
Abstract
Individuals can perceive the properties of an attached or grasped object by wielding it through muscular effort—an ability referred to as dynamic or effortful touch. Sensitivity to the forces required to move such objects and to the resulting global patterns of tissue deformation underlies such perception. Given that perception via dynamic touch is movement-based, we hypothesized that manipulations that affect the ability to produce and control muscular movements might affect perception via dynamic touch. Cooling muscles from 40 to 10 °C impedes the development and transmission of muscular force and diminishes muscle stretch-reflex sensitivity. Accordingly, we anticipated that changes in hand temperature would alter the ability to detect patterns of tissue deformation and thus perception of the properties of wielded objects. In two experiments, participants wielded dowels with different lengths and rotational inertias (Experiment 1) and objects with identical lengths and different rotational inertias (Experiment 2). They reported perceived lengths of these objects, in the absence of vision, in cool (~ 10 °C), neutral (~ 30 °C), and warm temperature conditions (~ 40 °C). Actual length predicted perceived length of the dowels (Experiment 1), and rotational inertia predicted perceived length of the objects (Experiment 2); perceived lengths were longer in the warm condition than in the cool condition. In consideration of known temperature-induced changes in tissue structure and function, our results support the hypothesis that comparable processes underlie the control of movement and perception via dynamic touch.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Individuals can perceive the properties of objects and surfaces through mechanical contact by cutaneous touch, haptic touch, and dynamic or effortful touch. Cutaneous touch refers to the ability to perceive touch, pressure, vibration, temperature, or pain through passive skin contact (Mountcastle 2005; Jones and Lederman 2006). Haptic touch refers to the ability to perceive the properties of objects or surfaces through active exploration by fingers, especially enclosure (Mountcastle 2005; Jones and Lederman 2006). Dynamic or effortful touch refers to the ability to perceive properties of the body or objects attached to it by means of movement via muscular effort. Dynamic touch can be used to perceive the movement and disposition of the limbs (proprioception), the length, width, and shape of an attached or grasped object (exteroception), the orientation of such objects in the hand (exproprioception), and the location of the hand on such objects (proexteroception) (Carello and Turvey 2000; Turvey and Carello 2011). Consequently, dynamic touch is central to attaining or maintaining posture and to grasping, holding, moving, and manipulating objects and using tools.
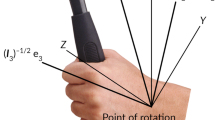
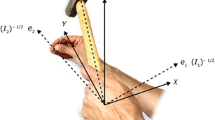
An invariant mechanical property, rotational inertia, I—the differential resistance of an object to angular acceleration in different directions—provides the informational support for dynamic touch [refer to Fitzpatrick et al. (1994) for a comprehensive account]. I can be represented in a 3 × 3 matrix—the “inertia tensor”—quantifying the differential resistance of an attached or grasped object to angular acceleration in different directions (Fig. 1). The eigenvectors e 1, e 2, and e 3 describe the symmetry axes of the mass distribution of the hand–object system and thus specify the orientation of (the mass distribution of) the hand–object system (Pagano and Turvey 1992; Turvey et al. 1992). The eigenvalues I 1, I 2, and I 3 describe the resistance to angular acceleration about the symmetry axes and thus specify the extent of such objects along each of those axes (Fitzpatrick et al. 1994; Turvey et al. 1998). The largest and the smallest eigenvalues, I 1 and I 3, specify the length and width, respectively, of such objects. Finally, the ratio of I 1 and I 3 specifies the shape of such objects (Burton et al. 1990).

Adapted from Mangalam et al. (2007). Copyright 2017 by the Springer US
Eigenvalues (I 1, I 2, and I 3) and eigenvectors (e 1, e 2, and e 3) of the inertia tensor of the hand–object system. The origin of the coordinate system is at a point in the wrist.
Whereas cutaneous and haptic touch seem to be based in the localized activity of mechanoreceptors, thermoreceptors, and nociceptors in the skin (Mountcastle 2005; Jones and Lederman 2006), dynamic touch seems to be based in more global patterns of tissue deformation (Turvey and Fonseca 2014). Movement of a limb and any object attached to it alters the tensile states of the muscles, tendons, and connective tissues of the hand, arm, shoulder, torso, and other body parts. This deformation, in turn, alters the patterning of afferent activity in a time-invariant manner.
