
Abstract
Handedness is often thought of as a hand “preference” for specific tasks or components of bimanual tasks. Nevertheless, hand selection decisions depend on many factors beyond hand dominance. While these decisions are likely influenced by which hand might show performance advantages for the particular task and conditions, there also appears to be a bias toward the dominant hand, regardless of performance advantage. This study examined the impact of hand selection decisions and workspace location on reaction time and movement quality. Twenty-six neurologically intact participants performed targeted reaching across the horizontal workspace in a 2D virtual reality environment, and we compared reaction time across two groups: those selecting which hand to use on a trial-by-trial basis (termed the choice group) and those performing the task with a preassigned hand (the no-choice group). Along with reaction time, we also compared reach performance for each group across two ipsilateral workspaces: medial and lateral. We observed a significant difference in reaction time between the hands in the choice group, regardless of workspace. In contrast, both hands showed shorter but similar reaction times and differences between the lateral and medial workspaces in the no-choice group. We conclude that the shorter reaction times of the dominant hand under choice conditions may be due to dominant hand bias in the selection process that is not dependent upon interlimb performance differences.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Handedness reflects a functional asymmetry in humans that is apparent in nearly all aspects of daily life, with approximately 90% of the population being right-handed. However, the decision of which hand to use for a given unimanual task or component of a bimanual task is not simply explained by which hand is dominant (Gilbert and Wysocki 1992). In simple reaching tasks, right-handed individuals often select their non-dominant left hand when reaching areas far-left from the body midline. In contrast, they most often choose the dominant right hand for areas surrounding midline and far-right from the midline (Przybyla et al. 2013). This dominant right hand preference for targets close to the body midline is present despite contralateral reaches into the left workspace being less efficient in kinematics, energetics, and time when compared to ipsilateral reaches (Coelho et al. 2013; Liang et al. 2018).
Various factors, including task performance advantages, are attributed to this observed dominant hand preference across the workspace. For example, Przybyla et al. (2013) demonstrated that right-handed participants’ hand selection decisions during a task requiring reaching many targets across the frontal workspace depended on visual feedback conditions. The choice to use the left hand was greater when visual feedback of the cursor was unavailable than when cursor feedback was available. This increase in left-hand choice was associated with an accuracy advantage of the non-dominant hand for most of the workspace under no-vision conditions. We subsequently confirmed that the non-dominant hand has a substantial speed-accuracy advantage early in task practice under no-vision conditions. In contrast, the dominant hand’s advantage emerges only after considerable practice under predictable conditions (Dexheimer and Sainburg 2021). Together, these findings suggested that hand choice in a particular task might be directly linked to hand performance advantages. Further supporting this idea, Stoloff (2011) showed that virtual performance outcomes modulated an individual hand’s selection frequency to favor one hand or the other. Thus, hand selection decisions are likely dependent on a planning process that compares predictions about each hand’s movement outcomes, including parameters such as task accuracy and efficiency. Such a process might reflect Bayesian-like decisions biased by previous outcomes (Rosenbaum and Kornblum 1982; Schweighofer et al. 2015).
While it is likely that hand selection decisions are primarily influenced by which hand might show performance advantages in a particular task under specific conditions, there also seems to be a bias toward the dominant hand, regardless of performance advantage. For example, Philip et al. (2021) recently showed that individuals who experienced a peripheral nerve injury affecting their dominant hand continued to select this hand at rates similar to healthy controls, even when the dominant hand was substantially less dexterous due to nerve damage. This observed separation of performance and preference supports the concept of an overall dominant hand bias for hand selection. Thus, it is reasonable to conclude that the hand-selection process might compare predicted movement outcomes, such as accuracy and mechanical efficiency. Nonetheless, hand dominance also contributes to bias in this process. Poole et al. (2018) investigated the neural process that might underlie this decision process, measuring responses to transcranial magnetic stimulation (TMS) in the motor cortex during a choice reaction time task. They measured motor evoked potential (MEP) amplitude during a warning cue immediately before movement initiation. In trials where participants selected the dominant hand, the corresponding corticospinal system did not show increased excitation immediately prior to hand selection. However, in trials where participants selected their non-dominant hand, increased corticospinal excitation occurred contralaterally, with corticospinal inhibition, measured ipsilaterally, during the ready cue prior to movement. Thus, corticospinal excitability might reflect a dominant hand selection bias, suggesting that greater neural resources are required when the non-dominant hand is selected.
These findings suggest a dominant hand bias that partially separates performance from preference in the hand selection decision-making process before reaching movements. However, the relative roles of dominant hand bias and performance advantages in hand selection decisions remain poorly understood. Performance advantages can conflict with the dominant hand bias when the non-dominant hand shows performance advantages in a task or vice-versa. To address this question, we now measure reaction time before dominant and non-dominant hand reaching movements. Previous research has measured reaction time to evaluate conflicts or consistency with biases in decision-making processes, as reflected by the Simon effect (Iani et al. 2014; Rubichi and Nicoleti 2006; Simon and Rudell 1967). In the seminal work of Simon and Rudell (1967), participants responded to an auditory stimulus by pressing a left or right key as quickly as possible. These stimuli were presented in either their left or right ear and consisted of the words “left” or “right,” indicating which key they should press in response. The results showed that the reaction times were shortest when the verbal stimulus presented matched the ear (i.e., left ear and the word “left”), while they were slowest when the verbal stimulus presented was contralateral to the ear (i.e., left ear and word “right”). This difference in reaction time between ipsilateral vs. contralateral stimuli is known as the Simon effect. Other studies have also demonstrated this reaction time difference with stimuli that do not have spatial representations, such as shapes (Rubichi and Nicoleti 2006) and auditory pitches (Simon and Small Jr 1969). The Simon effect appears the largest in the dominant workspace. For stimuli presented in the right workspace (for right-handers), reaction times of the dominant right hand are significantly shorter than those of the left hand. However, there is a smaller difference between the right and left hand’s reaction times for stimuli presented in the left workspace (Rubichi and Nicoleti 2006; Tagliabue et al. 2007). Thus, in the non-dominant workspace, the difference in reaction times between the hands was lower than in the dominant workspace, supporting a role of a dominant hand bias during hand selection.
