
Abstract
Explicit (strategic) and implicit (unconscious) processes play a role in visuomotor adaptation (Bond and Taylor, J Neurophysiol 113:3836–3849, https://doi.org/10.1152/jn.00009.2015, 2015; Werner et al., PLoS ONE 10:1–18, https://doi.org/10.1371/journal.pone.0123321, 2015). We investigated the contributions of explicit and implicit processes to visuomotor adaptation when awareness was manipulated directly vs. indirectly, and asked how these contributions changed over time. Participants were assigned to a Strategy or No-Strategy group. Those in the Strategy group were made aware of the visuomotor distortion directly. Participants were further subdivided into groups to train with a large (60°), medium (40°) or small (20°) visuomotor distortion, providing the potential for awareness to develop indirectly. Participants reached with their respective distorted cursor, followed by a series of no-cursor reaches to assess the contributions of explicit and implicit processes to visuomotor adaptation after every 30 reach training trials. Within the no-cursor reaching trials, participants reached (1) with any strategies they had gained during training (explicit + implicit processes), and (2) as accurately to the target as possible (implicit processes). Results showed that implicit contributions were greatest in the No-Strategy group, took time to develop, and were transient, as partial decay was seen following a 5-min rest. As well, implicit contributions were similar (i.e., plateaued), regardless of the rotation size participants trained with. In contrast, explicit contributions were greatest in the Strategy group, increased with rotation size, and remained consistent over time. Taken together, results reveal that there are notable differences in the stability of explicit and implicit processes and their potential to contribute to visuomotor adaptation depending on if awareness is provided directly.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
To carry out goal-directed movements, the central nervous system needs to transform sensory information related to hand and target locations into appropriate motor commands for the environment in which one is reaching (Wolpert and Kawato 1998; Flanagan et al. 1999; Jeannerod 1999; Kawato 1999). The sensory information used to plan and execute reaching movements is primarily visual and proprioceptive in nature. In general, these two sensory modalities are aligned, such that the position one sees their hand is the same as the position at which they feel their hand. However, situations can arise in which these signals differ, and the visual and proprioceptive signals conflict (e.g., reaching to an object under water). When sensory signals conflict, humans tend to rely on the visual estimate of hand location as opposed to the “felt” or actual position (Rock and Victor 1964; Mon-Williams et al. 1997; Hagura et al. 2007). Thus, movements are corrected based on the visual estimate of the hand.
Evidence for this reliance on visual information can be seen from results arising from visuomotor adaptation paradigms in the laboratory. In these paradigms, participants may train to reach to a visual target while seeing a cursor that is misaligned from their actual hand position (i.e., the cursor is rotated or translated relative to the hand). Initially, participants’ movements are deviated such that the cursor deviates from the target direction. However, participants rapidly adapt their movements so that the cursor will successfully land on the target. Furthermore, when participants reach in the absence of visual feedback, they demonstrate persistent movement deviations in the opposite direction of the visual distortion experienced. For example, if the cursor was rotated clockwise (CW) relative to the hand, participants reach counter-clockwise (CCW) relative to the target. These deviations reflect persistent changes in the motor system and are referred to as-reach aftereffects (Baraduc and Wolpert 2002; Buch et al. 2003; Simani et al. 2007; Cressman and Henriques 2009).
Current models would suggest that, in general, there are two processes that work in parallel but with different time scales to contribute to motor adaptation (Hwang et al. 2006; McDougle et al. 2015; Redding and Wallace 1996, 2001, 2002; Smith et al. 2006). Specifically, Redding and Wallace (2001) proposed that visuomotor adaptation arises due to a strategic perceptual-motor control process and an adaptive spatial alignment process. The first strategic process is an explicit process that contributes quickly to motor adaptation by reducing initial reach errors. In contrast, the second process unconsciously reduces reach errors over time and is present throughout adaptation. Furthermore, this unconscious or implicit process engaged during reach training is responsible for the errors seen in aftereffect trials when participants are instructed to aim to the target. In line with models detailing explicit and implicit contributions to visuomotor adaptation, Smith et al.’s (2006) two-state model indicates that there are two processes that contribute to motor adaptation, which operate on different time scales. In their model, they suggest that a fast process (that quickly reduces initial direction error and decays quickly) and a slow process (that reduces movement error slowly and is relatively stable over time) underlie motor adaptation. Recently, McDougle et al. (2015) have linked these fast and slow processes to explicit and implicit processes in visuomotor adaptation respectively.
The current models, as discussed above, would predict that the implicit process (i.e., adaptive spatial realignment or the slow process) takes time to develop and remains stable over time. In contrast, explicit processes would arise quickly and decay across adaptation trials, as shown by Taylor et al. (2014). While the time course of explicit and implicit processes have been probed and modeled across motor learning trials in which participants continue to reach with perturbed sensory feedback (Smith et al. 2006; Taylor et al. 2014; McDougle et al. 2015), the stability of these processes over time in the absence of altered sensory feedback (i.e., aftereffect trials) is unclear. As suggested above, aftereffect trials are assumed to reflect implicit adaptation, a persistent change in performance. However, recent results would suggest that errors observed on aftereffect trials decay quickly (McDougle et al. 2015). In fact, Zbib et al. (2016) have shown a significant decay in aftereffects following a 5-min break in which participants sat quietly. It is unclear if this decrease in aftereffects is due to changes in implicit and/or explicit processes, as implicit and explicit contributions to aftereffect trials have not previously been probed over time.
