
Abstract
Two behavioral estimates of interhemispheric transfer time, the crossed-uncrossed difference (CUD) and the unilateral field advantage (UFA), are thought to, respectively, index transfer of premotor and visual information across the corpus callosum in neurotypical participants. However, no attempt to manipulate visual and motor contingencies in a set of tasks while measuring the CUD and the UFA has yet been reported. In two go/no-go comparison experiments, stimulus pair orientations were manipulated. The hand of response changed after each correct response in the second, but not the first experiment. No correlation was found between the CUD and the UFA, supporting the hypothesis that these two measures index different types of information transfer across hemispheres. An effect of manipulation of stimulus pair orientation on UFAs was attributed to the homotopy of callosal fibers transferring visual information, while an effect of hand switching on CUDs was attributed mostly to spatial compatibility.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The Poffenberger paradigm was invented to estimate the time required for the critical transfer of information from one hemisphere to the other. Poffenberger (1912) presented visual stimuli in the left and right halves of the visual field, and required a motor response by the left or right hand. Visual information from the left visual field is processed in the right hemisphere, whereas visual information from the right visual field is processed in the left hemisphere. Likewise, motor responses from the left hand are controlled by the right hemisphere, and motor responses from the right hand are controlled by the left hemisphere. If the side of the visual field and the side of the hand are discordant (crossed trial), both hemispheres are involved in the task. Interhemispheric transfer of information is then necessary for the production of the motor response. If the side of the visual field and the side of the motor response are concordant (uncrossed trial), only one brain hemisphere’s processing capacity is required during the task, and interhemispheric transfer of information is not strictly necessary. Poffenberger inferred that the crossed-uncrossed difference (CUD) in reaction time (RT) represented an estimate of interhemispheric transfer time (IHTT).
Dimond (1969) created another tachistoscopic method for the study of interhemispheric dynamics. He presented pairs of visual stimuli which had to be compared. On bilateral trials, one stimulus was positioned within each visual hemifield, and a transfer of visual or cognitive information was necessary to decide whether the stimulus pair formed a matching pair. On unilateral trials, both stimuli were positioned within the same visual hemifield, and no such transfer of information was necessary for the stimuli to be compared. A subtraction of RTs from unilateral and bilateral trials for correct responses was thought to yield an estimate of the efficiency of interhemispheric communication.
There are important differences between effects found in the Poffenberger and Dimond paradigms which follow logically from their respective structures. For instance, Poffenberger’s interhemispheric inference does not specify the type of information, visual or premotor, which is transferred across hemispheres, and thus usually considers only simple RT. In contrast, Dimond’s interhemispheric inference does not consider the possibility of a transfer of premotor information. Dimond tasks are typically go/no-go tasks. To determine whether a motor response must be made, the two stimuli must first be compared. The information that is transferred during bilateral trials must be information that allows for such a comparison, such as visual or cognitive information.
Poffenberger first proposed that the CUD was an estimate of IHTT. As expected from this inference, mean CUD for the population has been positive in all Poffenberger tasks reported in two separate meta-analyses, reflecting faster performance when interhemispheric communication is not strictly necessary (Marzi et al. 1991; Braun 1992). Moreover, CUDs are extremely long in callosotomised patients, clearly implicating the corpus callosum as a structure necessary for interhemispheric communication in Poffenberger’s paradigm (Savazzi et al. 2007).
Dimond’s measure cannot be interpreted as a simple estimate of IHTT. RTs of bilateral trials are often shorter than RTs of unilateral trials (Banich and Belger 1990; Belger and Banich 1992; Leblanc-Sirois and Braun 2014), reflecting an advantage linked to cooperation between both hemispheres in these tasks rather than a cost of interhemispheric relay. Dimond’s so-called measure of interhemispheric dynamics must be interpreted as a combination of two sources of delay (Leblanc-Sirois and Braun 2014), one linked to interhemispheric transfer and one linked to processing delays within the hemispheres. A modest correlation between Poffenberger’s and Dimond’s measures, however, would still be expected if both CUD and the bilateral–unilateral difference partly reflected interhemispheric transfer of visual information.
Attempts to obtain correlations between different measures thought to be influenced by IHTT have been made, and have included visual evoked potential (VEP) estimates of IHTT. The onset of stimulus display in visual tasks creates a pattern of electrical activity in the visual cortex of the hemisphere contralateral to stimulus presentation. An echo of this pattern of activity with reduced amplitude also occurs in the hemisphere ipsilateral to presentation (Lines et al. 1984; Rugg et al. 1985; Saron and Davidson 1989), approximately 3 ms later when measured in central electrodes and 10–20 ms later when measured over the occipital cortex. This echo is absent in patients whose corpus callosum has been surgically sectioned (Brown et al. 1999), reflecting the important role of the corpus callosum for the generation of VEP-IHTT.
Two studies (Saron and Davidson 1989; Friedrich et al. 2017) have attempted to document correlations between CUD and VEP-IHTT, but failed to obtain significant effects. Electric activity measured at frontal, not posterior, electrode sites has also been shown to better predict CUD (Thut et al. 1999), supporting the hypothesis that CUD reflects the latency of transfer of premotor information, not visual information. In contrast, VEP-IHTT measured at posterior electrodes were correlated with Dimond’s measure in two separate studies (Brown and Jeeves 1993; Larson and Brown 1997), supporting the view that the transfer of visual information, not premotor information, is important in simple versions of the Dimond task.