According to a recent proposal (Turvey and Fonseca 2014), the anatomical basis of dynamic touch is best described as a multifractal tensegrity (MFT) system. The skin, connective tissue net, muscles, tendons, bones, joints, and nerve fibers together comprise a delicately balanced interconnected mechanical structure held together by the finely tuned interaction among elements under tension or compression. Localized mechanical forces bring a global realignment of compression and tension forces throughout the system, and therefore, local forces (e.g., at a particular anatomical site) are registered globally (i.e., at the level of the system as a whole). Accordingly, global patterns of tissue deformation, rather than the identity of the locally deformed tissues, underlie perception via dynamic touch. In other words, perception via dynamic touch is task-specific and anatomically independent. For example, individuals can perceive the length of an object by grasping it with minimal movement (Burton and Turvey 1990; Carello et al. 1992), they can perceive the whole or partial length of an object by grasping it at different positions along its length (Solomon et al. 1989; Pagano et al. 1994; Cooper et al. 2000), and they can perceive the length of an object by wielding it about their wrist, elbow, or shoulder (Pagano et al. 1993), and by wielding it with their limbs, torso, or head (Hajnal et al. 2007a, b; Palatinus et al. 2011; Wagman and Hajnal 2014a; Wagman et al. 2017). All these findings show that the patterns of stimulation relevant for dynamic touch occur not at the level of individual mechanoreceptors, but rather at the level of tissue deformations across the entire system.
Moreover, dynamic touch abilities are much less affected by aging (Carello et al. 2000, 2008) than cutaneous or haptic touch abilities (Kenshalo 1986; Stevens and Choo 1996), despite changes in the morphology and deterioration in the physiological function of somatosensory structures (myelinated sensory nerve fibers and receptors) underlying exploratory movements (Shaffer and Harrison 2007). Dynamic touch abilities are also less affected by neurological conditions such as peripheral neuropathies than either cutaneous or haptic touch. For example, a 40-year-old female with lesions in her dorsal column system could neither feel (cutaneous touch) nor identify an object (haptic touch), but could wield it and perceive its length with her affected hand (Carello et al. 2006). In this case, perceived length more closely reflected actual length with the unaffected arm compared to the affected arm, but the scaling between inertial properties and perceived length remained unchanged across the two arms. All these findings further indicate that the patterns of stimulation relevant for dynamic touch do not occur at the level of individual mechanoreceptors, but rather at the level of tissue deformations across the entire system.
Given that perception via dynamic touch is movement-based, we hypothesized that manipulations that affect the ability to produce and control muscular movements might affect perception via dynamic touch. Cooling muscles from 40 to 10 °C impedes the development and transmission of muscular force (Clarke et al. 1958; Ranatunga et al. 1987; Cornwall 1994; de Ruiter et al. 1999) and diminishes muscle stretch-reflex sensitivity (Ottoson 1965; Michalski and Séguin 1975). Accordingly, we hypothesized that changes in hand temperature while wielding an object would affect perception of properties of that object via dynamic touch.
In two experiments, participants wielded dowels with different lengths and rotational inertias (Experiment 1) and objects with identical lengths and different rotational inertias (Experiment 2). They reported perceived lengths, in the absence of vision, when wielding those objects immersed in cool (~ 10 °C), neutral (~ 30 °C), and warm (~ 40 °C) water. Perception of length requires the registration of muscular forces for rotation about a joint while wielding. Given the nature of informational support for dynamic touch and the nature of the medium for dynamic touch, we expected that the information for perception of length would remain unchanged across changes in temperature but that the overall attunement of such information may be diminished in colder water compared to warmer water. Therefore, we expected that perceived lengths of wielded dowels/objects would be shorter at lower hand temperatures than at higher hand temperatures but that the scaling relationship between actual length and perceived length (Experiment 1), and between rotational inertia and perceived length (Experiment 2), would remain constant across changes in hand temperature.
Experiment 1
We examined the influence of hand temperature on the perception of the lengths of objects via dynamic touch. Participants submerged their hand in water at three different temperatures: ~ 10 °C (cool condition), ~ 30 °C (neutral condition), and ~ 40 °C (warm condition) and reported perceived lengths of dowels of different lengths and rotational inertias. We tested two specific hypotheses. First, given that detection of an invariant mechanical property underlies perception via dynamic touch (Carello and Turvey 2000; Turvey and Carello 2011), we expected that the scaling relationship between actual and perceived lengths would remain constant across changes in hand temperature. Despite this constant scaling relation, given how perception of the length of a wielded object via dynamic touch is affected by peripheral neuropathies (Carello et al. 2006), we expected that perceived length would generally be shorter in cooler than in warmer conditions. That is, both actual length and temperature would influence perceived length, and actual length would influence perceived length in the same manner across temperature conditions.
Methods
Participants
Eight adult men and four adult women (mean ± SD age = 28.6 ± 4.9 years; range 21–38 years; 11 right-handed and 1 left-handed) participated in the present study. Each participant signed a consent form with information regarding the purposes of the study, the procedures, and the potential risks and benefits of participation. The Institutional Review Board (IRB) at the University of Georgia (Athens, GA, USA) approved the present study.
Stimuli rods
The participants wielded seven maple wood dowels (lengths: 30.0, 35.0, 40.0, 45.0, 50.0, 55.0, and 60.0 cm, diameter = 1.2 cm). We coated each dowel with a thin layer of epoxy to prevent water absorption.