In the current research, we examined the stability of implicit and explicit processes to visuomotor adaptation over time. Implicit and explicit processes were evaluated in a series of aftereffect trials when participants were not provided with feedback and hence could not update their movements on a trial-to-trial basis based on visual error experienced. The aftereffect trials were completed using the process dissociation procedure (PDP) introduced by Jacoby (1991) and adopted by Werner et al. (2015). Within this PDP framework, participants are instructed to use the behavior they have learned during the reach training trials (“use the strategies that you have developed from what was learned during training”) or to repress the learned behavior (“do not use any strategies, aim to the target as you would have during baseline”), while they reach to targets in the absence of visual feedback of their hand position (i.e., complete two types of aftereffect trials with specific instructions provided). The inclusion trials, in which participants are to reach using any strategy they picked up during training, are assumed to reflect explicit + implicit processes, whereas the exclusion trials are assumed to reflect implicit processes (Werner et al. 2015). To assess the stability of explicit and implicit contributions to visuomotor adaptation over time, participants completed the exclusion and inclusion reaches without visual feedback within the PDP framework at eight different time points over the course of the experiment. Specifically, PDP trials were completed immediately following a set of 30 reach training trials, and a second time after a 5-min rest. There were four sets of reach training trials. Awareness of the visuomotor distortion was manipulated directly and indirectly across participants, as awareness has been shown to influence the contribution of implicit and explicit processes to visuomotor adaptation (Benson et al. 2011; Werner et al. 2015). To manipulate awareness directly, half of our participants were provided with a strategy regarding the visuomotor distortion, like in Benson et al.’s study (2011), while the other half of participants were not provided with a strategy (Strategy vs. No-Strategy group). Furthermore, we manipulated awareness of the distortion indirectly by changing the magnitude of the distortion experienced by introducing participants to one of three different distortion sizes (20°, 40°, or 60°). We hypothesized that implicit processes would take time to develop, but would remain relatively stable (i.e., across the 5-min testing delay interval). As well, implicit processes were expected to contribute more to visuomotor adaptation for participants in the No-Strategy group than participants in the Strategy group (Benson et al. 2011; Werner et al. 2015). In contrast, explicit processes were expected to decay over time (Mazzoni and Krakauer 2006) and be greater for participants provided with instructions on the perturbation, (i.e., when awareness was provided directly), compared to participants in the No-Strategy group (Benson et al. 2011; Werner et al. 2015).
Methods
Participants
Sixty-three participants (27 males), aged 18–27 years old (M = 22, SD = 2.62), were recruited from the University of Ottawa community. Participants were naïve to the purpose of the study and had no previous experience with the task. Prior to beginning testing, all participants were confirmed to be right-handed by the Edinburgh Handedness Inventory (M = 76.42, SD = 24.37) (Oldfield 1971). In addition, participants self-reported to have normal or corrected-to-normal vision, and no neurological, sensory or motor dysfunction. All participants provided written informed consent and the experiment was approved by the University of Ottawa Research Ethics Board. Each participant completed testing in one session, which lasted approximately 70 min. Following testing, one participant’s data from the Strategy group who trained with the 60° rotation was excluded from analyses due to the participant reporting that they stopped following instructions part way through the experiment.
Apparatus
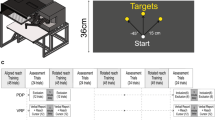
Participants were seated in front of the experimental apparatus. They were instructed to adjust the height and distance of chair to ensure they were seated comfortably and could reach to all possible target locations during the testing session (Fig. 1a). Participants were asked to firmly grip a vertical handle of a two-joint robot manipulandum (BKIN technologies; Kingston, Canada) with their right hand. They were then required to make reaching movements to three visual targets within a 70 cm × 36 cm workspace (Fig. 1b). The targets were projected onto a horizontal reflective surface from a downward facing computer monitor (EzSign model 47LD452B; refresh rate: 60 Hz; LG, Seoul, South Korea), that was placed 20.5 cm above the reflective surface. The robot manipulandum was 20.5 cm below the reflective surface, so that the visual stimuli appeared to lie in the same plane as the reaching hand. When they were ready to begin testing, participants positioned their hand, represented by a visual cursor (green circle, 1 cm in diameter), at the home position. The home position was indicated by a white circle, 1 cm in diameter, at the midline of the body of each participant, approximately 20 cm in front of the participant’s chest. The three visual targets, represented by yellow circles, 2 cm in diameter, appeared 15 cm from the home position at positions corresponding to straight ahead, 40° clockwise (CW) and 40° counter-clockwise (CCW) relative to straight ahead. Participants were prevented from seeing their right hand by a black cover placed between their shoulders and apparatus. The light in the room was turned off.

Experimental set-up and design. a Side-view of the experimental set-up. Participants were instructed to grasp the handle of the robot manipulandum with their right hand. b Top-view of the apparatus illustrating the dimensions of the experimental workspace. The home position (as represented by the light gray circle) was not visible to participants throughout testing. The three reach targets (filled white circle plus two gray circles, 2 cm in diameter), were located straight ahead, 40° clockwise (CW) and 40° counter-clockwise (CCW) relative to straight ahead, at a distance of 15 cm from the home position. During reach training trials with the rotation, the cursor's trajectory (solid black line) was rotated 20°, 40° or 60° CW relative to the participant's hand trajectory (dashed black line). c Overall breakdown of testing blocks completed by all participants, and breakdown of one set of PDP trials. The order of “exclusion” and “inclusion” trials was counterbalanced across participants. In addition, the order of these trials was the same for Time 1 (Initial) and Time 2 (5-min delay), such that half of the participants started with exclusion trials, the other half started with inclusion trials
Experimental procedure
Participants were pseudo-randomly assigned to one of six subgroups. Specifically, participants were assigned based on Aiming Instructions, such that half of the participants were assigned to the Strategy and No-Strategy groups (n = 32 and n = 31 in each group, respectively). These groups were then further subdivided into three smaller groups that trained with a cursor that was rotated 20° (Strategy: n = 10 and No-Strategy: n = 11), 40° (Strategy: n = 11 and No-Strategy: n = 10) or 60° (Strategy: n = 10 and No-Strategy n = 10) CW relative to hand motion.
Once a participant verbally confirmed that they were seated comfortably, the experimenter briefly explained the tasks that would be completed during the testing session. All participants completed four testing Blocks within the testing session (Fig. 1c). Within each testing Block there was 1 set of 30 reach training trials followed by 36 process dissociation procedure (PDP) trials. In the first set, the 30 reach training trials were completed with the cursor accurately representing the participant’s hand position (i.e., the cursor was aligned with his or her actual hand position). In the remaining three sets (Blocks 2–4), the reach training trials were completed such that the cursor representing the hand was rotated by 20°, 40° or 60° CW relative to the hand’s reaching trajectory depending on the subgroup the participant was assigned to. Therefore, participants completed 120 reach training trials in total and 144 PDP trials. Finally, the experiment concluded with participants completing a post-experiment questionnaire, as used by Benson et al. (2011), to assess their awareness of the distortion.
