
Abstract
The two-visual stream hypothesis posits that the dorsal stream is less susceptible than the ventral stream to the effects of illusions and visual priming. While previous studies have separately examined these perceptual manipulations, the present study combined the effects of a visual illusion and priming to examine the possibility of dorsally guided actions being susceptible to the perceptual stimuli due to interactions between the two streams. Thirty-four participants were primed with a ‘long’ or ‘short’ version of the Sander Parallelogram illusion and were asked to either reach out and grasp or manually estimate the length of a rod placed on a version of the illusion that was on some trials the same as the prime (congruent) and on other trials was the inverse (incongruent). Due to the context-focused nature of ventral processing, we predicted that estimations would be more susceptible to the effects of the illusion and priming than grasps. Results showed that while participants’ manual estimations were susceptible to both priming and the illusion, the grasps were only affected by the illusion, not by priming. The influence of the illusion on grip aperture was greater during manual estimations than it was during grasping. These findings support the notion that the functionally distinct dorsal and ventral streams interact under the current experimental paradigm. Outcomes of the study help better understand the nature of stimuli that promote interactions between the dorsal and ventral streams.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Every day we encounter diverse and complex environments. Vision allows us to identify the components and features in our surrounding environments and interact with these features in a meaningful way. The seamless processing and integration of visual information that enables us to navigate and engage with our surroundings have been explained by Goodale and Milner’s (1992) two visual streams hypothesis. This model explains that the cortical visual system is divided into a dorsal stream and a ventral stream. The ventral stream (“what” stream) transmits visual information to the inferior temporal cortex (Goodale and Milner 1992; Ungerleider and Mishkin 1982). The ventral stream controls perception and conscious visual judgements (Kravitz et al. 2013), allowing us to store visual information in memory (Goodale and Milner 2006). The dorsal stream (“how” stream) transmits to the posterior parietal cortex (Goodale and Milner 1992; Ungerleider and Mishkin 1982), mediates the planning and execution of visually guided actions and operates in real time (Buneo and Andersen 2006; Goodale et al. 2004). The division of labor between the two streams detailed by Goodale and Milner (1992) has been widely supported by several studies that have used a variety of methods, including neuropsychological experiments (James et al. 2003), single cell recordings (Fogassi et al. 2001) and neuroimaging studies (Cavina-Pratesi et al. 2010; Culham et al. 2003).
However, emerging research has challenged the amount of functional distinction between the two streams. Most notably it has been found that the dorsal stream is not only involved in the perception of object shape and structure (Freud et al. 2015, 2018) but is also involved in primarily perceptual tasks like face and object recognition in humans (Jeong and Xu 2016; Zachariou et al. 2017). Furthermore, there is evidence of dorsal stream activity that occurs prior to the planning and execution of actions in both human (Faillenot et al. 1999; Kourtzy and Kanwisher 2000) and non-human primates (Srivastava et al. 2009). One possible explanation for the lack of clear functional distinction between the two streams could lie in the interactions between the dorsal and ventral streams. Anatomical studies have shown the presence of white matter projections between the dorsal and ventral visual areas (Yeatman et al. 2014), projections between the inferior temporal and parietal lobes (Zhong and Rockland 2003) and the dorsal anterior intraparietal area having connections to the superior and middle temporal gyri of the ventral stream (Borra et al. 2007).
While evidence for communication between the two streams has been found using anatomical methods, information regarding the functional interactions between the two streams can be gleaned using visual illusions. Illusory images manipulate spatial context to influence perceptions of the size of a central target (Dickinson et al. 2019). Support for the two streams theory comes from Agliotti et al. (1995), who found that ventrally guided perceptual tasks (e.g., manual estimations) were affected by illusions, while dorsally guided actions (e.g., grasping) were not. Differences in illusory susceptibility suggest that the visually guided actions operate under two different systems of control: an ‘offline’ or memory-guided system that captures all aspects of a visual scene, which relies on ventral stream processing (Kravitz et al. 2013), and an ‘online’ system that is constantly updated in real time and does not rely on memory, but instead on active dorsal stream processing (Cesanek et al. 2018; Goodale 2008; Foley et al. 2015).
Existing research on ventral and dorsal actions has shown contradictory results. Some studies have found that while perceptual size estimations are influenced by illusory backgrounds, grasping actions toward the stimulus are resistant to illusions and are consistent with the actual size of the stimulus (Agliotti et al. 1995; Cesanek et al. 2018; Haffenden and Goodale 1998; Katsumata 2019; Westwood and Goodale 2003; Whitwell et al. 2018). These results indicate that the dorsal stream uses real-time information to guide action and is not influenced by ventral processing. Other studies have shown evidence supporting interactions between the two visual streams, where both perception and action were found to be susceptible to illusions (Bruno and Franz 2009; Heath et al. 2005; Kopiske et al. 2016). These results have been attributed to functional interactions between the two streams, where information from the ventral stream influences dorsally guided actions toward the target stimuli.
One possible explanation for these differing results has been identified as the availability of visual information during the grasping actions. Heath et al. (2005) observed that closed-loop actions, where vision is available throughout the grasp, are resistant to illusions since visual feedback can be used to adjust online and future movements to reduce errors. Open-loop grasps, where visual feedback is occluded after movement onset are better suited for illusion studies since they strengthen the effect of the illusion by promoting interaction between the perception and action streams (Heath et al. 2005; Milner and Goodale 1992; Whitwell et al. 2018). Occlusion of vision during the grasp leads to both ventral and dorsal representations being accessed to compensate for limited visual information. Additionally, open-loop grasps ensure that participants avoid using visual feedback to adapt to the illusion and consciously reduce illusion-induced errors (Whitwell et al. 2018).
The division of labor between the dorsal and ventral streams has also been tested through visuomotor priming studies. Priming involves the presentation of a visual stimulus (a prime) before the presentation of a target stimulus to influence actions toward that target (Roche et al. 2015). Per the two visual streams hypothesis, the dorsal stream operates in real time to process visual information that would control actions toward an object (Milner and Goodale 2008). If there was functional dissociation between the two streams, visually guided grasps controlled by dorsal processing would be resistant to effects of priming as they would not account for memory of the prime stimulus while memory-guided grasps that access ventral stream representations would be affected by priming (Seegelke et al. 2016). However, the majority of existing research exploring effects of priming on motor actions toward 3D objects has shown effects of priming during both visually guided and memory-guided grasps (Hesse et al. 2008; Masson et al. 2011; Roche et al. 2015; Seegelke et al. 2016). These results indicate that while the two streams may operate on different time scales, dorsal and ventral representations interact to guide actions. Existing action priming studies have shown that prime stimuli that are congruent with a target, facilitate grasps toward the target (Hesse et al. 2008; Roche and Chainay 2013; Pisu et al. 2020; Seegelke et al. 2016). Conversely, when the prime and target are incongruent, actions toward the target are slower or less accurate (Seegelke et al. 2016).