Experimental setup
We filled a 84 × 51 × 34 cm PVC container with water at the temperature of approximately 10 °C (cool condition), 27 °C (neutral condition), or 40 °C (warm condition) to a height of 50 cm (Fig. 2). Fluctuation in water temperature during testing in each temperature condition remained under ± 1.5°. We cut a circular hole (diameter = 12 cm) at each end of the lid so that both the experimenter, M. M., and the participant could insert their hand into the container. A black curtain blocked the container from the participant’s view. A 30-cm vertical slit in the curtain allowed the participants to access the container. The participant could report perceived length of the dowel on a meter scale vaulted at his/her shoulder height. The measurements on the meter scale were facing the other side and thus out of view of the participant.

Adapted from Mangalam et al. (2007). Copyright 2017 by the Springer US
Experimental setup from the top view (although not shown here, the participant’s forearm and hand, and the wielded dowel/object were completely immersed in the water and visible through the transparent lid).
Experimental procedure
We tested each participant individually in a 60- to 75-min session during which we conducted 21 trials (7 dowels × 3 trials/dowel) in each of the cool, neutral, and warm conditions. We randomized the order of temperature conditions across participants. We blocked trials in each temperature condition, but randomized the sequence of trials in each of the three temperature conditions for each of the 12 participants. We attached using surgical tape the thermal sensor of a stress thermometer (Bio-Medical Instruments, Inc., model SC911) on the back of the participant’s dominant hand. The participant submerged his/her hand in the water. In pilot trials, the skin temperature reached the water temperature in 3–4 min and thus we began the trials after 5 min. On each trial, the participant grasped the dowel at approximately 5 cm from the proximal end. The participant wielded the dowel without hitting the walls of the container and reported perceived length on the meter scale. M.M. recorded the measurement on the meter scale, the participant handed the dowel back to M.M., and a new trial began. M.M. recorded the water and skin temperatures at the beginning and after every three trials. After having completed all 21 trials for one temperature condition, the participant took a 10-min break. During the break, M.M. emptied the container and filled it with water per the subsequent temperature condition. After the break, the participant immersed his/her hand in the water at the current temperature condition for 5 min, and the trials began.
Statistical analysis
The values of r 2 lie between 0–1 and are not normally distributed. We thus transformed the values of r into Fischer’s z′. We performed all statistical analyses using two-tailed tests in SPSS 21 (IBM, Inc.) and considered outcomes statistically significant at the alpha level of 0.05.
Results and discussion
We conducted a 3 (temperature condition) × 7 (actual length) analysis of variance (ANOVA) on perceived length with within-subject factors of temperature condition and actual length. There was a main effect of temperature condition, F 2,22 = 4.796, p = 0.019, η 2 = 0.304. Follow-up t-tests with Bonferroni corrections revealed that perceived lengths were longer in the warm (40 °C) condition (mean ± SD = 35.8 ± 9.5 cm) than in the cool (10 °C) condition (32.4 ± 8.4 cm; p = 0.027). There was also a main effect of actual length, F 6,66 = 124.467, p < 0.001, η 2 = 0.919. In general, perceived length increased with actual length (all ps < 0.05). The interaction effect of temperature condition × actual length was not significant, F 12,132 = 1.505, p = 0.130, η 2 = 0.120.
A multiple regression revealed that actual length (in cm) and temperature (in °C) accounted for 94% of the variance in perceived length (p < 0.001), with actual length accounting for a much larger portion of the variance (β = 0.96, p < 0.001) and temperature accounting for a much smaller but still significant portion of the variance [β = 0.15, p < 0.05; perceived length = (0.71 × actual length) + (0.09 × temperature) + 0.618].
Figure 3 depicts the linear relationships between actual and perceived lengths of the seven dowels in each of the three temperature conditions for each of the 12 participants. In general, perceived lengths of the dowels did not exceed their actual lengths (Table 1), but the participants showed inter-individual differences. Perceived length increased linearly with actual length for each participant in each temperature condition (range of values of r 2 in Fig. 3: cool condition: r 2 = 0.689–0.989; neutral condition: r 2 = 0.815–0.990; warm condition: r 2 = 0.492–0.992). A one-way ANOVA with within-subject factors on the values of Fischer’s z′ (obtained by transforming the values of r) revealed no differences across the cool (mean ± SD = 2.020 ± 0.343), neutral (2.255 ± 0.416), and warm (2.058 ± 0.596) conditions, F 2,22 = 0.053, p = 0.948, η 2 = 0.005. A one-way ANOVA with within-subject factors on the values of the slopes also revealed no differences across the cool (mean ± SD = 0.697 ± 0.697), neutral (0.755 ± 0.191), and warm (0.783 ± 0.246) conditions, F 2,22 = 1.664, p = 0.212, η 2 = 0.131. Finally, a one-way ANOVA with within-subject factors on the values of the intercepts also revealed no differences across the cool (mean ± SD = 1.047 ± 7.684), neutral (0.752 ± 6.828), and warm (– 0.397 ± 9.431) conditions, F 2,22 = 0.547, p = 0.586, η 2 = 0.047.