Reach training trials
Participants were instructed to reach to one of three visual targets as quickly and accurately as possible while grasping the vertical robot handle with a firm, but comfortable grip. The hand, as represented by the cursor, started at the home position and was maintained at this position for 500 ms, after which one of the three visual targets appeared with equal probability. Once the target appeared, the home position disappeared and was not shown again during the rest of the reach training trials. The cursor feedback appeared once the hand had moved 7 cm from the home position, which corresponded with the point where participants generally achieved peak velocity. In the first set of reach training trials in Block 1 (baseline set), the cursor was aligned with the actual position of the participant’s hand. Thus, if the participant moved his or her hand directly to the target, the cursor moved directly to the target as well. During the next three sets of reach training trials (Blocks 2–4), the cursor was rotated relative to the hand’s reach trajectory by either 20°, 40° or 60° CW, depending on the subgroup the participant was assigned to. During the 30 reach training trials, participants would randomly reach to all three targets ten times. Upon movement completion, (i.e., when the cursor successfully landed on the target), both the target and cursor disappeared. Participants then moved their hand back to the home position by way of a linear grooved path, such that if they attempted to move outside of the designated path, a resistance force [proportional to the depth of penetration with a stiffness of 2 N/mm and a viscous damping of 5 N/(mm/s)] was produced perpendicular to the grooved wall. The position of the robot handle was recorded at all times at a sampling rate of 1000 Hz and a spatial accuracy of 0.1 mm.
Strategies provided during the reach training trials
As mentioned above, participants were assigned to either the Strategy or No-Strategy groups. During a 5-min break before the first set of rotated reach training trials (in Block 2, see Fig. 1c), the Strategy group was provided with a strategy on how to counteract the rotation that would be introduced. They were also reminded of this strategy before each subsequent set of reach training trials with the rotated cursor (i.e., in Blocks 3 and 4). Instructions were similar to those provided by Benson et al. (2011). Specifically, participants were shown a picture of a modified clock face with the numbers 0–17 around the perimeter, separated by 20°. The experimenter then explained the nature of the distortion (i.e., a CW rotation of 20°, 40° or 60°) using the clock face and showing how the distortion should be counteracted at one of the three potential target locations. For example, participants in the 20° Rotation subgroup were told to aim to the 15 in order for the cursor to land on the left target (located at 16). The participant then drew on the clock face where they had to reach to in order to counter the rotation for the other two target locations. Once participants could accurately draw on the clock face where they were required to reach to and they verbally confirmed that they understood the task, they began the rotated reach training trials (all participants achieved these criteria after one instruction trial, except for one participant who needed the instructions repeated a second time). During this period, the No-Strategy group was not provided with a strategy regarding the upcoming perturbation. Instead, they were instructed to sit quietly for 5 min (Fig. 1c). Before beginning their sets of reach training trials, they were instructed to aim their cursor to the target.
Process dissociation procedure (PDP trials)
These trials were used to dissociate explicit and implicit contributions to visuomotor adaptation. Within each Block of trials, participants completed nine “inclusion” and nine “exclusion” PDP trials two times, equally distributed across the three targets that randomly appeared (Fig. 1c). Similar to Werner et al. (2015), in the inclusion trials participants were instructed to “use what you have learned during training trials,” and in the exclusion trials they were instructed to “refrain from using what was learned during training trials, and perform the movements as you were during the baseline.” Based on results attained by Werner et al. (2015) and the current results presented below, it is evident that participants were able to turn off their strategy in the exclusion trials as instructed. The order in which these inclusion and exclusion trials were completed was counterbalanced across participants. Participants completed 18 trials (nine inclusion + nine exclusion trials) immediately after each set of reach training trials and then the other 18 trials again after a 5-min rest (Fig. 1c). The order of exclusion and inclusion trials was the same before and after the 5-min rest interval, as well as across all Blocks of testing.
All PDP trials were similar to the reach training trials, such that participants were instructed to continue grasping the robot handle with a comfortable, yet firm grip. The hand position was maintained at the home position for 500 ms before a visual target appeared in one of the three target positions. Participants were then instructed to reach as quickly and accurately as possible to the visual target using either what they had learned during training (Inclusion trials) or like they had during the baseline Block (Exclusion trials). However, during this task, no visual feedback (i.e., no cursor) representing the hand was provided. Participants’ reaches were considered complete once they had maintained a final hand position for 1000 ms. Upon movement completion, the visual target disappeared and the participant moved his or her hand back to the home position by way of the linear grooved path.
Post-experiment questionnaire
At the end of the experiment, participants answered a post-experiment questionnaire to assess their awareness of the perturbation and any strategy used to counteract it. The questionnaire was adopted from Benson et al. (2011), and asked questions such as: (1) did you notice the task get harder at any point? (2) Do you know why it became harder? (3) How many degrees (was it rotated) did you push? Based on their responses, participants were classified as aware or unaware of the distortion. As done by Benson et al. (2011), participants were required to indicate the presence of a perturbation and accurately describe the perturbation to be classified as being aware (i.e., followed the left path on the diagram shown in Benson et al. (2011) and claimed to have used a strategy to counteract the perturbation, reported the difference in the intended cursor trajectory or characterized a perturbation as being a rotation). All other participants were classified as not aware. At the end of the questionnaire, participants were debriefed about the nature and purpose of the study. In addition, they verbally confirmed the use of their data and were thanked for their participation.
Data and statistical analysis
Reaching performance during all trials was analyzed using custom written programs in MATLAB. First, the start and endpoint of each reaching movement (all Reach Training trials and PDP trials), were defined using a velocity criterion, such that the start and end of the movement were selected as the points in time when velocity first increased above or decreased below 0.01 m/s for 50 ms, respectively. For each of these reaching profiles, we then determined the angular error at peak velocity (PV), where PV angular error was defined as the angular difference between a movement vector (from the home position to peak velocity) and a reference vector (joining the home position and the target).
