
Abstract
Damage to the temporal-parietal cortex in the right hemisphere often leads to spatial neglect—a disorder in which patients are unable to attend to sensory input from their contralesional (left) side. Neglect has been associated with both attentional and premotor deficits. That is, in addition to having difficulty with attending to the left side, patients are often slower to initiate leftward vs. rightward movements (i.e., directional hypokinesia). Previous research has indicated that a brief period of adaptation to rightward shifting prisms can reduce symptoms of neglect by adjusting the patient’s movements leftward, toward the neglected field. Although prism adaptation has been shown to reduce spatial attention deficits in patients with neglect, very little work has examined the effects of prisms on premotor symptoms. In the current study, we examined this in healthy individuals using leftward shifting prisms to induce a rightward shift in the egocentric reference frame, similar to neglect patients prior to prism adaptation. Specifically, we examined the speed with which healthy participants initiated leftward and rightward reaches (without visual feedback) prior to and following adaptation to either 17° leftward (n = 16) or 17° rightward (n = 15) shifting prisms. Our results indicated that, following adaptation, participants were significantly faster to initiate reaches towards targets located in the direction opposite the prism shift. That is, participants were faster to initiate reaches to right targets following leftward prism adaptation and were faster to initiate reaches to left targets following rightward prism adaptation. Overall, these results are consistent with the idea that prism adaptation can influence the speed with which a reach can be initiated toward a target in the direction opposite the prism shift, possibly through altering activity in neural circuits involved in reach planning.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Damage to the right temporo-parietal cortex often leads to spatial neglect—a heterogenous disorder in which patients are unable to attend to sensory stimuli (visual, auditory, tactile) on their contralesional (i.e., left) side, and may even cause the patient to neglect the left side of their own body (Danckert and Ferber 2006; Husain and Rorden 2003). In addition to an impairment in processing sensory events occurring on the contralesional side, patients with neglect are also slower to initiate leftward movements toward targets in left space (for a review see Coulthard et al. 2006). Neglect is a debilitating condition that is present in roughly 50% of patients following a right hemisphere stroke (Buxbaum et al. 2004), and the severity of neglect is correlated with a poorer overall functional recovery (Cherney et al. 2001; Di Monaco et al. 2011). As such, many different techniques have been developed to try and reduce symptoms of neglect in patients with the disorder (for a review see Luaute et al. 2006).
One technique that has been extensively investigated as a potential therapy for patients with neglect is the prism adaptation (PA) procedure pioneered by Rossetti and colleagues (Pisella et al. 2006; Rossetti et al. 1998). Prior to PA the patients are asked to close their eyes and point straight-ahead of their body midline. Typically, a patient with neglect will point far to the right of midline, as their egocentric reference frame has been shifted toward their ipsilesional (right) side (Karnath et al. 1998). The patient is then asked to wear a set of rightward shifting prism lenses that induce a 10° (or greater) rightward visual shift. While wearing the prisms, the patient is asked to reach toward targets that are placed to the left and right of their body midline. Initially, when the patient reaches toward the target they miss far to the right because of the rightward shift induced by the prisms. However, on subsequent reaches the patient learns to adjust their movements leftward to compensate for the rightward visual shift (Redding and Wallace 2006; Rossetti et al. 1998). After 5–10 min of pointing to targets while wearing the prisms, the glasses are removed and the patient is asked to close their eyes and point straight-ahead once again. Because the patient had to adjust their movements leftward to compensate for the rightward visual shift, their egocentric reference frame also shifts leftward, closer to true center (Pisella et al. 2006; Rossetti et al. 1998). In a seminal study, Rossetti et al. (1998) demonstrated that this leftward realignment of the egocentric reference frame in patients with neglect led to significant improvements on clinical tests of neglect such as line bisection, cancelation and figure copying.
Since the initial demonstration by Rossetti et al. (1998) numerous studies over the past 18 years have demonstrated that rightward PA can reduce a number of neglect symptoms such as the leftward disengage deficit (Nijboer et al. 2008; Schindler et al. 2009; Striemer and Danckert 2007), the rightward attentional bias (Berberovic et al. 2004; Striemer et al. 2007), tactile extinction (Maravita et al. 2003), oculomotor biases (Angeli et al. 2004; Serino et al. 2006), deficits in posture and balance (Tilikete et al. 2001), as well as problems with visual imagery (Rode et al. 2001).
In addition, a number of studies in healthy adults over the past 15 years have demonstrated that leftward shifting prisms can shift attention and perceptual biases rightward, in the direction opposite the prism shift (e.g., Colent et al. 2000). Using this leftward PA technique, several studies have demonstrated a small rightward shift in performance on tasks like line bisection and the landmark task (Colent et al. 2000; Herlihey et al. 2012; Michel and Cruz 2015; Michel et al. 2003a; Schintu et al. 2014; Striemer and Danckert 2010a; Striemer et al. 2016), covert visual attention (Striemer et al. 2006), posture and balance (Michel et al. 2003b), and tactile (Girardi et al. 2004) and visual exploration (Ferber and Murray 2005). Although leftward shifting prims can induce a small rightward shift in performance (Colent et al. 2000; Michel et al. 2003a; Schintu et al. 2014; Striemer et al. 2016), most previous studies in healthy adults have failed to observe any significant effects of rightward shifting prisms on tasks that measure attention or spatial biases; however, see Striemer et al. (2006) and Berberovic and Mattingley (2003) for notable exceptions. Interestingly, Goedert et al. (2010) have suggested that the effectiveness of leftward and rightward shifting prisms to induce after-effects in the direction opposite the prism shift may be related to pre-existing attentional biases such that participants with a larger leftward attentional bias (as measured by line bisection) are more likely to be influenced by left prisms, and vice versa for participants with a rightward attentional bias. Studying how PA can alter attention and spatial biases in healthy adults is important for two reasons: (1) it affords a more thorough understanding of how the visuomotor system is linked with other cognitive functions (e.g., attention, spatial perception), and (2) it may offer a better understanding of how PA influences the behavior of patients with neglect.