Linear relationship between actual and perceived lengths of the dowels (n = 7) for each of the three temperature conditions for each of the 12 participants in Experiment 1
At the level of the mean data, simple linear regressions examining the relationships between actual and perceived lengths of the seven dowels across all 12 participants confirmed significant linear relationships in each temperature condition: (1) cool condition: perceived length = 0.696 × actual length + 1.093, in centimeters, F 1,5 = 545.216, p < 0.001, with an r 2 of 0.989; (2) neutral condition: perceived length = 0.754 × actual length + 0.818, in centimeters, F 1,5 = 688.838, p < 0.001, with an r 2 of 0.991; and (3) warm condition: perceived length = 0.782 × actual length + 0.618, in centimeters, F 1,5 = 868.481, p < 0.001, with an r 2 of 0.993 (Fig. 4; Table 1).

Linear relationship between actual and perceived lengths of the dowels (n = 7) for each of the three temperature conditions across all 12 participants in Experiment 1
Overall, both actual length and hand temperature influenced the perception of the lengths of the dowels. Perceived lengths were longer (and more closely reflected actual lengths) in the warm (40 °C) condition than in the cold (10 °C) condition. However, the scaling relationship between actual and perceived lengths remained constant across temperature conditions. This finding supports the hypothesis that changes in hand temperature do not change the invariant patterns of tissue deformation that support perception of the length of a wielded object via dynamic touch but may subtly change the ability to register (i.e., the attunement to) those patterns.
Experiment 2
In Experiment 1, changes in hand temperature influenced the perception of the length of a wielded object but did not affect the scaling of perceived length to actual length. However, Experiment 1 did not explicitly examine the influence of hand temperature on detection of or attunement to the invariant—rotational inertia, I—that supports perception via dynamic touch. To address this question, one needs to dissociate rotational inertia from actual length. We did this by attaching stacked steel rings to dowels of identical length and diameter, and of distinct material compositions. The rotational inertias of these “composite” objects were independent of their lengths. As in Experiment 1, we expected that perceived lengths would be shorter in colder water (i.e., at lower hand temperatures) than in warmer water (i.e., at warmer hand temperatures). However, we also expected that the scaling relationship between I 1 and perceived length would remain constant across changes in hand temperature.
Methods
Participants
Seven adult men and five adult women (mean ± SD age = 26.8 ± 3.8 years; range 21–29 years; 11 right-handed and 1 left-handed) participated in the present study. Each participant signed a consent form with information regarding the purposes of the study, the procedures, and the potential risks and benefits of participation. The Institutional Review Board (IRB) at the University of Georgia (Athens, GA, USA) approved the present study.
Stimuli objects
The participants wielded 12 objects with distinct values of I 1 (Fig. 5; Table 2). Each object consisted of a dowel (length = 50.0 cm, diameter = 1.2 cm) of distinct material composition (mass = pine wood: 27 g; maple wood: 45 g; hollow aluminum: 75 g; solid aluminum: 171 g) with a particular number of stacked steel rings (inner diameter = 1.4 cm, outer diameter = 3.4 cm; height = 0.2 cm, mass = 14 g) attached to it along its length. We attached two rings to the pine wood dowel (height = 0.4 cm, mass = 28 g), three rings to the maple wood dowel (height = 0.6 cm, mass = 42 g), four rings to the hollow aluminum dowel (height = 0.8 cm, mass = 56 g), and five rings to the solid aluminum dowel (height = 1.0 cm, mass = 70 g) at 25.0, 35.0, or 45.0 cm along the length of each dowel. To prevent water absorption, we coated the pine and maple wood dowels and sealed both openings of the hollow aluminum dowel with epoxy. Further, to prevent perception of the composition of a dowel via cutaneous touch, we put a weightless 15.0 cm rubber grip (inner diameter = 2.0 cm, outer diameter = 3.4 cm) of negligible mass on each dowel.

Stimuli objects (see text for details)
Experimental setup
The experimental setup was same as in Experiment 1.
Experimental procedure
We tested each participant individually in a 75- to 90-min session during which we conducted 36 trials (12 objects × 3 trials/object) in each of the cool, neutral, and warm conditions. We randomized the order of temperature conditions across participants. We blocked trials in each temperature condition, but randomized the sequence of trials in each of the three temperature conditions for each of the 12 participants.
The rest of the experimental procedure was same as in Experiment 1.
Statistical analysis
The values of I 1 were highly skewed and, therefore, we log transformed the values of I 1. Accordingly, we also log transformed the values of perceived length. The values of r 2 lie between 0 and 1 and are not normally distributed. We thus transformed the values of r into Fisher’s z′. We performed all statistical analyses using two-tailed tests in SPSS 21 and considered outcomes statistically significant at the alpha level of 0.05.
