
Abstract
In addition to the action potentials generated by the ongoing activation of single dorsal horn neurons in the anesthetized cat, we often recorded small negative field potentials with a fast-rising phase and a slow decay (dIFPs). These potentials could be separated in different classes, each with a specific and rather constant shape and amplitude. They were largest in spinal laminae III–V and gradually faded at deeper locations, without showing the polarity reversal displayed at these depths by the focal potentials produced by stimulation of muscle and cutaneous afferents. We propose that the dIFPs are postsynaptic field potentials generated by strongly coupled sets of dorsal horn neurons displaying a spatial orientation that generates closed field potentials in response to stimulation of high-threshold cutaneous and muscle afferents. These neuronal sets could form part of the spinal inhibitory circuitry that mediates presynaptic inhibition and Ib non-reciprocal postsynaptic inhibition and could be involved in the sensory–motor transformations activated by stimulation of high-threshold cutaneous afferents.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Previous studies made in anesthetized cats have shown that the magnitude of the ongoing synchronization displayed by the different sets of dorsal horn neurons has a non-random configuration suggesting structured patterns of functional connectivity between the different neuronal sets. Quite interestingly, this configuration is changed, also in a non-random manner, during the state of central sensitization induced by the intradermic injection of capsaicin, and is transiently reverted to its pre-capsaicin condition by the systemic injection of small amount of lidocaine (Contreras-Hernández et al. 2015, 2018 and Martin et al. 2019).
While performing these studies, we recorded in the dorsal horn, in addition to the ongoing action potentials generated by individual neurons, a substantial number of spontaneous negative field potentials with a fast-rising phase followed by a slow decay (henceforth termed “discrete intraspinal field potentials” or dIFPs).
The spontaneous dIFPs recorded in a given spinal location could be separated in different classes, each with a particular shape and amplitude. We could establish in few experiments that the dIFPs were inhibited by electrical stimulation of low-threshold cutaneous afferents and activated by stimulation of high-threshold (Aδ) and of group II muscle afferents. We found in addition that most dIFPs were preceded by a small brief volley similar to the incoming afferent terminal potential (ATP) that precedes the postsynaptic field potentials recorded in the motor nucleus following stimulation of single Ia afferents (Munson and Sypert 1979a, b; Sypert et al. 1980) and in the cord dorsum by stimulation of single cutaneous afferents (Koerber et al. 1990). Quite interestingly, each class of dIFPs displayed the same amplitude and shape, regardless of whether or not it was preceded by ATPs. This suggested that most of the dIFPs were generated by discrete populations of strongly interconnected neurons activated by incoming action potentials from muscle and/or cutaneous afferents. Quite interestingly, a significant fraction of the dIFPs occurred in synchrony with the ongoing negative–positive cord dorsum potentials (npCDPs), a finding suggesting that the neurons producing the dIFPs could form part of the circuitry that modulates transmission along the spinal pathways mediating PAD and presynaptic inhibition (Contreras-Hernández et al. 2015).
A systematic analysis of the spontaneous and of the sensory evoked dIFPs recorded at different spinal depths showed that the dIFPs generated by a given set of neurons were largest in spinal laminae III–V and gradually faded at deeper locations, without showing the polarity reversal displayed by the focal potentials generated by stimulation of cutaneous and muscle nerves. This suggested further that the neuronal sets that produce the dIFPs display a spatial orientation that generates closed field potentials, as expected for neurons with a spatially confined distribution (see Lorente de No 1947 and Klee and Rall 1977).
Based on the present observations, we propose that the dIFPs are postsynaptic field potentials generated by tightly coupled, spatially confined sets of dorsal horn neurons responding to the ongoing activity elicited in single II muscle and Aδ cutaneous afferents. These neurons could form part of the dorsal horn neuronal ensembles that mediate the presynaptic and postsynaptic inhibitory actions involved in specific sensorimotor transformations (see Schouenborg 2002, 2008; Levinsson et al. 2002; Bizzi and Cheung 2013); Previous observations were published in abstract form (Contreras–Hernández et al. 2011).
Methods
Ethical approval
Cats were bred and housed under veterinarian supervision at the Institutional Animal Care unit (SAGARPA permission AUT-B-C-0114-007). All experiments were approved by the Institutional Ethics Committee for Animal Research (Protocol no. 126–03) and comply with the ethical policies and regulations of The Journal of Physiology (Drummond 2009; Grundy 2015) and of the National Institutes of Health (Bethesda, MD, USA; Animal Welfare Assurance no. A5036-01). The Guide for the Care and Use of Laboratory Animals (National Research Council 2010) was followed in all cases.
General procedures
Preparation
The present set of observations was obtained from 14 young cats of either sex used in a previous study (Contreras-Hernández et al. 2015). These animals were initially anesthetized with pentobarbitone sodium (40 mg kg–1 I.P.). The carotid artery, radial vein, trachea and urinary bladder were cannulated. Additional doses of pentobarbitone sodium were given I.V. (5 mg kg–1 h–1) to maintain an adequate level of anesthesia, tested by assessing that withdrawal reflexes were absent, that the pupils were constricted, and that arterial blood pressure was between 100 and 120 mm Hg. A solution of 100 mM of sodium bicarbonate with 5% glucose was given I.V. (0.03 ml min−1) to prevent acidosis (Rudomin et al. 2007). When necessary, 10% dextran was administered to keep blood pressure above 100 mm Hg.
The lumbosacral and low thoracic spinal segments were exposed by laminectomy and opening of the dura mater. In all experiments, the left superficial peroneal (SP) and in three experiments, the posterior biceps and semitendinosus (PBSt) and the gastrocnemius (GS) nerves were dissected free, left in continuity and prepared for stimulation. After the main surgical procedures, the animals were transferred to a stereotaxic metal frame allowing immobilization of the head and spinal cord, and pools were made with the skin flaps to prevent desiccation of the exposed tissues. The pools were filled with paraffin oil and maintained between 36 and 37 °C by means of radiant heat.
Subsequently, neuromuscular block was induced with pancuronium bromide (0.1 mg kg–1), and the animals were artificially ventilated. The tidal volume was adjusted to maintain 4% CO2 in the expired air. During neuromuscular block, the adequacy of anesthesia was ensured with supplementary doses of anesthetic (2 mg kg–1 h–1) and by assessing that the pupils remained constricted, and that heart rate and blood pressure were not changed following a noxious stimulus (paw pinch).
At the end of the experiments, the animals were euthanized with an overdose of pentobarbitone. The spinal cord was removed, leaving the recording micropipette(s) in place. After fixation and dehydration, the lumbosacral segments were placed in a solution of methyl salicylate for clearing; subsequently, they were cut transversely to obtain a section containing the micropipette(s) and photographed. Location of the recording sites was estimated from the depth readings made during the experiment.
Recording and stimulation
Glass micropipettes filled with 2 M NaCl (2–6 MΩ) were used to record intraspinal field potentials as well as extracellular neuronal action potentials by means of an Axoclamp 2B amplifier (Molecular Devices, Sunnyvale, CA, USA), filters set from 0.1 to 5 kHz. Usually one and in some experiments two recording micropipettes were inserted in the left side of the L5–L6 segments and gradually displaced downward within the dorsal horn until recording spontaneous action potentials and slow field potentials (dIFPs).
Cord dorsum potentials (CDPs) were simultaneously recorded by means of silver ball electrodes placed on the surface of the L4–L7 segments on both sides of the spinal cord against an indifferent electrode inserted in the paravertebral muscles, using AC amplifiers with filters set from 0.3 to 1 kHz. Intraspinal and cord dorsum potentials were sampled at 10 kHz, visualized online, and stored digitally for offline analysis using software written in MatLab (Mathworks, Natick, MA, USA) and Octave (Eaton et al. 2018).
In several experiments the SP, as well as the PBSt and GS nerves were stimulated with single pulses of 0.1 ms duration with strengths ranging from 1.15 to 2.0xT and from 2 to 5xT, respectively.
Sorting of neuronal spikes and dIFPs
We selected for analysis the ongoing action potentials and dIFPs recorded in the left dorsal horn in segments L5–L6 within laminae III–VI. We used principal component analysis (PCA) to group the recorded potentials in separate classes according to their amplitude and shape (see Lewicki 1998).
Selection of spontaneous nCDPs and npCDPs
In previous work, we examined the functional relations between the ongoing activity of single dorsal horn neurons and the negative and negative–positive cord dorsum potentials (nCDPs and npCDPs, respectively; see Contreras-Hernández et al. 2015). We now examined the relations of the dIFPs with the ongoing nCDPs and npCDPs. The method for selection and classification of the nCDPs and npCDPs is detailed in a previous publication from our laboratory (Contreras-Hernández et al. 2015). Briefly, ongoing CDPs recorded in a specific segment that exceeded a preset amplitude were selected for offline analysis. The discrimination level was adjusted to include a wide range of small and large CDPs. Subsequently, each CDP was passed through nCDP-shaped and npCDP-shaped dot templates. The selected CDPs were considered as either nCDPs or npCDPs depending on their best fit to the templates. To ensure high quality in the selection of nCDPs and npCDPs, a final visual inspection was made to reject from the sample those CDPs with other shapes.
Coefficients of variation
Quite often, the neuronal action potentials and dIFPs recorded with the same intraspinal micropipette showed considerable variations in amplitude and duration. We used the coefficient of variation (CV) as normalized measure of the dispersion of the amplitude as well as of the duration (half-height width) for these two types of neuronal potentials.
The CV is defined as follows: \(CV = {\raise0.7ex\hbox{$\sigma $} \!\mathord{\left/ {\vphantom {\sigma \mu }}\right.\kern-\nulldelimiterspace} \!\lower0.7ex\hbox{$\mu $}},\) where σ and µ represent the standard deviation and the mean.