Although prisms may be a useful way to reduce some symptoms of neglect, and can be used to induce changes in attention and perceptual biases in healthy adults, other research suggests that prisms may have different effects depending on the nature of the experimental tasks used. Specifically, several studies suggest that prisms may have a greater influence on tests that require responses using the motor effectors that are directly influenced by adaptation (i.e., the eyes or the hand), but prisms may have less of an influence on tests that require purely perceptual responses (Danckert 2014; Dijkerman et al. 2003; Ferber et al. 2003; Sarri et al. 2010; Striemer and Danckert 2010a, b, 2013; Striemer et al. 2013). Furthermore, some of our recent work (Striemer et al. 2016) examining the effects of different magnitudes of leftward PA in healthy adults demonstrated that a larger leftward prism shift (17°) led to a greater rightward shift in manual line bisection compared to a smaller leftward prism shift (8.5°). In fact, the magnitude of the after-effect induced by PA (i.e., the rightward shift in straight-ahead pointing) was positively correlated with the magnitude of the observed rightward shift in manual line bisection. In contrast, magnitude of prism shift had no differential effect on a perceptual equivalent of the line bisection task (i.e., the landmark task). Thus, even in healthy adults, PA may have differential effects on motor vs. perceptual responses (Striemer et al. 2016).
One motor deficit that is commonly observed in patients with neglect is directional hypokinesia—a slowed ability to initiate reaches towards targets in contralesional (i.e., left) space (Buxbaum et al. 2004; Coulthard et al. 2006; Heilman et al. 1985; Husain et al. 2000; Mattingley et al. 1992, 1998; Sapir et al. 2007). Specifically, it has been observed that patients with neglect have a direction-specific reaching deficit such that they are slower to initiate leftward (but not rightward) reaches toward targets located in left space (Husain et al. 2000; Mattingley et al. 1998). Although directional hypokinesia is a symptom commonly associated with neglect (Buxbaum et al. 2004; Coulthard et al. 2006), it is important to note that this deficit can also be observed in right brain damaged patients without neglect (Rossit et al. 2009).
In the current study, we investigated whether PA could be used to induce a direction-specific alteration in the speed with which healthy participants could initiate reaches to left vs. right targets. To examine this possibility, in the current study, participants completed a target reaching task in which they had to point to targets located to the left or right of body midline as quickly and accurately as possible prior to, and following adaptation to either 17° leftward (n = 16), or 17° rightward (n = 15) shifting prisms. As mentioned previously, prior research examining the effects of prisms on tests of attention and spatial biases in healthy adults has commonly observed significant effects following leftward, but not rightward PA (e.g., Colent et al. 2000; Michel et al. 2003a; Schintu et al. 2014; Striemer et al. 2016). Given this, we predicted that leftward PA would have a significant influence on the speed with which participants could initiate reaches to targets in the direction opposite the prism shift (i.e., to right targets). Furthermore, given that rightward shifting prisms typically do not lead to significant changes on tasks that measure attention or spatial biases in healthy adults (for a recent review see Michel 2015), we predicted that rightward shifting prisms would have little or no effect on the speed with which participants could initiate reaches toward targets.
Methods
Participants
This study consisted of 31 right-handed (by self-report) undergraduate students (9 males; mean age 20.67 years; SD 3.91) from the MacEwan University psychology participant pool who participated for course credit. All participants had normal or corrected to normal vision. The participants provided informed consent prior to participating in the study. If the participant agreed to take part in the study they were randomly assigned into either the leftward (n = 16; 5 males, mean age 20.75, SD 3.97), or rightward shifting prisms group (n = 15; 4 males, mean age 20.6, SD 3.98). The experimental protocol was approved by the MacEwan University Research Ethics Board in accordance with the ethical standards of the 1964 Declaration of Helsinki (last update: Seoul, 2008).
Apparatus and procedure
Reaching task
In this study participants completed a reaching task in which they were asked to make leftward and rightward reaches to targets on a touch screen once prior to, and once following PA. The reaching task used in the current study was modified from Sapir et al. (2007). During the task participants were asked to wear special goggles (PLATO, Translucent Technologies) that could be made transparent to allow a view of the workspace, or opaque to eliminate the participant’s view of the workspace (Fig. 1). Participants sat with their head in a chin rest 45 cm away from a horizontally mounted 32″ widescreen touch screen (ELO touch systems) located at their midsagittal plane. A button (a serial mouse) that was aligned with the center of the touch screen was placed directly in front of the participant and served as the start position for each trial. During the task the touch screen constantly displayed a central fixation cross that was flanked by two unfilled squares (2.5 cm × 2.5 cm) with their centers 12° to the right and left of center (Fig. 1). All stimuli were white and were presented on a uniform black background in a dimly lit room.