It is important to note that in visuomotor priming studies, kinematic variables such as maximum grip aperture are more useful in understanding interactions compared to variables such as initiation time. Movement initiation time has been identified as a variable that is more biased toward ventral processing, as it accounts for action planning, motor control and decision-making processes all of which need to be carried out prior to the execution of the action (Hesse et al. 2008; Roche et al. 2015; Wong et al. 2015). Focusing on dorsal processing during a grasp is beneficial as it allows us to evaluate the extent of the effect the ventrally stored prime has on the action and thus better understand the conditions that promote interactions between the two streams. Variables such as grip aperture (Seegelke et al. 2016; Valyear et al. 2011), grip orientation (Hesse et al. 2008; Roche et al. 2015) and time to maximum grip aperture are more representative of dorsal stream processing as they do not account for the action planning that occurs before an action is initiated. These variables account for the action planning that occurs after an action is initiated and reflect adjustments made during the course of the action (van Sonderen and Denier van der Gon 1991). The purpose of the current experiment was to explore if the two visual streams interact to influence visually guided actions when priming and visual illusions are combined, and how interactions between the two streams influence visually guided actions.
The Sander Parallelogram illusion was used in the current experiment due to its robust illusory effect and lack of distractor elements (Whitwell et al. 2018). Participants were primed with either the ‘long’ or ‘short’ version of the illusion. Following a brief window of time, participants were presented with a three-dimensional target rod placed on a version of the illusion that was either in the same orientation as the prime (congruent) of the inverse orientation (incongruent). All participants performed two experimental tasks: a manual estimation task and a grasping task. In the manual estimation task, participants used their thumb and index finger to estimate the length of the metal rod placed on the central diagonal line of the two-dimensional Sander Parallelogram illusion. The grasping task required participants to reach out, grasp and lift the metal rod placed on the illusion using a precision grip. Grasping was performed with partial visual feedback, where vision of the target was removed upon action initiation, while estimations were performed with full visual feedback of the target available for the duration of the task. The estimation task was used to analyze the effects of illusions and priming on the ventral stream, while the grasping task was used to determine the effects of the illusion and priming on the dorsal stream (Whitwell et al. 2018).
Consistent with the two visual systems hypothesis, we hypothesized that overall, manual estimations would be more susceptible to the effects of the illusion and priming than grasps (Carther-Krone et al. 2020; Whitwell et al. 2018). Manual estimations of the perceived size of the rod were expected to be larger for the perceptually ‘long’ illusion compared to the perceptually ‘short’ illusion. Previous studies have consistently found that manual size estimations are susceptible to illusory backgrounds (Foster et al. 2012; Ozana and Ganel 2020; Whitwell et al. 2018), this has been attributed to reliance on the ventral stream processing which considers visual information of the target as well as the surrounding illusory background, thereby increasing susceptibility to illusory effects during estimations. Since the dorsal stream provides real-time visual feedback, no illusion-based differences in maximum grip aperture (MGA) were expected during grasping trials (Haffenden and Goodale 1998; Goodale 2008; Katsumata 2019; Whitwell et al. 2018). With respect to priming, we predicted that incongruent priming to lead to larger grip apertures in both estimations and grasps due to the compensatory adjustments required from participants when interacting with a target that is different from the previously seen prime (Roche et al. 2015; Seegelke et al. 2016). This is based on the notion that if the target does not match the prime stimulus, participants will compensate for the difference in length by widening their grip to ensure that accuracy of the grasp (Volcic and Domini 2016). We expected the same to occur when the target was perceptually shorter, as normal grasp mechanics typically start with a wider aperture that scales down to fit the size of the target (Keefe et al. 2019). Finally, as prior studies have found that the memory of a prime influences the perceived size of a target (Seegelke et al. 2016), we predicted that the average grip apertures for both grasps and estimations when the prime was the ‘short’ illusion, and the target was the ‘long’ illusion, would be smaller compared to the average grip apertures for both estimations and grasps where the prime was the ‘long’ illusion, and the target was the ‘short’ illusion.
Methods
Participants
Thirty-eight undergraduate psychology students were recruited for this study and received course credit for their participation. Four participants (1 male) of the original 38 participants were excluded from data analyses due to them not being able to perform successful grasps throughout the study. Thus, the final sample consisted of 34 participants (9 males; 18–25 years old; (M = 19.6 years). All participants had normal or corrected to normal vision, and were right-hand dominant, as determined by a modified version of the Edinburgh Handedness Inventory (Oldfield 1971). All experimental procedures were approved by the Psychology/Sociology Research Ethics Board (PSREB) at the University of Manitoba.
Stimuli and materials
PLATO liquid crystal shutter goggles (Plato Translucent Technologies, Toronto, ON) were programmed to vary the amount of vision provided to the participant over the course of the reach. Grip aperture was recorded using an Optotrak Certus motion capture camera (Northern Digital Inc., Waterloo, ON, Canada) at a frequency of 175 Hz. Six infrared light-emitting diodes (IREDS) were attached to the participants' right hand and wrist (2 IREDs each placed on the proximal edge of cuticle of the index finger, the proximal edge of the cuticle of the thumb, and on the distal radius of the wrist). Only one IRED at each position was used to analyze the participants’ movement. If there was a significant loss of data using the first IRED at one of these locations (e.g., missing or extreme values due to rotation of the hand), the second IRED was used for the analysis of that participant. MotionMonitor software (Innovative Sports Training Inc., Chicago, IL, USA) was used to record spatial and temporal data from the Optotrak, control the time intervals at which the PLATO shutter goggles became transparent or opaque, as well as to generate the auditory tone (350-Hz), which served as a cue for the participants to perform the grasping or estimation tasks. This software was run on an Inspiron 545 Dell computer (Duo Core 3.16 GHz).
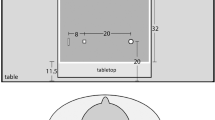
The illusion underlying the Sander Parallelogram is that the diagonal line bisecting the larger parallelogram is perceived as longer than the line bisecting the smaller parallelogram, even though both lines are the same length. Therefore, while the length of the target was constant in all trials, the illusory background made the target appear longer or shorter than its actual length. The Sander Parallelogram illusion was used to examine illusory and priming effects on a ventrally based perceptual estimation task and dorsally driven grasping task. The stimuli used in this study included a black, three-dimensional metal rod (6 cm × 0.3 cm × 0.3 cm) and the Sander Parallelogram illusion (Fig. 1), which was presented on a white background on a Lenovo X201 ThinkPad tablet screen (26 cm × 16 cm). Participants were instructed to either grasp and lift or estimate the length of the metal target rod placed on the tablet screen over the diagonal line in the upper half of the Sander Parallelogram illusion. The illusory background shown by the tablet screen was alternated between the ‘long’ and ‘short’ illusory configurations, depending on the experimental condition. Even though the illusory background alternated between ‘long’ and ‘short’ configurations on the screen, the metal rod that was placed on top of the tablet screen was in the same location and orientation in all trials. This ensured that participants were reaching and viewing the target at the same angle and distance for each trial.