PV angular errors were averaged across three consecutive reach training trials for each participant to form ten bins of reach training trials in each Block. From these reaching errors at PV, we first looked to establish that all participants reached in a similar manner during aligned reach training, (i.e., during Block 1, the baseline set of reach training trials), using a 2 Aiming Instructions (Strategy vs. No-Strategy) × 3 Rotation Size (20°, 40° vs. 60°) × 2 Bin (average of first 3 bins vs. average of last 3 bins) mixed analysis of variance (ANOVA) with repeated measures (RM) on the last factor. Having established that all participants had similar PV angular errors during the baseline set of reach training trials, we then looked to analyze the sets of reach training trials completed when a cursor rotation was introduced (i.e., in Blocks 2–4), to determine if performance on reach training trials was dependent on aiming instructions and/or rotation size. Mean initial reach errors for each of these three sets of reach training trials were determined for each participant by averaging their PV angular errors over the first three bins of trials (i.e., nine trials in total) and then subtracting their average PV angular error achieved over the last three bins (i.e., nine trials in total) in the aligned reach training trials performed in Block 1. Similarly, mean final reach errors in each set of rotated reach training trials were determined for each participant by averaging their PV angular errors over the last three bins of trials and then subtracting their average PV angular error achieved over the last three bins in the aligned reach training trials performed in Block 1. Mean errors were then subtracted from the magnitude of the distortion introduced (i.e., 20°, 40° or 60°), such that if a participant reached with a mean PV angular error of 60° and the cursor distortion introduced was 60°, then the participant would have a mean PV angular error of 0°. These reach errors with respect to the rotation size were submitted to a 2 Aiming Instructions (Strategy vs. No-Strategy) × 3 Rotation Size (20°, 40° vs. 60°) × 3 Block (the 1st vs. 2nd vs. 3rd set of 30 rotated reach training) × 2 Bin (average of first 3 bins vs. average of last 3 bins) mixed ANOVA, with RM on the last two factors.
With respect to the PDP trials, PV angular errors were collapsed across target positions and averaged across the 9 (1) inclusion or (2) exclusion trials completed at the same time for each participant. Thus, in total, there were eight averages for each of the two types of trials for each participant. Average PV angular errors were used to compute the index (i.e., the magnitude) of implicit and explicit contributions to visuomotor adaptation according to the following formulas:
These indices at each of the two time points (initial vs. 5-min delay) following each of the sets of rotated reach training trials were then calculated relative to the corresponding index achieved following training with the aligned cursor in Block 1. Specifically:
To determine if the Implicit and Explicit Indices changed over time, with instructions, and/or the magnitude of the distortion, we performed a 2 Aiming Instructions (Strategy vs. No-Strategy) × 3 Rotation Size (20°, 40° vs. 60°) × 3 Block (following the 1st vs. 2nd vs. 3rd set of 30 rotated reach training trials) × 2 Time (initial vs. 5-min delay) mixed ANOVA with RM on the last two factors for both the Implicit and Explicit Indices. Of note, data for one participant’s explicit adaptation indices following their first set of rotated reach training trials (initial and 5-min delay) were more than three standard deviations away from the group’s mean indices, and were, therefore, replaced by average performance across the remaining participants in the corresponding group (the 60° Strategy group). For all analyses, differences with a probability of less than 0.05 were considered to be significant and Bonferonni post hoc tests corrected for multiple comparisons were administered to find the locus of these differences for all pre-planned comparisons. In addition to the data reported below, further performance measures are reported in the Supplementary Analyses and Results file (i.e., data related to reaction time (RT), movement time (MT), and adaptation indices expressed as a proportion of reach adaptation and relative to each target).
Results
Post-experiment questionnaire data
We first present our data from the post-experiment questionnaire to show that participants in the No-Strategy group became aware of the distortion indirectly when training with the larger rotations sizes. According to the post-experiment questionnaire, most participants were aware of the visuomotor distortion, as they reported (1) being aware of the distortion, and (2) claimed to have used a strategy (i.e., “pushed” his or her hand left to successfully land the cursor on the target). Specifically, all participants in the Strategy group were classified as being aware of the presence of a distortion, which was expected since they had been provided with a strategy to counter their respective rotation. As well, for the No-Strategy group, 9 out of 10, and 8 out of 10 participants training with the 40° and 60° Rotation Sizes, respectively, were classified as aware. However, only 2 of 11 participants in the No-Strategy group were classified as aware when training with a 20° Rotation. Therefore, it seems when participants trained with the larger rotation sizes, they became aware of the distortion indirectly.
Reach performance during reach training trials
In Fig. 2, we present changes in mean PV angular errors over reach training trials when training with the 20° (A), 40° (B) or 60° (C) cursor rotation for participants in the Strategy group (dark gray line) and No-Strategy group (light gray line). For trials during Block 1 (baseline), when participants reached with a cursor that was aligned with their hand, we see that in general, all participants were fairly accurate in reaching to the target, regardless of aiming instructions or rotation size. Since the Strategy group only received their strategy following the baseline trials, we would expect there to be no differences between the Strategy and No-Strategy groups during Block 1. As expected, ANOVA revealed that there was no significant main effects of Aiming Instructions [F(1, 56) = 0.366, p = .547], Rotation Size [F(1, 56) = 2.302, p = .109], or Bin [F(1, 56) = 0.406, p = .527] and no significant interactions (all ps > 0.139). Thus, all participants reached similarly during baseline, with a mean reach error of 2.55° (SD = 1.87) left of the target.

Reach Training Performance. Mean PV angular errors when participants trained with a cursor that was aligned (Block 1) or rotated clockwise relative to the hand (Blocks 2, 3, 4) by 20° (a), 40° (b), or 60° (c). If participants adjusted their movements to counteract the distortion, reach errors would be around the dotted line in each figure. The results are presented such that each point represents average performance across three trials (i.e., 1 bin). The black lines correspond to reach errors of the Strategy group and gray lines correspond to reach errors of the No-Strategy group. Error bars represent standard error of the mean
In contrast to the fairly accurate performance when training with the aligned cursor, we see that when the cursor distortion was introduced, participants’ reaches began to deviate to counteract the imposed cursor rotation. Moreover, from Fig. 2, it appears that participants who were in the Strategy group had a performance advantage in that they compensated earlier on (i.e., in less trials) for the imposed rotation than those in the No-Strategy group. However, by the end of each set of rotated reach training trials, participants appeared to have similar errors regardless of whether they were provided with a strategy or not. In fact, by the end of the first set of rotated reach training trials with the 20° cursor rotation, participants in the Strategy group were reaching with an average PV angular error of 2.97° (SD = 0.80) compared to 3.99° (SD = 1.23) for the No-Strategy group. In accordance with these observations, ANOVA revealed that while there was no main effect of Rotation Size [F(2, 56) = 0.465, p = .631], there were significant Aiming Instructions × Rotation Size × Block [F(4, 56) = 5.219, p = .002], and Aiming Instructions × Rotation Size × Bin [F(2, 56) = 5.197, p = .009] interactions. Post hoc analyses revealed that there were no significant differences for the 20° Rotation between the Strategy and No-Strategy groups at any of the time points tested (all ps > 0.164). While the Strategy and No-Strategy 20° Rotation groups displayed similar errors at each time point, the lack of a significant difference does not imply that the same processes were used regardless of instructions. In support of different processes being used depending on instructions, initial errors in the first set of rotated reach training trials (i.e., average of the first 3 bins) for the 40° and 60° Rotation Sizes were larger in the No-Strategy group compared to the corresponding Strategy groups (both ps < 0.001). For the 40° Strategy and No-Strategy groups, no further differences were found (all ps > 0.200), such that initial and final reach errors were similar in the second (Block 3) and third set (Block 4) of rotated reach training trials. As for the 60° Rotation Size, final errors differed between the Strategy and No-Strategy groups at the end of the first set of rotated reach training (Block 2, p < .001) and initial errors in the second (Block 3, p < .001) and third (Block 4, p = .004) set of rotated reach training differed as well, showing a continued advantage for the Strategy group. However, reach errors at the end of the second and third sets of rotated reach training (Block 3 and 4) did not differ between the groups (both ps > 0.139).
