, Giachelli C, Hruska KA, Ketteler M, Levin A, Massy Z, McCarron DA, Raggi P, Shanahan CM, Yorioka N (2004) Vascular calcification in chronic kidney disease. Am J Kidney Dis 43(3):572–579
Johnson RC, Leopold JA, Loscalzo J (2006) Vascular calcification: pathobiological mechanisms and clinical implications. Circ Res 99(10):1044–1059
Price PA, Faus SA, Williamson MK (1998) Warfarin causes rapid calcification of the elastic lamellae in rat arteries and heart valves. Arterioscler Thromb Vasc Biol 18(9):1400–1407
Price P, William MK, Nguyen TMT, Than TN (2004) Serum levels of the fetuin-mineral complex correlate with artery calcification in the rat. J Biol Chem 279(3):1594–1600
Speer MY, McKee MD, Guldberg RE, Liaw L, Yang H-Y, Tung E, Karsenty G, Giachelli CM (2002) Inactivation of the Osteopontin gene enhances vascular calcification of matrix Gla protein–deficient mice: evidence for osteopontin as an inducible inhibitor of vascular calcification in vivo. J Exp Med 196(8):1047–1055
Watson KE, Bostrom K, Ravindranath R, Lam T, Norton B, Demer LL (1994) TGF-beta 1 and 25-hydroxycholesterol stimulate osteoblast-like vascular cells to calcify. J Clin Invest. 93(5):2106–2113
Bostrom MK, Watson KE, Horn S, Wortham C, Herman IM, Demer LL (1993) Bone morphogenetic protein expression in human atherosclerotic lesions. J Clin Investig 91(4):1800–1809
Reynolds JL, Joannides AJ, Skepper JN, McNair R, Schurgers LJ, Proudfoot D, Jahnen-Dechent W, Weissberg PL, Shanahan CM (2004) Human vascular smooth muscle cells undergo vesicle-mediated calcification in response to changes in extracellular calcium and phosphate concentrations: a potential mechanism for accelerated vascular calcification in ESRD. J Am Soc Nephrol 15(11):2857–2867
Hsueh Yang, Gabrielle Curinga, Giachelli CM (2004) Elevated extracellular calcium levels induce smooth muscle cell matrix mineralization in vitro. Int Soc Nephrol 66:6
Hruska KA, Mathew S, Saab G (2005) Bone morphogenetic proteins in vascular calcification. Circ Res 97(2):105–114
Zhou H, Hammonds RG Jr, Findlay DM, Martin TJ, Ng KW (1993) Differential effects of transforming growth factor-beta 1 and bone morphogenetic protein 4 on gene expression and differentiated function of preosteoblasts. J Cell Physiol 155(1):112–119
Wang EA, Israel ID, Kelly S, Luxenberg DP (1993) Bone morphogenetic protein-2 causes commitment and differentiation in C3H10T1/2 and 3T3 cells. Growth Factors 9(1):57–71
Byon CH, Javed A, Dai Q, Kappes JC, Clemens TL, Darley-Usmar VM, McDonald JM, Chen Y (2008) Oxidative stress induces vascular calcification through modulation of the osteogenic transcription factor Runx2 by AKT signaling. J Biol Chem 22:15319–15327
Ewence AE, Bootman M, Roderick HL, Skepper JN, McCarthy G, Epple M, Neumann M, Shanahan CM, Proudfoot D (2008) Calcium phosphate crystals induce cell death in human vascular smooth muscle cells: a potential mechanism in atherosclerotic plaque destabilization. Circ Res 5:28–34
Collett GDM, Sage AP, Kirton JP, Alexander MY, Gilmore AP, Canfield AE (2007) Axl/phosphatidylinositol 3-kinase signaling inhibits mineral deposition by vascular smooth muscle cells. Circ Res 100(4):502–509
Yang Lei, Aditi Sinha, Nasim Nosoudi, Ankit Grover, Naren Vyavahare (2014) Hydroxyapatite and calcified elastin induce osteoblast-like differentiation in rat aortic smooth muscle cells. Exp Cell Res 323(1):198–208
Orlandi A, Ehrlich HP, Ropraz P, Spagnoli LG, Gabbiani G (1994) Rat aortic smooth muscle cells isolated from different layers and at different times after endothelial denudation show distinct biological features in vitro. Arterioscler Thromb Vasc Biol 14(6):982–989
Shroff Rukshana, Long David A, Shanahan Catherine (2013) Mechanistic insights into vascular calcification in CKD. J Am Soc Nephrol 24:179–189
Cuny GD, Yu PB, Laha JK, Xing X, Liu J-F, Lai CS, Deng DY, Sachidanandan C, Bloch KD, Peterson RT (2008) Structure–activity relationship study of bone morphogenetic protein (BMP) signaling inhibitors. Bioorg Med Chem Lett 18(15):4388–4392
Covic A, Kanbay M, Voroneanu L, Turgut F, Serban DN, Serban IL, Goldsmith DJ (2010) Vascular calcification in chronic kidney disease. Clin Sci 119(3):111–121
Wang J, Zhou HY, Salih E, Xu L, Wunderlich L, Gu X, Hofstaetter JG, Torres M, Glimcher MJ (2006) Site-specific in vivo calcification and osteogenesis stimulated by bone sialoprotein. Calcif Tissue Int 79(3):179–189