Previous attempts to manipulate Poffenberger’s CUD by modifying visual characteristics of Poffenberger tasks have generally not been successful. A meta-analysis comparing CUD between tasks with different characteristics (Braun 1992) did find that larger distances between the stimuli and the vertical meridian and decreased luminance yielded longer CUDs. However, most relevant studies failed to obtain a significant effect of stimulus eccentricity or luminance on CUDs (Berlucchi et al. 1971, 1977; Milner and Lines 1982; Lines et al. 1984; Clarke and Zaidel 1989). Moreover, the strongest effect documented by Braun’s (1992) meta-analysis was a shorter CUD when the thumb, rather than the index, was used for responses. The same meta-analysis found no effect of stimulus size on CUD.
However, the manipulation of motor demands within the Poffenberger task has yielded significant results within single studies. Braun et al. (2004) found that CUDs were halved in a simple reaction time task when arms were crossed, though Berlucchi et al. (1977) found no such effect in a similar task and Anzola et al. (1977) obtained this effect only in a choice reaction time task, not in a simple reaction time task. Larocque et al. (2001) found that CUDs in a simple reaction time task were longer when a sequence of three keypresses was necessary rather than just one keypress, and were also longer when the finger of response alternated between the index and the middle finger. In a test exploring both motor and visual task manipulations, (Iacoboni and Zaidel 1995) obtained an effect of response complexity on CUD in the absence of an effect of stimulus complexity.
Unlike Poffenberger’s measure, Dimond’s measure has shown sensitivity to manipulation of visual elements of the task. In particular, when the invisible line separating two stimuli of a pair was oblique relative to the horizon, a larger unilateral field advantage (UFA) was obtained than when it was horizontal, in four separate experiments (Desjardins et al. 2009; Braun et al. 2011, 2015; Leblanc-Sirois and Braun 2015). This effect was attributed to the homotopy of callosal connections, perpendicular to the falx, which more often link areas of visual cortex that process parts of the visual field that are symmetric across the vertical meridian.
Poffenberger’s and Dimond’s measures of interhemispheric dynamics have previously been obtained in the same task, and attempts to explore the relation between the two have been made. In a Dimond task with bilateral presentations, unilateral left field presentations, and unilateral right field presentations, Braun et al. (1997) previously reported significant correlations between Poffenberger’s and Dimond’s “measures”. However, Poffenberger’s measure in a “go on same” condition requiring identical stimuli correlated with Dimond’s measure in a “go on different” condition requiring non-identical stimuli, and vice-versa. Moreover, the significant correlations were between different dependent variables: no correlation was reported between these two RT measures within one experimental condition. To the best of our knowledge, this result has not been replicated, and no attempt to experimentally separate Dimond’s and Poffenberger’s measures within the same set of tasks has yet been made.
Purpose of the current experiments
The current set of experiments aimed to study differences between two measures of interhemispheric transfer of information, Dimond’s measure and Poffenberger’s CUD, during visual go/no-go tasks. More precisely, the experiments aimed to demonstrate that Dimond’s measure primarily indexes interhemispheric transfer of visual information, while Poffenberger’s CUD primarily indexes transfer of premotor information. For this purpose, one visual element was manipulated within each of two experiments, and a difference between the motor demands separated both experiments.
Experiment 1
Methods
Task
Twelve right handed participants (6 male, 6 female) aged 18 to 30 were recruited among university students. Handedness was tested with the Collin and Braun questionnaire (unpublished), a checklist of 19 questions about the preferred hand for actions such as cutting bread, holding a racquet, and writing. Participants had no history of neurological or psychiatric problems, and were tested for adequate vision with an Optec Vision Tester (Stereo Optical Co., Inc), with a focus on acuity and correct tonic vergence, prior to the experiment.
The go/no-go Dimond task was programmed in ePrime version 1.1 software. Participants viewed the computer screen display through a flat black 57 cm long tube preventing reflections and properly shaping the stimulus field as a circle (see Fig. 1). Stimuli were dark (rgb: 66, 66, 66) and medium (rgb: 128, 128, 128) grey disks displayed on a light grey (rgb: 198, 195, 198) background. Stimuli were presented in pairs. For 80% of all trials, both stimuli of a pair were identical. This high frequency of go trials was chosen to maximize the number of trials for which RT was available. For 20% of all trials, they were different. Stimulus pairs were presented in one of four quadrants positioned above, below, to the left, or to the right of the center-screen fixation point. An equal number of stimulus pairs were presented in each of the four quadrants. The center of each quadrant was positioned 6.4 cm away from the fixation cross. Stimulus pairs were separated by an invisible horizontal, or vertical, oblique slash, or oblique backslash line. An equal number of stimulus pairs were presented in each of the four stimulus pair orientations. The distance between two stimuli of a pair was kept constant at 5.25 cm.