Two configurations of the Sander Parallelogram illusion were used in the study. The upper, diagonal line in the “short” illusion appears perceptually shorter than the upper diagonal line in the ‘long’ illusion, even though both lines are the same length. The metal rod was always placed in the same location and orientation (i.e. on the upper diagonal line), and only the illusory background was alternated between the ‘long’ and ‘short’ configurations for each trial. The four conditions included: congruent stimuli on long illusion (a), congruent stimuli on short illusion (b), prime on long illusion and target on short illusion (c), prime on short illusion and target on long illusion (d)
Procedure
After completing the consent form and handedness questionnaire, participants had the IREDs attached to their index finger, thumb, and wrist using adhesive tape. Participants performed the experiment wearing the PLATO goggles while standing in front of a tabletop, where the stimuli were displayed along the participant’s sagittal plane. The stimuli included the tablet screen that showed the Sander parallelogram illusion (either ‘long’ or ‘short’ depending on the condition) and the 6-cm metal rod. The metal rod was placed over the diagonal line on the upper half of the illusion during viewing of the prime and viewing of the target. On each trial, participants were required to start with their index finger and thumb pinched together, touching a designated ‘starting spot’ on the table located 20 cm away from the participant and 10 cm away from the stimulus display. Figure 2 shows the procedure used in the study. The goggles were opaque at the start of each trial and remained opaque for 4.25 s, after which they turned translucent for 0.5 s allowing participants to view the prime stimulus in either the ‘long’ or ‘short’ configuration of the Sander Parallelogram illusion. The goggles then turned opaque for 1.5 s during which the display background either remained the same for the congruent prime condition, or was alternated in the incongruent condition, so the perceived length was different than the prime. This time duration was less than the two second duration it takes for information to decay in short-term memory (Hesse and Franz 2009; Roche et al. 2015) ensuring that the visual information from the prime remained in memory for the subsequent task. The goggles then became translucent which was accompanied by an auditory tone of 350 Hz. Upon hearing the tone participants performed either the grasping or estimation tasks.

Procedural timing used in the experiment. Following the presentation of the prime participants head a tone that indicated the start of one of three tasks: estimation, grasping with partial visual feedback or grasping with full visual feedback
The procedure for the grasping and estimation tasks followed the procedure outlined above, with the exception that participants had full vision in the estimation block but only had partial vision in the grasping block. In the estimation trials, participants used their index and thumb to indicate how long they perceived the target rod to be and let the experimenter know through a verbal signal (saying “ok”) once they made a satisfactory estimation and their grip aperture remained stable. Once the verbal signal was provided, the experimenter concluded the trial and the participant returned their hand to the starting spot. In the grasping trials, participants were required to reach out grasp and lift the rod off the tablet screen to a height of around 2.5 cm and place it back down. The trial was ended by the experimenter once the rod was lifted off the surface.
For the grasping trials, visual information was only available until grasp initiation, which was defined as when the wrist IREDs surpassed the 20 cm mark, indicating the original ‘starting spot’, and reaching a velocity of 5 cm/s (Roche et al. 2015). At this point, the goggles turned opaque. The difference in availability of visual information for the two tasks was necessary due to prior research showing that illusory effects are minimized when there is visual feedback of the hand contacting the target (Bruno and Franz 2009; Heath et al. 2006, 2005). Since illusory effects are minimized when visual feedback is available due to online feedback mechanisms that operate to reduce any errors caused by the illusion, removing visual feedback at the time of contact reduces the likelihood of participants using error minimization strategies and promotes a stronger reliance on ballistic visual processing until the hand moves (Whitwell et al. 2018).
To ensure that the illusion was influencing participants, grasping trials with visual feedback for the full duration of the movement were included. Grasping trials with full visual feedback followed the same sequence as the partial visual feedback grasps. The goggles were opaque for 4.25 s, they then turned translucent for 0.5 s to view the prime. The goggles turned opaque again for a duration of 1.5 s. After the 1.5 s elapsed, the goggles turned translucent and a 250 Hz tone was sounded to cue the participant to initiate grasping task. In the full visual feedback grasping trials, the goggles remained translucent from the moment the auditory tone was played until the time participant grasped the target and raised it to a height of 2.5 cm. Unlike in the partial visual feedback tasks, the goggles did not turn opaque once movement was initiated but were translucent for the full duration of the grasping motion. All estimation trials were performed with full visual feedback available after the onset of the tone. Since there was no physical contact with the target in the estimation trials, there was no need to remove visual feedback for this task. Furthermore, since estimations occur on a slower timescale than grasps (Franz 2003), it was beneficial to provide visual feedback for the entire duration of the estimation task.
Design
In total, four different conditions were present in the experiment (Fig. 1): congruent prime and target stimuli on perceptually long illusion, congruent prime and target stimuli on perceptually short illusion, incongruent prime on short illusion with target on long illusion and incongruent prime on long illusion with target on short illusion. The estimation block consisted of twenty trials, where each of the four stimulus conditions was performed five times in a randomized order. The grasping block had 42 trials, seven trials for each of the four stimulus conditions with partial visual feedback (28 trials with partial visual feedback available) and seven trials each for congruent long and short illusions with full visual feedback available (14 trials with full visual feedback available). In total, there were 62 experimental trials. To control for task priming effects half of the participants were randomly assigned to perform the grasping task before the estimation task, while the remaining participants performed the estimation task first (Whitwell et al. 2018).
Data analyses
Estimation trials were analyzed using a 2 (Congruency: prime and target congruent, prime and target incongruent) × 2 (Illusion: target on perceptually long illusion, target on perceptually short illusion), repeated measures ANOVA to examine whether there were any differences in final grip aperture (FGA). This measure was defined as the vector distance between the thumb and index finger once the participant had made a satisfactory estimation and was used to determine if participants perceive the rod to be its actual length (6 cm) or its illusory length. For the five trials in each of the four conditions, grip aperture data were averaged, and condition means were used in the ANOVA analysis.