Reaction time has also been used to investigate decision-making during the movement planning process. In general, choice ambiguity, or the amplitude of difference between two outcomes, has also been shown to modulate reaction time, where greater ambiguity, or uncertainty, coincides with a linear increase in reaction time (Bartz 1971; Bernstein et al. 1967; Bonnet et al. 2008; Caplan 2002; Wifall et al. 2016). Suppose hand selection decisions involve a comparison of predicted performance outcomes for each hand. In that case, choice ambiguity may modulate reaction time, with longer reaction times occurring in regions of space where predicted performance is similar. In regions where predicted outcomes between the hands show a larger difference, such as in the far-right or far-left workspaces, reaction times may be shorter. Reaction time has also been shown to increase linearly during decision making as the logarithm of the number of required decisions increases, a phenomenon known as Hick’s Law (Hick 1952). Wright et al. (2019) recently demonstrated Hick's Law in the context of hand selection by comparing reaction times among two different response selection tasks. In the stylus task, participants used one hand to move a stylus to a corresponding target location. In contrast, participants placed fingers from both hands on a keyboard and pressed corresponding keys as targets appeared in the key-press task. The authors showed an increase in reaction time consistent with Hick’s Law in the key-press task: as the number of stimulus–response alternatives doubled, reaction time increased by an average of 91 ms for expert typists and 51 ms for novice typists (compared to 50 and 34 ms, respectively, for the stylus task). The authors concluded that the reaction time was substantially longer in the effector selection tasks when compared to tasks where response involved only a single effector. Similarly, Rosenbaum (1980) compared the reaction times across reaches requiring various movement planning decisions: hand selection, reach direction, and reach extent. Participant reaction times increased the most in trials for which a hand selection decision was required compared to trials requiring only a decision on reach direction or extent. Thus, one could expect a longer reaction time in a condition in which there is a choice of which hand to use. Further, reaction time would modulate with choice ambiguity, perhaps due to differences in the predicted performance of each hand.
In summary, the studies above indicate that reaction time represents a planning process that includes decisions about the use of intended effectors in the context of the impending task. However, the extent to which these decisions are weighed by previous performance or rely on a dominant hand bias requires further investigation. In the present study, we have designed a hand selection task to examine this relationship: we distributed target locations across each participant’s left, middle, and right workspace, and we compared reach performance and reaction time across two groups: those selecting which hand to use on a trial-by-trial basis (termed the choice group) and those performing the task with a preassigned hand (the no-choice group). To our knowledge, no previous study has analyzed how reaction times during hand selection decisions are modulated both by differing workspace locations and which hand is selected. Under the hypothesis that a dominant hand bias is present for hand selection decisions, we would expect to see reaction times favor the dominant hand in choice conditions only. We would also expect this bias to persist across all workspace areas. However, under the hypothesis that reaction times of hand selection decisions vary based on similarity or difference between predicted performance of each effector (choice ambiguity), we would expect to see a dependence on spatial location for reaction times in the choice group. Space regions reached most often by one hand (i.e., far-right workspace) should demonstrate reaction times similar to those in the no-choice group, indicating a shorter aspect of total planning time associated with the hand selection process.
Methods
Participants
Twenty-six healthy young adults (18–35 years old) participated in this study. Informed consent was obtained for each participant. All procedures were approved by the institutional review board of The Pennsylvania State University and conducted according to the Declaration of Helsinki ethical guidelines. All participants were right-hand dominant, as confirmed by 100% right-handedness scores on the Edinburgh Handedness Inventory (Oldfield 1971). This questionnaire asks participants to self-report their preferred hand for various daily tasks. Any potential participant scoring less than 100% on this questionnaire was excluded from the study.
Experimental setup
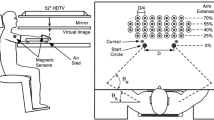
Participants were randomized to complete either the choice or no-choice condition, which are outlined individually in detail below. Both experiments involved a reaching task conducted in a 2-D virtual reality workspace, as shown in Fig. 1A. Each participant’s forearms were supported in the horizontal plane with an air sled to limit arm fatigue and reduce friction, and the participant’s wrist and fingers were immobilized in a resting splint. Participants were unable to see their arms during the task but could view images reflected onto a chin-level mirror from an LCD screen above, including a cursor representing each hand’s fingertip location, as well as a start position and a target. We placed two 6-DOF magnetic sensors (trakSTAR, NDI) on both the left and right dorsal hand and upper arm of each participant, and bony landmarks were digitized relative to each sensor.

(A) Side view of virtual reality workspace set-up; (B) Overhead view of workspace, including target locations and normalized values for target spacing and start circle placement. Note: variable ‘D’ in B (distance between the fingertips when internal elbow angle was 75° and external shoulder angle was 25°) was used to calculate column spacing
Experimental task
Regardless of whether the participant was randomized to experiment #1 or #2, each individual had to place the cursor for each hand into its respective start circle (2.5 cm diameter) to initiate each trial, see Fig. 1B. This start circle was placed at the location of the fingertip cursor when the participant’s internal elbow angle was 75°, and their external shoulder angle was 25°. Once the participant remained in this start circle for 300 ms, an auditory tone signaled the start of the trial, and a target appeared. Targets were 3.5 cm in diameter and would appear in one of 32 different target locations. These locations were normalized to percentages of the participant’s maximal forward reach, as shown in Fig. 1B. Target rows were placed at 25%, 40%, 55%, and 70% of reach, respectively, while target columns were spaced at 25% of the total distance between each start circle (represented by ‘D’ in Fig. 1B). The instruction was to reach the target in one discrete motion as quickly and accurately as possible. Participants had visual feedback of their cursor location for the duration of the reach. Reward points were awarded based on the reach’s final position accuracy, and following the reach, participants would receive feedback on their cursor path and final cursor position. They would then return their cursor to the start circle to begin the next trial. The entire experiment lasted approximately 40 min, and each participant completed 320 total trials, reaching to the 32 different target locations distributed across the left and right workspace in a pseudorandom order.