The layout of reaching task. Participants sat in front of a horizontally mounted touch screen with their head in a chin rest. a During the task participants wore PLATO goggles (Translucent Technologies) that could be made opaque to eliminate their view of the workspace following reach onset. At the beginning of a trial the participant fixated on the central cross while holding down a start button. After a variable period of time (1500–3000 ms) a target stimulus (an asterisk) appeared in one of the peripheral boxes. b Following the onset of the target the participant reached to the target as quickly and accurately as possible while maintaining central fixation. Immediately after movement onset (i.e., the release of the start button) the PLATO goggles turned opaque thereby eliminating any visual feedback or knowledge of the accuracy of their reach with respect to the target. This was done to prevent any de-adaptation from the prisms during the reaching task
Throughout the task participants were told to always maintain fixation on the central cross, and to not look at the peripheral boxes. Fixation was directly monitored by the experimenter using a video camera that was positioned to monitor participant’s eyes. Participants had very little trouble maintaining fixation during the task. Each trial began with the participant holding down the start button with their right index finger while fixating on the central cross with the PLATO goggles open (i.e., transparent) to allow a clear view the workspace. Then, after a random delay interval of 1500–3000 ms, a target stimulus (i.e., a white asterisk: 1.1 cm by 1.1 cm) was presented for 2000 ms in the center of either the left or right box. The target appeared in one of the two boxes with equal probability in a random order. Upon presentation of the target, participants were instructed to release the start button and reach toward and touch the target as quickly and accurately as possible with their right index finger while maintaining central fixation. Importantly, upon release of the start button the PLATO goggles immediately (i.e., within 5 ms) switched from a clear to an opaque state, thereby eliminating all visual feedback and knowledge of endpoint accuracy during each trial (Fig. 1b). This was done to prevent de-adaptation from the prisms during the post-prisms reaching task. For each trial, we recorded reaction time (i.e., the time between the presentation of the target and the release of the start button), movement time (i.e., the time between the release of the start button and contact with the touch screen) and endpoint error (distance between the endpoint of the reach and the target location). After a touch response was registered by the screen there was a 2000-ms delay in which the PLATO goggles remained opaque (i.e., blocking vision of the workspace). Once the participant pressed and held down the start button, the goggles returned to a clear (i.e., transparent) state prior to commencing the next trial. A single block contained 50 trials (25 left and 25 right targets) in random order. Each participant completed two blocks (i.e., 100 trials) prior to and following PA with a short break in between each block to allow participants to rest.
Prism adaptation procedure
Following the baseline reaching task, participants were adapted to 30 diopter prisms (Bernell Products) that were used to induce either a 17° leftward (n = 16), or 17° rightward (n = 15) shift of the visual field.Footnote 1 The experimental procedure was identical for the two groups. During the PA procedure participants sat with their head in a chin rest in front of the touch screen with their right index finger on the start button. The initial starting position of the hand was partially obscured by the chinrest; however, participants had full vision of their hand during the reach (i.e., concurrent feedback). During each trial a circular target (1 cm in diameter) appeared either 10 cm to the left, or 10 cm to the right of the participant’s body midline. Participants were instructed to reach out and touch the target as quickly and accurately as possible with their right index finger. After touching the screen, the participant returned their finger to the start button and another target was presented. In total, participants completed 200 pointing trials (100 left targets and 100 right targets) presented in a random order while wearing the prisms. The PA task took roughly 7–10 min to complete.
In order to measure PA after-effects participants completed a straight-ahead pointing task before, and immediately after PA. In addition, a final straight-ahead pointing measurement was taken at the conclusion of the study to determine if participants remained adapted throughout the experiment. During the straight-ahead pointing task, the participant sat with their head in a chin rest. Participants were instructed to make five pointing movements straight-ahead of their body midline (i.e., subjective midline) with their eyes closed. The endpoint of each of the pointing movements was recorded by the surface of the touch screen. These endpoints were used to calculate subjective straight-ahead pre-PA, immediately post-PA and once again at the conclusion of the experiment.
To summarize, in the current study participants completed the tasks in the following order: (1) baseline (pre-prisms) reaching task, (2) baseline (pre-prisms) subjective straight-ahead pointing, (3) PA procedure (either 17° leftward or 17° rightward shifting prisms), (4) post-prisms subjective straight-ahead pointing, (5) post-prisms reaching task, and (6) final subjective straight-ahead pointing.
Data analysis
For the reaching task, average reaction (RT) and movement times (MT) were calculated [in milliseconds (ms)] for each trial for each participant. Any trials in which the RT was below 150 ms or more than 2SD above the participant’s mean RT for that condition were discarded as outliers. Outliers accounted for less the 5% of the data in each participant. For the endpoint error data from the reaching task, we calculated horizontal pointing error [in centimeters (cm)] by subtracting the finger landing position from the actual target location where negative values were errors to the left of the target location, and positive values were errors to the right of the target location. Initially, the data from the reaching tasks were analyzed with the use of a three-way mixed-model ANOVA with prisms group (left, right) as a between-subject factor, and time (pre, post), and target (left, right) as within-subject factors. We also carried out planned comparisons (using paired-samples t tests) examining the difference in reaction times for left targets pre vs. post prisms and right targets pre vs. post prisms. In addition, given that we expected a significant change in RT when initiating reaches toward targets in the direction opposite the prism shift, we also carried out a subsequent ANOVA analysis with the with prism group (left vs. right) as a between-subject factor, and time (pre vs. post) and target (ipsilateral or contralateral to the prism shift) as within-subject factors. Post-hoc tests were carried out using either paired-samples or independent-samples t tests with Bonferroni correction.