Grasping trials were analyzed using 2 (Congruency: prime and target congruent, prime and target incongruent) × 2 (Illusion: target on perceptually long illusion, target on perceptually short illusion) repeated measures ANOVAs to examine whether there were any differences in maximum grip aperture (MGA) and time to maximum grip aperture. MGA was defined as the maximum vector distance between the index finger and thumb between the start and end of the reach and was analyzed to help determine if participants plan for a wider grasp when the long illusion is shown compared to when the short illusion is shown. The start of the reach was defined as the time point at which the wrist sensor moved faster than a velocity of 5 cm/s and had passed the ‘starting spot’ located 20 cm away from the edge of the table. The end of the reach was defined as the time point where the wrist velocity was slower than 5 cm/s after the reach had been initiated. Time to MGA was defined as the length of time in seconds between the start of the movement and MGA. This variable was analyzed to examine how priming and illusions affect the execution of the action plan during the grasp and to further understand how grip aperture was influenced in each of the conditions. For the first 42 trials in the grasping block, MGA and time to MGA were averaged across each of the seven trials in each of the 4 conditions and the condition means were used in two ANOVA analyses.
An additional analysis was also performed to directly compare the overall effect of the illusion across conditions in the grasping and estimation tasks. This refers to the difference in grip aperture between perceptually long and short stimuli (Whitwell et al. 2018). For this analysis condition means representing perceptually ‘long’ stimuli and perceptually “short” stimuli were computed for each participant in each of the following 5 conditions: grasping with congruent priming, grasping with incongruent priming, estimation with congruent priming, estimation with incongruent priming and grasping under full vision with congruent priming. The overall effect of the illusion was then calculated by subtracting the mean grip aperture for perceptually long stimuli from the mean grip aperture for the perceptually short stimuli such that positive values indicate a response consistent with the direction of the illusion. These values were then submitted to a 2 (Task: grasping vs. estimation) × 2 (Prime: congruent vs. incongruent) repeated-measures within-subject ANOVA to determine the role of priming in affecting the illusion across task condition. The overall effect of the illusion for each of the 5 conditions was also examined using one-sample t tests against zero to determine whether the illusion influenced grasping and manual estimations for each condition individually using the Holm (1979) multiple comparisons procedure.
Finally, MGA and time to MGA were compared across conditions in which full vision and partial vision were available. The last fourteen trials of the grasping block in which full vision was available consisted of trials showing congruent primes and targets. These were compared the fourteen congruent partial visual feedback trial using 2 (Visual feedback: full visual feedback, partial visual feedback) × 2 (Illusion: target on perceptually long illusion, target on perceptually short illusion), repeated measures ANOVAs. Consistent with the above analyses, the 7 trials in each of the 4 conditions were averaged and the condition means were submitted to the ANOVA analyses. All analyses were carried out using alpha = 0.05 and all post hoc comparisons were performed using a Bonferroni correction (p < 0.05).
Results
Estimation
A 2 (Prime: congruent vs. incongruent) × 2 (Illusion: target on perceptually long illusion, target on perceptually short illusion) repeated measures ANOVA was performed to explore the presence of any effects of priming and illusion on the final grip aperture (FGA) in the estimation task. As can be seen in Fig. 3 there was a statistically significant main effect of the illusion on the manual estimation task, F (1,33) = 12.118, p = 0.001, ηp2 = 0.269, but there was no effect of priming, F (1,33) = 1.016, p = 0.321, ηp2 = 0.030. A post hoc analysis for the main effect of the illusion showed that estimations were larger for targets on the long illusory background (M = 6.8 cm, SE = 0.10 cm) compared to the short illusory background (M = 6.5 cm, SE = 0.10 cm). There was a statistically significant interaction between illusions and primes, F (1,33) = 19.792, p < 0.001, ηp2 = 0.375. Post hoc tests for the estimation task showed that when the prime and target were congruent, grip aperture was larger for the long illusory background (M = 6.9 cm, SE = 0.10 cm) compared to short illusory background (M = 6.4 cm, SE = 0.20 cm), p < 0.001. There was no significant difference between the long (M = 6.7, SE = 0.20) and short (M = 6.6 cm, SE = 0.20 cm) illusory backgrounds when the prime and target were incongruent, p = 0.321. When estimations of the target length were made for the long illusory background the grip aperture was larger when the prime and target were congruent (M = 6.9, SE = 0.10 cm) compared to when the prime and target were incongruent (M = 6.7 cm, SE = 0.20 cm), p < 0.001. Size estimations of the target on the short illusory background were larger for the incongruent prime (M = 6.6 cm, SE = 0.20 cm) compared to the congruent prime (M = 6.4 cm, SE = 0.20 cm), p = 0.001.

Final grip aperture results (cm) for the estimation task. When the prime and target were congruent, estimations were significantly larger for the target on the ‘long’ background compared to the ‘short’ background. For targets on the long background, estimations were larger when the prime and target were congruent. Error bars reflect the standard error of the mean
Grasping
Data from the grasping task were analyzed using a 2 (Prime: congruent vs. incongruent) × 2 (Illusion: target on perceptually long illusion, target on perceptually short illusion) repeated measures ANOVA to explore any effects of priming and illusion on the maximum grip aperture (MGA). As seen in Fig. 4, there was a main effect of illusion on grip aperture, F (1,33) = 11.429, p = 0.002, ηp2 = 0.257, but there was no significant effect of the priming, F (1,33) = 1.759, p = 0.194, ηp2 = 0.051. A post hoc analysis showed that the grip aperture was larger when grasping the target on the long illusory background (M = 7.4 cm, SE = 0.10 cm) compared to the short illusory background (M = 7.3 cm, SE = 0.10 cm). There was no significant interaction effect between prime and length on MGA, F (1,33) = 1.125, p = 0.297, ηp2 = 0.033.

Maximum grip aperture results (cm) for the grasping task. Grip apertures were significantly larger when the target was on the ‘long’ background compared to the ‘short’ background for both congruent and incongruent primes. There was no significant effect of priming. Error bars reflect the standard error of the mean
A 2 (Prime: congruent vs. incongruent) × 2 (Illusion: target on perceptually long illusion, target on perceptually short illusion) repeated measures ANOVA was performed to explore any effects of priming and the illusion on the time to maximum grip aperture (MGA) in the grasping task. Results showed no significant effect of priming on the time to MGA during grasps, F (1,33) = 1.760, p = 0.194, ηp2 = 0.051. The illusory background had no significant effect on the time to MGA, F (1,33) = 1.731, p = 0.197, ηp2 = 0.050. There was no significant interaction effect between priming and illusion on time to MGA, F (1,33) = 0.001, p = 0.981, ηp2 < = 0.001.