Participants randomized to experiment #1, the choice condition, were instructed to reach a target displayed at each trial with whatever hand they preferred. Thus, participants selected which hand to use for each reach on a trial-by-trial basis.
Participants randomized to experiment #2, the no-choice condition, were instructed to complete the task with the assigned hand. This hand was predetermined prior to the start of the task. Table 1 outlines participant group sizes. Because previous research has shown that the left hand is selected significantly less frequently than the right in hand selection reaching tasks (Przybyla et al. 2013), we designed the study with a larger sample size for the choice experiment (ten participants) when compared to the no-choice experiment (eight participants using the left hand, eight participants using the right hand) to account for the smaller expected amount of left hand reaches within this group.
Data analysis
Kinematic analysis of reaching movements was coded and executed in a scientific data analysis software environment (Igor Pro 8.04, WaveMetrics). Displacement data of each reach were smoothed by filtering using an 8 Hz low-pass Butterworth filter (3rd-order, dual-pass) and differentiated to yield velocity and acceleration profiles. The first minimum of tangential velocity under 8% of peak velocity defined the movement onset. Similarly, movement offset was defined as the first minimum of tangential velocity occurring after peak velocity that was lower than 8% of peak velocity.
The movement performance was quantified using the following measures: initial direction error (direction error at peak velocity), final position error, deviation from linearity, magnitude of peak velocity, and reaction time. The initial direction error was defined as the angle between the vector from the start circle to the target and the vector from the start position and the 2-D fingertip position at peak velocity. Peak velocity generally occurs mid-way through the duration of the reach in simple point-to-point reaching movements and scales with target distance, so this measure provides information on online trajectory errors (Georgopoulos 1986; Gordon et al. 1994; Messier and Kalaska 1999). The final position error was defined as the 2-D Euclidean distance from the final cursor location to the center of the target at movement offset and provides information on online error corrective mechanisms (Desmurget and Grafton 2000). The deviation from linearity was defined as the ratio between the minor and major axis of the hand path and provides information on curvature of the hand path throughout the entire duration of the reach. The reaction time was calculated as the difference between the “go” stimuli and the movement onset determined from the first minimum of tangential velocity below 8% of peak velocity.
Statistical analysis
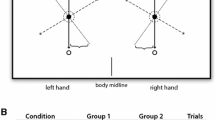
All statistical analyses were done using SAS Version 9.4 (SAS Institute Inc., 2013) where significance was defined as a two-sided alpha ≤ 0.05. Our primary objective was to analyze non-dominant (i.e., left) and dominant hand (i.e., right) performance differences within the choice and no-choice experiments. Because of the large difference in contralateral reach frequency between the right and left hands in the choice condition, we limited our analyses to ipsilateral reaches only (see Fig. 1B). For this reason, the midline row of targets was not included, a restriction that controlled for hand performance differences purely due to reach type (ipsilateral vs. contralateral). To analyze the effect of workspace location on reaction time, we then separated ipsilateral reaches into two subcategories: medial target columns and lateral target columns (see Fig. 2A). Mean performance measures were evaluated with a mixed model analysis using the MIXED procedure in SAS, and included person-specific random intercepts that adjusted for the correlation of the repeated observations contributed by each participant. Explanatory variables in this analysis included: experiment (choice, no choice), hand (left, right), and workspace (lateral, medial).

(A) Example reaching paths for a single subject and target row along with their associated tangential velocity profiles. Other target locations for the task are shown in gray and separated into two categories for analysis: medial target columns and lateral target columns; (B) Reaching frequency for the choice experiment, separated by hand and reach type (contralateral, ipsilateral), along with mean percentage ± SE displayed
Results
Hand selection patterns
Hand selection patterns for the choice experiment and example hand paths are shown in Fig. 2. Example reaches within one participant for the left and the right hand are displayed in Fig. 2A. Figure 2B shows mean percentages of each reach type (ipsilateral, contralateral) performed by each subject. Participants reached with their dominant right hand, on average, 81.9% of the time. Almost two-thirds of those reaches were to ipsilateral target locations. Conversely, participants selected their non-dominant left hand an average of 18.1% of the time, with 93.4% of those being reaches to ipsilateral locations. Thus, participants selected their right hand to reach for left workspace targets just as much, if not more, than their left hand, despite this being a contralateral reach. However, this trend was not observed in the far-right workspace, where contralateral left-hand reaches occurred only sporadically. Due to the small number of contralateral reaches performed by the left hand, we limited the remaining analyses in this study to ipsilateral reaches only (outlined further in Fig. 1B) to appropriately compare performance differences between the hands. These reaches were then separated into two categories for analysis: medial target columns and lateral target columns (Fig. 2A). Of the ipsilateral reaches included in the analysis, 44% and 56% were to lateral and medial target locations, respectively.
Ipsilateral reach performance differences: direction error, position error, deviation from linearity, and peak velocity
Ten right-handed participants completed the choice experiment, performing the entire task with both hands and selecting which hand to use on a trial-by-trial basis, while 16 right-handed participants completed the no-choice experiment, performing the entire task with a preassigned hand, either their right or left. Figure 3 shows the performance means for each experiment, hand, and workspace. Initial direction error was measured as the difference in degrees between the linear path to the target and the cursor’s position at the time of peak velocity (Fig. 3A). The analysis revealed a main effect of workspace [F(1, 42) = 5.6; p = 0.02], along with experiment X workspace [F(1, 40) = 7.0; p = 0.01] and hand X workspace [F(1, 40) = 8.7; p = 0.005] interactions. These interactions reflect significantly higher direction errors in the medial vs. lateral workspace in the choice experiment (1.7 ± 0.4 deg; p = 0.0004), but no difference between the workspaces in the no-choice experiment (p = 0.93). The left hand had significantly higher overall direction errors in the medial workspace when compared to the lateral (1.8 ± 0.5; p = 0.0005), while the right hand performed with similar direction errors across both workspaces (p = 0.76).