Coefficients of correlation
As in previous work (Chávez et al. 2012), the changes in correlation were estimated by means of the Pearson correlation coefficient (ρ), as follows
where X = {xi} and Y = {yi} are records of paired sets of CDPs and/or IFPs.
Results
The dIFPs are produced by synchronous activation of small sets of dorsal horn neurons
As mentioned in the Introduction, in addition to the ongoing action potentials generated by individual neurons located in the deep dorsal horn, we recorded a substantial number of spontaneous small negative field potentials with a fast-rising phase followed by a slow decay. This section provides a more detailed account on the basic features of these potentials.
The traces displayed in Fig. 1Aa show the cord dorsum potentials (CDPs) recorded in the left side of the L4–L7 segments as well as the intraspinal potentials recorded at the same time with a glass micropipette placed in the left L6 dorsal horn at 1150 µm depth (Fig. 1Ab). The intraspinal potentials that exceeded the discrimination threshold (100 µV, horizontal dotted line) were selected and sorted using PCA (see Methods). Figure 1B shows the representation of these potentials in the space of the three first principal components, where it can be seen that they were grouped in three main clusters (s1, s2 and s3), together with some dispersed undefined potentials (us).

Sorting of intraspinal potentials using principal component analysis (PCA). Aa, ongoing cord dorsum potentials recorded in the left side of the L4–L7 segments. Ab, simultaneous recording of action potentials and slow neuronal fields (dIFPs) in the dorsal horn at 1150 µm depth, within Rexed’s lamina IV. Intraspinal potentials exceeding a predetermined threshold (see horizontal dashed line) were selected for further processing with PCA. B, PCA space representation of the selected potentials. Each point is plotted according to its covariance with the first three principal components. The selected potentials were grouped in three well-defined clusters (s1, s2, and s3) and in another dispersed, undefined group (us), as indicated. C, superposed traces of the potentials included in the s1, s2 and s3 clusters. Means are shown in white. In all figures, negativity is downward in the intraspinal recordings. Further explanations in text
The potentials that contributed to the s1 and s2 clusters displayed in Fig. 1C were rather brief, biphasic and had the features of the extracellular recordings of action potentials generated by single neurons (Gold et al. 2006; Buzsáki et al. 2012; Lindén et al. 2014). In contrast, the potentials included in the s3 cluster (dIFPs) had an initial negative fast-rising phase of 0.98 ± 0.15 ms (mean and S.D.) followed by a slow decay, resembling the monosynaptic field potentials generated by the activation of single muscle afferents (Munson and Sypert 1979a, b; Sypert et al. 1980). The half-height width of the s3 dIFPs varied between 1.8 and 4.8 ms (3.1 ± 0.62 mean ± SD) and was clearly longer than the duration of the s1 and s2 spikes that varied between 0.25 and 0.5 ms, respectively.
The rise time of the dIFPs recorded in 4 different experiments varied between 0.5 and 1.0 ms (0.79 ± 0.21 ms). This value is in fair agreement with the rise times of the homonymous monosynaptic EPSPs recorded in motoneurons after stimulation of single muscle spindle afferents reported by Munson and Sypert (1979b). Namely, 0.6 ± 0.03 ms; range 0.11–2.8 ms.
We used the coefficient of variation (CV) to compare the fluctuations in the amplitude and duration of the dIFPs with the fluctuations of the neuronal action potentials recorded in eight experiments. Interestingly, the amplitude CVs of the dIFPs (0.13 ± 0.05) and those of the neuronal action potentials (0.14 ± 0.04) were small and highly similar (two-sample Kolmogorov–Smirnov test, 5% significance level). In contrast, the half-height width CVs of the dIFPs were consistently larger (0.42 ± 0.14) than those of the action potentials (0.17 ± 0.08).
Based on these observations, we propose that the dIFPs are postsynaptic field potentials produced in small aggregates of dorsal horn neurons (see below).
The dIFPs can be preceded by afferent terminal potentials
Figure 2A shows superposed traces of the potentials comprised in the s3 cluster of Fig. 1C. It may be seen in the red traces of Fig. 2B and C that a significant fraction of the individual dIFPs (105 of 135 in this set of recordings) was preceded by a brief potential resembling the afferent terminal potentials (ATPs) recorded in the motor nucleus after electrical stimulation of single muscle spindle afferents by Munson and Sypert (1979a, b) and by Sypert et al. (1980) and in the cord dorsum by stimulation of single cutaneous afferents (Koerber et al. 1990).

dIFPs and afferent terminal potentials. A, s3 superposed dIFPs (same as those displayed in Fig. 1C). These potentials were visually separated into six different sets according to the amplitude of their afferent terminal potentials (ATPs). Ba and Bb show dIFPs generated without preceding ATPs (blue traces). Bc-Bf, dIFPs grouped according to the amplitude of their ATPs (red traces). Black arrow points at ATPs. Sample size is indicated. C, superposed traces of the dIFPs illustrated in Ba and Bf, with higher amplification and expanded time scale. Red vertical lines in C indicate the afferent terminal potential (black arrow) and the onset of the dIFPs. Negativity downward. See text for further explanations
Some of the ATPs were relatively small (Figs. 2Bc, 2Bd) and others larger (Fig. 2Be, f). We found also that 30 out of the 135 recorded dIFPs appeared not to be preceded by ATPs (Fig. 2Ba and 2Bb and Fig. 2Ca with a faster time scale). Yet, these dIFPs had basically the same amplitude and shape as those preceded by the ATPs, suggesting that both sets of dIFPs reflected postsynaptic potentials evoked in the same neuronal population (see below).
Altogether, we examined in 10 experiments 19 sets of dIFPs, each comprising PCA selected potentials with the same shape and amplitude. The fraction of dIFPs recorded in the dorsal horn that was preceded by an ATP varied over a wide range (between 3 and 100%). The latency between the ATPs and the onset of the dIFPs recorded in different experiments varied between 0.17 and 0.57 ms (0.37 ± 0.1 ms). These values resembled those reported by Munson and Sypert (1979a, b) for motoneurons activated by single muscle afferents (0.46 ± 0.01 ms) and suggest a monosynaptic linkage (see “Discussion”).
Intraspinal distribution and segmental coupling of the dIFPs
The observations depicted in Fig. 2 already suggest that a significant fraction of the recorded dIFPs are postsynaptic field potentials produced by afferent fibers. Yet, they provide limited information on their location within the dorsal horn and of their functional links with other neuronal populations in the same or in adjacent spinal segments.
In one experiment, we were able to record the dIFPs simultaneously from two closely placed micropipettes inserted in the dorsal horn. We could then examine the functional relations between the neuronal sets producing these two sets of potentials. The data depicted in Fig. 3 were obtained by placing one micropipette at 1226 µm (micro1, red traces) and the other slightly rostral, at 1432 µm depth, both in the region between Rexed’s laminae IV and V (micro2, green traces, see insert in Fig. 4B). We found that for each spontaneous PCA selected dIFP recorded with micropipette 1, a concurrent dIFP was recorded with the other electrode.

Correlated activation of dIFPs. A, paired sets of dIFPs were simultaneously recorded with two micropipettes, one at placed at 1226 µm (micropipette 1, red colors) and the other at 1432 µm depth (micropipette 2, green colors) Vertical gray bars indicate time of occurrence of the ATPs. Note that dIFPs recorded with micropipette 2 displayed in panels 1–4 were not preceded by the afferent terminal potential (marked with asterisks), in contrast with the dIFPs recorded with micropipette 1, all of which showed the afferent terminal potential. The red arrows point to a neuronal action potential recorded with both electrodes. B, superposed traces of the paired sets of dIFPs displayed in panels A1–A4. That is, of those pairs in which the dIFPs recorded with micropipette 2 had no distinct ATPs. C, superposed traces of the paired sets of dIFPs displayed in panels A5–A10, both of them with preceding ATPs (indicated by the double-headed arrows). Further explanations in the text

Intraspinal distribution of dIFPs. A–C, dIFPs were simultaneously recorded with two micropipettes inserted in the dorsal horn. Same experiment as that of Fig. 3. Micropipette 1 was left in a fixed position (1226 µm depth; red dot in histology) while micropipette 2 was positioned at various depths (between 1180 and 1800 µm; green dots in histology). The traces in A1 and A2 show the dIFPs simultaneously recorded with the fixed micropipette and with the mobile micropipette positioned at three different depths, as indicated. A3 shows that the amplitude normalized traces displayed in A1 and A2 had the same shape. B, plot of mean amplitude versus depth of the dIFPs recorded at different depths with micropipette 2 (green dots and graph) while micropipette 1 was left in a fixed position (1226 µm red dots). C1, correlation between the dIFPs recorded with micropipette 1 (fixed) and micropipette 2 positioned at different depths (see the vertical scale on the left side). C2 and C3, correlation between the dIFPs recorded with the mobile micropipette 2 and the L6 and L5 CDPs, respectively. D–F, same format as in A–C, but recordings made with micropipette 2 left in a fixed position (at 1432 µm) and micropipette 1 placed at different depths within Rexed’s laminae IV–V, as indicated in the inserts. Further explanations in text
Figure 3A displays 10 pairs of simultaneously recorded dIFPs. It may be seen that in some pairs, the ATPs were clearly visible in both dIFPs (traces 5–10 in Fig. 3A), while in other pairs the ATPs recorded with micro2 were not evident, even though the ATPs recorded with microelectrode 1 were clearly discernible (traces 1–4). Yet, we found no case where both dIFPs appeared without a preceding ATP, but this could be to the relatively small sample size or perhaps to the interference by the background noise.