Illusion effect
A 2 (Task: grasping vs. estimation) × 2 (Prime: congruent vs. incongruent) repeated-measures within-subject ANOVA was performed to compare the effect of the illusion across the grasping and estimation tasks (Fig. 5). There was a significant effect of task, F (1,33) = 5.132, p = 0.030, ηp2 = 0.135, as well as a significant interaction effect between tasks and prime, F (1,33) = 9.303, p = 0.004, ηp2 = 0.220. There was no significant effect of priming on the illusion effect, F (1,33) = 24.092, p = < 0.001, ηp2 = 0.422. The effect of illusion was larger in estimations (M = 0.316 cm, SE = 0.091) compared to grasps (M = 0.104 cm, SE = 0.031), p = 0.030. Post hoc analyses show that the illusion effect was greater when the prime and target were congruent for estimations (M = 0.510 cm, SE = 0.101) compared to grasps (M = 0.131 cm, SE = 0.043), p = 0.030. Furthermore, in the estimation task the effect of the illusion was greater when the prime and target were congruent (M = 0.510 cm, SE = 0.101) rather than incongruent (M = 0.122 cm, SE = 0.100), p = 0.030. Importantly, one-sample t tests (Table 1) showed that there was a significant effect of the illusion during grasps and estimations with congruent priming.

Effect of the illusion on all five conditions. The illusion influenced both estimations and grasps, but the effect on estimations was significantly larger than on grasping. When the prime and target were congruent, the illusion had a greater effect on estimations compared to grasps. For estimations, the illusion had a greater effect when the prime and target were congruent rather than incongruent. Error bars reflect the standard error of the mean
Visual feedback
In the full vision trials, participants were able to see their hand throughout the reach and grasp motion. In the partial vision trials, participants were only provided visual feedback of their hand during the first half of the reach and were not able to see their hand make contact with the target. As can be seen in Fig. 6, there was a significant effect of the availability of visual feedback on grip aperture, F (1,33) = 10.787, p = 0.002, ηp2 = 0.246. Participants had larger grip apertures when they had to grasp the target with partial visual feedback (M = 7.4 cm, SE = 0.10 cm) compared to when they had full visual feedback (M = 7.1 cm, SE = 0.10 cm). There was a significant effect of the illusion, F (1,33) = 9.742, p = 0.004, ηp2 = 0.228, where participants had larger grip apertures for targets on the long illusory background (M = 7.3 cm, SE = 0.10 cm) compared to the short illusory background (M = 7.2 cm, SE = 0.10 cm). There were no significant interaction effects between vision and illusion, F (1,33) = 1.311, p = 0.260, ηp2 = 0.038.

Average grip aperture for different vision conditions in the grasping task (cm). In both full and partial vision conditions grip apertures were larger when the target was on the long illusory background compared to the short illusory background. Average grip apertures were larger when participants had partial vision during the reach compared to when they had full vision throughout the reach. Error bars reflect the standard error of the mean
There was a difference in time to MGA for partial vision and full vision conditions, F (1,33) = 34.426, p < 0.01, ηp2 = 0.511. Participants grasped the target faster when they had partial visual feedback (M = 5.355 s, SE = 0.055 s) compared to when they had full visual feedback (M = 5.705 s, SE = 0.079 s). There was no effect of the illusion on time to MGA, F (1,33) = 0.18, p = 0.895, ηp2 = 0.001, or an interaction effect between illusion condition and vision on time to MGA, F (1,33) = 2.855, p = 0.100, ηp2 = 0.080.
Discussion
The purpose of this study was to understand the nature of interactions between the dorsal and ventral streams by examining how grasping and manual estimations are influenced by the combined effects of visual priming and a visual illusion. To accomplish this, participant grip aperture was examined during a perception-based estimation task and an action-based reach and grasp task toward a three-dimensional target placed over an illusory background. Results from this experiment showed that while manual estimations were susceptible to the influence of both illusions and priming, grasping was influenced by the illusion but not by priming. Overall, these results indicate that while the dorsal and ventral streams function separately in certain instances, they do interact to complete goal-directed movements (Carther-Krone et al. 2020; Goodale and Haffenden 1998; Whitwell et al. 2018).
Perception
Results of the perceptual estimation task showed that grip apertures were larger for targets on the long illusion compared to the short illusion. This finding is consistent with existing literature on illusions and ventral stream-based actions (Haffenden and Goodale 1998; Katsumata 2019; Whitwell et al. 2016; Whitwell et al. 2018). This finding provides evidence to support the claim that perception-based tasks like estimations, which are processed predominantly by the ventral stream, are highly susceptible to the effects of illusions (Foley et al. 2015). Since manual estimations require no action toward the target, the visual information needed to estimate the length of the target comes from the ventral stream. Ventral stream processing accounts for dimensions of the scene as a whole (i.e. target and illusory background) rather than only for specific objects (i.e. the target) (Kravitz et al. 2013), it is, therefore, more susceptible to the illusory effect. This results in estimations being influenced by the illusion.
It was also hypothesized that an influence of the prime would be observed on manual size estimations. Estimations were predicted to be shorter when the prime was the ‘short’ illusion and the target was the ‘long’ illusion compared to when the prime was the ‘long’ and the target was the ‘short’ illusion. Results from this experiment showed that while there was no significant effect of priming by itself on estimations, there was a statistically significant interaction effect between illusions and priming on estimations. When the prime and target were congruent, targets on the long background were estimated to be larger than targets on the short background. No such effect was observed in the incongruent priming condition. These results suggest that the memory of the prime stimulus influences the estimation of the target length. For example, when participants are primed with the short illusion and have to estimate the length of the target on the long illusory background, they estimate it to be shorter than they would if the prime was of the long illusion and the target was on the long illusion.
This interaction effect is particularly interesting due to participants having a full vision of their hands while making estimations. It would have been expected that the visual information available while making an estimation would have replaced any influence of the memory of the prime. However, in this case, an additive effect was observed where a congruent long illusion made estimations larger and a congruent short illusion made estimations smaller. This finding may be attributed to the time duration between the prime viewing period and start of that estimation task, which was 175 ms. This time duration was less than the 2-s duration it takes for information to decay in short-term memory (Hesse and Franz 2009; Roche et al. 2015). Additionally, there is evidence that actions that occur after a time delay are more likely to rely on perceptual representations stored in memory rather than real-time information (Hu et al. 1999). Therefore, the 175 ms time delay may have contributed to the interaction between the memory of the prime and the perceived size of the illusion.
Action
In Goodale and Milner’s (1992), two visual stream hypothesis, the dorsal stream utilizes visual information to guide our actions. The dorsal stream operates in real time (Buneo and Andersen 2006; Goodale et al. 2004; Goodale and Milner, 1992) and due to the precise maneuvering required to carry out most actions, it is expected to be more resistant to perceptual influences (Goodale 2008) . The findings from the present study indicate that grasps are affected by perceptual influences in partial vision conditions (Aglioti et al. 1995; Carther-Krone et al., 2020; Goodale and Haffenden 1998; Whitwell et al. 2018), but not when full visual information is available. It was observed that grip apertures in the grasping task were influenced by the illusory background but not priming. Participants opened their hands wider when the target was on the long illusory background compared to the short illusory background. Previous research on the effects of illusions on grasping is mixed, with some studies finding that grasps are resistant to the effects of illusions (Agliotti et al. 1995; Cesanek et al. 2018; Goodale and Haffenden 1998; Katsumata 2019; Whitwell et al. 2018) and other studies showing that grasps are influenced by illusions (Bruno and Franz 2009; Heath et al. 2005; Kopiske et al. 2016). The present experiment also showed that the time to MGA was not affected by either illusions or priming, supporting the idea that overall, there was less of an influence of perceptual factors on grasping.