For the straight-ahead pointing task, we calculated the endpoint (in cm) for each of the five straight-ahead pointing movements in each session (pre, post, final) and then averaged across them. The straight-ahead pointing data were initially analyzed using a mixed-model ANOVA with group (left vs. right prisms) as a between-subject factor and time (pre, post, final) as a within-subject factor. This overall ANOVA was then followed up by separate one-way within-subject ANOVAs for the left and right prisms groups with time (pre vs. post vs. final) as the within-subject factor. We then used planned comparisons (using paired-samples t tests) to contrast the straight-head pointing data for the pre vs. post sessions, and the pre vs. final sessions, to determine if participants adapted, and remained adapted to the prisms.
Results
Prism adaptation
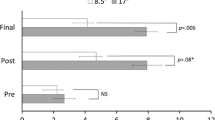
To determine whether participants successfully adapted to prisms we initially examined straight-ahead pointing performance using a mixed-model ANOVA with group (left vs. right prisms) as a between-subject factor and time (pre, post, final) as a within-subject factor. This analysis revealed main effects of time [F(2,58) = 10.58 p < 0.0001, η 2p = 0.27] and group [F(1,29) = 7.06, p = 0.013, η 2p = 0.20], as well as a group × time interaction [F(1,29) = 42.28, p < 0.0001, η 2p = 0.59]. To investigate the main effect of time we used paired samples t tests to carry out planned comparisons. The main effect of time indicated that, overall, the pre-prisms pointing session (0.85 cm) was significantly different from the post (4.25 cm) pointing session [t(30) = 4.30, p < 0.0001, d = 0.77]. In addition there was a trend for the pre-prisms pointing session (0.85 cm) to differ from the final pointing session [2.58 cm; t(30) = 1.96, p = 0.06, d = 0.35]. Finally, there was no significant difference between the post (4.25 cm) and final (2.58 cm) pointing sessions [t(30) = 1.66, p = 0.11, d = 0.29]. The main effect of group revealed that, overall, straight-ahead pointing judgments were larger for the left (4.97 cm) compared to the right (−0.013 cm) prisms group.
To further examine the group × time interaction we analyzed the effect of time separately for the left and right shifting prisms groups using within-subject ANOVAs with time (pre, post, final) as the within-subject factor. For the leftward shifting prisms group (n = 16) there was a significant effect of time [F(2,30) = 13.22, p < 0.0001, η 2p = 0.47; Fig. 2]. Planned comparisons revealed that, following leftward PA, straight-ahead pointing was shifted significantly rightward post (5.7 cm) in comparison to pre-adaptation] 1.9 cm; t(15) = 3.104, p = 0.007, d = 0.78]. We also found a difference in straight-ahead pointing between the pre (1.9 cm) and final pointing sessions [7.29 cm; t(15) = 6.298, p < 0.001, d = 1.57] indicating that participants were still adapted to prisms at the conclusion of the experiment.

The straight-ahead pointing data for the left (white bars) and right (gray bars) prisms groups as a function of time (pre, post, final). All data are reported in centimeters (cm) with negative values indicating a leftward shift and positive values indicating a rightward shift. *A statistically significant difference (p < 0.05). Error bars represent the within-subject standard error (Loftus and Masson 1994)
For the rightward shifting prisms group (n = 15) we also observed a significant effect of time [F(2,28) = 13.14, p < 0.0001, η 2p = 0.48; Fig. 2]. Planned comparisons revealed a significant difference in straight-ahead pointing pre (−0.31 cm) vs. post prisms [2.72 cm; t(14) = −2.909, p = 0.011, d = 0.75], but in the direction opposite than was expected. That is, participants initially pointed further to the right after rightward PA. However, by the end of the experiment there was a significant leftward shift in straight-ahead pointing when comparing the pre (−0.31 cm) and final pointing sessions [−2.44 cm; t(14) = 2.771, p = 0.015, d = 0.71; Fig. 2].
Finally, to compare the magnitude of adaptation effects between the two prisms groups we computed difference scores for the straight-ahead pointing data for pre vs. final and post vs. final. This analysis revealed that, for the pre vs. final pointing sessions, the leftward PA group (5.35 cm) had a larger change in straight-ahead pointing than the rightward PA group [−2.12 cm; t(29) = 6.50, p < 0.0001, d = 2.34]. However, if one considers the fact that immediately post PA the rightward PA group demonstrated a rightward shift in straight-ahead pointing (2.72 cm), but then demonstrated a significant leftward shift in straight-ahead pointing by the end of the experiment (−2.44 cm), then the total shift in straight-ahead pointing between the post and final sessions in the right PA group was (5.15 cm). Importantly, the total amount of shift post vs. final in the rightward PA group (5.15 cm) was no different than the total shift in straight-ahead pointing observed in the leftward PA group pre vs. post PA [5.35 cm; t(29) = 0.14, p = 0.89, d = 0.05]. Thus, the total amount of change in straight-ahead pointing between the groups in response to PA was the same by the end of the experiment.
Reaching task: reaction time
To analyze reaction time in the reaching task we used a three-way mixed-model ANOVA with group (left prisms, right prisms) as a between-subject factor, and time (pre, post), and target (left, right) as within-subject factors. We observed main effects of time and target. The main effect of time [F(1,29) = 5.724, p = 0.023, η 2p = 0.17] indicated that participants were faster to initiate reaches post-adaptation (433 ms) in comparison to pre-adaptation (450 ms). The main effect of target [F(1,29) = 46.666, p < 0.001, η 2p = 0.62] indicated that participants were faster to initiate reaches toward right targets (433 ms) in comparison to left targets (450 ms). There was also a significant time × target × group interaction [F(1,29) = 12.999, p < 0.001, η 2p = 0.31].