Performance variables across the experiments, hands, and ipsilateral workspaces (lateral and medial): (A) Initial direction error; (B) Final position error; (C) Deviation from linearity; (D) Peak velocity
Final position error is displayed in Fig. 3B, and this performance metric was calculated as the 2-D distance from the final cursor location to the center of the intended target at the end of the reach. The analysis demonstrated a main effect of hand [F(1, 42) = 9.9; p = 0.003], with overall higher final position errors observed in left hand reaches (0.28 ± 0.09 cm). There were no significant main effects of experiment (p = 0.71) and workspace (p = 0.95).
Deviation from linearity describes hand path curvature during the reach, with higher values representing more curved reaches, and this performance measure is shown in Fig. 3C. The analysis revealed a main effect of both workspace [F(1, 42) = 13.6; p = 0.0006] and hand [F(1, 42) = 15.5; p = 0.0003], along with a significant hand X workspace interaction [F(1, 41) = 7.0; p = 0.01]. The left hand performed significantly more curved reaches overall in the medial workspace when compared to the lateral (0.03 ± 0.007 au; p < 0.0001), while the right hand performed similarly across the workspaces (p = 0.34). These interlimb differences consistent across both the choice and no-choice experiments have previously been attributed to hemispheric differences in control of limb dynamics (Bagesteiro and Sainburg 2002).
Peak velocity is shown in Fig. 3D. The analysis revealed no significant performance differences between the hands (p = 0.40), but a main effect of both workspace [F(1, 42) = 14.5; p = 0.0004] and experiment [F(1, 42) = 11.3; p = 0.002]. For both the left and right hands, reaches to the lateral workspace were significantly faster when compared to the medial workspace (mean difference: 0.06 ± 0.01 m/s), and reaches in the no-choice experiment were significantly faster than those in the choice experiment (0.15 ± 0.04).
Reaction time differences: movement preparation
Reaction time differences for each experiment are displayed in Fig. 4. The analysis revealed a main effect of both experiment [F(1, 42) = 10.7; p = 0.0021] and hand [F(1, 42) = 22.7; p < 0.0001], along with a significant experiment X hand interaction [F(1, 42) = 11.4; p = 0.0016]. Reaction times for the left hand in the choice group were significantly longer than right hand choice reaches (mean difference ± SE: 43 ± 8 ms; p < 0.0001) and left-hand no-choice reaches (78 ± 15; p < 0.0001). Right hand reaction times were not significantly different between the choice and no-choice experiments (p = 0.24). There was no effect of workspace on reaction time (p = 0.10).

Reaction time of each hand and workspace for both the choice and no-choice experiments
Discussion
In the present study, we conducted two experiments to examine performance and preparation time associated with hand selection in a reaching task. In the first experiment, participants reached various targets with the hand of their choice (choice experiment). In the second experiment, participants reached targets with a preassigned hand (no-choice experiment). Within each experiment, we compared each hand’s performance and reaction time, separated into two ipsilateral workspaces: lateral workspace and medial workspace. Our results support the hypothesis of an underlying dominant arm bias present during hand selection decisions.
Interlimb differences in hand selection behavior
In this study, the pattern of hand selection was substantially biased towards the choice of the dominant right hand. Participants selected the dominant right hand for approximately 80% of total reaches when given a choice. This finding is consistent with previous reports demonstrating the dominant right-hand preference for targets located ipsilaterally and in the midline (Bryden and Roy 2006; Coelho et al. 2013; Przybyla et al. 2013). We also observed the dominant right-hand preference for reaching contralateral targets in about 30% of all reaching movements to the left space, despite contralateral reaches being mechanically less efficient (Liang et al. 2018). In contrast, contralateral reaches with the non-dominant hand happened only sporadically, approximately 1% of the time. This finding is consistent with previously reported non-dominant hand selection rates in similar reaching tasks (Coelho et al. 2013; Przybyla et al. 2013). Furthermore, contralateral reaching with the dominant hand was more frequent than ipsilateral reaching with the non-dominant hand.
Why are reaction times asymmetric under choice conditions?
In the current study, we observed an asymmetry in reaction times for ipsilateral reaching when a hand selection decision was required. This was evident in both the dominant and non-dominant hands. In the choice experiment, the non-dominant hand demonstrated significantly longer reaction times when compared to the dominant hand. In contrast, there were no significant differences between the hands in the no-choice experiment. This asymmetry between the hands under choice and no-choice conditions demonstrates that the overall reaction time required for a hand selection decision cannot simply be attributed to additional preparation requirements regardless of the hand selected or decision itself. We have previously demonstrated, under the same reaching conditions to the same set of targets, that ipsilateral reaches are substantially more efficient in kinematics, energetics, and time, when compared to contralateral reaches (Liang et al. 2018; Przybyla et al. 2013). Thus, the decision within our choice experiment to use the ipsilateral hand, whether dominant or non-dominant, was not surprising. However, why did this decision take longer for the non-dominant hand when compared to the dominant?
While hand selection decisions are likely influenced by which hand might show performance advantages in a particular task or condition, the current findings also support the concept of a dominant hand bias. In the choice experiment, we observed a longer reaction time in the non-dominant hand but no significant differences in this reaction time across the lateral and medial workspaces. In contrast, the no-choice experiment revealed an overall difference in reaction time when compared to the choice experiment, but no difference between the hands or workspaces. We propose that a dominant hand bias may explain this observed asymmetry within the hand selection process itself, which causes increased reaction times when the controller undergoes the process of selecting the non-dominant hand.