Possible stimulus pair positions in experiments 1 and 2. U Unilateral presentations. B Bilateral presentations. For each trial, two stimuli were presented simultaneously within one quadrant, in one of four pair orientations located to the left, right, top or bottom of the central fixation cross (+). Dotted lines were not visible for the participants. Black, white, and grey disks illustrate the possible location of stimulus pairs, and do not represent the stimuli used during the study. Black and white disk pairs represent cardinal pair orientations, only white disk pairs represent horizontal pair orientations, and both types of grey disk pairs represent oblique pair orientations
Participants had to fixate the fixation cross (+) and press the space bar on the keyboard when two stimuli presented simultaneously in peripheral vision were identical. A minimum of 2560 trials per participant, distributed between 8 trial runs separated by short breaks, was used. Response hand did not change during a trial run. An AB BA BA AB counterbalancing was used for response hand. One-half of the participant group began with the left hand, while the other half began with the right hand.
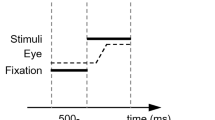
Each trial began with a fixation point displayed between 350 and 2000 ms, according to a logarithmic ramp favoring shorter durations. A pair of stimuli was then displayed for 60 ms. The fixation remained on screen during stimulus display. Following stimulus onset, participants had 750 ms to respond. After 750 ms or after a response was registered, the fixation remained on screen for 500 ms if the response or non-response was correct. If a commission error (responding on a no-go trial) was registered, a “wrong response” error message was instead displayed for 500 ms. If an omission error (failure to respond on a go trial) was registered, a “too long” error message was displayed for 500 ms.
Analyses of unilateral-bilateral differences for this task have previously been reported only for RT (Leblanc-Sirois and Braun 2015). RT results are reported again here to allow a comparison with Experiment 2 during which motor demands are different. Accounts of both experiments are here accompanied by new analyses of error rates and focus on different measures. Analyses of left–right hand and visual field interactions in RT and error rates for this task have not previously been reported in the scientific literature.
Statistical methods
Two separate series of analyses of variance, respectively, meant to test the significance of Dimond’s and Poffenberger’s measures. The first series of analyses of variance was meant to investigate effects of unilateral and bilateral stimulus pair presentations, and included 4 factors of interest. A 4-level block factor contrasted the four pairs of trial runs in temporal order and provided a measure of practice or fatigue effects. A 2-level response hand factor contrasted left-handed and right-handed responses. A 2-level bilateral vs unilateral factor contrasted pairs of stimuli presented on different sides of the midline of the visual field against stimulus pairs presented wholly within either the left or right visual field. Finally, a 3-level pair orientation factor contrasted pairs of stimuli separated by an invisible line in the shape of a slash, horizontal line, or backslash. Because vertical pairs are likely processed in both brain hemispheres at once during bilateral presentations (Bourne 2006) due to the individual stimuli being on the vertical meridian, they were not used during this analysis. Their inclusion in the task served to distribute attention equally to cardinal and oblique orientations. Because the callosal homotopy hypothesis predicts that horizontal pair orientations should yield a smaller UFA than both oblique pair orientations, the quadratic trend was reported for “pair orientation” and all its interactions. RT was the dependent variable of main interest, but secondary analyses of variance were also run with the rate of commission and omission errors as dependent variables. Only effects of bilateral vs unilateral, pair orientation, and their interactions with other factors were of interest for the present report.
The second series of analyses of variance was meant to obtain a significance measure for Poffenberger’s CUD, and also included 4 factors of interest. There were a 4 level block factor, a 2 level response hand factor and a 2 level visual field factor, contrasting the left and right visual fields. For this analysis, data from stimulus presentations in the top and bottom quadrant were not used. Finally, a 2 level pair orientation factor was used, distinguishing both oblique from both cardinal orientations. The response hand * visual field interaction was of primary interest, because it was an index of Poffenberger’s CUD: Stimulus pairs presented to the left of fixation are expected to yield shorter RTs for left hand responses, and stimulus pairs presented to the right of fixation are expected to yield shorter RTs for right hand responses. The dependent variables were RT as in the previous series of analyses. To avoid repetition, only effects of hand and visual field and their interactions with other variables were reported.
Results
Dimond analyses
Mean RT of all trials of Experiment 1 included in the Dimond analysis was 419 ms. An effect of bilateral vs unilateral presentation was found (F(1,11) = 6.03, p = 0.032), with longer RTs for bilateral presentations than unilateral presentations. An effect of pair orientation was also found (quadratic trend: F(1,11) = 11.18, p = 0.007, partial η2 = 0.465), with horizontal pair RTs being shorter than RTs for the pairs of stimuli positioned in oblique orientations. A significant interaction between bilateral vs unilateral and pair orientation was obtained (quadratic trend: F(1,11) = 9.58, p = 0.01), with a non-significant 1 ms BFA for horizontal presentations and a significant (F(1,11) = 12.15, p = 0.005) 7 ms UFA for oblique orientations. The only other interaction involving the two factors of main interest was an interaction between block and bilateral vs unilateral (F(1,11) = 4.11, p = 0.041), with a larger UFA at the beginning of the task which became shorter as time on task accrued (linear trend: F(1,11) = 6.39, p = 0.028). UFAs of 6, 8, 0 and 2 ms were obtained during the four blocks. This linear trend has previously been interpreted as an effect potentially attributable to callosal plasticity in the same dataset (Leblanc-Sirois and Braun 2015).