During the partial viewing condition, it could be argued that the illusion is influencing the grasp because the information in the first part of the grasp is seen more recently than the originally presented prime. The dorsal stream’s action plan may be updated to represent the information seen during grasp initiation rather than the memory of the prime. However, once visual feedback was removed, the dorsal stream was no longer able to use online feedback to correct the action plan. To compensate for this missing information, the representation of the target in the context of the illusion seen at the start of the grasp may have been accessed during the grasp causing an effect of the illusion. Consequently, grasps were affected by the illusory background but not priming. This representation may have been processed by both the dorsal and ventral streams to execute the rest of the grasp. While this result by itself is insufficient in understanding the exact mechanisms of dorsal and ventral interactions during grasping, this finding does provide novel insight into the outcome of the combined effects of illusions and priming on grasps.
Analyses of the effects of the illusion further reinforced the above findings on how the two visual streams interact to mediate grasping. Overall, illusions had a stronger effect on ventral stream-guided estimations than dorsal stream-guided grasps, supporting Milner and Goodale’s (1992) two visual stream model. We observed that priming plays a role in mediating the influence of the illusion on estimations. The difference in manual estimations for congruent priming was significantly greater than for incongruent priming. When participants saw a target that did not match the previously shown prime, it perhaps caused them to revise their stored representation of the stimulus. When participants were shown a target congruent to the previously shown prime it reinforced the stored representation and strengthened the effect of the illusion.
Despite there being a significant effect of the illusion on grasping, there was no effect of priming during grasps. The lack of priming effect on grasping may support the notion of the dorsal stream processing information in real time, and priming being ineffective for a purely dorsal stream-based action, thus supporting dissociation between the two visual streams. However, we previously noted that the illusory effect on grasping during partial vision grasps may have been due to the influence of the ventral stream on a dorsal stream action. The lack of effect of priming may support the dissociation between the streams and reject the idea that the two streams interact. However, it is important to note that despite the difference in statistical effects, the actual differences in grip aperture are minimal (± 1 mm). As such, this result may not be sufficient to draw a conclusion as to whether the lack of priming effect on grasping is supportive of the two-stream hypothesis or not.
One other way to interpret the effect of priming on estimations and grasps is that incongruent priming minimizes the effect of the illusion. This was observed in estimations where MGAs were significantly smaller for incongruent conditions compared to the congruent conditions. While there were no significant effects of priming on grasps overall, incongruent priming led to smaller grasps on average compared to congruent priming. If it were the case that incongruent primes diminished the strength of the illusion, one possible explanation might be that the memory of the incongruent prime may have been viewed as unreliable once movement toward the target was initiated. The incongruent prime may have prepared the participants for an action that requires a slightly different motor plan than what is required to interact with the target. However, as previously mentioned the difference in grip aperture between incongruent priming is minimal and is not sufficient to make a conclusion. Regardless, this highlights the importance for future studies to examine how multiple perceptual factors together influence dorsally driven tasks. Future studies can address these possibilities by using larger targets in a similar context. The priming paradigm applied in this experiment differs from those used in previous studies (Hesse et al. 2008; Roche and Chainay 2013; Seegelke et al. 2016) in that the present study manipulated the target context but not the target. This differing application of priming was used to focus on the effect of the illusion on grasping, rather than the participant’s ability to grasp targets that vary in size.
Visual feedback
The effect of visual feedback on grasps was analyzed in this study to determine if the Sander Parallelogram illusion was effective enough to influence actions toward it when not under experimentally induced constraints. As such we did not explore the role of priming in conjunction with visual feedback on the illusion. Consistent with previous studies, we found that when full vision is provided for the entire duration of the grasp, the MGA is closer to the real size of the target (Agliotti et al. 1995; Carther-Krone et al. 2020). A previous study by our lab has shown that the grasps are affected by the Sander Parallelogram illusion when vision is available in the early stages of the grasp (625 ms after tone) and when visual information is not available during the grasp (Carther-Krone et al. 2020). The effect of the illusion when vision was available for the early stages of the grasp is like the effect of the illusion on partial vision grasps in the present study where vision was available in the early part of the grasp until wrist velocity reached 5 cm/s.
In the present study, we found that there was a significant effect of the illusion in the full vision condition, indicating that the illusion was indeed influencing the perception of the size of the target. However, it is important to note that participants had smaller grip apertures in the full vision condition compared to the partial vision condition. This shows that when full vision was available the grip aperture was closer to the actual size of the target compared to the partial vision condition. It was also found that participants took longer to reach MGA in the full vision condition compared to the partial vision condition. The delay in time to MGA observed in the full vision condition may be due to full vision grasps occurring with more input from the dorsal stream. When there is full visual feedback available the dorsal stream keeps using this available information to make online adjustments to the grip aperture throughout the grasp to increase grasp accuracy, as shown by the smaller average grip apertures. Partial vision grasps, which have less visual information available have limited information and opportunity to make online adjustments. Consequently, these grasps have a faster time to MGA and are less accurate, as shown by larger average grip apertures (Keefe et al. 2019).
Conclusion
The goal of the present study was to examine the extent to which dorsally processed actions like grasps are resistant or susceptible to ventrally processed stimuli such as illusions and primes as well as understand the nature of interactions between the dorsal and ventral stream. This novel study provides insight into the complex human visual processing system by exploring the nature of interactions between the streams in guiding goal-directed actions. Results showed that ventral stream-guided estimations were more susceptible to perceptual influences than dorsal stream-guided grasps. Specifically, estimations were influenced by both primes and the illusion, while grasps in the partial vision condition were only influenced by the illusion. These results support the notion that in situations where both dorsal and ventral stream input is necessary to perform an action, such as when visual feedback is removed upon action initiation, the visuomotor system uses the most recent visual information made available through online corrections from the dorsal steam as well as the previously stored information in memory from the ventral stream to complete the action as accurately and efficiently as possible. Future studies could explore the role of visual feedback on size perceptions of familiar objects and tools that are used in everyday situations, as well as look at where individuals fixate when interacting with everyday objects. Visual processing plays an important role in our daily activities by enabling us to interact meaningfully with our environment. By exploring the effects of priming and illusion on visual processing and visually guided actions, we can better understand how this complex system operates and how our vision guides our actions.