PDP data
From Fig. 3a, b, it appears that implicit processes took time to develop and were not that stable (i.e., partial decay of implicit adaptation was evident over a 5-min rest interval). In accordance with these observations, ANOVA revealed a main effect of Block [F(2, 112) = 9.36, p < .001], and post hoc tests indicated that implicit adaptation indices after the second set of rotated reach training (Block 3) were greater than after the first set (Block 2, p < .001). There was no significant difference following the second and third set of rotated reach training trials (Blocks 3 and 4, p = 1). As well, ANOVA revealed a main effect of Time [F(1, 56) = 37.66, p < .001], suggesting that implicit processes decreased from when initially tested to when tested a second time following a 5-min rest. With respect to our manipulation of awareness, as shown in Fig. 3c, d, ANOVA revealed a main effect of Aiming Instructions [F(1, 56) = 5.29, p = .025], such that the No-Strategy group (Time 1: 20° Rotation Size M = 9.3°, SD = 1.35, 40° Rotation Size M = 8.9°, SD = 1.54 and 60° Rotation Size M = 7.2°, SD = 2.3) had a greater implicit adaptation index compared to the Strategy group (Time 1: 20° Rotation Size M = 7.8°, SD = 1.63, 40° Rotation Size M = 5.0°, SD = 1.11 and 60° Rotation Size M = 4.15°, SD = 2.72). Interestingly, as shown in Fig. 3E, F, we found no significant main effect of Rotation Size [F(2, 56) = 2.56, p = .086] and no significant interactions (all ps > 0.168), suggesting implicit processes were similar in magnitude across the three rotation sizes, regardless of the larger distortion size leading to participants becoming indirectly aware.

Implicit adaptation indices. a Mean Implicit index for the No-Strategy group. b Mean implicit index for the Strategy group. For each plot, the solid lines represent indices for participants who trained with the 20° distortion, the large dash lines represent indices for participants who trained with the 40° distortion and the small dash lines represent indices for participants who trained with the 60° distortion [Data related to Time 1 (Initial = black lines) and Time 2 (5-min delay = gray lines) are slightly offset along the horizontal axis]. c, d Mean Implicit Index over Blocks 2, 3 and 4 for participants in the No-Strategy (gray line) and Strategy groups (black line) at Time 1 (c) and Time 2, following a 5-min delay (d). Data is collapsed across rotation sizes. e, f Mean Implicit Index for Block 4 at Time 1 (e) and Time 2, following a 5-min delay (f), for participants in the No-Strategy (gray bars) and Strategy groups (black bars). Data is shown for each of the different rotation sizes. Error bars represent standard error of the mean
Explicit adaptation indices are shown in Fig. 4. There were no significant main effects of Block [F(2, 112) = 0.398, p = .673], or Time [F(1, 56) = 0.496, p = .484], indicating that explicit indices were stable over time. ANOVA did reveal a main effect of Aiming Instructions [F(1, 56) = 15.70, p < .001], such that the Strategy group showed a greater explicit adaptation index than the No-Strategy group. ANOVA also revealed a main effect of Rotation Size [F(2, 56) = 73.57, p < .001], with post hoc analyses revealing that the explicit contribution was greater after training with a 60° Rotation compared to a 40° Rotation (p < .001), which was significantly greater than after training with a 20° Rotation (p < .001). There were no significant interactions (all ps > 0.133), suggesting that as rotation size increased, explicit adaptation indices increased in a similar manner, regardless of whether participants were in the Strategy or No-Strategy group, with participants in the Strategy group displaying the greatest explicit adaptation index (Rotation Size of 20°: Strategy M = 10.3°, SD = 1.6 and No-Strategy M = 0.274°, SD = 1.03; Rotation Size of 40°: Strategy M = 28.95°, SD = 3.9, and No-Strategy M = 20.91°, SD = 4.1; Rotation Size of 60°: Strategy M = 51.23°, SD = 7.67 and No-Strategy M = 37.69°, SD = 2.72).

Explicit adaptation indices. a Mean Explicit index for the No-Strategy group and b Mean Explicit index for the Strategy group. For each plot, the solid lines represent indices for participants who trained with the 20° distortion, the large dash lines represent indices for participants who trained with the 40° distortion and the small dash lines represent indices for participants who trained with the 60° distortion [Time 1 (Initial) = black lines and Time 2 (5-min delay) = gray lines]. Error bars represent standard error of the mean
Discussion
The main objective of the current study was to assess the stability of explicit and implicit contributions to visuomotor adaptation over time when awareness was manipulated directly and indirectly. Participants were assigned to a Strategy or No-Strategy group, and assigned to train with one of three Rotation Sizes (20°, 40°, or 60°). Following sets of 30 reach training trials, participants completed reaches without visual feedback within a process dissociation procedure (PDP) framework. Using this framework, and testing at eight different time points during the experiment, we were able to establish changes in contributions of explicit and implicit processes to visuomotor adaptation over time. Our results revealed that regardless of the distortion size, participants successfully adapted to their respective rotation size during reach training trials, and that these changes were mostly accounted for (~ 93%) by the sum of explicit and implicit indices seen in the PDP trials (i.e., performance on the PDP inclusion trials, see Supplementary Figure S.1). As determined by the PDP trials, implicit learning took time to develop over blocks, and partially decayed following a 5-min break. In general, participants assigned to the No-Strategy group demonstrated greater implicit learning than participants within the Strategy group. Interestingly, the magnitude of participants’ implicit adaptation indices did not vary across rotation sizes. This indicates a similar degree overall of implicit learning, regardless of the size of reach error initially experienced. In contrast, explicit contributions were greater for participants in the Strategy compared to No-Strategy group and relatively stable over time. Together, our results suggest that while implicit contributions change over time and are transient, there is a stability to explicit contributions. Moreover, providing awareness directly modifies the contribution of both explicit and implicit processes to visuomotor adaptation over time, whereas manipulating rotation size influences only the contribution of explicit processes.