Mean commission and omission error rates were 26 and 2%, respectively. With commission error rate as the dependent variable, only the bilateral vs unilateral effect was significant (F(1,11) = 8.88, p = 0.013), revealing more errors for bilateral than unilateral presentations. With omission error rate as the dependent variable, the bilateral vs unilateral effect was again significant (F(1,11) = 8.69, p = 0.013), revealing more omission errors for bilateral over unilateral presentations. There was an effect of pair orientation (quadratic trend: F(1,11) = 15.22, p = 0.002). Fewer errors of omission were made following horizontal pair presentations than following oblique pair presentations. A significant bilateral vs unilateral * pair orientation interaction was also obtained (quadratic trend: F(1,11) = 12.20, p = 0.005). Decomposing this effect yielded no significant bilateral or unilateral field advantage for horizontal pair presentations, but yielded a significant UFA (F(1,11) = 11.53, p = 0.007) for oblique presentations. Mean RTs and error rates for bilateral and unilateral presentations, and for horizontal and oblique pair orientations of Experiment 1, are available in Table 1.
Poffenberger analyses
With RT of Experiment 1 as the dependent variable, there were no significant response hand (F(1,11) = 1.22, p = 0.29) or visual field (F(1,11) = 1.96, p = 0.19) effects. However, a significant response hand * visual field interaction was found (F(1,11) = 27.4, p < 0.0005). A CUD of 5 ms was recorded. No other interactions were significant.
For commission errors, no effects of response hand (F(1,11) = 0.66, p = 0.43) or visual field (F(1,11) = 2.19, p = 0.16) were found, but there was a significant interaction between the two (F(1,11) = 6.01, p = 0.032), yielding fewer commission errors for crossed presentations than for uncrossed presentations. The only other significant interaction involving a factor of primary interest was a significant block * visual field interaction (F(1,11) = 3.8, p = 0.039), with a higher proportion of commission errors occurring for right visual field stimuli as time on task accrued. No response hand or visual field effects or interactions were found in omission error rates.
The correlation between RT measures of CUD and UFA was calculated, and was found to be non-significant (r = − .023, p = 0.94). The correlation between RT CUD and the length of the UFA difference between horizontal and oblique presentations was also non-significant (r = − 0.153, p = 0.64). Mean RTs and error rates for crossed and uncrossed presentations in Experiment 1 are available in Table 1.
Experiment 2
In simple reaction time tasks, the complexity of the motor response has been shown to influence a measure of interhemispheric transfer time, the CUD (Braun et al. 2004; Iacoboni and Zaidel 1995). In particular, CUDs lengthened significantly when participants had to alternate between the left and right hands within a block, after each correct response. Experiment 2 was an attempt to investigate whether the effects of callosal homotopy remained significant when participants have to alternate between hands after each correct go trial. This second experiment was also made as similar as possible to Experiment 1 to allow a comparison between conditions with and without hand switch after each correct go trial.
Methods
Twelve right handed participants (6 male, 6 female) aged 18 to 30 were recruited among university students. Participants had no history of neurological or psychiatric problems, and were tested for adequate vision prior to the experiment.
The experimental procedure was similar to the task of Experiment 1, but response hand varied within trial runs rather than between trial runs. Participants were told to switch response hands after each correct response to a go trial. The left index finger on the “Z” key was used for left hand responses. The right index finger on the “2” key on the numeric pad was used for right hand responses. These two keys were chosen for their left and right position and for their equidistance to the edge of the keyboard. To facilitate responses, a keyboard on which the keys surrounding the “Z” and the “2” were absent was used during the experiment.
To help participants respond with the correct hand, the fixation cross on each trial had to be replaced by a bracket (“>” or “<”) pointing to the side of the response hand for the current trial. To draw attention to the change of hands, a fixation blink in between trials was created by decreasing the duration of the blank display with fixation (or error message during failed trials) from 500 to 400 ms, and by adding a blank display without fixation for 100 ms afterwards, separating the end of each trial from the beginning of the next trial.
Commission and omission errors were followed by feedback to the participant as in the previous task. “Incorrect hand” errors on go trials were also recorded and indicated to the participant by a “wrong hand” error message. As with omission errors, incorrect hand responses were not counted as correct, and trials yielding incorrect hand responses were replaced within each trial run. Statistical methods were identical to those of Experiment 1, with the addition of analyses of variance on incorrect hand error rates.
Results
Dimond analyses
Mean RT of Experiment 2 for all data included in the Dimond analysis was 429 ms. With RT data, there were no significant simple effects of bilateral vs unilateral (F(1,11) = 1.98, p = 0.187) and pair orientation (quadratic trend: F(1,11) = 0.002, p = 0.963, partial η2 = 0.435). However, there was a significant interaction between bilateral vs unilateral and pair orientation (quadratic trend: F(1,11) = 8.46, p = 0.014) in the expected direction: there was a non-significant 4 ms BFA for horizontal pair orientations (F = 1.29, p = .28) and a significant 4 ms UFA (F(1,11) = 11.34, p = 0.006) for oblique pair orientations.