This bias may also explain the performance differences observed in the current study. Contralateral reaching by the dominant hand occurred for approximately 30% of total reaches in the choice experiment, despite these reaches being less mechanically efficient when compared to non-dominant hand ipsilateral reaching (Coelho et al. 2013; Liang et al. 2018; Przybyla et al. 2013). Suppose hand selection decisions reflect a comparison of predicted task performance outcomes for each hand, which is modulated by a dominant hand bias. In that case, we might expect the dominant hand’s selection under conditions where the opposite hand’s performance is more advantageous in mechanical efficiency, accuracy, or speed. Liang et al. (2018) previously showed that, in a reaching task, selection of the dominant hand for contralateral reaches increased when participants were asked to complete reaches with rising demands for memory and visual search. Because these reaches across the midline are less mechanically and kinematically efficient than ipsilateral reaches with the non-dominant hand, those findings suggested that increased cognitive load reduced the efficiency of the hand-decision process, reverting decisions to the influence of a simple dominant hand bias. This bias might also explain the large number of contralateral reaches we observed in our current study under choice conditions. A more profound effect of dominant hand bias in hand selection was recently reported by Philip et al. (2021), who showed that individuals with peripheral nerve injury continue to select their injured dominant hand for a block building task at rates similar to healthy controls, despite the non-dominant hand showing substantially greater dexterity in standardized tests of hand function. This study emphasized the importance of understanding hand-selection conditions for clinical conditions affecting unilateral hand and hand function.
In the current study, we also observed significant interlimb differences in performance across the workspaces. The left hand reached with higher initial direction errors and greater deviations from linearity for the medial workspace when compared to the lateral. The right hand did not demonstrate significant differences in these performance measures between the medial and lateral workspaces. We have previously shown that, in a reaching task to target locations requiring increasing amounts of shoulder joint excursion, the non-dominant arm demonstrated greater deviations in linearity that scaled with increasing shoulder joint excursion. The dominant arm’s trajectory linearity remained consistent regardless of target location (Bagesteiro and Sainburg 2002; Sainburg and Kalakanis 2000). We have proposed that the dominant hemisphere is specialized for the prediction of interaction torques between limb segments, while the non-dominant hemisphere is specialized for impedance control mechanisms resulting in accurate steady-state positioning (Bagesteiro and Sainburg 2002, 2003; Sainburg 2002). These specializations may have contributed to the patterns of interlimb differences in performance across the workspaces observed in the present study: the non-dominant hand reached with higher initial direction and final position errors and greater deviations from linearity to target locations requiring greater shoulder joint excursion, while the dominant hand demonstrated consistent performance across the workspaces.
It is important to note that these interlimb differences in performance did not vary with reaction time. Previous research has detailed a relationship between reaction time and task accuracy, such that faster reaction times coincide with lower rates of task-specific accuracy. In contrast, longer reaction times predict higher rates of accuracy. This phenomenon has been demonstrated in a variety of tasks, such as button pressing (Spieser et al. 2017), reading comprehension/word recognition (Rinkenauer et al. 2004), and visuomotor adaptations for reaching (Fernandez-Ruiz et al. 2011). Thus, we might expect that the increase in reaction time for left-hand reaches in the choice experiment might coincide with more advantageous task performance when compared to the no-choice experiment. However, we did not observe this pattern in the present study. Overall, the non-dominant left hand reached with significantly higher initial direction error and deviation from linearity in the medial vs. lateral workspace but showed no difference in reaction time between these workspaces. The dominant right hand performed with consistent levels of final position error, initial direction error, and deviation from linearity across the workspaces, along with consistent reaction times. While previous research has suggested that movements for each hand are planned in parallel and “compete” for the selection (Cisek and Kalaska 2010; Fitzpatrick et al. 2019; Hirayama et al. 2021), the findings of the present study demonstrate a reaction time difference during the hand selection process that suggests the non-dominant hand takes a longer time to be selected for the task. Although speculative, we suggest that dominant hand bias may disrupt or delay the motor planning process for the non-dominant hand, and this may explain both the reaction time and performance differences observed in the current study: the choice condition left hand showed a disrupted relationship between reaction time and task-specific performance when compared to the no-choice left hand.
Under the concept of choice ambiguity, we would have expected to observe a modulation in reaction time for hand selection decisions based on the ambiguity of predicted outcomes (i.e., Hick’s Law). For example, in the choice group, decisions with low ambiguity might be workspace areas where predicted performance for the right and left hands are very different, such as for a target in the far-left or far-right workspace. Therefore, we might expect shorter reaction times for this hand selection decision. Areas of the workspace with similar predicted performance, such as for a target location along the midline, may have higher choice ambiguity and coincide with relatively longer reaction times. In the present study, we observed an overall difference in reaction time between the hands in the choice group, but this did not vary with the region of space or any specific performance variable we measured. It is possible that the weighing of predicted outcomes in the decision-making process might involve a variable or set of variables not reported in this study, such as mechanical efficiency or the end-state comfort effect. For example, Coelho et al. (2014) has previously demonstrated the impact of the end-state comfort on a hand selection task. Participants were asked to grasp and position one of their chosen hands onto a dowel for manipulation into a target. They selected their non-dominant hand to position in a thumbs-up grasp more frequently than they used their dominant hand in a thumbs-down grasp, a position the authors deemed to have “lower” end-state comfort than the thumbs-up grasp. The authors concluded that the end-state comfort might be weighed and prioritized over hand dominance during hand selection decisions, depending on task demands. Consistent with this conclusion, it has also previously been suggested that action selection, in general, may dynamically prioritize the amount in which different variables, such as the end-state comfort, inform choice depending on the specific task demands (Rosenbaum et al. 2001). In the present study, while we did not observe hand selection and performance patterns consistent with the concept of choice ambiguity for the variables we measured, further investigation is required to determine the extent to which choice ambiguity in hand selection decision-making affects reaction time.