Figure 3B and C shows the dIFPs displayed in Fig. 3A. They were visually separated in two different classes, according to whether the dIFPs recorded with micropipette 2 showed or not preceding ATPs at the time marked with double-headed arrows. It may be seen that four dIFPs showed rather small or no preceding ATPs (green traces in Fig. 3B), while in the other six recordings both sets of dIFPs showed clear terminal potentials (green traces in Fig. 3C). But even so, both sets of the dIFPs had a similar shape and amplitude.
The finding that the two sets of dIFPs had basically the same shape and amplitude, regardless of whether or not they were preceded by ATPs, suggests that both reflected postsynaptic potentials evoked in the same neuronal population, either by the ongoing afferent inputs and /or by the sporadic activation of some interneurons.
Based on the assumption that both sets of dIFPs were produced by the same set of dorsal horn neurons (see above) we could then examine the intraspinal distribution of the dIFPs recorded with one of the micropipettes using as reference the PCA selected dIFPs recorded with the fixed micropipette. The red traces in Fig. 4A1 provide three samples of the dIFPs recorded with the micropipette (micro1 fixed) that remained positioned at 1226 µm depth (Rexed’s lamina V), while Fig. 4A2 shows the dIFPs recorded at the same time with the other micropipette that was placed at 3 different depths, as indicated (green traces, micro2 mobile; see insert in Fig. 4B). Since the dIFPs recorded with micro1 were selected using PCA, they had rather similar amplitudes and shapes, in contrast with the unselected dIFPs recorded at the same time with micro2, whose amplitude varied with the recording depth.
To examine the extent of functional coupling between the neurons generating the two sets of dIFPs, we calculated the average Pearson correlation (using consecutive time-bins of ~ 1.5 ms) between the dIFPs recorded simultaneously with the two micropipettes, one fixed and the other placed at different depths.
As shown in Panel 1 in Fig. 4C, both sets of dIFPs were highly correlated at the time bin of their peaks (second column) and decayed during their falling phase (3rd–11th bins). We found in addition that the correlation between both sets of dIFPs was higher when micropipette 2 was within the superficial layers (1200–1650 µm) and smaller, but still appreciable when positioned at deeper layers (1650–1800 µm).
Figure 4B displays the mean amplitude of the dIFPs recorded at different depths with micro2 (green dots), together with the dIFPs recorded with micro1 left in a fixed position (red dots; see insert). It may be seen that the dIFPs recorded with micro2 in the most superficial layers were rather small, increased at deeper layers, became largest at 1500 µm and decreased thereafter, until they disappeared at 1700 mm depth, without showing a polarity reversal, while the amplitude of the dIFPs recorded with micro1 (red dots) remained rather constant throughout the whole observation (270 ± 13 µV, n = 268).
We assumed, as a working hypothesis, that the gradual decline in amplitude without a polarity reversal of the dIFPs recorded with micro2 at deeper locations was an indication that the dIFPs were produced by a set of neurons displaying a spatial distribution that generated closed field potentials during their synaptic activation. This means that within the region of the synaptically activated neuronal population the potential would be everywhere negative, and isopotentially zero everywhere outside the population (see Lorente de No 1947 and Klee and Rall, 1977 and “Discussion”). Based on this assumption, we expected that the dIFPs recorded at one site with micro1 and the dIFPs recorded at different depths with micro2, would all have the same shape, despite their different size. The amplitude normalized recordings depicted in Fig. 4A3 show that this was indeed the case.
It should be noted that the dIFPs recorded with the mobile electrode were largest between 1400 and 1650 µm depth, well within the projection region of cutaneous afferents (Manjarrez et al. 2000; Levinsson et al. 2002). They were still recorded 200 µm away from the site of maximum amplitude, in contrast with the narrow spatial spread of the extracellular recordings of neuronal action potentials, whose amplitude sharply decreased 30–50 microns away from the neuronal cell body (see Gold et al. 2006).
We also asked the question on the extent to which the current flows generated during the occurrence of individual dIFPs would be seen in the cord dorsum recordings. As in Fig. 4C1, we calculated the average correlation between the dIFPs recorded at different depths and the simultaneously recorded L5 and L6 CDPs. Figure 4C2 and C3 shows that the dIFPs recorded at different depths with the mobile micropipette were more correlated with the CDPs recorded in the same L6 segment (correlation coefficient 0.53 ± 0.08 at the time of the peak, Fig. 4C2) than with the CDPs recorded in the L5 adjacent segment (0.10 ± 0.11; Fig. 4C3). The increased correlation was seen only at the time of the peak and was not evident later on, during the falling phase of the dIFPs.
It thus seems that the dorsal horn neuronal ensembles generating the dIFPs had a spatially restricted distribution, in this case confined within a single segment. Yet, it should be noted in Fig. 4C2 that although the correlation with the L6CDPs was increased at the time of the peak of the dIFPs, the mean L6 CDPs showed no distinctive peaks at this time. This is not at all surprising because, as discussed above, activation of the neurons that produce the dIFPs generate closed field potentials with a rather circumscribed spatial distribution. These fields would be barely detectable at the cord dorsum, particularly after averaging a relatively small number of events, as in the present set of observations. In fact, as reported by Koerber et al. (1990) rather large samples (1024) were required to disclose the CDPs produced by stimulation of single cutaneous afferents.
Figure 4D1 and 4D2 show recordings obtained in the same experiment when micro2 was left in a fixed position while micro1 was placed at different depths within Rexed’s laminae IV–V (see insert in Fig. 4E). As in Fig. 4E, the dIFPs recorded deeper within the dorsal horn reduced their amplitude and basically disappeared, also without showing a polarity reversal (Fig. 4E). When amplitude normalized the potentials recorded with both micropipettes also had the same shape (Fig. 4D3). The two sets of potentials were also correlated at the time of their peak (Fig. 4F1). As shown in Fig. 4F2, the dIFPs recorded with micro1 at different depths also showed a mild correlation with the CDPs produced in the L6 segment (0.50 ± 0.13) and weak correlation with the CDPs recorded in the L5 segment (0.11 ± 0.09; Fig. 4F3).
These findings further support the proposal that the neuronal networks involved in the generation of the dIFPs were mostly local, in the sense that their stronger functional connections were with neurons in the same segment than with neurons in neighboring segments, suggesting actively rather than passively evoked synaptic actions by these interneurons.
Neuronal activation during the dIFPs
Quite often, the ongoing dIFPs also displayed distinct action potentials during their falling phase. The experiment of Fig. 5 was particularly interesting because we were able to record with a single microelectrode left in a fixed position (1208 µm depth within Rexed’s lamina IV) five separate classes of dIFPs and four different sets of clearly defined neuronal action potentials. We could then examine the patterns of neuronal activation during each class of dIFPs.

Neuronal activation during ongoing dIFPs. A, mean of L5 and L6 CDPs (upper pair of traces) and of the intraspinal potentials recorded with a single micropipette inserted at 1208 µm depth in the left L6 segment (lower trace). Superposed traces in the middle show dIFPs together with neuronal action potentials and below, two single traces, in this case not preceded by ATPs. B, same as A, but the dIFPs were preceded by a terminal potential (red arrows in inserts). C–E, other three sets of dIFPs still recorded with the same intraspinal electrode. dIFPs in A and B had the same shape and amplitude. The dIFPs displayed in C–E were preceded by terminal potentials. Sample size is indicated. Bottom, superposed traces of four different sets of action potentials, together with the histograms of their activation during the different classes of dIFPs, as indicated. Time zero set to the onset of the dIFPs (indicated by the vertical dotted lines). Further explanations in text
The upper pair of traces in Fig. 5A–E shows the averages of the L5 and L6 CDPs associated with each class of the selected dIFPs whose means are shown under the CDPs. The traces below show the superposed recordings of the dIFPs and neuronal action potentials included in each cluster, and below two single traces, both displayed with an expanded time base. It may be seen in the superposed traces that the neuronal action potentials appeared only during the falling phase of the dIFPs and not before and that they displayed different activation patterns during each class of dIFPs.
Quite interestingly, the dIFPs illustrated in panel A were generated without a preceding ATP and displayed different patterns of neuronal activation than the dIFPs shown in panels B–E that were preceded by ATPs (see red arrows in insets). Moreover, unlike the dIFPs shown in Fig. 5A, the dIFPs of Fig. 5B and those of Fig. 5E appeared synchronized with the L5 and L6 CDPs (see next section for more details).
We could identify four different sets of neuronal action potentials (shown as inserts in the left side of the bottom panels) and examine their occurrence during each of the five classes of dIFPs (see histograms). Neuron 1 (red) was activated during the dIFPs displayed in panels A, B, D and E and not activated during the dIFPs displayed in panel C. In contrast, neuron 2 (green) exhibited similar activation patterns during the dIFPs displayed in panels A, B and C and was silent during the dIFPs in panels D and E. Neuron 3 (blue) was activated in association with all the dIFPs, particularly during the dIFPs displayed in panels C and D, while neuron 4 (purple) appeared more often during the dIFPs displayed in panels D and E.
The latencies of the neuronal action potentials relative to the onset of the dIFPs were quite variable, but still within a given range for each class of dIFP. Some were relatively short (3.0–8.5 ms for neurons 1 and 2 activated during class D and C dIFPs, respectively). Others were longer (5.8–11 for neuron 3 during C, D and E dIFPs and 11–21 ms for neuron 4 during D and E dIFPs), suggesting variable but rather structured synaptic linkages of the neurons generating the dIFPs with other neuronal sets.
dIFPs are generated during the spontaneous negative and negative–positive CDPs
In previous work, we showed that the temporal relations between the ongoing activity of the dorsal horn neurons determines, to some extent, the patterns of functional connectivity between neurons in the network and their interaction with other neuronal populations (Contreras-Hernández et al. 2015). Namely, during low levels of synchronization, the dorsal horn neurons acquire a configuration that leads to the generation of negative cord dorsum potentials (nCDPs) and there is a preferential activation of the pathways mediating Ib non-reciprocal postsynaptic inhibition. During higher levels of synchronization, the activation of the dorsal horn neurons produces negative–positive potentials (npCDPs) together with increased transmission along the pathways mediating PAD and presynaptic inhibition.