Mean commission error rate was 28%. With commission errors, there was a significant effect of bilateral vs unilateral (F(1,11) = 5.46, p = 0.039), with higher rate of commission errors for bilateral than for unilateral trials. This result constituted a significant UFA obtained with commission errors. A trend towards a bilateral vs unilateral * pair orientation interaction was also detected (quadratic trend: F(1,11) = 4.78, p = 0.051) in a direction concordant with RT data, with a non-significant BFA for horizontal pair orientations and a non-significant UFA for oblique pair orientations. Mean omission error rate was 2.4%, with no significant effects or interactions involving bilateral vs unilateral or pair orientation. Mean “incorrect hand” error rate was 1.1%. With “incorrect hand” error data, there was a significant effect of bilateral vs unilateral (F(1,11) = 22.21, p = 0.001), showing more hand side errors for unilateral over bilateral presentations. However, there was no interaction between bilateral vs unilateral and pair orientation (quadratic trend: F(1,11) = 0.46, p = 0.51). Mean RTs and error rates for bilateral and unilateral presentations, and for horizontal and oblique pair orientations of Experiment 2, are available in Table 1.
Poffenberger analyses
With RT data of Experiment 2, there was a significant effect of response hand (F(1,11) = 5.97, p = 0.033), with shorter RT for the right over the left hand. A significant effect of visual field was also obtained (F(1,11) = 9.88, p = 0.009), with an advantage of the left visual field over the right visual field. There was a significant interaction between response hand and visual field (F(1,11) = 25.5, p < 0.0005), yielding a CUD of 25 ms.
With commission error data, there were no effects of response hand (F(1,11) = 2.17, p = 0.169) or visual field (F(1,11) = 3.4, p = 0.092), but an interaction between the two was found (F(1,11) = 5.95, p = 0.033), indicating more commission errors for uncrossed over crossed presentations. The direction of this inverse CUD was opposite to that of the CUD obtained in RT data. With omission error data, there were no effects of response hand (F(1,11) = 0.004, p = 0.95) or visual field (F(1,11) = 0.92, p = 0.359), but there was a significant interaction between the two (F(1,11) = 13.51, p = 0.004), indicating fewer omission errors for uncrossed over crossed presentations, an effect in the same direction as RT data. With incorrect hand error rates as the dependent variable, there were no effects of response hand (F(1,11) = 0.004, p = 0.95) or visual field (F(1,11) = 0.32, p = 0.585), but there was a significant interaction between the two (F(1,11) = 24.72, p < 0.0005), indicating fewer “incorrect hand” errors for uncrossed presentations than for crossed presentations, indicating better performance on uncrossed trials as with RT and omission errors.
The correlation between RT measures of CUD and UFA was calculated, and was found to be non-significant (r = 0.17, p = 0.60). The correlation between RT CUD and the length of the UFA difference between horizontal and oblique presentations was also non-significant (r = 0.22, p = 0.49). Mean RTs and error rates for crossed and uncrossed presentations in Experiment 2 are available in Table 1.
Both experiments combined
As previously stated, the principal purpose of the two experiments was to test the hypothesis that manipulating the visual characteristics of a Dimond task would modify the UFA or BFA without influencing Poffenberger’s CUD, while manipulating the motor contingencies would change Poffenberger’s CUD without altering the UFA or BFA. For this purpose, the same statistical analyses realized in the two experiments separately were run again on the data from the two experiments combined. A two level experiment between subjects factor, contrasting the first experiment during which response hand changed between trial runs and the second experiment during which response hand changed within trial runs, was added to the analyses of variance. Only effects of experiment and its interactions will be of interest for the next section of the present report.
Results
Dimond analyses
With RT as the dependent variable, the main effect of experiment on RT was non-significant (F(1,22) = 0.69, p = 0.42). There was no interaction between experiment and bilateral vs unilateral (F(1,22) = 0.71, p = 0.407) and no triple interaction between experiment, bilateral vs unilateral, and pair orientation (quadratic trend: F(1,22) < 0.01, p = 0.95). There were no other significant interactions with experiment in RT data. Experiment had no significant effect and did not significantly interact with other factors in commission error rates or omission error rates.
Poffenberger analyses
With RT as the dependent variable, there was no interaction between response hand and experiment (F(1,22) = 3.06, p = 0.09) and no interaction between visual field and experiment (F(1,22) = 0.76, p = 0.39). However, a significant triple interaction between response hand, visual field, and experiment was obtained (F(1,22) = 15.55, p = 0.001, partial η2 = 0.414), showing a larger CUD when response hand varied within blocks. No other significant interactions involving experiment were found in RT data.
With commission error rate, no significant interactions involving experiment were found with commission error rate as the dependent variable. However, with omission error rate as the dependent variable, there was a significant triple interaction between response hand, visual field, and experiment (F(1,22) = 10.05, p = 0.004). As with RT data, the CUD in omission errors was larger when response hand varied within blocks rather than between blocks. The partial correlation, controlling for experiment, between RT-CUD and RT-UFA remained non-significant (r = 0.97, p = 0.66).