To further investigate the time × target × group interaction, we computed time × target ANOVAs separately for the leftward and rightward shifting prisms groups. For the leftward shifting prisms group we found a main effect of target [F(1,15) = 10.43, p = 0.006, η 2p = 0.41] and a significant time × target interaction [F(1,15) = 5.820, p = 0.029, η 2p = 0.28; Fig. 3a]. The main effect of target indicated that participants in the leftward shifting prisms group were faster to initiate movements toward right targets (428 ms) in comparison to left targets (442 ms) overall. Furthermore, in reference to the time × target interaction, planned comparisons indicated that the difference in reaction time for right targets pre (440 ms) vs. post prisms (415 ms) was significant [t(15) = 2.42, p = 0.029, d = 0.60], such that participants were faster to initiate rightward movements following leftward PA. However, there was no significant change in reaction times to left targets pre (451 ms) vs. (431 ms) post-adaptation [t(15) = 1.75, p = 0.10, d = 0.44; Fig. 3a].

a The reaction time (RT) data in milliseconds (ms) from the reaching task for the leftward and rightward shifting prisms groups as a function of time (pre vs. post) and side of target (left vs. right). b The RT data (in ms) from the reaching task as a function time (pre vs. post) and side of target with respect to direction of prism shift (i.e., ipsilateral vs. contralateral). *A statistically significant difference (p < 0.05). Error bars represent the within-subject standard error (Loftus and Masson 1994)
The same analysis for the rightward shifting prisms group also revealed a main effect of target [F(1,14) = 61.252, p < 0.001, η 2p = 0.81], and a significant time × target interaction [F(1,14) = 7.467, p = 0.016, η 2p = 0.35; Fig. 3a]. The main effect of target in the rightward shifting prisms group indicated that participants in the rightward shifting prisms group were faster to initiate movements toward right targets (438 ms) in comparison to left targets (459 ms). As for the time × target interaction, planned comparisons failed to demonstrate a significant difference for reaction times when initiating reaches toward right targets before versus after PA [t(14) = 0.524, p = 0.609, d = 0.13]. However, we did observe a non-significant trend toward faster reaction times to initiate reaches to left targets post (450 ms) compared to pre-PA [468 ms; t(14) = 1.838, p = 0.087, d = 0.47; Fig. 3a].Footnote 2
To further analyze the effects of PA on reaction times to targets in the direction opposite the prism shift, we conducted a mixed-model ANOVA with group (left vs. right prisms) as a between-subject factor, and time (pre vs. post) and target (ipsilateral or contralateral to the direction of prism shift) as within-subject factors. Following this analysis we observed a significant main effect of time [F(1,29) = 5.72, p = 0.023, η 2p = 0.17], such participants were faster to respond post (433 ms) vs. pre (450 ms) PA. In addition, we also observed significant target × group [F(1,29) = 46.66, p < 0.0001, η 2p = 0.62] and time × target [F(1,29) = 12.99, p = 0.001, η 2p = 0.31] interactions.
The target × group interaction demonstrated that the difference in reaction time to ipsilateral minus contralateral targets was significant between the two groups. Specifically, overall, the leftward shifting prisms group was faster for contralateral (428 ms) vs. ipsilateral targets (442 ms; diff. = −14 ms), whereas the rightward shifting prisms group, overall, was faster for ipsilateral (438 ms) vs. contralateral targets [459 ms; diff. = 21 ms; t(29) = 6.8, p < 0.0001, d = 2.47]. Note that this is, essentially, the same as the main effect of target demonstrated in the previous analysis such that participants were faster overall to initiate movements right vs. left targets. Specifically, the left prisms group was faster overall for contralateral (i.e., right targets), whereas the right prisms group was faster overall for ipsilateral (i.e., right) targets.
More importantly, the time × target interaction (Fig. 3b) revealed that there was a significant reduction in RTs for targets presented on the side opposite the prism shift (i.e., contralateral targets) when comparing pre PA (454 ms) vs. post PA [432 ms; (t(30) = 3.07, p = 0.01, Bonferroni corrected, d = 0.56]. However, there was no significant reduction in RT for targets presented on the same side as the prism shift (i.e., ipsilateral) when comparing pre PA (446 ms) vs. post PA [434 ms; t(30) = 1.71, p = 0.18, Bonferroni corrected, d = 0.31]. This interaction clearly demonstrates that PA resulted in a reduction in RTs for initiating movements toward targets presented on the side opposite the prism shift for both the left and the right prisms groups. Furthermore, this effect was of equal magnitude for both the leftward and rightward shifting prisms groups as there was no three-way interaction with group (p = 0.20).
Reaching task: movement time data
To analyze movement time in the reaching task we used a three-way mixed-model ANOVA with group (left prisms, right prisms) as between-subject factor, and time (pre, post), and target (left, right) as within-subject factors. We found a main effect of target [F(1,29) = 107.92, p < 0.001, η 2p = 0.78], and a marginally significant target × group interaction [F(1,29) = 3.96, p = 0.056, η 2p = 0.12]. The main effect of target indicated that, overall, participants were faster to execute reaches to right (785 ms) compared to left (842 ms) targets. Given that we had no specific hypotheses regarding the possible effects of PA on MT, we did not carry out any additional analyses.