The data described in Fig. 5 already indicated that some, but not all dIFPs increased their activity during the L6 and L5 CDPs. It thus seemed important to examine the extent to which there was some functional linkage between the neurons involved in generation of the nCDPs and npCDPs and the neuronal ensembles producing the dIFPs.
Figure 6A shows superposed traces of the L6-left nCDPs that were selected using the appropriate templates (see Methods). The traces below show a single nCDP and a single intraspinal field with superposed dIFPs recorded at 1456 µm depth in Rexed’s lamina V. The rasters and histograms of the dIFPs illustrated in Fig. 6B and C show that most of the ongoing dIFPs were activated during the peak of the negative component of the nCDPs. The blue superposed traces displayed at the bottom of panel A show the dIFPs recorded at that time. These dIFPs were visually separated in two groups, one occurring without a preceding ATP and another with clear ATPs (see arrow). Likewise, as shown in Fig. 6D–F, the same set of dIFPs was activated during the negative phase of the npCDPs. The red traces below panel D show the dIFPs, also separated in two groups, one with and the other without a preceding ATP. It should be noted that the dIFPs associated either with the nCDPs or with the npCDPs had similar amplitude and time course, regardless of whether they were or not preceded by ATPs.

Generation of dIFPs during nCDPs and npCDPs. A, superposed traces of ongoing nCDPs recorded in the rostral part of the left L6 segment together with a single nCDP and an intraspinal field potential with superposed dIFPs recorded at 1456 µm depth. B, C, rasters and corresponding histograms of the dIFPs generated during the L6-nCDPs. D–F, same format as in A–C, but for the dIFPs generated during the L6-npCDPs. Inserts show the superposed traces of the dIFPs generated during the nCDPs and npCDPs with and without preceding ATPs (arrows). See text for additional details
We found in 10 experiments that the proportion of dIFPs with and without preceding ATPs generated during the nCDPs and npCDPs was not significantly different (p-value = 0.1826, t-test).
Changes in dIFP frequency during ongoing nCDPs and npCDPs
We have shown in the previous section that the dIFPs increased their spontaneous firing during both nCDPs and npCDPs. In Contreras-Hernández et al. (2015) we suggested that the npCDPs are generated during a state of increased synchronization along a longitudinally distributed ensemble of dorsal horn neurons. It, thus, seemed of interest to examine the changes in the frequency of the dIFPs generated at different times before, during and after the nCDPs and npCDPs.
Figure 7A shows that the ongoing dIFPs appeared more often during the npCDPs than during the nCDPs recorded in the same segment (L5 or L6; n = 15, Two-sample Kolmogorov–Smirnov test, p < 0.05). In contrast, as shown in Fig. 7B, the increments in the spontaneous occurrence of the dIFPs during the nCDPs and npCDPs recorded in the adjacent segment were smaller and similar.

Changes in dIFP frequency during ongoing nCDPs and npCDPs A, mean frequency of dIFPs associated with both nCDPs and npCDPs (n + , np +) recorded in the same segment (L5 or L6). B, same as A, but changes measured relative to the spontaneous CDPs recorded in the adjacent segment. Upper traces are sham nCDPs and npCDPS used as reference. C, number of dIFPs associated with nCDPs and npCDPs (n + , np +), with nCDPs only (n + , np0), with npCDPs only (n0, np +) or to none of them (n0, np0). D–F, spinal location of the dIFPs associated with nCDPs and/or npCDPs, as indicated. Note that most of the recorded dIFPs were located within Rexed´s laminae IV–VI. Further explanations in the text
These observations indicate that during the spontaneous npCDPs, the dIFPs are generated more often than during the nCDPs. Therefore, it is reasonable to assume that paired sets of spontaneous dIFPs would be more correlated during the npCDPs when there is preferential activation of the pathways mediating PAD than during the nCDPs, as it has been shown for individual dorsal horn neurons (Contreras-Hernández et al. 2015). It is, therefore, possible that the dIFPs were also involved in the generation of PAD, particularly during the states of the increased neuronal synchronization induced by nociception (see “Discussion” and Contreras-Hernández et al. 2018 and Martin et al. 2019).
Altogether, we examined the changes in the probabilities of occurrence of 53 sets of dIFPs during the spontaneous the nCDPs and npCDPs recorded in the L5-L6 segments. Figure 7C shows that 15 dIFPs increased their spontaneous firing during both the nCDPs and the npCDPs (n + , np +), 7 dIFPs were activated only during the nCDPs (n + , np0), 2 only during the npCDPs (n0, np +) and 29 dIFPs did not change their spontaneous frequency during the nCDPs or the npCDPs (n0, np0). It, thus, seems that about half of the examined dIFPs occurred in synchrony with the nCDPs and/or npCDPs. As shown in Fig. 7D–F, there were no differences in the location of the recording sites of the different sets of dIFPs. All of them were recorded in the dorsal horn, most of them in Rexed’s laminae IV–VI.
dIFP responses produced by electrical stimulation of cutaneous and muscle nerves
Based on the findings of Munson and Sypert (1979a, b) and of Koerber et al. (1990), we assumed that the ATPs that preceded the dIFPs were produced by incoming afferent action potentials. Hence, we tested in several experiments the possible activation of the dIFPs by electrical stimulation of cutaneous and muscle nerves. Seen in retrospect this approach was not the best choice because stimulating the whole nerve produced complex focal potentials where identification of single dIFPs was rather difficult. Ideally, we should have examined the responses produced by stimulating single afferents as it was done by Munson and Sypert (1979a, b), but this required another experimental design that we hope to pursue in the future. Nevertheless, we have been able to show in several experiments that the ongoing dIFPs were inhibited by stimulation of low-threshold Aβ cutaneous afferents while they were activated by stimulation of Aδ cutaneous and group II muscle afferents.
Figure 8A provides details on the intraspinal responses produced by SP nerve stimulation, in this case with single pulses 1.6xT applied while the intraspinal recording electrode was inserted in the dorsal horn, at 1668 µm from the cord surface in Rexed’s lamina V (see insert in Fig. 8D). It may be seen that SP stimulation produced a slow positive–negative focal potential. Stronger stimuli (1.8xT) produced in addition to the evoked focal potentials, several small potentials resembling the dIFPs (arrow in Fig. 8B).

Activation of dIFPs by stimulation of SP afferents in the Aδ range. A and B, CDPs and IFPs produced by stimulation of the SP nerve with two different strengths, as indicated. At this depth (1668 µm, Rexed’s lamina V), the focal potentials produced with 1.6xT stimuli were positive–negative. Stronger stimuli (1.8xT) show in addition short-latency dIFPs (see arrow). C and D, the same but recordings made deeper (2200 µm, Rexed’s lamina VII). Note sign reversal of the focal potentials. E and F, superposed traces of single CDPs and of IFPs recorded at both depths following SP stimulation with 1.6 and 1.8xT pulses. Note faster time base. Brackets in F indicate dIFP onset latencies. SP stimuli were delivered at 1 Hz. Insert shows location of recording sites. Further explanations in the text
Figure 8C and D shows the SP-induced responses recorded when the intraspinal electrode was inserted 532 µm deeper along the same track (red dot in histology). At this location (2200 µm; Rexed´s lamina VII), stimulation of the SP nerve with pulses 1.6xT and 1.8xT produced focal potentials with an already reversed polarity. Yet, the dIFPs produced by the 1.8xT stimuli still remained negative (arrow in Fig. 8D).
Figure 8E and F shows, with an expanded time base, single traces of the CDPs together with the focal potentials recorded in the superficial (black traces) and deep locations (red traces). As shown by the black horizontal brackets displayed in Fig. 8F, the onset latency of the earliest neuronal responses produced by the SP 1.8xT recorded at the superficial location was 6.25 ms (measured from the time of arrival of the earliest volley recorded in the cord dorsum). Likewise, the latency of the neuronal responses recorded at the deeper location was 6.8 ms respectively (red horizontal bracket). The onset latencies of the dIFPs produced by SP stimulation recorded in 2 experiments varied between 4.55–7.35 ms and were within the range expected for activation by Aδ fibers (Coombs et al. 1956).
Already in our 2000 paper, we reported that stimulation of the SU and SP nerves with relatively weak strengths (1.2xT) induced a pronounced inhibition of the ongoing CDPs lasting between 100 and 150 ms (see Fig. 10 in Manjarrez et al. 2000). Subsequent studies also indicated that the spinal potentials produced by skin stimulation with pulses of 3.2xT strength were also inhibited when preceded by spontaneous nCDPs or npCDPs (see Fig. 9 of Contreras-Hernández et al. 2015). This finding raised the question on whether the ongoing dIFPs would be similarly inhibited during SP stimulation with relatively low strengths. The observations depicted in Fig. 9 address to this question.