General discussion
Unilateral field advantage influenced by stimulus orientation
In both experiments reported here, UFA was longer for oblique than for horizontal orientations. This effect of stimulus orientation on the UFA, a measure thought to reflect the latency of interhemispheric transfer of non-motor information, was interpreted as consequence of the homotopy of callosal fibers transporting visual information. The current experiments constitute two successful replications of this previously obtained effect (Desjardins et al. 2009; Braun et al. 2011, 2015) in similar tasks, and show that this effect resists manipulation of motor demands of the task.
Several researchers (Zaidel 1983; Brown et al. 1998) have previously argued that accuracy should also be lower when interhemispheric transfer of information is necessary, as a result of signal degradation attributable to the limited number of callosal connections and the longer distances between communicating neurons located in opposing hemispheres. Lower error rates in oblique rather than horizontal bilateral presentations relative to their respective unilateral control presentations had yet to be found. Both the significant homotopy effect obtained with omission errors in Experiment 1 and the trend towards a homotopy effect in Experiment 2 suggest a larger reduction in precision when bilaterally presented stimuli are not symmetric across the vertical meridian.
Spatial compatibility influences Poffenberger’s CUD in go/no-go tasks
Poffenberger’s CUD was also significant in both experiments, despite the relative complexity of the current task relative to simple reaction time or go/no-go tasks traditionally used to estimate interhemispheric transfer time. Its mean value fell very close to the usual 2–4 ms estimates of CUD obtained with simple reaction time and simple go/no-go tasks (Marzi et al. 1991; Braun 1992), so long as response hand was kept constant during each trial run. However, when response hand alternated within each trial run, a very large mean CUD of 25 ms was obtained. This CUD value falls outside the normal range for behavioral estimates of IHTT, and even falls outside the longer 10–20 ms range for estimates of VEP-IHTT (Saron and Davidson 1989).
Two explanations based on directed visual attention vs spatial compatibility were considered for these large CUDs in Experiment 2. An attentional explanation proposes that participants’ visual attention deviates towards the side of the visual field ipsilateral to the hand required for the next response, perhaps because of the hint towards the hand of response given at the site of the fixation (Hommel et al. 2001). The higher rate of commission and incorrect hand errors for uncrossed trials were consistent with this explanation. However, the attentional explanation also predicted that omission errors would be more frequent in uncrossed trials, but the opposite result was obtained. This result corresponds to a response bias towards the side of response, not to higher accuracy towards the side of response.
The spatial compatibility explanation instead proposes that participants more often evaluated trials as matches if the sides of the stimulus pairs and of the response were spatially concordant. The spatial compatibility explanation is consistent with the direction of CUDs obtained for RTs and all three error types of Experiment 2. The spatial compatibility effect occurred even though similarities between both stimuli, not spatial concordance, separated go from no-go trials. Spatial compatibility effects occurred even though no correlation between hand side and stimulus pair side existed.
The CUD has previously been shown to be in part determined by interhemispheric transfer time and in part by spatial compatibility effects (Braun et al. 2004) which are also callosally mediated (Di Stefano et al. 1992; Aglioti et al. 1996). Because the CUDs of Experiment 2 are extremely long, a very large part of the increase in CUD resulting from the more complex motor condition is likely attributable to spatial compatibility. Longer delays attributable to spatial incompatibility in Experiment 2 may result from the need to actively process stimulus location and response location on each trial, whereas in Experiment 1 response location remains fixed and does not need to be determined on every trial.
Dimond’s measure and Poffenberger’s CUD represent different types of interhemispheric transfer
The logic of Dimond’s inference regarding interhemispheric dynamics excludes premotor information. Accordingly, neither Dimond’s “measure” nor the homotopy effect were affected by changes in the task’s motor demands. Additionally, two arguments emerging from data collected during the two experiments strongly suggest that a transfer of heavily cognitively processed information can be excluded as well. Stated otherwise, the interhemispheric transfer indexed in the current perceptual matching task is almost certainly a transfer of visual information. First, a short UFA was found in Experiment 1, while a non-significant UFA was found in Experiment 2, with no significant difference between these two results (See Fig. 2). Secondly, short UFAs in Dimond tasks have previously been associated with transfer of visual information, while transfer of information that has been verbally encoded normally yield relatively large and significant BFAs (Banich and Belger 1990; Belger and Banich 1992), usually longer than 20 ms. A recent meta-analysis of Dimond tasks showed that to be the case in 23 out of 25 experiments (Leblanc-Sirois and Braun 2014). Moreover, the bilateral–unilateral differences were affected by a manipulation of stimulus pair orientation, in both experiments. Information pertaining to stimulus location obviously does not require verbal encoding. Indeed, such a robust UFA as obtained in both current experiments would be expected only if stimuli were not verbally encoded before information is transferred across the corpus callosum.