Reaction time costs and the emergence of lateralization
Regardless of the exact mechanisms surrounding hand selection, the phylogenetic emergence of handedness may have arisen, in part, as a mechanism to reduce planning time. Specialization of each hemisphere/limb system could bias this process for different aspects of unimanual tasks (i.e., holding projectiles with the non-dominant hand while throwing with the dominant hand) or bimanual tasks (i.e., stabilizing a stone with the non-dominant hand while hammering with the dominant hand). Our laboratory has previously reported performance differences across the hands, leading to a bi-hemispheric model of motor control termed the dynamic dominance hypothesis (Bagesteiro and Sainburg 2003; Mutha et al. 2013; Sainburg 2002; Yadav and Sainburg 2011, 2014). This hypothesis proposes that the dominant and non-dominant hemispheres are specialized for different but complementary aspects of motor control. Specifically, the dominant hemisphere and its contralateral arm are specialized in predictive control of intersegmental dynamics, which results in straighter, more mechanically efficient movements (Bagesteiro and Sainburg 2002; Przybyla et al. 2012), while the non-dominant hemisphere/arm is specialized in impedance control, which results in better performance under unpredictable movement perturbations when compared with the dominant arm (Bagesteiro and Sainburg 2003; Dexheimer and Sainburg 2021; Schabowsky et al. 2007). In the absence of these performance asymmetries, deciding which hand to use for each function could be taxing and result in relatively long planning delays and a more significant cognitive load for relatively simple tasks. Indeed, asymmetry in motor function does not seem to be a necessary result of asymmetry in hemispheric motor control processes. Language is a strongly lateralized process in which the behavioral expression of verbal language does not express the separation of functions reflected by each hemisphere (i.e., prosody and emotional expression by the right hemisphere and lexicon and syntax by the left hemisphere). Our previous research suggests that the functional neuroanatomy that underlies hemispheric specializations for motor control is reflected by tertiary cortical areas that are not directly connected with spinal cord ascending and descending systems. Thus, it should be possible for each arm to reflect contributions from each hemisphere. Why, then, should hemispheric lateralization for motor control lead to behavioral asymmetries? We now suggest that this behavioral asymmetry may have conferred substantial advantages in hand selection during the course of evolution.
This hemispheric lateralization may have allowed for parallel processing in each hemisphere, thus optimizing neural resources for each process and, in turn, reducing cognitive load and planning time. Similar neurobehavioral asymmetries have been elaborated for a large range of animals and behaviors, including affective, communication, prey–predator behavior, and visual processing domains, among others (MacNeilage et al. 2009). The ability to plan different aspects of control in a parallel rather than in series could have been advantageous to survival during the course of evolution (Rogers 2000; Vallortigara and Rogers, 2005). For example, an animal would benefit from the timely discrimination between dangerous and non-threatening stimuli while simultaneously developing an appropriate response to that stimulus. In the context of motor control, fMRI studies have shown bilateral activation of many relevant brain areas during both unimanual and bimanual movement, including the thalamus, premotor cortex, and posterior parietal cortex (Culham et al. 2003; Fitzpatrick et al. 2019; Kertzman et al. 1997; Vingerhoets 2014).
Furthermore, lesion studies have shown that specific movement deficits manifest bilaterally, despite the unilateral nature of the lesion, perhaps indicating a disruption in this parallel processing (Maenza et al. 2021; Mani et al. 2013; Noskin et al. 2008; Schaefer et al. 2007). Our model of motor lateralization is a bi-hemispheric model that is consistent with the idea that each hemisphere contributes different processes to motor planning and control, as reflected by previously published simulation studies (Jayasinghe et al. 2020; Yadav and Sainburg 2011, 2014). We now suggest that, during the course of evolution, reduction in reaction time through hand specialization and selection bias may have aided in the performance of many primitive tasks, such as rapid escape and combative behaviors.
Conclusion
The present study investigated the processes that underlie hand selection in motor behaviors. We observed differences in reaction time between the dominant and non-dominant hands under choice conditions but no difference in reaction time between the hands under no-choice conditions. While the exact mechanisms behind this asymmetry remain unknown, we propose that a dominant hand bias for hand selection may have contributed to increased non-dominant hand reaction times observed during reaching under the hand-choice condition. Further investigation is necessary to resolve the extent to which a dominant hand bias interacts with predicted performance advantages to influence the hand selection decision-making process.
Data availability
The datasets generated and analyzed during the current study are available from the corresponding author on reasonable request.