Ongoing dIFPs are inhibited by stimulation of Aβ and activated by SP Aδ afferents. A–C, CDPs and IFPs produced by stimulation of the SP nerve with 3 different strengths, as indicated. Panel 1, superposed CDPs and IFPs. Panel 2, samples of individual intraspinal recordings. Note that in addition to the focal potentials produced by SP stimulation these records show dIFPs (marked with small black vertical bars). Panel 3, raster displays of the dIFPs (dots) together with the bars shown in panel 2. A, 1.3xT SP stimuli barely affected the occurrence of the dIFPs. B, 1.6xT SP stimuli prevented dIFP generation for about 20 ms. C, stimuli 1.8xT instead activated the dIFPs with latencies between 4.5 and 7.3 ms after the arrival of the afferent volley. Intraspinal field potentials were recorded at 2.2 mm depth. Data from same experiment as Fig. 8. See text for additional information
Figure 9A provides several samples of intraspinal recordings made at 2200 µm depth during the SP stimulation with single pulses 1.3xT strength applied once per second. It may be seen that in addition to the rather small focal potentials produced by the SP stimulus, the occurrence of the ongoing dIFPs (marked with small black vertical bars) was barely changed after the stimulus (see superposed traces in Panel A1 and raster display in panel A3). In contrast, 1.6xT SP stimulation prevented the occurrence of the dIFPs during 20 ms after the arrival of the afferent volley (Fig. 9B), while stronger stimuli (1.8xT) led instead to the occurrence of dIFPs with an onset latency of 6.2–6.8 ms relative to the arrival of the afferent volley that was followed by a prolonged silent period (Fig. 9C).
Figure 10 illustrates the responses produced by PBSt stimulation with single pulses 2 and 5xT strength recorded in the same experiment as that of Fig. 8. It may be seen in Fig. 10A and B that the cord dorsum potentials (CDPs) produced by PBSt 2xT and 5xT stimulation had two components, one brief potential with a rather short latency due to activation of gr I fibers (black arrow) and a delayed slow potential due to activation of gr II fibers (black double-headed arrow; see Lindblom and Ottosson, 1955).

IFPs and dIFPs produced by activation of gr. II PBST afferents. Same experiment and format as in Fig. 8. A, B, CDPs and IFPs produced with stimuli 2xT and 5xT PBSt stimuli recorded at 1668 µm depth. Red arrow in B points at the shortest latency dIFPs. C, D, same but recording electrode placed deeper (2200 µm depth). Note sign reversal of the PBSt evoked focal potentials but not of the dIFPs that remain negative. E, F, superposed traces of CDPs and IFPs recorded at both depths with faster time resolution. Brackets in F show estimated latencies of the first and second dIFPs in the series. Single arrow points at the gr I volley recorded in the cord dorsum following PBSt stimulus. Double headed arrow points at the CDP produced by the gr. II fibers. Further explanations in text
Both the 2 and 5xT stimuli produced a slow field potential resembling the CDP ascribed to the activation of gr II afferents as well as a series of small superposed wavelets that we have attributed to field potentials rather than to neuronal spikes. At this depth (1668 microns, black traces), the slow gr II field potential was basically negative (Fig. 10A, B and black traces in Fig. 10E and F) and became slightly positive at deeper locations, particularly with the 5xT stimuli (2200 microns Fig. 10C, D and red traces in Fig. 10 E and F). Yet, and quite remarkably, the wavelets ascribed to the dIFPs retained their negative polarity at both depths. The latency of the first set of wavelets measured from the time of arrival of the gr I volley was 6.1 ms (range 6.1–6.7 ms; black bracket in Fig. 10F) suggests it was produced by activation of muscle afferents with conduction velocities in the range of group II fibers (Edgley and Jankowska, 1987; Bras et al. 1989). The onset latency of the second set was around 10.3 ms (red bracket, range 9.7–10.6 ms) and suggests oligosynaptic activation.
In two additional experiments, we were able to examine the activation of individual dIFPs following relatively strong electrical stimulation (1.8xT–2xT) of cutaneous and muscle nerves without the interference of the field potentials produced by nerve stimulation. In these cases, the onset latencies of the dIFPs produced by SP stimulation were 6.2 and 9.1 ms (n = 2) and of 5.1 ms by SU nerve stimulation (n = 1). Stimulation of the PBST and GS nerve with 5xT stimuli produced dIFPs with onset latencies of 6.1 ms and of 8.3 ms (n = 1), respectively.
Discussion
The dIFPs are generated by synchronous activation of discrete sets of dorsal horn neurons
As indicated in the Introduction, in addition to the action potentials produced by individual dorsal horn neurons, we also recorded small ongoing negative field potentials with a fast-rising phase followed by a slow decay (dIFPs) resembling the intraspinal postsynaptic focal potentials produced in the motor nuclei by stimulation of single Ia afferent fibers (Munson and Sypert 1979a,b) as well as the cord dorsum potentials recorded in several spinal segments after electrical stimulation of single cutaneous afferents (Koerber et al. 1990).
In contrast with the intraspinal fields produced by the neuronal action potentials that decrease rapidly with distance (Buzaki et al. 2012; Linden et al. 2014; Gold et al. 2006), the dIFPs have displayed a wider intraspinal distribution (300–400 µm; see Figs. 4B and E) that fits well with the distribution displayed by the cutaneous afferent fibers arriving at the spinal cord, that extend their axonal processes within III-IV Rexed's laminae and synapse with a population of neurons whose dendritic and axonal processes remain confined within these laminae (see Brown 1982; Koerber et al. 1990; Granmo et al. 2008; Bannatyne et al. 2009).
An important finding was that the amplitude of the dIFPs was reduced at deeper intraspinal locations until it faded without showing a polarity reversal (see green graph in Fig. 4B and red graph in Fig. 4E, as well as Figs. 8F and 10F). This is in contrast with the polarity reversal of the field potentials generated during the nCDPs and npCDPs and of the focal potentials evoked by electrical stimulation of cutaneous and muscle nerves as shown in Figs. 8 E, F and 10E, F (see also Fig. 5 in the paper of Chávez et al. 2012).
Since we were unable to find a polarity reversal of the dIFPs recorded in the deepest regions including the superficial region of the ventral horn (up to 2500 µm), we propose that the examined dIFPs were produced by small populations of functionally coupled neurons whose dendrites and cell bodies have a spatial orientation producing closed fields when synaptically activated. This implies that within the region of the synaptically activated neuronal population the potential would be everywhere negative, and isopotentially zero everywhere outside the population (see Lorente de No 1947; Klee and Rall 1977). In other words, the dIFPs would be generated by the synaptic activation of small sets dorsal horn neurons displaying a circumscribed spatial distribution. Nevertheless, we must be aware that this just a working hypothesis that requires precise anatomical information on the spatial orientation of the neurons leading to these potentials, as pointed out by Klee and Rall (1977).
The ATPs that precede the dIFPs are afferent terminal potentials
One additional and important finding disclosed by the present study was that a significant fraction of the dIFPs was preceded by small spike-like terminal potentials (ATPs) resembling those recorded by Koerber et al. (1990) and by Munson and Sypert (1979a, b) following stimulation of single cutaneous and muscle afferents.
At present, our observations provide limited indications on the possible origin of these ATPs, because electrical stimulation of cutaneous and of muscle nerves with strengths up to 5xT produced in most cases focal potentials with several components that precluded proper identification of individual dIFPs. Clearly, a better approach would have been to stimulate single afferents, as was done by Sypert et al. (1980) while recording the dIFPs, or else by recording the action potentials of single afferents from nerves left in continuity with the peripheral receptors as was done by Solodkin et al. (1991), but the present series of experiments was not designed for that purpose. Nevertheless, the results obtained so far indicate that a significant fraction of the dIFPs was activated both by gr II muscle and by Aδ cutaneous afferents (see Figs. 8, 9, 10).
It could be argued that some ATPs, if not all, could be also produced by the ongoing activation of local glutamatergic interneurons interposed in the gr II pathways because, as shown by Jankowska and Roberts (1972) the activation of premotor interneurons produces in motoneurons field potentials with a presynaptic volley resembling the potential generated by Ia afferents (see also Bannatyne et al. 2009), but this remains to be investigated. Yet, for the time being, we propose that the presently recorded ATPs were intraspinal fields generated by the action potentials invading the terminal arborizations of the afferent fibers (see below).
Variability of the ATPs suggests a fluctuating level of presynaptic depolarization
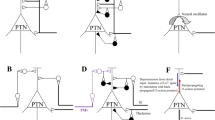
Another important finding was that the ATPs that preceded a particular class of the dIFPs recorded with a single micropipette left in a fixed position were not constant but showed clear fluctuations, both in shape and amplitude. Based on our previous studies that demonstrated the local character of the GABAergic PAD (Quevedo et al. 1997; Lomeli et al. 1998), it is possible that the different amplitudes and shapes, as well as the absence of the ATPs, reflected changes in impulse conduction, including possible failures in some of the intraspinal collaterals of the afferent fibers synapsing with the network, either spontaneously (Luscher et al. 1983; Debanne et al. 2011; Hari et al. 2021) or during the PAD elicited by stimulation of sensory nerves (Rudomin and Dutton 1969; Wall and McMahon 1994; however, see Lomelí et al. 2000). This possibility is considered in the diagram of Fig. 11A (see Figure legend for details).