Manipulation of visual and motor conditions yields differences in Dimond’s and Poffenberger’s measures, respectively. a UFAs are larger for oblique stimulus pair presentations than for horizontal stimulus pair presentations, irrespective of motor demands. b CUDs are longer when participants must switch hands within trial runs than when response hand is fixed within a trial run, irrespective of stimulus pair orientation. Error bars represent standard error of the mean. *Statistically significant
Moreover, data collected from both experiments confirm that Poffenberger’s CUD was not tied to the transfer of visual information. In both experiments, a manipulation of the visual scene known to affect the latency of the transfer of visual information did not modify CUDs significantly (See Fig. 2). Data instead confirmed a link between Poffenberger’s CUD and the transfer of premotor information, because a manipulation of motor demands of the task yielded a change in the length of CUDs. However, it is probable that a large part of this CUD difference between both experiments can be accounted for by spatial compatibility. The current set of experiments did not precisely determine to which extent the CUD difference represented a change in IHTT.
A correlation between UFA and CUD measures obtained for each participant was also expected if the transfer of visual information accounted for both measures. The non-significance of this correlation in both experiments also provided an argument for the idea that different types of information transfer across hemispheres influence Dimond’s and Poffenberger’s measures.
In conclusion, results obtained in both experiments indicate that Dimond’s measure of interhemispheric dynamics and Poffenberger’s CUD do not index the same type of interhemispheric transfer of information. The balance between unilateral and bilateral presentations was found to be sensitive to a manipulation of stimulus pair orientation, but not to the addition of frequent changes in response hand. The difference between crossed and uncrossed trials was instead sensitive to a manipulation of motor demands, but was not sensitive to a change in stimulus pair orientation.
References
Aglioti S, Tassinari G, Berlucchi G (1996) Spatial stimulus – response compatibility in callosotomy patients and subjects with callosal agenesis. Neurosci Biobehav Rev 20:623–629. https://doi.org/10.1016/0149-7634(95)00075-5
Anzola GP, Bertoloni G, Buchtel HA, Rizzolatti G (1977) Spatial compatibility and anatomical factors in simple and choice reaction time. Neuropsychologia 15:295–302. https://doi.org/10.1016/0028-3932(77)90038-0
Banich MT, Belger A (1990) Interhemispheric interaction: how do hemispheres divide and conquer a task? Cortex 26:77–94. https://doi.org/10.1016/S0010-9452(13)80076-7
Belger A, Banich MT (1992) Interhemispheric interaction affected by computational complexity. Neuropsychologia 30:923–929. https://doi.org/10.1016/0028-3932(92)90036-L
Berlucchi G, Heron W, Hyman R, Rizzolatti G, Umiltà AG (1971) Simple reaction time of ipsilateral and contralateral hand to lateralized visual stimuli. Brain 94:419–430. https://doi.org/10.1093/brain/94.3.419
Berlucchi G, Crea F, Di Stefano M, Tassinari G (1977) Influence of spatial stimulus—response compatibility on reaction time of ipsilateral and contralateral hand to lateralized light stimuli. J Exp Psycho Hum Percept Perform 3:505–517. https://doi.org/10.1037/0096-1523.3.3.505
Bourne VJ (2006) The divided visual field paradigm: Methodological considerations. Laterality 11:373–393. https://doi.org/10.1080/13576500600633982
Braun CMJ (1992) Estimation of interhemispheric dynamics from simple unilateral reaction time to extrafoveal stimuli. Neuropsychol Rev 3:321–365. https://doi.org/10.1007/BF01108415
Braun CMJ, Collin I, Mailloux C (1997) The “Poffenberger” and “Dimond” paradigms: interrelated approaches to the study of interhemispheric dynamics? Brain Cogn 34:337–359. https://doi.org/10.1006/brcg.1997.0912
Braun CMJ, Larocque C, Achim A (2004) Experimental disentangling of spatial-compatibility and interhemispheric-relay effects in simple reaction time (Poffenberger paradigm). Exp Brain Res 157:442–456. https://doi.org/10.1007/s00221-004-1858-5
Braun CMJ, Achim A, Gauvin G, Desjardins S, Roberge C, Suffren S (2011) New variations of intrahemispheric and interhemispheric processing indexed by the Dimond paradigm. Am J Psychol 124:163–176. https://doi.org/10.5406/amerjpsyc.124.2.0163
Braun CMJ, Achim A, Roberge C, Gauvin G (2015) Callosal sensitivity to short-range stimulus orientation and long-range stimulus context orientation: tachistoscopic evidence. Am J Psychol 128:355–365. https://doi.org/10.5406/amerjpsyc.128.3.0355
Brown WS, Jeeves MA (1993) Bilateral visual field processing and evoked potential interhemispheric transmission time. Neuropsychologia 31:1267–1281. https://doi.org/10.1016/0028-3932(93)90097-J
Brown WS, Bjerke MD, Galbraith GC (1998) Interhemispheric transfer in normals and acallosals: latency adjusted evoked potential averaging. Cortex 34:677–692. https://doi.org/10.1016/S0010-9452(08)70772-X