Reaching task: endpoint error data
To analyze horizontal pointing error in the reaching task, we used the same mixed-model ANOVA with group (left vs. right prisms) as the between-subject factor and time (pre vs. post) and target (left vs. right) as the within-subject factors. This analysis revealed a main effect of target [F(1,29) = 37.08, p < 0.0001, η 2p = 0.56] such that, overall, pointing errors were greater for left (−2.45 cm) compared to right targets (0.89 cm). In addition, there was also a significant time × group interaction [F(1,29) = 226.56, p < 0.0001, η 2p = 0.89; Fig. 4]. The time × group interaction demonstrated that, although there was no difference in pointing errors between the groups prior to PA [left prisms = −0.96 cm vs. right prisms = −0.84 cm; t(29) = 0.143, p = 0.89, d = 0.05], there was a significant difference in pointing errors following PA. Specifically, pointing errors for the leftward PA group were to the right of the target (4.16 cm) post PA, whereas pointing errors for the rightward PA group were to the left of the target post PA [−5.5 cm; t(29) = 10.74, p < 0.0001, d = 1.96]. Importantly, the difference in pointing errors pre vs. post adaptation were also significantly different between the two groups [left prisms = −5.13 cm vs. right prisms = 4.61 cm; t(29) = 15.05, p < 0.0001, d = 2.75] indicating that the effect of PA on pointing errors was in opposite directions for the two groups (i.e., in the direction opposite the prism shift).

The horizontal endpoint error data in centimeters (cm) as a function of group (left prisms vs. right prisms) and time (pre vs. post). *A statistically significant difference (p < 0.05). Error bars represent the within-subject standard error (Loftus and Masson 1994)
Correlation analysis
To examine the possible relationship between changes in straight-ahead pointing and changes in RT before and after PA we conducted a correlation analysis on pre-post difference scores separately for the leftward and right shifting prisms groups. Specifically, if larger changes in straight-head pointing were related to larger changes in RT post PA then one would expect a positive correlation between the two variables (see Striemer et al. 2016). Thus, for straight-ahead pointing we subtracted data from the ‘final’ pointing session (at the end of the experiment) from the ‘pre’ pointing session. For the RT data, given that we observed the largest effects of prisms on RTs to targets contralateral vs. ipsilateral to the prism shift, we subtracted RTs from targets contralateral to the prism shift, from RTs to targets ipsilateral to the prism both pre and post PA. We then computed a final difference score that examined the change in the difference in RTs for targets ipsi vs. contralateral to the prism shift before vs. after PA (i.e., we subtracted the pre ipsi vs. contra difference score from the post ipsi vs. contra difference score).
For the leftward PA group this analysis revealed that there was a tendency for larger changes in straight-head pointing (i.e., a larger rightward shift) to be correlated with larger changes in RT to targets ipsilateral vs. contralateral to the prism shift pre vs. post PA [r(16) = 0.43, p = 0.085, one-tailed]. Furthermore, when the same analysis was conducted for the rightward PA group there was no such trend toward a positive correlation [r(15) = 0.05, p = 0.85, one-tailed]. It is important to emphasize that these correlations need to be interpreted with caution as they are based on a small number of data points (Fig. 5).

The correlation between changes in straight-ahead pointing (final–pre; cm) and changes in the difference in reaction times between targets presented ipsilateral vs. contralateral (ipsi–contra; ms) to the prism shift pre vs. post prisms for the left (a) and right (b) prisms groups
Discussion
In the present study, we were interested in whether adaptation to leftward or rightward shifting prisms could influence the speed with which healthy adults could initiate reaches to left and right targets. Numerous previous studies in healthy adults have demonstrated that, following leftward PA, there is a rightward shift in performance on tests of attentional and spatial biases (for a recent review see Michel 2015). In addition, most previous studies have failed to demonstrate any consistent effects of rightward shifting prisms on attentional and spatial biases in healthy adults (e.g., Colent et al. 2000; Michel et al. 2003a; Schintu et al. 2014; Striemer et al. 2016). Given these previous findings, we predicted that leftward PA would alter the speed with which participants could initiate reaches toward targets in the direction opposite the prism shift (i.e., to right targets). However, we expected no effects following rightward PA.
The results from the current study indicated that, similar to previous studies (Colent et al. 2000; Michel et al. 2003a; Schintu et al. 2014; Striemer et al. 2016), there was a significant rightward shift in straight-ahead pointing following leftward PA (Fig. 2). However, in the rightward shifting prisms group, there was initially a rightward shift in straight-ahead pointing immediately post-prisms, followed by a significant leftward shift by the end of the experiment. The initial rightward shift in straight-ahead pointing (i.e., immediately post PA) was unexpected as most (if not all) previous studies to our knowledge have demonstrated a significant leftward shift in straight-ahead pointing following rightward PA (e.g., Colent et al. 2000; Michel et al. 2003a; Schintu et al. 2014; Striemer et al. 2016). We are unsure as to why we observed this effect and, given that is has not been observed in previous studies, it may be a spurious finding. Nevertheless, we do not feel that the initial rightward shift in straight-ahead pointing following rightward PA had a negative impact on our study for two reasons. First, participants in the rightward PA group demonstrated the expected leftward after-effect by the end of the experiment. In fact, if you compare the difference in straight-ahead pointing between the post PA and final pointing sessions for the rightward PA group, it is of the same magnitude of that observed in the leftward PA group when comparing pre vs. final. This suggests that the overall amount of shift in straight-ahead pointing was the same between the two groups. The second reason we do not feel that the initial rightward shift in pointing in the right PA group influenced our results comes from our analysis of the endpoint error data from the post-prisms reaching task (Fig. 4). Specifically, if you look at the endpoint errors for reaches during the post-prisms reaching task, they are in the direction opposite the prism shift for both groups (i.e., left errors for right prisms and vice versa for left prisms). This is exactly what one would expect from someone who had successfully adapted to a rightward prism shift. Thus, we are confident that, overall, participants adapted equally to left and right prisms in the two groups and that they remained adapted throughout the experiment. This is particularly important in the current study as participants completed 100 reaching movements in the post-prisms reaching task which may have been expected to result in de-adaptation. However, because we were able to remove all visual feedback during the reach using the PLATO goggles, we were able to limit de-adaptation from the prisms.