Origin of the ATP amplitude variations. The drawings show the intraspinal branches of the afferent fibers that activate the tightly coupled set of neurons that generates the dIFPs (the neurons are not shown), as well as the field potentials recorded with two micropipettes placed close to the neuronal cluster. Panel A shows that under condition 1, all the intraspinal collaterals of the afferent fiber conduct action potentials. Hence, the two micropipettes would record similar dIFPs, both of them preceded by ATPs. Under condition 2, a fraction of the intraspinal terminals of the afferent fibers would be subjected to the effects produced by the intermittent activation of a discrete population of GABAergic neurons. This would produce PAD in some of the collaterals of the afferent fiber and reduce the amplitude or even suppress the afferent volley that precedes the dIFPs. However, since the neurons in this cluster are strongly coupled, activation of few neurons would be sufficient to activate the whole ensemble. Panel B illustrates a case where the neurons generating the dIFPs would be activated by more than one afferent fiber, each displaying a particular pattern of intraspinal branching. The presence or absence of ATPs preceding the dIFP would then depend on the location of the recording microelectrode relative to the intraspinal collaterals of the afferent fibers. Under condition 1 all the collaterals in that particular afferent would be activated and both electrodes would record dIFPs with preceding ATPs. Under condition 2, the collaterals of the activated afferent would have a more restricted spatial distribution. One electrode would then record dIFPs preceded by ATPs and the other dIFPs preceded by smaller or no ATPs. The dIFPs in this figure were taken from Fig. 3B and C. Further explanations in text
To the extent to which the variations in the shape and amplitude of the ATPs result from spatial and temporal variations in PAD, the resulting presynaptic inhibition exerted on the cutaneous fibers that activate the dIFPs could well be the expression of a mechanism that allows transmission of information from the periphery either “as a simple unit or in a fractional manner”, to activate specific sets of spinal neurons, as suggested by the data depicted in Fig. 5 (see Rudomin and Schmidt 1999; Rudomin 2009; Côté et al. 2018).
However, as shown in Fig. 11B, an alternative explanation to the variations in the amplitude and shapes of the potentials preceding the same set of dIFPs could be that the size of the extracellularly recorded presynaptic volleys depends on the distance between the tip of the electrode and the actual site of the synaptic contacts. If only a small number of afferent fibers connect with single or a few neurons, the effects of distance may vary considerably, especially if the synapses were on dendrites, we would record dIFPs without preceding ATPs.
It is of course possible that the dIFPs generated without an afferent terminal potentials were produced by the sporadic activation of some neurons within the ensemble, that would induce a synchronized activation of the whole set of highly coupled neurons, as it has been shown in cultured neurons by Soriano et al. (2008), but this remains as an open question.
The dIFPs are postsynaptic field potentials generated by tightly coupled sets of neurons
Regardless of whether the dIFPs are activated by afferent and/or interneuronal ongoing activity, the present set of observations suggests that the dIFPs are synchronized postsynaptic potentials displayed by spatially confined sets of tightly coupled neurons. Yet, at present it is not clear which mechanisms underlie such a strong neuronal coupling. One possible mechanism could involve robust excitatory connections between neurons in the ensemble as in the olfactory bulb (Karnup et al. 2006). It is also possible that the strong neuronal coupling involves electrical synapses (Chapman et al. 2013; Nagy et al. 2004; Bautista et al. 2012; Nakamura et al. 2015).
Clearly, these two possibilities are not mutually exclusive, and the high synchronization could be mediated by mixed synapses (Rash et al. 1996). In this context, the observations of Bautista et al, (2012) are particularly relevant. They show that transgenic mice lacking connexin36, a protein that mediates electrical communication via gap junctions between neurons, suffer a severe impairment of presynaptic inhibition. Quite interestingly, similar effects can be produced in normal mice with drugs that disrupt gap junction function. In their view, gap junction coupling may be extensive within subsets of interneurons organized into functionally discrete groups. Moreover, Bautista et al. (2012) provide evidence showing that immunofluorescence labeling of Cx36 is associated with neurobiotin-coupled neurons in the deep dorsal horn and intermediate zone of mice at P11 with a confined spatial distribution.
In fact, as shown here, a significant fraction of dIFPs is activated during the npCDPs, which means they are functionally related with the generation of PAD. It is, therefore, possible that these are the same sets of neurons as those examined by Bautista et al. (2012), but so far this remains as an open question.
Another possibility would be that the neurons generating the dIFPs are the di3 spinal interneurons involved in the cutaneous control of hand grasp (see Bui et al. 2013). These interneurons appear to have a modular organization, convey inputs from low- and medium-threshold (1–4xT) cutaneous afferents and are mostly located in the deep dorsal horn and in intermediate laminae. Alternatively, they could be inhibitory neurons, as suggested by the observations of Bourane et al. (2015). These interneurons are defined by the expression of neuropeptide Y::Cre (NPY::Cre) and are assumed to gate mechanical itch. They are activated by light mechanical stimulation of the skin and are located in laminae III and IV. Yet, these two classes of dorsal horn neurons may not be good candidates because we found that the dIFPs we have so far recorded are activated by high-threshold cutaneous and gr II muscle afferents and inhibited by stimulation low-threshold cutaneous afferents (Figs. 8, 9, 10).
In short, the identity of the neurons producing the dIFPs is still unclear as it is the possibility that the dorsal horn neurons generate different classes of dIFPs, each with specific actions on the different spinal pathways. Alternatively, it is also possible that depending on the patterns of neuronal synchronization, the same set of dorsal neurons generates the different classes of dIFPs, as it seems to be the case with the nCDPs and npCDPs (see Contreras-Hernández et al. 2015), and this remains to be investigated.
Distributed and discrete organization of dorsal horn neuronal ensembles
In a previous study, Contreras-Hernández et al. (2015) reported that a significant proportion of the ongoing dorsal horn neuronal activity displayed higher synchronization during the npCDPs and was more susceptible to activation by low-threshold cutaneous afferents than during the nCDPs. We then proposed that this set of dorsal horn neurons was part of a segmentally distributed ensemble of dorsal horn neurons whose interaction with other spinal pathways varied according to its patterns of functional synchronization (see also Chávez et al. 2012; Manjarrez et al. 2003; Lomelí et al. 1998).
Although the dIFPs share many characteristics of the segmentally distributed ensemble of dorsal horn neurons leading to the nCDPs and npCDPs (see Contreras-Hernández et al. 2015), they have some distinctive features: (a) less than half of the whole sample of dIFPs was associated with nCDPs and/or npCDPs compared with individual dorsal horn neurons where most (75%) were associated with spontaneous nCDPs and/or npCDPs, (b) the probability of response of the dorsal horn neurons to stimulation of low-threshold cutaneous afferents was higher for the neurons associated with the CDPs (65%). In contrast, the presently examined dIFPs reduced their activity following stimulation of low-threshold cutaneous afferents, while they responded to stimulation of high-threshold muscle and cutaneous fibers, suggesting that the neurons producing the dIFPs form part of other, but still functionally related networks. c) The activity of the dorsal horn interneurons involved in the generation of the nCDPs and npCDPs is correlated with the activity of other neuronal populations located in the same and in other spinal lumbar segments. In contrast, the dIFPs appear to be associated with the activation of neuronal populations confined within the same segment. Nevertheless, the observation that a significant fraction of the dIFPs is activated during the nCDPs and npCDPs suggests that these particular sets of neurons could be involved in the generation of Ib non-reciprocal postsynaptic inhibition and of PAD and presynaptic inhibition (see below).
Altogether, the present set of observations suggests that the dIFPs are not produced by the activation of scattered groups of neurons but rather by the activation of small ensembles of tightly coupled neurons. Functionally coherent groups of neurons or modules have been also described in a variety of structures in the nervous system such as cortical columns (Mountcastle 1997), hippocampus (Bazelot et al. 2010, 2016) or the olfactory bulb, where the glomeruli act as discrete functional units encoding different odors (Johnson and Leon 2000; Wachowiack and Shipley 2006).
In the case of the spinal cord, even though there is no general consensus for the meaning of "module", many papers refer to it as a functional and/or anatomical group of neurons mediating specific features of motor actions (Hägglund et al. 2013; Wyart et al. 2009; Hart and Giszter 2010; Bizzi and Cheung 2013; Bizzi et al. 2008) or units of sensory-motor transformations that simplify the central processing of multiple sensory inputs producing specific outputs (Tresch et al. 1999; Schouenborg 2008; Chopek et al. 2018).
By labeling the terminations of coarse and thin afferent fibers, Levinsson et al. (2002; see also Granmo et al. 2008) proposed that first cutaneous relay neurons and reflex encoders conform to a robust functional anatomic column in the dorsal horn. In this regard, the electrical synapses (see above) could have a key role in the appropriate timing during the integration of the sensory information (see Wachowiak and Shipley 2006). They could also participate in the processing of the premotor (and/or motor) activity where the chemical and/or electrical synapses facilitate the highly coordinated activation between motoneurons (Tresch and Kiehn 2000; Bhumbra and Beato 2018).
One interesting feature of the dIFPs pertains the activation of the same set of neurons during the different classes of dIFPs illustrated in Fig. 5. We have assumed, also as a working hypothesis, that the action potentials that are generated during the falling phase of the dIFPs could well represent the patterned activation of premotor neurons (Levinsson et al. 2002) related to the spinal networks involved in the generation of the structured organization of the withdrawal reflexes analyzed by Schouenborg (2002, 2008). Alternatively, they could form part of the distributed dorsal horn neuronal ensemble that modulates the input of sensory information by changing, in a dynamic and flexible manner, the functional connectivity between their individual elements (Jankowska 2001; Contreras-Hernández et al. 2015), and thus affect transmission along a variety of inhibitory pathways (Contreras-Hernández et al. 2015).
This raises the question on the functional role of a modular architecture (dIFPs) and a distributed neuronal configuration operating simultaneously in the dorsal horn (see Contreras-Hernández et al. 2015). We believe that these arrangements are not mutually exclusive but represent different modes of operation in the spinal cord.
A distributed ensemble would allow the dorsal horn neurons to respond in a very flexible manner to the incoming sensory inputs (Seki et al. 2003; Contreras-Hernández et al. 2015). Such a distributed system would permit the nervous system to adapt itself and operate in response to a variety of sensory stimuli without the need for additional specialized circuits (Ekeberg 1993; Ekeberg et al. 1995). One example of this situation has emerged from the analysis of the changes in the activity of the dorsal horn neurons involved in the generation of the nCDPs and npCDPs. These neurons are distributed along several spinal segments (L4–L7). During states of low neuronal synchronization there is a preferential generation of nCDPs and Ib-non-reciprocal postsynaptic inhibition. Increased neuronal synchronization in the same ensemble would instead lead to the generation of npCDPs and of presynaptic inhibition (Contreras-Hernández et al. 2015).