Brown WS, Jeeves MA, Dietrich R, Burnison DS (1999) Bilateral field advantage and evoked potential interhemispheric transmission in commissurotomy and callosal agenesis. Neuropsychologia 37:1165–1180. https://doi.org/10.1016/S0028-3932(99)00011-1
Clarke JM, Zaidel E (1989) Simple reaction times to lateralized flashes: varieties of interhemispheric communication routes. Brain 112:849–870. https://doi.org/10.1093/brain/112.4.849
Desjardins S, Braun CMJ, Achim A, Roberge C (2009) A choice reaction time index of callosal anatomical homotopy. Brain Cogn 71:46–51. https://doi.org/10.1016/j.bandc.2009.04.002
Di Stefano M, Saurerwein HC, Lassonde M (1992) Influence of anatomical factors and spatial compatibility on the stimulus-response relationship in the absence of the corpus callosum. Neuropsychologia 30:177–185. https://doi.org/10.1016/0028-3932(92)90026-1
Dimond SJ (1969) Hemisphere function and immediate memory. Psychonomic Sci 16:111–112
Friedrich P, Ocklenburg S, Mochalski L, Schlüter C, Güntürkün O, Genc E (2017) Long-term reliability of the visual EEG Poffenberger paradigm. Behav Brain Res 330:85–91. https://doi.org/10.1016/j.bbr.2017.05.019
Hommel B, Pratt J, Colzato L, Godijn R (2001) Symbolic control of visual attention. Psychol Sci 12:360–365. https://doi.org/10.1111/1467-9280.00367
Iacoboni M, Zaidel E (1995) Channels of the corpus callosum: Evidence from simple reaction times to lateralized flashes in the normal and the split brain. Brain 118:779–788. https://doi.org/10.1093/brain/118.3.779
Larocque C, Achim A, Braun CMJ (2001) Refutation of Poffenberger’s inference: An existence proof and a countermodel. Brain Cogn 46:188–191. https://doi.org/10.1016/S0278-2626(01)80062-2
Larson EB, Brown WS (1997) Bilateral field interactions, hemispheric specialization and evoked potential interhemispheric transmission time. Neuropsychologia 35:573–581. https://doi.org/10.1016/S0028-3932(96)00099-1
Leblanc-Sirois Y, Braun CMJ (2014) Intra and inter hemispheric dynamics revealed by reaction time in the Dimond paradigm: a quantitative review of the literature. Neuropsychologia 58:1–13. https://doi.org/10.1016/j.neuropsychologia.2014.03.012
Leblanc-Sirois Y, Braun CMJ (2015) Inter-hemispheric dynamics revealed by reaction time in the Dimond paradigm. Q J Exp Psychol 68:165–180. https://doi.org/10.1080/17470218.2014.940984
Lines CR, Rugg MD, Milner AD (1984) The effects of stimulus intensity on visual evoked potential estimates of interhemispheric transmission time. Exp Brain Res 57:89–98. https://doi.org/10.1007/BF00231135
Marzi CA, Bisiacchi P, Nicoletti R (1991) Is interhemispheric transfer of visuomotor information asymmetric? Evidence from a meta-analysis. Neuropsychologia 29:1163–1177. https://doi.org/10.1016/0028-3932(91)90031-3
Milner AD, Lines CR (1982) Interhemispheric pathways in simple reaction time to lateralized light flash. Neuropsychologia 20:171–179. https://doi.org/10.1016/0028-3932(82)90007-0
Poffenberger AT (1912) Reaction time to retinal stimulation with special reference to the time lost in conduction through nerve centers. Arch Psychol 23:1–73
Rugg MD, Lines CR, Milner AD (1985) Further investigation of visual evoked potentials elicited by lateralized stimuli: Effects of stimulus eccentricity and reference site. EEG Clin Neurophysiol 62:81–87. https://doi.org/10.1016/0168-5597(85)90019-X
Saron CD, Davidson RJ (1989) Visual evoked potential measures of interhemispheric transfer time in humans. Behav Neurosci 103:1115–1138. https://doi.org/10.1037/0735-7044.103.5.1115
Savazzi S, Fabri M, Rubboli G, Paggi A, Tassinari CA, Marzi CA (2007) Interhemispheric transfer following callosotomy in humans: role of the superior colliculus. Neuropsychologia 45:2417–2427. https://doi.org/10.1016/j.neuropsychologia.2007.04.002
Thut G, Hauert C-A, Morand S, Seeck M, Landis T, Michel C (1999) Evidence for motor-level transfer in a simple reaction time task: an EEG study. Exp Brain Res 128:256–261. https://doi.org/10.1007/s002210050
Zaidel E (1983) Disconnection syndrome as a model for laterality effects in the normal brain. In: Hellige J (ed) Cerebral hemisphere asymmetry. A.R. Liss, New York, pp 95–151
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The author declares that there is no conflict of interest.
Ethical approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards. Experimental procedures for the first experiment were approved by the Institutional Review Board of the Université du Québec à Montréal Psychology Department. Experimental procedures for the second experiment were approved by the Institutional Review Board of the Université du Québec à Montréal Faculty of Human Sciences.
Informed consent
Informed consent was obtained from all individual participants included in the study.
Rights and permissions
About this article

Cite this article
Leblanc-Sirois, Y., Braun, C.M.J. & Elie-Fortier, J. Effects of stimulus pair orientation and hand switching on reaction time estimates of interhemispheric transfer. Exp Brain Res 236, 1593–1602 (2018). https://doi.org/10.1007/s00221-018-5243-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-018-5243-1