References
Bagesteiro LB, Sainburg RL (2002) Handedness: dominant arm advantages in control of limb dynamics. J Neurophysiol 88(5):2408–2421. https://doi.org/10.1152/jn.00901.2001
Bagesteiro LB, Sainburg RL (2003) Nondominant arm advantages in load compensation during rapid elbow joint movements. J Neurophysiol 90(3):1503–1513. https://doi.org/10.1152/jn.00189.2003
Bartz A (1971) Reaction time as a function of stimulus uncertainty on a single trial. Percept Psychophys 9(1):95–96. https://doi.org/10.3758/BF03213036
Bernstein IH, Schurman DL, Forester G (1967) Choice reaction time as a function of stimulus uncertainty, response uncertainty, and behavioral hypotheses. J Exp Psychol 74(4, Pt.1):517–524. https://doi.org/10.1037/h0021279
Bonnet C, Ars JF, Ferrer SE (2008) Reaction times as a measure of uncertainty. Psicothema 20(1):43–48
Bryden PJ, Roy EA (2006) Preferential reaching across regions of hemispace in adults and children. Dev Psychobiol 48(2):121–132. https://doi.org/10.1002/dev.20120
Caplan S (2002) The effect of uncertainty on reaction time. Proc Human Factors Ergonomics Soc Annu Meeting 46(8):805–809. https://doi.org/10.1177/154193120204600810
Cisek P, Kalaska JF (2010) Neural mechanisms for interacting with a world full of action choices. Annu Rev Neurosci 33(1):269–298. https://doi.org/10.1146/annurev.neuro.051508.135409
Coelho CJ, Przybyla A, Yadav V, Sainburg RL (2013) Hemispheric differences in the control of limb dynamics: a link between arm performance asymmetries and arm selection patterns. J Neurophysiol. https://doi.org/10.1152/jn.00885.2012
Coelho CJ, Studenka BE, Rosenbaum DA (2014) End-state comfort trumps handedness in object manipulation. J Exp Psychol Hum Percept Perform 40(2):718–730. https://doi.org/10.1037/a0034990
Culham JC, Danckert SL, DeSouza JFX, Gati JS, Menon RS, Goodale MA (2003) Visually guided grasping produces fMRI activation in dorsal but not ventral stream brain areas. Exp Brain Res. https://doi.org/10.1007/s00221-003-1591-5
Desmurget M, Grafton S (2000) Forward modeling allows feedback control for fast reaching movements. Trends Cogn Sci 4(11):423–431. https://doi.org/10.1016/S1364-6613(00)01537-0
Dexheimer B, Sainburg R (2021) When the non-dominant arm dominates: the effects of visual information and task experience on speed-accuracy advantages. Exp Brain Res. https://doi.org/10.1007/s00221-020-06011-6
Fernandez-Ruiz J, Wong W, Armstrong IT, Flanagan JR (2011) Relation between reaction time and reach errors during visuomotor adaptation. Behav Brain Res 219(1):8–14. https://doi.org/10.1016/j.bbr.2010.11.060
Fitzpatrick AM, Dundon NM, Valyear KF (2019) The neural basis of hand choice: an fMRI investigation of the Posterior Parietal Interhemispheric Competition model. Neuroimage. https://doi.org/10.1016/j.neuroimage.2018.10.039
Georgopoulos AP (1986) On reaching. Annu Rev Neurosci 9(1):147–170
Gilbert AN, Wysocki CJ (1992) Hand preference and age in the United States. Neuropsychologia. https://doi.org/10.1016/0028-3932(92)90065-T
Gordon J, Ghilardi MF, Cooper SE, Ghez C (1994) Accuracy of planar reaching movements: II. Systematic extent errors resulting from inertial anisotropy. Exp Brain Res 99(1):112–130. https://doi.org/10.1007/BF00241416
Hick WE (1952) On the rate of gain of information. Q J Exp Psychol 4(1):11–26. https://doi.org/10.1080/17470215208416600
Hirayama K, Koga T, Takahashi T, Osu R (2021) Transcranial direct current stimulation of the posterior parietal cortex biases human hand choice. Sci Rep 11(1):204. https://doi.org/10.1038/s41598-020-80611-8
Iani C, Milanese N, Rubichi S (2014) The influence of prior practice and handedness on the orthogonal Simon effect. Front Psychol. https://doi.org/10.3389/fpsyg.2014.00039
Jayasinghe SAL, Sarlegna FR, Scheidt RA, Sainburg RL (2020) The neural foundations of handedness: insights from a rare case of deafferentation. J Neurophysiol. https://doi.org/10.1152/jn.00150.2020
Kertzman C, Schwarz U, Zeffiro TA, Hallett M (1997) The role of posterior parietal cortex in visually guided reaching movements in humans. Exp Brain Res. https://doi.org/10.1007/PL00005617
Liang J, Wilkinson K, Sainburg RL (2018) Is Hand selection modulated by cognitive–perceptual load? Neuroscience. https://doi.org/10.1016/j.neuroscience.2017.11.005
MacNeilage PF, Rogers LJ, Vallortigara G (2009) Origins of the left & right brain. Sci Am. https://doi.org/10.1038/scientificamerican0709-60
Maenza C, Wagstaff DA, Varghese R, Winstein C, Good DC, Sainburg RL (2021) Remedial training of the less-impaired arm in chronic stroke survivors with moderate to severe upper-extremity paresis improves functional independence: a pilot study. Front Hum Neurosci 15:133
Mani S, Mutha PK, Przybyla A, Haaland KY, Good DC, Sainburg RL (2013) Contralesional motor deficits after unilateral stroke reflect hemisphere-specific control mechanisms. Brain 136(4):1288–1303. https://doi.org/10.1093/brain/aws283
Messier J, Kalaska JF (1999) Comparison of variability of initial kinematics and endpoints of reaching movements. Exp Brain Res 125(2):139–152. https://doi.org/10.1007/s002210050669
Mutha PK, Haaland KY, Sainburg RL (2013) Rethinking motor lateralization: specialized but complementary mechanisms for motor control of each arm. PLoS ONE. https://doi.org/10.1371/journal.pone.0058582
Noskin O, Krakauer JW, Lazar RM, Festa JR, Handy C, O’Brien KA, Marshall RS (2008) Ipsilateral motor dysfunction from unilateral stroke: implications for the functional neuroanatomy of hemiparesis. J Neurol Neurosurg Psychiatry. https://doi.org/10.1136/jnnp.2007.118463
Oldfield RC (1971) The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia. https://doi.org/10.1016/0028-3932(71)90067-4