The most interesting results from the current study were observed in the reaction time data from the reaching task. Similar to previous studies (Sapir et al. 2007) participants were, overall, faster when reaching toward right compared to left targets with their right hand. In addition, we observed that, following PA, participants were faster to initiate reaches toward targets in the direction opposite the prism shift. That is, following leftward PA, participants were faster to initiate reaches to right targets, whereas following rightward PA participants were faster to initiate reaches to left targets (Fig. 3a, b). This finding was further emphasized in a subsequent analysis that examined the influence of PA as a function of target side (i.e., ipsilateral vs. contralateral) with respect to direction of prism shift (i.e., for left prisms left targets = ipsilateral, right targets = contralateral, and vice versa for right prisms). Specifically, this analysis revealed a significant time (pre vs. post PA) by target (ipsilateral vs. contralateral) interaction such that participants were faster to initiate reaches toward contralateral targets (i.e., targets in the direction opposite the prism shift) following PA. Importantly, there was no interaction with group (i.e., direction of prism shift) indicating that the effects of PA on reach RT were equivalent for both the left and right PA groups (Fig. 3b).
Analysis of the movement time (MT) data indicated that, overall, participants were faster to initiate reaches to right compared to left targets when reaching with the right hand. These results are consistent with the RT data reported earlier. In addition, although PA was able to speed RT to initiate reaches to targets in the direction opposite the prism shift, PA had no systematic effects on MT (i.e., no differences emerged pre vs. post PA).
Finally, in order to examine the relationship between changes in straight-ahead pointing pre vs. post PA and changes in performance in the reaching task we conducted a correlation analysis on difference scores. Specifically, we correlated changes in straight-ahead pointing (final minus pre) with changes in the difference in RT to targets that were ipsilateral vs. contralateral to the prism shift before vs. after PA. Interestingly, for the leftward (but not the rightward) PA group, there was a trend [r(16) = 0.43, p = 0.085, one-tailed] toward a positive correlation between changes in straight-ahead pointing, and changes in the difference in RT for targets ipsilateral vs. contralateral to the prism shift pre vs. post PA. That is, larger rightward shifts in straight-ahead pointing following leftward PA were correlated with larger changes in the difference in RT between targets ipsilateral vs. contralateral to the prism shift pre vs. post PA.
Although this trend needs to be interpreted with caution because it is only based on a small number of data points, it is worth pointing out that the relationship is in the direction one would expect if changes in straight-ahead pointing were linked with changes in RT post-prisms. Furthermore, this trend is also consistent with recent data from our lab in which we observed a significant positive correlation between changes in straight-ahead pointing, and changes in manual line bisection pre vs. post PA (Striemer et al. 2016). These data also support the more general notion that the beneficial effects of prism on attention and spatial biases may be directly tied to changes in motor performance that are induced by PA (Striemer and Danckert 2010a, b). Finally, it is also worth pointing out that the trend toward a correlation between changes in straight-ahead pointing and changes in RT was only observed in the leftward shifting prisms group. Although it is not clear why this trend was only observed for the leftward shifting prisms group, it may be related to the fact that changes in straight-ahead pointing in the expected direction took longer to develop in the rightward shifting prisms group.
Although we have demonstrated that prisms can speed RTs to initiate reaches toward targets in the direction of the prism after-effect, it is still unclear how PA was able induce this effect. Specifically, PA could speed reach RTs toward targets in the direction opposite the prism shift by either, (1) inducing a bias in attention in the direction of the prism after-effect, thereby speeding target detection, and hence, motor RT or (2) by increasing the speed with which the participant could plan movements towards targets in the direction of the prism after-effect once the target was detected. Although, in theory, either explanation could account for our effects, we believe that it is more likely the case that prisms speeded the initiation of the reach towards the target, rather than speeding target detection per se. We will consider each of these theories in turn.
The notion that PA might alter the distribution of spatial attention by rebalancing activity between the two hemispheres has been suggested since the original studies of PA in patients with neglect and healthy adults were first published (for reviews of possible mechanisms see Clarke et al. 2016; Michel 2006; Pisella et al. 2006; Redding and Wallace 2006; Striemer and Danckert 2010b). The first evidence that PA might alter the allocation of attention in healthy adults was reported by Colent et al. (2000) and Michel et al. (2003a). Specifically, using manual line bisection and landmark tasks they demonstrated a significant rightward shift in manual bisection (Michel et al. 2003a) and a rightward shift in the perceived midpoint of the line in the landmark task (Colent et al. 2000; Michel et al. 2003a) following leftward, but not rightward PA. This supported the idea that, in healthy adults, leftward PA led to a rightward shift in attention by increasing activity in the left posterior parietal cortex (PPC) and simultaneously decreasing activity in the right PPC (for a review see Michel 2006). A rightward shift in attention following leftward PA (and vice versa following rightward PA) could provide a simple explanation for the results of the current study. Namely, following leftward PA participants may have experienced a rightward shift in attention which led to faster detection of right targets, and hence, larger changes in motor RTs for right, but not left targets. The same mechanism (in the opposite direction) could explain the results for the rightward shifting prisms group. That is, a speeding of RTs for left but not right targets following rightward PA.