Results and discussion
We conducted a 3 (temperature condition) × 7 (logI 1) ANOVA on log(perceived length) (logL perceived) with within-subject factors of temperature condition and logI 1. There was a main effect of temperature condition, F 2,22 = 6.271, p = 0.007, η 2 = 0.363. Follow-up t-tests with Bonferroni corrections revealed that perceived lengths were longer in the warm (40 °C) condition (mean ± SD = 45.0 ± 12.2 cm) than in the cool (10 °C) condition (41.0 ± 12.3 cm; p = 0.005). There was also a main effect of logI 1, F 11,121 = 22.097, p < 0.001, η 2 = 0.668. In general, perceived length increased with logI 1 (all ps < 0.05). The interaction effect of temperature condition × logI 1 was not significant, F 22,242 = 1.419, p = 0.105, η 2 = 0.114.
A multiple regression revealed that logI 1 (in g·cm2) and temperature (in °C) accounted for 88% of the variance in perceived length (p < 0.001), with logI 1 accounting for a much larger portion of the variance (β = 0.90, p < 0.001) and temperature accounting for a smaller but still significant portion of the variance [β = 0.27, p < 0.001; perceived length = (21.2 × logI 1) + (0.13 × temperature) − 65.03].
Figure 6 depicts the linear relationships between logI 1 and perceived lengths of the objects (n = 12) in each of the three temperature conditions for each of the 12 participants. The participants showed inter-individual differences. Perceived lengths increased linearly with logI 1 for all participants (except participant 2 in Fig. 5) in each temperature condition (range of values of r 2 in Fig. 5: cool condition: r 2 = 0.004–0.946; neutral condition: r 2 = 0.085–0.919; warm condition: r 2 = 0.068–0.907). A one-way ANOVA with within-subject factors on the values of Fisher’s z′ (obtained by transforming the values of r) revealed that the values differed across the three temperature conditions (cool: mean ± SD = 0.995 ± 0.542, neutral: 1.268 ± 0.544, and warm: 0.998 ± 0.454), F 2,22 = 3.613, p = 0.044, η 2 = 0.247. However, follow-up t-tests with Bonferroni corrections failed to find specific differences between any two conditions. A one-way ANOVA with within-subject factors on the values of the slopes also revealed no differences across the cool (mean ± SD = 0.182 ± 0.0.5), neutral (0.179 ± 0.124), and warm (0.153 ± 0.100) conditions, F 2,22 = 1.047, p = 0.368, η 2 = 0.087. Finally, a one-way ANOVA with within-subject factors on the values of the intercepts also revealed no differences across the cool (mean ± SD = 0.694 ± 0.468), neutral (0.571 ± 0.653), and warm (0.769 ± 0.502) conditions, F 2,22 = 2.473, p = 0.107, η 2 = 0.184.

Linear relationship between logI 1 and logL perceived (i.e., log[perceived length]) of the objects (n = 12) for each of the three temperature conditions for each of the 12 participants in Experiment 2
At the level of the mean data, simple linear regressions examining the relationship between logI 1 and logL perceived of the 12 across all 12 participants confirmed significant linear relationships in each of the three temperature conditions: (1) cool condition: logL perceived = (0.210 × logI 1) + 0.572, F 1,10 = 47.485, p < 0.001, with an r 2 of 0.809; (2) neutral condition: logL perceived = (0.229 × logI 1) + 0.497, F 1,10 = 146.952, p < 0.001, with an r 2 of 0.930; (3) warm condition logL perceived = (0.192 × logI 1) + 0.700, F 1,10 = 73.371, p < 0.001, with an r 2 of 0.868 (Fig. 7; Table 3). This implies that perceived length of an object was equal to (1) cool condition: 3.733 × I 1 0.210 cm; (2) neutral condition: 3.141 × I 1 0.229 cm; (3) warm condition: 5.012 × I 1 0.192 cm. These scaling relationships are comparable to those obtained in a previous experiment in which participants reported perceived lengths of objects wielded in water (Mangalam et al. 2017).

Linear relationship between logI 1 and logL perceived (i.e., log[perceived length]) of the objects (n = 12) for each of the three temperature conditions across all 12 participants in Experiment 2
Overall, both logI 1 and hand temperature influenced the perception of the lengths of the objects. Perceived lengths were longer in the warm (40 °C) condition than in the cold (10 °C) condition. The scaling relationship between logI 1 and log(perceived length) remained constant across the three temperature conditions. This finding supports the hypothesis that changes in hand temperature do not change the invariant patterns of tissue deformation that support perception of the length of a wielded object via dynamic touch, but may subtly change the ability to register (i.e., the attunement to) those forces.
General discussion
The present study aimed to identify whether and how temperature influences perception of the length of a wielded object via dynamic touch. We reasoned that since perception via dynamic touch requires the production of muscular forces, manipulations that affect the ability to produce and control muscular movements might affect perception via dynamic touch. Specifically, given that changes in tissue temperature influence the transmission of muscular forces, we hypothesized that changes in hand temperature might alter the ability to detect patterns of tissue deformation and thus affect the perception of length via dynamic touch. We expected that perceived lengths would be shorter in colder water (i.e., at lower hand temperatures) than in warmer water (i.e., at warmer hand temperatures). However, we also expected that the scaling relationship between I 1 and perceived length would remain constant across changes in hand temperature. In two experiments, participants wielded dowels with different lengths and rotational inertias (Experiment 1) and objects with different rotational inertias and identical lengths (Experiment 2). The participants reported perceived lengths of these objects, in the absence of vision, at three different hand temperatures (about 10, 30, and 40 °C). In general, the results supported our hypothesis (see Figs. 3, 4, 6, 7).