Philip BA, Thompson MR, Baune NA, Hyde M, Mackinnon SE (2021) Failure to compensate: nerve injury patients use their injured dominant hand, even when their non-dominant is more dexterous. Arch Phys Med Rehabil. https://doi.org/10.1016/j.apmr.2021.10.010
Poole BJ, Mather M, Livesey EJ, Harris IM, Harris JA (2018) Motor-evoked potentials reveal functional differences between dominant and non-dominant motor cortices during response preparation. Cortex 103:1–12. https://doi.org/10.1016/j.cortex.2018.02.004
Przybyla A, Good DC, Sainburg RL (2012) Dynamic dominance varies with handedness: reduced interlimb asymmetries in left-handers. Exp Brain Res 216(3):419–431. https://doi.org/10.1007/s00221-011-2946-y
Przybyla A, Coelho CJ, Akpinar S, Kirazci S, Sainburg RL (2013) Sensorimotor performance asymmetries predict hand selection. Neuroscience 228:349–360. https://doi.org/10.1016/j.neuroscience.2012.10.046
Rinkenauer G, Osman A, Ulrich R, Müller-Gethmann H, Mattes S (2004) On the locus of speed-accuracy trade-off in reaction time: inferences from the lateralized readiness potential. J Exp Psychol Gen 133(2):261–282. https://doi.org/10.1037/0096-3445.133.2.261
Rogers LJ (2000) Evolution of hemispheric specialization: advantages and disadvantages. Brain Lang. https://doi.org/10.1006/brln.2000.2305
Rosenbaum DA (1980) Human movement initiation: specification of arm, direction, and extent. J Exp Psychol Gen 109(4):444–474. https://doi.org/10.1037/0096-3445.109.4.444
Rosenbaum DA, Kornblum S (1982) A priming method for investigating the selection of motor responses. Acta Physiol (oxf) 51(3):223–243. https://doi.org/10.1016/0001-6918(82)90036-1
Rosenbaum DA, Meulenbroek RJ, Vaughan J, Jansen C (2001) Posture-based motion planning: applications to grasping. Psychol Rev 108(4):709–734. https://doi.org/10.1037/0033-295X.108.4.709
Rubichi S, Nicoleti R (2006) The Simon effect and handedness: evidence for a dominant-hand attentional bias in spatial coding. Percept Psychophys 68(7):1059–1069. https://doi.org/10.3758/BF03193709
Sainburg RL (2002) Evidence for a dynamic-dominance hypothesis of handedness. Exp Brain Res. https://doi.org/10.1007/s00221-001-0913-8
Sainburg RL, Kalakanis D (2000) Differences in control of limb dynamics during dominant and nondominant arm reaching. J Neurophysiol. https://doi.org/10.1152/jn.2000.83.5.2661
SAS Institute Inc. (9.4) (2013) SAS User’s Guide—Procedures. SAS Institute Inc., Cary, NC
Schabowsky CN, Hidler JM, Lum PS (2007) Greater reliance on impedance control in the nondominant arm compared with the dominant arm when adapting to a novel dynamic environment. Exp Brain Res 182(4):567–577. https://doi.org/10.1007/s00221-007-1017-x
Schaefer SY, Haaland KY, Sainburg RL (2007) Ipsilesional motor deficits following stroke reflect hemispheric specializations for movement control. Brain. https://doi.org/10.1093/brain/awm145
Schweighofer N, Xiao Y, Kim S, Yoshioka T, Gordon J, Osu R (2015) Effort, success, and nonuse determine arm choice. J Neurophysiol. https://doi.org/10.1152/jn.00593.2014
Simon JR, Rudell AP (1967) Auditory S-R compatibility: the effect of an irrelevant cue on information processing. J Appl Psychol 51(3):300–304. https://doi.org/10.1037/h0020586
Simon JR, Small A Jr (1969) Processing auditory information: interference from an irrelevant cue. J Appl Psychol 53(5):433
Spieser L, Servant M, Hasbroucq T, Burle B (2017) Beyond decision! Motor contribution to speed–accuracy trade-off in decision-making. Psychon Bull Rev 24(3):950–956. https://doi.org/10.3758/s13423-016-1172-9
Stoloff RH (2011) Effect of reinforcement history on hand choice in an unconstrained reaching task. Front Neurosci. https://doi.org/10.3389/fnins.2011.00041
Tagliabue M, Vidotto G, Umiltà C, Altoè G, Treccani B, Spera P (2007) The measurement of left—right asymmetries in the Simon effect: a fine-grained analysis. Behav Res Methods 39(1):50–61. https://doi.org/10.3758/BF03192843
Vallortigara G, Rogers LJ (2005) Survival with an asymmetrical brain: advantages and disadvantages of cerebral lateralization. Behav Brain Sci. https://doi.org/10.1017/S0140525X05000105
Vingerhoets G (2014) Contribution of the posterior parietal cortex in reaching, grasping, and using objects and tools. Front Psychol. https://doi.org/10.3389/fpsyg.2014.00151
Wifall T, Hazeltine E, Toby Mordkoff J (2016) The roles of stimulus and response uncertainty in forced-choice performance: An amendment to Hick/Hyman Law. Psychol Res 80(4):555–565. https://doi.org/10.1007/s00426-015-0675-8
Wright CE, Marino VF, Chubb C, Mann D (2019) A model of the uncertainty effects in choice reaction time that includes a major contribution from effector selection. Psychol Rev 126(4):550–577. https://doi.org/10.1037/rev0000146
Yadav V, Sainburg RL (2011) Motor lateralization is characterized by a serial hybrid control scheme. Neuroscience. https://doi.org/10.1016/j.neuroscience.2011.08.039
Yadav V, Sainburg RL (2014) Handedness can be explained by a serial hybrid control scheme. Neuroscience. https://doi.org/10.1016/j.neuroscience.2014.08.026
Funding
The project described was supported by National Institutes of Health Award Number R01HD059783 to RLS, along with the National Center for Advancing Translational Sciences, Grant TL1 TR002016, and Grant UL1 TR002014. This research was also supported by an endowment from Dorothy Foehr Huck and J. Lloyd Huck Institutes for Life Sciences at Penn State University to RLS. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Additional information
Communicated by Bill J Yates.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Dexheimer, B., Przybyla, A., Murphy, T.E. et al. Reaction time asymmetries provide insight into mechanisms underlying dominant and non-dominant hand selection. Exp Brain Res 240, 2791–2802 (2022). https://doi.org/10.1007/s00221-022-06451-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-022-06451-2