During reach training trials with distorted visual feedback of the hand, knowledge of the novel visuomotor environment has been shown to lead to participants correcting their movements in fewer trials compared to participants without knowledge of the perturbation (i.e., faster visuomotor adaptation during reach training trials; Benson et al. 2011). However, by the end of the reach training trials with misaligned visual feedback, the extent of visuomotor adaptation tends to be comparable across participants, regardless of the initial distortion size. In other words, all participants can successfully reach to the target, regardless of initial rotation experienced (Thomas and Bock 2012; Werner et al. 2015; Bond and Taylor 2015). In accordance with these results, we found that participants in the Strategy group compensated more for the distortion when it was first introduced compared to participants in the No-Strategy group. Thus, during rotated reach training, participants who were provided with a strategy showed a performance advantage and compensated for the rotation earlier than those in the No-Strategy group who tended to show greater reaching errors initially. It is important to note that by the end of the rotated reach training trials, all participants reached in a similar manner, regardless of being provided with a strategy or not.
From these reach training trials, it is evident that explicit awareness can be of benefit during visuomotor adaptation. Traditionally, explicit (and hence implicit) contributions to visuomotor adaptation have been assessed by comparing performance between participants who are classified as aware vs. unaware of the presence of the distortion based on post-experiment questionnaires (e.g., Benson et al. 2011). For example, participants are asked if they noticed a change in the task following completion of the experiment. If participants indicate that the task became more difficult, they are then further probed on the perturbation itself. If they are able to indicate and describe (or draw) how the cursor movement did not correspond with the intended cursor trajectory, they are classified as being “aware” of the perturbation (Benson et al. 2011; Heuer and Hegele 2008; Hwang et al. 2006; Werner et al. 2015). Results from participants classified as aware are then contrasted with participants classified as unaware and the role of explicit/implicit contributions to visuomotor adaptation determined. Using a similar questionnaire, we classified participants assigned to the Strategy group as aware, as well as the majority of participants in the No-Strategy group who trained with the 40° and 60° Rotation Sizes. Participants who trained with the 20° Rotation Size and were not instructed on the perturbation remained unaware of the distortion based on our post-experiment questionnaire.
This classification of participants as aware vs. unaware and hence the conclusions one would draw on the contribution of explicit and implicit processes to visuomotor adaptation, differed from the results attained in our PDP trials. Similar to Werner et al., our participants were provided with instructions during the no-cursor PDP reaching trials such that they were to: use what they had learned during reach training (Inclusion trials) or reach like they had during the baseline block (Exclusion trials). By contrasting results between these sets of trials, we were able to establish explicit and implicit contributions to visuomotor adaptation at different points during reach training (e.g., after every 30 trials and following a 5-min delay). Within these trials, we found that all participants provided evidence (albeit limited for some groups of participants) of explicit and implicit contributions to visuomotor adaptation, implying that the classification of participants as aware vs. unaware of the perturbation based on post-experiment questionnaires may be too strict. Specifically, participants classified as aware demonstrated both implicit and explicit contributions to visuomotor adaptation. As well, and somewhat surprisingly, a couple of participants classified as unaware demonstrated explicit adaptation when instructed to use what they had learned in the PDP inclusion trials (see Supplementary Figure S.2).
Time course of explicit and implicit contributions to visuomotor adaptation: implications for current models
Within the current study, we looked to evaluate explicit and implicit contributions to visuomotor adaptation across time. Recently, Taylor et al. (2014) showed that implicit processes took time to develop during reach training. In their paradigm, implicit processes were assessed during reach training trials in which participants had to verbally report the location they planned to aim to in order for the cursor to hit the target using landmarks provided on each trial prior to reaching. Taylor et al. (2014) then contrasted the reported location with the location participants aimed to in order to determine the extent of explicit and implicit contributions to visuomotor adaptation. Participants showed both explicit and implicit contributions to visuomotor adaptation, though the contributions of these processes changed over trials. Specifically, explicit contributions were greatest at the start of the reach training trials, while implicit processes gradually increased over the course of reach training. Similar to these results, we found that implicit processes took time to develop while training with a distortion, such that implicit processes were greater after 60 vs. 30 reach training trials with a rotated cursor. Surprisingly, we found that there was some decay in these implicit processes following a 5-min rest such that a significant decrease in implicit indices was found when tested for a second time following a 5-min break where participants sat quietly between the two sets of PDP trials. This partial decay in implicit processes is in line with the changes in aftereffects reported by Zbib et al. (2016) and shown by Bond and Taylor (2015). In contrast, we found that the contribution of explicit processes to visuomotor adaptation as assessed through PDP trials did not differ across the three blocks of rotated reach training and did not decay when a 5-min delay was inserted following reach training.
As previously discussed in the “Introduction” section, current models suggest there are two processes, which work in parallel, but on different timescales that contribute to motor adaptation (Hwang et al. 2006; McDougle et al. 2015; Redding and Wallace 1996, 2001, 2002). In line with these models, our results suggest that explicit and implicit processes work in parallel to contribute to visuomotor adaptation. Additionally, our results revealed that implicit processes took time to develop. However, these models do not seem to reflect, nor predict the partial decay we saw in implicit adaptation over time, nor the consistent contribution of explicit processes to visuomotor adaptation we observed in our PDP trials. One possible explanation is that the fast process put forth by Smith et al. (2006) has both explicit and implicit components, and does not necessarily reflect an explicit process alone [as was indicated by McDougle et al. (2015)]. Thus, as suggested by Zbib et al. (2016), the partial decay that we see in our implicit indices over a 5-min rest interval reflects a decay of an implicit component whose trial-to-trial behavior is typically accounted for by the fast process in the two rate model of Smith et al. (2006).