Data availability
Raw data are available at https://dataverse.lib.umanitoba.ca/dataset.xhtml?persistentId=doi:10.34990/FK2/JUCL7I.
References
Aglioti S, DeSouza JFX, Goodale MA (1995) Size-contrast illusions deceive the eye but not the hand. Curr Biol 5(6):679–685. https://doi.org/10.1016/S0960-9822(95)00133-3
Borra E, Belmalih A, Calzavara R, Gerbella M, Murata A, Rozzi S, Luppino G (2007) Cortical connections of the macaque anterior intraparietal (AIP) area. Cereb Cortex 18(5):1094–1111. https://doi.org/10.1093/cercor/bhm146
Bruno N, Franz VH (2009) When is grasping affected by the Muller-Lyer illusion? A quantitative review. Neuropsychologia 47(6):1421–1433. https://doi.org/10.3200/JMBR.37.3.179-185
Buneo C, Andersen R (2006) The posterior parietal cortex: sensorimotor interface for the planning and online control of visually guided movements. Neuropsychologia 44(13):2594–2606. https://doi.org/10.1016/j.neuropsychologia.2005.10.011
Carther-Krone TA, Senanayake SA, Marotta JJ (2020) The influence of the Sander parallelogram illusion and early, middle and late vision on goal-directed reaching and grasping. Exp Brain Res 238(12):2993–3003. https://doi.org/10.1007/s00221-020-05960-2
Cavina-Pratesi C, Monaco S, Fattori P, Galletti C, McAdam TD, Quinlan DJ, Culham JC (2010) Functional magnetic resonance imaging reveals the neural substrates of arm transport and grip formation in reach-to-grasp actions in humans. J Neurosci 30(31):10306–10323. https://doi.org/10.1523/jneurosci.2023-10.2010
Cesanek E, Campagnoli C, Taylor JA, Domini F (2018) Does visuomotor adaptation contribute to illusion-resistant grasping? Psychon Bull Rev 25(2):827–845. https://doi.org/10.3758/s13423-017-1368-7
Culham JC, Danckert SL, Souza JF, Gati JS, Menon RS, Goodale MA (2003) Visually guided grasping produces fMRI activation in dorsal but not ventral stream brain areas. Exp Brain Res 153(2):180–189. https://doi.org/10.1007/s00221-003-1591-5
Dickinson JE, Green RJ, Harkin GM, Tang MF, Badcock DR (2019) A new visual illusion aspect-ratio context. Vision Res 165:80–83. https://doi.org/10.1016/j.visres.2019.10.003
Faillenot I, Decety J, Jeannerod M (1999) Human brain activity related to the perception of spatial features of objects. Neuroimage 10(2):114–124. https://doi.org/10.1006/nimg.1999.0449
Fogassi L, Gallese V, Buccino G, Craighero L, Fadiga L, Rizzolatti G (2001) Cortical mechanism for the visual guidance of hand grasping movements in the monkey: a reversible inactivation study. Brain 124(3):571–586. https://doi.org/10.1093/brain/124.3.571
Foley R, Whitwell R, Goodale M (2015) The two-visual-systems hypothesis and the perspectival features of visual experience. Conscious Cogn 35(1):225–233. https://doi.org/10.1016/j.concog.2015.03.005
Foster RM, Kleinholdermann U, Leifheit S, Franz VH (2012) Does bimanual grasping of the Müller-Lyer illusion provide evidence for a functional segregation of dorsal and ventral streams?. Neuropsychologia 50(14):3392–3402. https://doi.org/10.1016/j.neuropsychologia.2012.09.026
Franz V (2003) Manual size estimation: a neuropsychological measure of perception? Exp Brain Res 151(4):471–477. https://doi.org/10.1007/s00221-003-1477-6
Freud E, Ganel T, Shelef I, Hammer MD, Avidan G, Behrmann M (2015) Three-dimensional representations of objects in dorsal cortex are dissociable from those in ventral cortex. Cereb Cortex 27(1):422–434. https://doi.org/10.1093/cercor/bhv229
Freud E, Robinson AK, Behrmann M (2018) More than action: The dorsal pathway contributes to the perception of 3-D structure. J Cogn Neurosci 30(7):1047–1058. https://doi.org/10.1162/jocn_a_01262
Goodale MA (2008) Action without perception in human vision. Cogn Neuropsychol 25(7–8):891–919. https://doi.org/10.1080/02643290801961984
Goodale MA, Haffenden AM (1998) The effect of pictorial illusion on prehension and perception. J Cogn Neurosci 10(1):122–136. https://doi.org/10.1162/089892998563824
Goodale MA, Milner AD (1992) Separate visual pathways for perception and action. Trends Neurosci 15:20–25. https://doi.org/10.1016/0166-2236(92)90344-8
Goodale MA, Westwood DA, Milner AD, Goodale MA (2004) Two distinct modes of control for object-directed action. Prog Brain Res 144:131–144. https://doi.org/10.1016/s0079-6123(03)14409-3
Heath M, Rival C, Westwood DA, Neely K (2005) Time course analysis of closed- and open-loop grasping of the Muller-Lyer illusion. J Mot Behav 37(3):179–185. https://doi.org/10.3200/JMBR.37.3.179-185
Heath M, Rival C, Neely K (2006) Visual feedback schedules influence visuomotor resistance to the Müller-Lyer figures. Exp Brain Res 168(3):348–356. https://doi.org/10.1007/s00221-005-0095-x
Hesse C, De Grave DJ, Franz VH, Brenner E, Smeets JB (2008) Planning movements well in advance. Cogn Neuropsychol 25(7–8):985–995. https://doi.org/10.1080/02643290701862399
Hesse C, Franz V (2009) Memory mechanisms in grasping. Neuropsychologia 47(6):1532–1545. https://doi.org/10.1016/j.neuropsychologia.2008.08.012
Holm S (1979) A simple sequentially rejective multiple test procedure. Scand J Stat 6:65–70
Hu Y, Eagleson R, Goodale M (1999) The effects of delay on the kinematics of grasping. Exp Brain Res 126(1):109–116. https://doi.org/10.1007/s002210050720
James TW, Culham J, Humphrey GK, Milner AD, Goodale MA (2003) Ventral occipital lesions impair object recognition but not object-directed grasping: an fMRI study. Brain 126(11):2463–2475. https://doi.org/10.1093/brain/awg248
Jeong SK, Xu Y (2016) Behaviorally relevant abstract object identity representation in the human parietal cortex. J Neurosci 36(5):1607–1619. https://doi.org/10.1523/JNEUROSCI.1016-15.2016
Katsumata H (2019) Attenuation of size illusion effect in dual-task conditions. Hum Mov Sci 67:102497. https://doi.org/10.1016/j.humov.2019.102497
Keefe B, Suray P, Watt S (2019) A margin for error in grasping: hand pre-shaping takes into account task-dependent changes in the probability of errors. Exp Brain Res 237(4):1063–1075. https://doi.org/10.1007/s00221-019-05489-z