The present findings are consistent with the hypothesis that the medium for dynamic touch is the muscular, connective tissue, and skeletal system organized as a multifractal tensegrity (MFT) system (Turvey and Fonseca 2014). In such a system, local forces (at a particular anatomical site) are registered globally (at the level of the whole system). Accordingly, global patterns of tissue deformation, rather than the identity of the locally deformed tissues, underlie perception via dynamic touch. Such a view of perception via dynamic touch explains how it is that the particular limb that wields an object is often ‘functionally transparent’ to the perceiver (Hajnal et al. 2007a, b; Wagman and Hajnal 2014a). The present findings indicate that such a view might also explain how the properties of the medium in which an object is wielded is also functionally transparent to the perceiver (Pagano and Donahue 1999; Pagano and Cabe 2003; Mangalam et al. 2017). In perceiving the length (or any other property) of a wielded object via dynamic touch, the differential tissue deformation brought about by manipulating that object is influential, whereas the uniform deformation brought about by the medium in which such manipulation occurs is less so.
The present findings have implications for applied ergonomics, particularly concerning the risk of injury while using handheld (power) tools at colder temperatures. Labor under colder temperatures increases the risk of musculoskeletal injury caused by mechanical stress and strain due to diminished grip strength, cutaneous and haptic sensitivity, and manual dexterity, among other components of human manual performance (reviewed in Parsons 2002). The present findings highlight that changes in perception of the length of a grasped tool with decrease in temperature may be an additional risk. In particular, changes in perception of the dimensions or disposition of a handheld power tool, or the disposition of the hand on the handheld power tool, at colder temperatures, might significantly contribute to risks of injury. Thus, it may be beneficial to incorporate the effects of ambient temperature on perception of the length of wielded objects into the design of power tools, especially those explicitly meant for use at colder temperatures.
Electromyographic (EMG) recordings have revealed that flexor muscles of the forearm play a particularly important role in the perceived heaviness of a handheld object and the activity of these muscles is associated with the invariant structure of the inertia tensor at the wrist (Waddell and Amazeen 2017). The long tendons of the flexor muscles are the prime movers of the wrist and descend to the wrist and into the hand (Jarmey and Sharkey 2003). Moreover, the tendons and the connective tissue (fascia) interconnect with superficial muscles (Schleip et al. 2012), and the primary locations of muscle spindles and Golgi tendon organs are the transitional zones between collagenous connective tissue and muscle (van der Wal and Drukker 1988). In both experiments, the participants’ hand and forearm, particularly the distal part, were completely immersed in water. We thus believe that changes in the temperature of muscle belly and distal tendons of the flexor muscles of the forearm, which contain muscle spindles and Golgi tendon organs, respectively, are responsible for the observed temperature-induced changes in perception of length via dynamic touch.
The present findings are also consistent with J.J. Gibson’s ecological approach to perception–action (Gibson 1966, 1979). According to the ecological approach, perception of a given environmental property is supported by the detection of an invariant stimulation pattern that is informative about that property. Objects and events lawfully structure patterned energy distributions (e.g., the optic array or the acoustic array) such that this structure is specific to its source. This lawful structuring of energy distributions entails that the informative stimulation pattern is invariant across both particular instances of perceiving and the particular anatomical components by which the stimulation pattern is detected. In other words, such invariance putatively underlies the ability to perceive a given property with different configurations of the same perceptual modality and by means of entirely different perceptual modalities (Palatinus et al. 2011; Wagman and Abney 2012; Wagman and Hajnal 2014a, b; Wagman et al. 2017). That is, to a large extent, perception of a given environmental property is independent of both anatomy and modality. Together with previous research (Pagano and Donahue 1999; Pagano and Cabe 2003; Mangalam et al. 2017), the results of the experiments reported here suggest that perception of the length of a wielded object is also independent of the properties of the medium.
Dexterous coordination of the multi-segmented body is founded upon the formation and regulation of perceptuomotor synergies, a task accomplished through dynamic touch (Bernstein 1967; Bernstein et al. 1996). However, our understanding of the physiology of dynamic touch is not sufficient considering its fundamental role in the control of movement. Several questions remain unanswered in this context. What is the structural architecture of deformable tissues? Whether and how do certain features of the structural architecture of deformable tissues allow specific patterns of tissue deformation? What is the relationship between the global patterns of tissue deformation and the patterns of stimulation in the mechanoreceptors infused in the skin, connective tissue (fascia), and muscles? Given the distributed nature of this haptic subsystem, the depth of understanding might depend on the degree to which we meet the challenge of developing theoretical frameworks and experimental paradigms to study dynamic touch, perhaps in certain clinical populations (Carello et al. 2008).