The influence of awareness on explicit and implicit contributions to visuomotor adaptation
Recent studies by Day et al. (2016), as well as McDougle et al. (2017), suggest that the extent of implicit adaptation observed in the workspace is direction dependent, such that the greatest implicit adaptation is observed when reaching to targets where one intends to move [i.e., around a target located at a position similar to explicit adaptation and not at the original reach target (e.g., at a position 30° from a target when training with a 45° distortion)]. We assessed implicit and explicit adaptation at the trained targets, not at intended targets. If aiming direction generalization of implicit adaptation was present in the current paradigm, results should reveal greater implicit adaptation at the left and center targets compared to the right target, since participants trained with rotations in the clockwise direction and would compensate for the distortion by reaching in the counter-clockwise direction. However, further analyses determined that implicit processes did not differ significantly between targets (see Supplementary Figure S.2). Thus, while the current study did not directly test for workspace differences in explicit and implicit adaptation, results suggest that the increase we see in implicit processes over time and consistency in explicit processes cannot be accounted for by generalization around one’s intended movement (as would be expected if generalization was dependent on aiming direction).
While we do not observe direction dependent differences in implicit adaptation, we did find that the contribution of implicit and explicit processes to visuomotor adaptation changed with manipulations of awareness. With respect to the role of explicit processes in visuomotor adaptation, we found greater explicit adaptation indices in our Strategy compared to No-Strategy group. In contrast to Werner et al. (2015), we found a greater contribution of implicit processes for our No-Strategy group compared to our Strategy group. Moreover, we found that the magnitude of implicit adaptation appeared to plateau, such that there was a maximum amount implicit processes could contribute to visuomotor adaptation. Recent research has also reported that implicit adaptation may reach a ceiling effect regardless of the rotation size, and hence error size experienced. Specifically, our results corroborate those from Bond and Taylor (2015) and Morehead et al. (2017), who show that in a series of aftereffect trials in which participants are instructed that the distortion had been removed and they are to aim to the target, implicit processes did not scale with the rotation size. For example, the extent of implicit adaptation observed in their aftereffects trials of Bond and Taylor was between 8° and 12°, regardless if participants trained with a cursor that was rotated 15° or 90°. The implicit adaptation index in our No-Strategy group was similar in magnitude at 6°–10°. In contrast, the implicit adaptation index was lower in our Strategy group (approximately 5° overall). While this difference between aiming instructions groups is not surprising, given that implicit processes have been suggested to play a smaller role in visuomotor when strategies are provided directly (Benson et al. 2011), Werner et al. (2015) showed no difference in implicit processes between their No-Strategy group compared to their Strategy group.
Potential differences in our paradigm from Werner et al. (2015) have to do with experimental design and the method of reporting the data. In their study, Werner et al. (2015) had participants train on all three perturbations, which differs from our current study. This training regime, and the potential recognition of a perturbation at different times during the testing session depending on the order in which the rotations were introduced, may have shifted how some of their participants behaved across the experiment. Specifically, it is possible for interference to have occurred across rotations sizes, as participants were required to learn three different visuomotor rotation tasks in the same session. For example, anterograde interference, which refers to impairment in learning a second task following learning of a first task, has been shown when participants were required to learn two force field perturbations one after the other (Sing and Smith 2010).
Our data also differs from the results presented by Werner et al. (2015) in that we show that the values reported in our implicit and explicit adaptation indices approximately sum to the extent of reach adaptation observed (i.e., the sum of implicit and explicit adaptation indices when expressed as a proportion of reach adaptation were approximately 1.08 for the Strategy group and 0.75 and for the No-Strategy group; see Supplementary Figure S.1), whereas the values reported by Werner et al. appear to be approximately 0.80 for their Strategy group and 0.55 for their No-Strategy group. Thus, it appears that the implicit and explicit indices reported by Werner et al. (2015) do not entirely account for the changes in reaches observed during the reach training trials. Unfortunately, data related to the magnitude of implicit and explicit contributions and reach adaptation are not provided in the study by Werner et al. (2015), but rather expressed in relation to each other. Given the differences in the data reported, it is difficult to determine if differences in results may also lie in the extent of reach adaptation achieved across the two experiments. Based on our results, it seems that reach adaptation can almost be entirely accounted for by Explicit and Implicit Adaptation indices, especially when participants are provided with a strategy. Moreover, it appears that when explicit processes are engaged from the beginning of training based on instructions provided, implicit processes do not have the same opportunity to develop and contribute to visuomotor adaptation, as when awareness is not provided directly.
It is important to note that within our current study and the studies by Taylor et al. (Taylor et al. 2014; Bond and Taylor 2015; Morehead et al. 2017), in which participants reported their aiming direction on each reach training trial and/or were instructed to ignore erroneous visual feedback, participants are required to think about the task continually. Specifically, in our study, participants were instructed to (a) reach to the target with a strategy or any information they may have picked up, and (b) to reach as they would have during baseline, after every 30 reach training trials. While we wanted to test for explicit and implicit contributions on an ongoing basis to establish a timeline, these trials may have indicated to our No-Strategy participants that the environment they were reaching in had, or was going to change, which may in turn have influenced their awareness of the distortion. Notably this assessment was also done in the baseline block, prior to reach training with a rotated cursor, which again could have cued participants that something was going to change. Future research should consider if providing such instructions influences performance or if implicit adaptation saturates even with larger rotation sizes, regardless if participants have become (indirectly) aware of the perturbation or not.
Conclusion
We found that explicit and implicit contributions to visuomotor adaptation differed over time. Specifically, implicit contributions took time to develop and were shown to partially decay following a 5-min rest interval, whereas explicit contributions did not change with time. We also established that modifying awareness directly via instructions influenced explicit and implicit contributions to visuomotor adaptation, such that providing a strategy resulted in greater explicit contributions to visuomotor adaptation and smaller implicit contributions. Importantly, implicit contributions did not increase in magnitude across rotation sizes, implying a ceiling level, whereas explicit contributions increased in magnitude with rotation sizes. These differences in the No-Strategy vs. Strategy groups were observed even though the majority of participants were classified as aware of the perturbation on a post-experiment questionnaire. Taken together, our results suggest that awareness is detrimental to implicit contributions to visuomotor adaptation, as explicit processes tend to dominate. Moreover, the results highlight the importance of instructions when evaluating reaching performance in no-cursor trials, as they can modulate reaching errors.