Zhou Y, Wang J-Y, Feng H, Wang C, Li L, Wu D, Lei H, Li H, Wu L-L (2014) Overexpression of C1q/tumor necrosis factor–related protein-3 promotes phosphate-induced vascular smooth muscle cell calcification both in vivo and in vitro. Arterioscler Thromb Vasc Biol 34(5):1002–1010
Fitzpatrick LA, Severson A, Edwards WD, Ingram RT (1994) Diffuse calcification in human coronary arteries. Association of osteopontin with atherosclerosis. J Clin Investig 94(4):1597–1604
Zebboudj AF, Imura M, Bostrom K (2002) Matrix GLA protein, a regulatory protein for bone morphogenetic protein-2. J Biol Chem 277(6):4388–4394
Diane Proudfoot, Shanahan CM (2006) Molecular mechanisms mediating vascular calcification: role of matrix Gla protein (Review Article). Nephrology 11(5):455–461
Luo G, Ducy P, McKee MD, Pinero GJ, Loyer E, Behringer RR, Karsenty G (1997) Spontaneous calcification of arteries and cartilage in mice lacking matrix GLA protein. Nature 386(6620):78–81
Chen NX, Duan D, O’Neill KD, Wolisi GO, Koczman JJ, LaClair R, Moe SM (2006) The mechanisms of uremic serum-induced expression of bone matrix proteins in bovine vascular smooth muscle cells. Kidney Int 70(6):1046–1053
Lee K-S, Kim H-J, Li Q-L, Chi X-Z, Ueta C, Komori T, Wozney JM, Kim E-G, Choi Je-Young RH-M, Bae S-C (2000) Runx2 is a common target of transforming growth factor β1 and bone morphogenetic protein 2, and cooperation between Runx2 and Smad5 induces osteoblast-specific gene expression in the pluripotent mesenchymal precursor cell line C2C12. Mol Cell Biol 20(23):8783–8792
Lau W, Ling P, Ashwini M, Sharon MG, Cecilia M (2011) Direct effects of phosphate on vascular cell function. Adv Chron Kidney Dis 18(2):105–112
Silver IA, Murrills RJ, Etherington DJ (1988) Microelectrode studies on the acid microenvironment beneath adherent macrophages and osteoclasts. Exp Cell Res 175(2):266–276
Chattopadhyay N, Yano S, Tfelt-Hansen J, Rooney P, Kanuparthi D, Bandyopadhyay S, Ren X, Terwilliger E, Brown EM (2004) Mitogenic action of calcium-sensing receptor on rat calvarial osteoblasts. Endocrinology 145(7):3451–3462
Dinithi Peiris, Ivan Pacheco, Craig Spencer, MacLeod RJ (2007) The extracellular calcium-sensing receptor reciprocally regulates the secretion of BMP-2 and the BMP antagonist Noggin in colonic myofibroblasts. Am J Physiol Gastrointest Liver Physiol 292:753–766
Barradas AMC, Fernandes HAM, Groen N, Chai YC, Schrooten J, van de Peppel J, van Leeuwen JPTM, van Blitterswijk CA, de Boer J (2012) A calcium-induced signaling cascade leading to osteogenic differentiation of human bone marrow-derived mesenchymal stromal cells. Biomaterials 33(11):3205–3215
Costa Jr JR, Abizaid A, Costa R, Feres F, Tanajura LF, Abizaid A, Mattos LA, Staico R, Siqueira D, Sousa A, Bonan GMR, Raoul Sousa JE (2008) Preliminary results of the hydroxyapatite nonpolymer-based sirolimus-eluting stent for the treatment of single de novo coronary lesions: a first-in-human analysis of a third-generation drug-eluting stent system. JACC 1(5):545–551
Acknowledgments
The authors would like to thank Jayesh Betala and Dr. Martine LaBerge at the Department of Bioengineering at Clemson University for the gift of rat aortic smooth muscle cells. This study was partially supported by NIH P20GM103444 grant and the Hunter Endowment to NV.
Conflict of Interest
Pranjal Nahar-Gohad, Neeraj Gohad, Chen-Chih Tsai, Rajendra Bordia, and Naren Vyavahare declare that there is no conflict of interest.
Human and Animal Rights and Informed Consent
No Human Subjects were involved in this study. The Clemson University Institutional Animal Care and Use Committee (IACUC) assures that animal care and use are in compliance with all federal, state and local regulations as well as University policy and assurances. This research study has been carried out in compliance with IACUC regulations.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Nahar-Gohad, P., Gohad, N., Tsai, CC. et al. Rat Aortic Smooth Muscle Cells Cultured on Hydroxyapatite Differentiate into Osteoblast-Like Cells via BMP-2–SMAD-5 Pathway. Calcif Tissue Int 96, 359–369 (2015). https://doi.org/10.1007/s00223-015-9962-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00223-015-9962-z