References
Bernstein NA (1967) The co-ordination and regulation of movement. Pergamon, Oxford
Bernstein NA, Latash ML, Turvey MT (1996) Dexterity and its development. Lawrence Erlbaum, Marwah
Burton G, Turvey MT (1990) Perceiving the lengths of rods that are held but not wielded. Ecol Psychol 2:295–324. https://doi.org/10.1207/s15326969eco0204_1
Burton G, Turvey MT, Solomon HY (1990) Can shape be perceived by dynamic touch? Percept Psychophys 48:477–487. https://doi.org/10.3758/BF03211592
Carello C, Turvey MT (2000) Rotational invariance and dynamic touch. In: Heller MA (ed) Touch, representation and blindness. Oxford University Press, New York, pp 27–66
Carello C, Fitzpatrick P, Domaniewicz I et al (1992) Effortful touch with minimal movement. J Exp Psychol Hum Percept Perform 18:290–302. https://doi.org/10.1037/0096-1523.18.1.290
Carello C, Thuot S, Turvey MT (2000) Aging and the perception of a racket’s sweet spot. Hum Mov Sci 19:1–20. https://doi.org/10.1016/S0167-9457(99)00044-5
Carello C, Kinsella-Shaw J, Amazeen EL, Turvey MT (2006) Peripheral neuropathy and object length perception by effortful (dynamic) touch: a case study. Neurosci Lett 405:159–163. https://doi.org/10.1016/j.neulet.2006.06.047
Carello C, Silva PL, Kinsella-Shaw JM, Turvey MT (2008) Muscle-based perception: theory, research and implications for rehabilitation. Braz J Phys Ther 12:339–350. https://doi.org/10.1590/S1413-35552008000500002
Clarke RSJ, Hellon RF, Lind AR (1958) The duration of sustained contractions of the human forearm at different muscle temperatures. J Physiol 143:454–473. https://doi.org/10.1113/jphysiol.1958.sp006071
Cooper MM, Carello C, Turvey MT (2000) Perceptual independence of whole length, partial length, and hand position in wielding a rod. J Exp Psychol Hum Percept Perform 26:74–85. https://doi.org/10.1037/0096-1523.26.1.74
Cornwall MW (1994) Effect of temperature on muscle force and rate of muscle force production in men and women. J Orthop Sport Phys Ther 20:74–80. https://doi.org/10.2519/jospt.1994.20.2.74
de Ruiter CJ, Jones DA, Sargeant AJ, de Haan A (1999) Temperature effect on the rates of isometric force development and relaxation in the fresh and fatigued human adductor pollicis muscle. Exp Physiol 84:1137–1150
Fitzpatrick P, Carello C, Turvey MT (1994) Eigenvalues of the inertia tensor and exteroception by the “muscular sense”. Neuroscience 60:551–568. https://doi.org/10.1016/0306-4522(94)90264-X
Gibson JJ (1966) The senses considered as perceptual systems. Houghton Mifflin, Boston
Gibson JJ (1979) The ecological approach to visual perception. Houghton Mifflin, Boston
Hajnal A, Fonseca S, Harrison S et al (2007a) Comparison of dynamic (effortful) touch by hand and foot. J Mot Behav 39:82–88. https://doi.org/10.3200/JMBR.39.2.82-88
Hajnal A, Fonseca S, Kinsella-Shaw JM et al (2007b) Haptic selective attention by foot and by hand. Neurosci Lett 419:5–9. https://doi.org/10.1016/j.neulet.2007.03.042
Jarmey C, Sharkey J (2003) The concise book of muscles. North Atlantic Books, Berkeley
Jones LA, Lederman SJ (2006) Human hand function. Oxford University Press, New York
Kenshalo DRS (1986) Somesthetic sensitivity in young and elderly humans. J Gerontol 41:732–742. https://doi.org/10.1093/geronj/41.6.732
Mangalam M, Barton SA, Wagman JB et al (2017) Perception of the length of an object through dynamic touch is invariant across changes in the medium. Atten Percept Psychophys 79:2499–2509. https://doi.org/10.3758/s13414-017-1403-9
Michalski WJ, Séguin JJ (1975) The effects of muscle cooling and stretch on muscle spindle secondary endings in the cat. J Physiol 253:341–356. https://doi.org/10.1113/jphysiol.1975.sp011193
Mountcastle VB (2005) The sensory hand. Harvard University Press, Cambridge
Ottoson D (1965) The effects of temperature on the isolated muscle spindle. J Physiol 180:636–648. https://doi.org/10.1113/jphysiol.1965.sp007721
Pagano CC, Cabe PA (2003) Constancy in dynamic touch: length perceived by dynamic touch is invariant over changes in media. Ecol Psychol 15:1–17. https://doi.org/10.1207/S15326969ECO1501_01
Pagano CC, Donahue KG (1999) Perceiving the lengths of rods wielded in different media. Percept Psychophys 61:1336–1344. https://doi.org/10.3758/BF03206184
Pagano CC, Turvey MT (1992) Eigenvectors of the inertia tensor and perceiving the orientation of a hand-held object by dynamic touch. Percept Psychophys 52:617–624. https://doi.org/10.3758/BF03211699