References
Baraduc P, Wolpert DM (2002) Adaptation to a visuomotor shift depends on the starting posture. J Neurophysiol 88:973–981
Benson BL, Anguera JA, Seidler RD (2011) A spatial explicit strategy reduces error but interferes with sensorimotor adaptation. J Neurophysiol 105:2843–2851. https://doi.org/10.1152/jn.00002.2011
Bond KM, Taylor JA (2015) Flexible explicit but rigid implicit learning in a visuomotor adaptation task. J Neurophysiol 113:3836–3849. https://doi.org/10.1152/jn.00009.2015
Buch ER, Young S, Contreras-Vidal JL (2003) Visuomotor adaptation in normal aging. Learn Mem 10:55–63. https://doi.org/10.1101/lm.50303
Cressman EK, Henriques DYP (2009) Sensory recalibration of hand position following visuomotor adaptation. J Neurophysiol 102:3505–3518. https://doi.org/10.1152/jn.00514.2009
Day KA, Roemmich RT, Taylor JA, Bastian AJ (2016) Visuomotor learning generalizes around the intended movement. Cogn Behav. https://doi.org/10.1523/ENEURO.0005-16.2016
Flanagan JR, Nakano E, Imamizu H et al (1999) Composition and decomposition of internal models in motor learning under altered kinematic and dynamic environments. J Neurosci 19:1–5
Hagura N, Takei T, Hirose S et al (2007) Activity in the posterior parietal cortex mediates visual dominance over kinesthesia. J Neurosci 27:7047–7053
Heuer H, Hegele M (2008) Adaptation to visuomotor rotations in younger and older adults. Psychol Aging 23:190–202. https://doi.org/10.1037/0882-7974.23.1.190
Hwang EJ, Smith MA, Shadmehr R (2006) Dissociable effects of the implicit and explicit memory systems on learning control of reaching. Exp Brain Res 173:425–437. https://doi.org/10.1007/s00221-006-0391-0
Jacoby LL (1991) A process dissociation framework: separating automatic from intentional uses of memory. J Mem Lang 30:513–541
Jeannerod M (1999) To act or not to act: perspectives on the representation of actions. Q J Exp Psychol 1999:1–29
Kawato M (1999) Internal models for motor control and trajectory planning. Curr Opin Neurobiol 9:718–727
Mazzoni P, Krakauer JW (2006) An implicit plan overrides an explicit strategy during yisuomotor adaptation. J Neurosci 26:3642–3645. https://doi.org/10.1523/JNEUROSCI.5317-05.2006
McDougle SD, Bond KM, Taylor JA (2015) Explicit and implicit processes constitute the fast and slow processes of sensorimotor learning. J Neurosci 35:9568–9579. https://doi.org/10.1523/JNEUROSCI.5061-14.2015
McDougle SD, Bond KM, Taylor JA (2017) Implications of plan-based generalization in sensorimotor adaptation. J Neurophysiol 118:383–393. https://doi.org/10.1152/jn.00974.2016
Mon-Williams M, Wann JP, Jenkinson M, Rushton K (1997) Synaesthesia in the normal limb. Proc Biol Sci 264:1007–1010
Morehead JR, Taylor JA, Parvin DE, Ivry RB (2017) Characteristics of implicit sensorimotor adaptation revealed by task-irrelevant clamped feedback. J Cogn Neurosci 29:1061–1074. https://doi.org/10.1162/jocn_a_01108
Oldfield RC (1971) The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9:97–113
Redding GM, Wallace B (1996) Adaptive spatial alignment and strategic perceptual-motor control. J Exp Psychol Hum Percept Perform 22:379
Redding GM, Wallace B (2001) Calibration and alignment are separable: Evidence from prism adaptation. J Mot Behav 33:401–412. https://doi.org/10.1080/00222890109601923
Redding GM, Wallace B (2002) Strategic calibration and spatial alignment: A model from prism adaptation. J Mot Behav 34:126–138. https://doi.org/10.1080/00222890209601935
Rock I, Victor J (1964) Vision and touch: An experimentally created conflict between the two senses. Science 143:594–596
Sing GC, Smith MA (2010) Reduction in learning rates associated with anterograde interference results from interactions between different timescales in motor adaptation. Plos Comput Biol 6:1–14
Smith MA, Ghazizadeh A, Shadmehr R (2006) Interacting adaptive processes with different timescales underlie short-term motor learning. PLoS Biol 4:e179. https://doi.org/10.1371/journal.pbio.0040179
Taylor JA, Krakauer JW, Ivry RB (2014) Explicit and implicit contributions to learning in a sensorimotor adaptation task. J Neurosci 34:3023–3032. https://doi.org/10.1523/JNEUROSCI.3619-13.2014
Thomas M, Bock O (2012) Concurrent adaptation to four different visual rotations. Exp Brain Res 221:85–91. https://doi.org/10.1007/s00221-012-3150-4
Werner S, van Aken BC, Hulst T et al (2015) Awareness of sensorimotor adaptation to visual rotations of different size. PLoS ONE 10:1–18. https://doi.org/10.1371/journal.pone.0123321
Wolpert DM, Kawato M (1998) Multiple paired forward and inverse models for motor control. Neural Netw 11:1317–1329
Zbib B, Henriques DYP, Cressman EK (2016) Proprioceptive recalibration arises slowly compared to reach adaptation. Exp Brain Res 234:2201–2213. https://doi.org/10.1007/s00221-016-4624-6
Acknowledgements
This work was supported by a Discovery Grand provided by the Natural Sciences and Engineering Research Council of Canada (E. K. Cressman).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
No conflicts of interest, financial or otherwise, are declared by the author(s).
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Neville, KM., Cressman, E.K. The influence of awareness on explicit and implicit contributions to visuomotor adaptation over time. Exp Brain Res 236, 2047–2059 (2018). https://doi.org/10.1007/s00221-018-5282-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-018-5282-7