In contrast, the activation of strongly functionally coupled sets of interneurons by specific neuronal inputs would be barely affected, even under high levels of synaptic noise produced by other neuronal networks. This feature could be involved in the processing of very specific sensory inputs in a shorter time-window, as it could be the case with the modular organization of the flexor reflexes described by Schouenborg (2002, 2008). We propose, as a working hypothesis, that the dIFPs fall within this category. Yet, it is still possible that the different classes of dIFPs are distributed along several spinal segments, just as the nCDPs and npCDPs. This seems unlikely because, as discussed above, the neurons that generate the dIFPs have functional connections with other neurons in the same but not in adjacent segments.
An intriguing question is how modular and distributed systems interact in the spinal cord. The segmental reorganization of dorsal horn neuronal ensembles in response to nociceptive inputs (Contreras-Hernández et al. 2018) is also associated with a non-random reversible reorganization of the segmental patterns of presynaptic inhibition (Chávez et al. 2012), suggesting that such distributed ensemble and specific local populations of GABAergic interneurons cooperate on the process of modulation and transmission of sensory information during nociceptive stimulation.
Concluding remarks
Studies made in anesthetized animals have provided valuable information on the non-random dynamic reorganization of the functional connectivity between the dorsal horn neurons during the activation of nociceptive pathways (Martin et al. 2015, 2017; Contreras-Hernández et al. 2018; see also Rodríguez et al. 2011) and how the nociceptive-induced changes are modulated by supraspinal pathways (Contreras-Hernández et al. 2018). These studies were mostly based on the analysis of the functional relations between the activity of individual dorsal horn neurons and the cord dorsum potentials generated by neuronal synchronous firing. A more detailed analysis of the dorsal horn field potentials has now disclosed the ongoing generation of a different kind of field potentials, the dIFPs that we have ascribed to the activation of strongly coupled sets of neurons that could be intrinsically active and also respond to stimulation of gr II muscle and Aδ cutaneous afferents.
Our observations suggest in addition that the neuronal ensembles generating the dIFPs display robust patterns of functional connectivity between the neurons involved in their generation. This view agrees with previous work from our laboratory (Martin et al. 2015, 2017, 2019) where it was shown that the spontaneous cord dorsum potentials (CDPs) produced by segmentally distributed dorsal horn neuronal ensembles can be grouped in discrete classes with specific shapes and durations. Each of these classes of spontaneous CDPs appeared with a specific non-random pattern of activation before and after the activation of nociceptive pathways.
Future work contemplates a more exhaustive analysis of the functional relevance of the neuronal groups that give rise to the dIFPs. In this context, a key issue will be the proper identification of the afferent fibers involved in the generation of the ATPs that precede the dIFPs, in terms of their origin and sensory modality as well as the connections of the dIFPs neurons with neurons in other spinal pathways. This information is expected to provide a more integral view on the functional relations between the interneurons involved in the generation of the dIFPs and the local and segmental populations of dorsal horn neurons involved in the presynaptic modulation of sensory information and in the organization of the flexor reflexes activated by these sensory inputs.
Abbreviations
- ATP:
-
Afferent terminal potential
- CDPs:
-
Cord dorsum potentials
- CV:
-
Coefficient of variation
- GS:
-
Gastrocnemius
- dIFPs:
-
Discrete intraspinal field potentials
- L:
-
Lumbar
- nCDPs:
-
Negative cord dorsum potentials
- npCDPs:
-
Negative–positive cord dorsum potentials
- PAD:
-
Primary afferent depolarization
- PCA:
-
Principal component analysis
- PBSt:
-
Posterior biceps and semitendinosus
- s:
-
Spontaneous
- SP:
-
Superficial peroneal
- SU:
-
Sural
References
Bannatyne BA, Liu TT, Hammar I, Stecina K, Jankowska E, Maxwell EJ (2009) Excitatory and inhibitory intermediate zone interneurons in pathways from feline group I and II afferents: differences in axonal projections and input. J Physiol 587:379–399
Bautista W, Nagy JI, Dai Y, McCrea D (2012) Requirement of neuronal connexin36 in pathways mediating presynaptic inhibition of primary afferents in functionally mature mouse spinal cord. J Physiol 590:3821–3839
Bazelot M, Dinocourt C, Cohen I, Miles R (2010) Unitary inhibitory field potentials in the CA3 region of rat hippocampus. J Physiol 588:2077–2090
Bazelot M, Teleńczuk MT, Miles R (2016) Single CA3 pyramidal cells trigger sharp waves in vitro by exciting interneurones. J Physiol 594:2565–2577
Bhumbra GS, Beato M (2018) Recurrent excitation between motoneurones propagates across segments and is purely glutamatergic. PLoS Biol 16(3):e2003586
Bizzi E, Cheung VC (2013) The neural origin of muscle synergies. Front Comput Neurosci 7:1–6
Bizzi E, Cheung VC, d’Avella A, Saltiel P, Tresch M (2008) Combining modules for movement. Brain Res Rev 57:125–133
Bourane S, Duan B, Koch SC, Dalet A, Britz O, Garcia-Campmany L, Kim E, Cheng L, Ghosh A, Ma Q, Goulding M (2015) Gate control of mechanical itch by a subpopulation of spinal cord interneurons. Science 350:550–554
Bras H, Cavallari P, Jankowska E, McCrea D (1989) Comparison of effects of monoamines on transmission in spinal pathways from group I and II muscle afferents in the cat. Exp Brain Res 76:27–37
Brown AG (1982) The dorsal horn of the spinal cord. Q J Exp Physiol 67:193–212
Bui TV, Akay T, Loubani O, Hnasko TS, Jessell TM, Brownstone RM (2013) Circuits for grasping: spinal dl3 interneurons mediate cutaneous control of motor behavior. Neuron 78:191–204
Buzsáki G, Anastassiou CA, Koch C (2012) The origin of extracellular fields and currents EEG, ECoG LFP and spikes. Nat Rev Neurosci 13:407–420
Chapman RJ, Lall VK, Maxeiner S, Willecke K, Deuchars J, King AE (2013) Localization of neurones expressing the gap junction protein Connexin45 within the adult spinal dorsal horn: a study using Cx45-eGFP reporter mice. Brain Struct Funct 218:751–765
Chávez D, Rodríguez E, Jiménez I, Rudomin P (2012) Changes in correlation between spontaneous activity of dorsal horn neurones lead to differential recruitment of inhibitory pathways in the cat spinal cord. J Physiol 590:1563–1584
Chopek JW, Nascimento F, Beato M, Brownstone RM, Zhang Y (2018) Sub-populations of spinal V3 interneurons from focal modules of layered pre-motor microcircuits. Cell Rep 25:146–156
Contreras-Hernández E, Chávez D, Hernández E, Rudomin P (2011) Spontaneous focal micropotentials produced by the synchronous activation of small sets of dorsal horn neurons and their relation with primary afferent depolarization in the cat spinal cord. Abs Soc Neurosci. 917.17/SS17
Contreras-Hernández E, Chávez D, Rudomin P (2015) Dynamic synchronization of ongoing neuronal activity across spinal segments regulates sensory information flow. J Physiol 593:2343–2363
Contreras-Hernández E, Chávez D, Hernández E, Velázquez E, Reyes P, Béjar J, Martín M, Cortés U, Glusman S, Rudomin P (2018) Supraspinal modulation of neuronal synchronization by nociceptive stimulation induces an enduring reorganization of dorsal horn neuronal connectivity. J Physiol 596:1747–1776
Coombs JS, Curtis DF, Landgren S (1956) Spinal cord potentials generated by impulses in muscle and cutaneous afferent fibres. J Neurophysiol 19:452–467
Côté MP, Murray LM, Knikou M (2018) Spinal control of locomotion: individual neurons, their circuits and functions. Front Physiol 25(9):784
Debanne D, Campanac E, Bialowas A, Carlier E, Alcaraz G (2011) Axon physiology. Physiol Rev 91:555–602
Drummond GB (2009) Reporting ethical matters in The Journal of Physiology: standards and advice. J Physiol 587:713–719
Eaton JW, Bateman D, Hauberg S, Wehbring R (2018) GNU Octave version 4.4.1, March 2019. Manual: a high-level interactive language for numerical computations. https://www.gnu.org/software/octave/doc/v4.4.1/
Edgley SA, Jankowska E (1987) An interneuronal relay for group I and II muscle afferents in the midlumbar segments of the cat spinal cord. J Physiol 389:647–674
Ekeberg Ö (1993) A combined neuronal and mechanical model of fish swimming. Biol Cyber 69:363–374
Ekeberg Ö, Grillner S, Lansner A (1995) The neural control of fish swimming studied through numerical simulations. Adapt Behav 3:363–384
Gold C, Henze DA, Koch C, Buzsáki G (2006) On the origin of the extracellular action potential waveform: a modeling study. J Neurophysiol 95:3113–3128
Granmo M, Petersson P, Schouenborg J (2008) Action-based body maps in the spinal cord emerge from a transitory floating organization. J Neurosci 28:5494–5503
Grundy D (2015) Principles and standards for reporting animal experiments in The Journal of Physiology and Experimental Physiology. J Physiol 593:2547–2549
Hägglund M, Dougherty KJ, Borgius L, Itohara S, Iwasato T, Kiehn O (2013) Optogenetic dissection reveals multiple rhythmogenic modules underlying locomotion. Proc Natl Acad Sci 110:11589–11594