Kopiske KK, Bruno N, Hesse C, Schenk T, Franz VH (2016) The functional subdivision of the visual brain: is there a real illusion effect on action? A multi-lab replication study. Cortex 79:130–152. https://doi.org/10.1016/j.cortex.2016.03.020
Kourtzi Z, Kanwisher N (2000) Cortical regions involved in perceiving object shape. J Neurosci 20(9):3310–3318. https://doi.org/10.1523/JNEUROSCI.20-09-03310.2000
Kravitz DJ, Saleem KS, Baker CI, Ungerleider L, Mishkin M (2013) The ventral visual pathway: an expanded neural framework for the processing of object quality. Trends Cogn Sci 17(1):26–49. https://doi.org/10.1016/j.tics.2012.10.011
Masson M, Bub D, Breuer A (2011) Priming of reach and grasp actions by handled objects. J Exp Psychol Hum Percept Perform 37(5):1470–1484. https://doi.org/10.1037/a0023509
Milner AD (2017) How do the two visual streams interact with each other? Exp Brain Res 235(5):1297–1308. https://doi.org/10.1007/s00221-017-4917-4
Milner AD, Goodale MA (2008) Two visual systems re-viewed. Neuropsychologia 46(3):774–785. https://doi.org/10.1016/j.neuropsychologia.2007.10.005
Milner D, Goodale M (2006) The Visual Brain in Action. Oxford University Press. https://doi.org/10.1093/acprof:oso/9780198524724.001.0001
Nielsen LS, Skov L, Jensen H (2007) Visual dysfunctions and ocular disorders in children with developmental delay. I. prevalence, diagnoses and aetiology of visual impairment. Acta Ophthalmol Scand 85(2):149–156. https://doi.org/10.1111/j.1600-0420.2006.00867.x
Oldfield R (1971) The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9(1):97–113. https://doi.org/10.1016/0028-3932(71)90067-4
Ozana A, Ganel T (2020) A double dissociation between action and perception in bimanual grasping: evidence from the Ponzo and the Wundt-Jastrow illusions. Sci Rep 10(1):14665. https://doi.org/10.1038/s41598-020-71734-z
Pisu V, Uccelli S, Riggio L, Bruno N (2020) Action preparation in grasping reveals generalization of precision between implicit and explicit motor processes. Neuropsychologia 141:107–406. https://doi.org/10.1016/j.neuropsychologia.2020.107406
Roche K, Chainay H (2013) Visually guided grasping of common objects: Effects of priming. Visual Cognition 21(8):1010–1032. https://doi.org/10.1080/13506285.2013.851136
Roche K, Verheij R, Voudouris D, Chainay H, Smeets J (2015) Grasping an object comfortably: orientation information is held in memory. Exp Brain Res 233(9):2663–2672. https://doi.org/10.1007/s00221-015-4360-3
Seegelke C, Güldenpenning I, Dettling J, Schack T (2016) Visuomotor priming of action preparation and motor programming is similar in visually guided and memory-guided actions. Neuropsychologia 91:1–8. https://doi.org/10.1016/j.neuropsychologia.2016.07.033
Srivastava S, Orban G, De Mazière P, Janssen P, Srivastava S (2009) A distinct representation of three-dimensional shape in macaque anterior intraparietal area: fast, metric, and coarse. J Neurosci 29(34):10613–10626. https://doi.org/10.1523/JNEUROSCI.6016-08.2009
Ungerleider LG, Mishkin M (1982) Two cortical visual systems. In: Goodale MA, Mansfield RJW (eds) DJ Ingle. MIT Press, Analysis of visual behavior Cambridge, pp 549–586
Valyear K, Chapman C, Gallivan J, Mark R, Culham J (2011) To use or to move: goal-set modulates priming when grasping real tools. Exp Brain Res 212(1):125–142.https://doi.org/10.1007/s00221-011-2705-0
van Sonderen JF, Denier van der Gon JJ (1991) Reaction-time-dependent differences in the initial movement direction of fast goal-directed arm movements. Hum Move Sci 10(6):713–726
Volcic R, Domini F (2016) On-line visual control of grasping movements. Exp Brain Res 234(8):2165–2177
Westwood D, Goodale MA (2003) Perceptual illusion and the real-time control of action. Spat Vis 16(3):243–254. https://doi.org/10.1163/156856803322467518
Whitwell R, Buckingham G, Enns J, Chouinard P, Goodal M (2016) Rapid decrement in the effects of the Ponzo display dissociates action and perception. Psychon Bull Rev 23(4):1157–1163. https://doi.org/10.3758/s13423-015-0975-4
Whitwell RL, Goodale MA, Merritt KE, Enns JT (2018) The Sander parallelogram illusion dissociates action and perception despite control for the litany of past confounds. Cortex 98:163–176. https://doi.org/10.1016/j.cortex.2017.09.013
Wong A, Haith A, Krakauer J (2015) Motor planning [Reviews]. Neuroscientist 21(4):385–398. https://doi.org/10.1177/1073858414541484
Yeatman JD, Weiner KS, Pestilli F, Rokem A, Mezer A, Wandell BA (2014) The vertical occipital fasciculus: a century of controversy resolved by in vivo measurements. Proc Natl Acad Sci 111(48):E5214–E5223. https://doi.org/10.1073/pnas.1418503111
Zachariou V, Nikas CV, Safiullah ZN, Gotts SJ, Ungerleider LG (2017) Spatial mechanisms within the dorsal visual pathway contribute to the configural processing of faces. Cereb Cortex 27(8):4124–4138. https://doi.org/10.1093/cercor/bhw224
Zhong Y, Rockland KS (2003) Inferior parietal lobule projections to anterior inferotemporal cortex (Area TE) in Macaque Monkey. Cereb Cortex 13(5):527–540. https://doi.org/10.1093/cercor/13.5.527
Acknowledgements
This research was supported by a grant from the Natural Science and Engineering Research Council of Canada (NSERC) (Grant no. 04964-2018) held by J.J.M.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Consent to participate
Informed consent was obtained from all individual participants included in the study.
Ethical standards
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Declaration of Helsinki and its later amendments or comparable ethical standards.
Additional information
Communicated by Melvyn A. Goodale.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Senanayake, S.A., Carther-Krone, T. & Marotta, J.J. Priming of the Sander Parallelogram illusion separates perception from action. Exp Brain Res 239, 2207–2220 (2021). https://doi.org/10.1007/s00221-021-06076-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-021-06076-x