Pagano CC, Fitzpatrick P, Turvey MT (1993) Tensorial basis to the constancy of perceived object extent over variations of dynamic touch. Percept Psychophys 54:43–54. https://doi.org/10.3758/BF03206936
Pagano CC, Kinsella-Shaw JM, Cassidy PE, Turvey MT (1994) Role of the inertia tensor in haptically perceiving where an object is grasped. J Exp Psychol Hum Percept Perform 20:276–285. https://doi.org/10.1037/0096-1523.20.2.276
Palatinus Z, Carello C, Turvey MT (2011) Principles of part-whole selective perception by dynamic touch extend to the torso. J Mot Behav 43:87–93. https://doi.org/10.1080/00222895.2010.538767
Parsons K (2002) Human thermal environments. CRC, Boca Raton
Ranatunga KW, Sharpe B, Turnbull B (1987) Contractions of a human skeletal muscle at different temperatures. J Physiol 390:383–395. https://doi.org/10.1113/jphysiol.1987.sp016707
Schleip R, Findley T, Huijing P (2012) Fascia: the tensional network of the human body. Elsevier, New York
Shaffer SW, Harrison AL (2007) Aging of the somatosensory system: a translational perspective. Phys Ther 87:193–207
Solomon HY, Turvey MT, Burton G (1989) Perceiving extents of rods by wielding: haptic diagonalization and decomposition of the inertia tensor. J Exp Psychol Hum Percept Perform 15:58–68. https://doi.org/10.1037/0096-1523.15.1.58
Stevens JC, Choo KK (1996) Spatial acuity of the body surface over the life span. Somatosens Mot Res 13:153–166. https://doi.org/10.3109/08990229609051403
Turvey MT, Carello C (2011) Obtaining information by dynamic (effortful) touching. Philos Trans R Soc London B Biol Sci 366:3123–3132. https://doi.org/10.1098/rstb.2011.0159
Turvey MT, Fonseca ST (2014) The medium of haptic perception: a tensegrity hypothesis. J Mot Behav 46:143–187. https://doi.org/10.1080/00222895.2013.798252
Turvey MT, Burton G, Pagano CC et al (1992) Role of the inertia tensor in perceiving object orientation by dynamic touch. J Exp Psychol Hum Percept Perform 18:714–727. https://doi.org/10.1037/0096-1523.18.3.714
Turvey MT, Burton G, Amazeen EL et al (1998) Perceiving the width and height of a hand-held object by dynamic touch. J Exp Psychol Hum Percept Perform 24:35–48. https://doi.org/10.1037/0096-1523.24.1.35
van der Wal JC, Drukker J (1988) The occurrence of muscle spindles in relation to the architecture of the connective tissues in the lateral cubical region of the rat. In: Hník P, Soukup T, Vejsada R, Zelená J (eds) Mechanoreceptors: development, structure, and function. Springer, Boston, pp 345–346
Waddell ML, Amazeen EL (2017) Evaluating the contributions of muscle activity and joint kinematics to weight perception across multiple joints. Exp Brain Res 235:2437–2448. https://doi.org/10.1007/s00221-017-4979-3
Wagman JB, Abney DH (2012) Transfer of recalibration from audition to touch: modality independence as a special case of anatomical independence. J Exp Psychol Hum Percept Perform 38:589–602
Wagman JB, Hajnal A (2014a) Task specificity and anatomical independence in perception of properties by means of a wielded object. J Exp Psychol Hum Percept Perform 40:2372–2391. https://doi.org/10.1037/xhp0000014
Wagman JB, Hajnal A (2014b) Getting off on the right (or left) foot: perceiving by means of a rod attached to the preferred or non-preferred foot. Exp Brain Res 232:3591–3599. https://doi.org/10.1007/s00221-014-4047-1
Wagman JB, Langley MD, Higuchi T (2017) Turning perception on its head: cephalic perception of whole and partial length of a wielded object. Exp Brain Res 235:153–167. https://doi.org/10.1007/s00221-016-4778-2
Author information
Authors and Affiliations
Contributions
MM and KMN conceived and designed research; MM performed experiments; MM analyzed data; MM and JBW interpreted results of experiments; MM prepared figures; MM, JBW, and KMN edited and revised manuscript; MM, JBW, and KMN approved final version of manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that no competing interests exist.
Rights and permissions
About this article

Cite this article
Mangalam, M., Wagman, J.B. & Newell, K.M. Temperature influences perception of the length of a wielded object via effortful touch. Exp Brain Res 236, 505–516 (2018). https://doi.org/10.1007/s00221-017-5148-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-017-5148-4