Hari K, Lucas-Osma A, Bennett DJ et al (2021) Nodal GABA facilitates axon spike transmission in the spinal cord. Neuron. https://doi.org/10.1101/2021.01.20.427494inpress
Hart CB, Giszter SF (2010) A neural basis for motor primitives in the spinal cord. J Neurosci 30:1322–1336
Jankowska E (2001) Spinal interneuronal systems: identification, multifunctional character and reconfigurations in mammals. J Physiol 533:31–40
Jankowska E, Roberts WJ (1972) Synaptic actions of single interneurones mediating reciprocal Ia inhibition of motoneurones. J Physiol 222:623–642
Johnson BA, Leon M (2000) Modular representations of odorants in the glomerular layer of the rat olfactory bulb and the effects of stimulus concentration. J Comp Neurol 422:496–509
Karnup SV, Hayar A, Shipley MT, Kurnikova MG (2006) Spontaneous field potentials in the glomeruli of the olfactory bulb: the leading role of juxtaglomerular cells. Neuroscience 142:203–221
Klee M, Rall W (1977) Computed potentials of cortically arranged populations of neurons. J Neurophysiol 40:647–666
Koerber HR, Brown PB, Mendell LM (1990) Correlation of monosynaptic field potentials evoked by single action potentials in single primary afferent axons and their bouton distributions in the dorsal horn. J Comp Neurol 294:133–144
Levinsson A, Holmberg H, Broman J, Zhang M, Schouenborg J (2002) Spinal sensorimotor transformation: relation between cutaneous somatotopy and a reflex network. J Neurosci 22:8170–8182
Lewicki MS (1998) A review of methods for spike sorting: the detection and classification of neural action potentials. Network 9:R53–R78
Lindblom UF, Ottosson JO (1955) Bulbar influence on spinal cord dorsum potentials and ventral root reflexes. Acta Phys Scand 35:203–214
Lindén H, Hagen E, Lęski S, Norheim ES, Pettersen KH, Einevoll GT (2014) LFPγ: a tool for biophysical simulation of extracellular potentials generated by detailed model neurons. Front Neuroinform 16:7–41
Lomelí J, Quevedo J, Linares P, Rudomin P (1998) Local control of information flow in segmental and ascending collaterals of single afferents. Nature 395:600–604
Lomelí J, Castillo L, Linares P, Rudomin P (2000) Effects of PAD on conduction of action potentials within segmental and ascending branches of single muscle afferents in the cat spinal cord. Exp Brain Res 135:204–214
Lorente de No R (1947) Analysis of the distribution of the action current of nerve in volume conductors. Stud Rockefeller Inst Med Res Repr 132:384–477
Luscher HR, Ruenzel P, Henneman E (1983) Composite EPSPs in motoneurons of different sizes before and during PTP: implications for transmission failure and its relief in Ia projections. J Neurophysiol 49:269–289
Manjarrez E, Rojas-Piloni JG, Jiménez I, Rudomin P (2000) Modulation of synaptic transmission from segmental afferents by spontaneous activity of dorsal horn spinal neurones in the cat. J Physiol 529:445–460
Manjarrez E, Jiménez I, Rudomin P (2003) Intersegmental synchronization of spontaneous activity of dorsal horn neurons in the cat spinal cord. Exp Brain Res 148:401–413
Martin M, Contreras-Hernández E, Bejar J, Esposito G, Chávez D, Glusman S, Cortes U, Rudomin P (2015) A machine learning methodology for the selection and classification of spontaneous spinal cord dorsum potentials allows disclosure of structured (non-random) changes in neuronal connectivity induced by nociceptive stimulation. Front Neuroinform 26:9–21
Martin M, Béjar J, Esposito G, Chávez D, Contreras-Hernández E, Glusman S, Cortés U, Rudomín P (2017) Markovian analysis of the sequential behavior of the spontaneous spinal cord dorsum potentials induced by acute nociceptive stimulation in the anesthetized cat. Front Comput Neurosci 1:11–32
Martin M, Béjar J, Chávez D, Ramírez-Morales A, Hernández E, Moreno L, Contreras-Hernández E, Glusman S, Cortés U, Rudomin P (2019) Supraspinal shaping of adaptive transitions in the state of functional connectivity between segmentally distributed dorsal horn neuronal populations in response to nociception and antinociception. Front Syst Neurosci. https://doi.org/10.3389/fnsys.2019.00047
Mountcastle VB (1997) The columnar organization of the neocortex. Brain 120:701–722
Munson JB, Sypert GW (1979a) Properties of single central Ia afferent fibres projecting to motoneurones. J Physiol 296:315–327
Munson JB, Sypert GW (1979b) Properties of single fibre excitatory post-synaptic potentials in triceps surae motoneurones. J Physiol 296:329–342
Nagy JI, Dudek FE, Rash JE (2004) Update on connexins and gap junctions in neurons and glia in the mammalian nervous system. Brain Res Rev 47:191–215
Nakamura Y, Morioka N, Zhang FF, Hisaoka-Nakashima K, Nakata Y (2015) Downregulation of connexin36 in mouse spinal dorsal horn neurons leads to mechanical allodynia. J Neurosci Res 93:584–591
National Research Council (2010) Guide for the care and use of laboratory animals, 8th edn. The National Academic Press, Washington
Quevedo J, Eguibar JR, Lomelí J, Rudomin P (1997) Patterns of connectivity of spinal interneurons with single muscle afferents. Exp Brain Res 115:387–402
Rash JE, Dillman RK, Bilhartz BL, Duffy HS, Whalen LR, Yasumura T (1996) Mixed synapses discovered and mapped throughout mammalian spinal cord. Proc Natl Acad Sci USA 93:4235–4239
Rodríguez EE, Hernández-Lemus E, Itzá-Ortiz BA, Jiménez I, Rudomin P (2011) Multichannel detrended fluctuation analysis reveals synchronized patterns of spontaneous spinal activity in anesthetized cats. PLoS ONE 6(10):e26449. https://doi.org/10.1371/journal.pone.0026449
Rudomin P (2009) In search of lost presynaptic inhibition. Exp Brain Res 196:139–151
Rudomin P, Dutton H (1969) Effects of muscle and cutaneous afferent nerve volleys on excitability fluctuations of Ia terminals. J Neurophysiol 32:158–169
Rudomin P, Schmidt RF (1999) Presynaptic inhibition in the vertebrate spinal cord revisited. Exp Brain Res 129:1–37
Rudomin P, Hernández E, Lomelí J (2007) Tonic and phasic differential GAB aergic inhibition of synaptic actions of joint afferents in the cat. Exp Brain Res 76:98–118
Schouenborg J (2002) Modular organisation and spinal somatosensory imprinting. Brain Res Rev 40:80–91
Schouenborg J (2008) Action-based sensory encoding in spinal sensorimotor circuits. Brain Res Rev 57:111–117
Seki K, Perlmutter SI, Fetz EE (2003) Sensory input to primate spinal cord is presypnaptically inhibited during voluntary movement. Nat Neurosci 6:1309–1316
Solodkin M, Jiménez I, Collins WF III, Mendell LM, Rudomin P (1991) Interaction of baseline synaptic noise and Ia EPSPs: evidence for appreciable negative correlation under physiological conditions. J Neurophysiol 65:927–945
Soriano J, Rodríguez Martínez M, Tlusty T, Moses E (2008) Development of input connections in neuronal cultures. Proc Natl Acad Sci USA 105:13758–13763
Sypert GW, Munson JB, Fleshman JW (1980) Effect of presynaptic inhibition on axonal potentials, terminal potentials, focal synaptic potentials, and EPSP in cat spinal cord. J Neurophysiol 44:792–803
Tresch MC, Kiehn O (2000) Motor coordination without action potentials in the mammalian spinal cord. Nat Neurosci 3:593–599
Tresch MC, Saltiel P, Bizzi E (1999) The construction of movement by the spinal cord. Nat Neurosci 2:162–167
Wachowiak M, Shipley MT (2006) Coding and synaptic processing of sensory information in the glomerular layer of the olfactory bulb. Semin Cell Dev Biol 17:411–423
Wall PD, McMahon SB (1994) Long range afferents in rat spinal cord. III. Failure of impulse transmission in axons and relief of the failure following rhizotomy of dorsal roots. Phil Trans Roy Soc Lond B343:211–233
Wyart C, Del Bene F, Warp E, Scott EK, Trauner D, Baier H, Isacoff EY (2009) Optogenetic dissection of a behavioural module in the vertebrate spinal cord. Nature 461:407–410
Acknowledgements
We want to thank L Mendell and R Brownstone for their comments to earlier versions of the text, E Rosales for excellent secretarial support and Cirilo León for technical support. This work was partly supported by CONACyT grant 50900. E.C.-H. was holder of a CONACyT fellowship for doctoral studies. Present address: Department of Biomedical Engineering, University of Texas at Austin. 107 W Dean Keeton St, Austin, TX 78712.
Author information
Authors and Affiliations
Contributions
Conception and design of experiments, E.C-H. and P.R.; data collection, E.C.-H., E.H. and D.Ch; data analysis, E.C-H.; interpretation of data and drafting of the article and reviewing it critically for important intellectual content: E.C-H. and P.R. All the authors have contributed to this study. Experiments were performed at the Department of Physiology, Biophysics and Neurosciences, Center of Research and Advanced Studies of the Instituto Politécnico Nacional, Mexico. All authors approved the final version of the manuscript for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Communicated by Winston D Byblow.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Contreras-Hernández, E., Chávez, D., Hernández, E. et al. Discrete field potentials produced by coherent activation of spinal dorsal horn neurons. Exp Brain Res 240, 665–686 (2022). https://doi.org/10.1007/s00221-021-06286-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-021-06286-3