If a simple visual attentional bias account could explain the results from the current study, then there should be ample evidence that PA can speed RTs to detect targets in the direction opposite the prism shift in healthy adults in previous studies; however, this is not the case. In a previous study Striemer et al. (2006) found that PA could speed the reorienting of reflexive covert attention away from a cue in the direction opposite the prism shift. That is, participants were faster to reorient attention away from a right cue to a left target following leftward PA and were faster to reorient attention away from a left cue to a right target following rightward PA (Striemer et al. 2006). It is important to note that, in the Striemer et al. (2006) study, PA had no effect on RTs to non-cued targets (i.e., targets that appeared without any preceding cue). This is consistent with previous work in healthy adults showing no changes in simple RTs to detect left or right targets following PA in the absence of a spatial cue (Berberovic et al. 2004; Morris et al. 2004). A recent study by Martin-Arevalo et al. (2016) lends further support to Striemer et al.’s (2006) findings. Specifically, Martin-Arevalo et al. (2016) found that leftward (but not rightward) PA reduced ERP amplitudes associated with shifting attention leftward, as well as detecting invalidly cued left targets. Note, however, that no behavioral (i.e., RT) effects were observed by Martin-Arevalo et al. (2016) in response to PA. Nevertheless, these two studies indicate that, following leftward PA, there are changes in the efficiency with which participants can redirect attention away from a cue on the side opposite the prism shift. However, there is no clear evidence that PA can influence RTs for non-cued targets. It is important to emphasize that, in the current study, participants were required to initiate a reach towards a non-cued target that appeared on the left or right side of the screen. Given that no previous studies in healthy adults have ever observed changes in simple RT to detect non-cued targets following PA, we feel that it is unlikely that our results can be explained by a simple visuospatial attentional bias account.
One aspect of the current study that is unique is that this is the first study (at least to our knowledge) that has directly examined changes in RT to initiate movements to left and right targets pre vs. post PA. Specifically, our study required participants not only to attend to and detect the target, but also to plan and execute a reach towards the same target (without visual feedback). Given that previous studies in healthy adults have only observed effects of prisms on attention tasks that require reorienting attention away from a cue (Martin-Arevalo et al. 2016; Striemer et al. 2006) and have failed to demonstrate any effects of PA on simple RT to detect left or right targets (Berberovic et al. 2004; Morris et al. 2004; Striemer et al. 2006), we are inclined to believe that prisms must have altered the speed with which participants were able to plan their movements to targets in the direction opposite the prism shift.
To clarify, we are not suggesting that attention was unaffected by prisms in the current study, only that a simple visuospatial attentional bias account post-prisms is unlikely to explain our effects. Interestingly, a good deal of previous work has suggested that the left hemisphere may be “dominant” for fine motor control, such that it is responsible for planning and executing skilled movements with both hands (Kimura, 1993; Perenin and Vighetto 1988; Rothi and Heilman 1997). In addition, numerous studies have suggested that visuospatial attention (i.e., attending to different locations in space) and so-called “motor attention” (i.e., the ability to plan motor actions to different locations) rely on separable cognitive and neural mechanisms (Jonikaitis and Deubel 2011; Rushworth et al. 1997, 2001a, b). Specifically, the right PPC (in particular the right inferior parietal lobe and temporal-parietal junction) is thought to be critical for reorienting visuospatial attention (e.g., Corbetta et al. 2000, 2002), whereas the left PPC (in particular, the left supramarginal gyrus) is thought to be important for controlling motor attention when planning reaching movements (Rushworth 1997, 2001a, b). Additional evidence in support of the separation of visual attention from motor attention comes from the fact that deficits in the speed with which one can detect the presence of a visual target (i.e., visuospatial attention) are not correlated with deficits in visuomotor control (Striemer et al. 2009). One possible explanation for the current findings is that, because participants adapted to prisms by reaching to targets with the right hand which is controlled by the left PPC, this may have influenced the left PPC’s ability to allocate motor attention (i.e., to plan movements), thereby speeding motor RTs toward targets in the direction opposite the prism shift. Note that this is consistent with several previous studies that have demonstrated that the left PPC is actively engaged during PA with the right hand (e.g., Clower et al. 1996; Danckert et al. 2008; Luaute et al. 2009).
Another potential mechanism to explain the results of the current study comes from a study by Magnani et al. (2014) who demonstrated that PA increases intracortical excitability [as measured via paired-pulse transcranial magnetic stimulation (TMS)] in the primary motor cortex (M1) contralateral to the prism-induced after-effect. That is, leftward PA, which induces a rightward after-effect, was found to increase the excitability of M1 in the left hemisphere. In contrast, rightward PA, which induces a leftward after-effect, was found to increase the excitability of M1 in right hemisphere. Critically, in the Magnani et al. study (2014; as in the current study), all reaches were made with the dominant right hand. Thus, the changes in M1 excitability observed by Magnani et al. (2014) cannot be explained by the hand used during adaptation, but instead are tied to the direction of the prism after-effect. Magnani et al. (2014) further postulated that the changes in M1 activity that arise following PA might possibly spread to other areas within the activated hemisphere. Thus, as per Magnani et al. (2014), an increase in M1 activity in the left hemisphere following leftward PA might then spread to other motor areas within the same hemisphere, creating a generally more active left hemisphere (and vice versa for rightward PA). Therefore, if leftward PA results in an increase in M1 excitability in the left hemisphere, and this effect then spreads to other motor related regions of the left hemisphere such as the PPC, then this could result in faster RTs to initiate movements to right targets. The reverse of this could also explain faster RTs to initiate movements to left targets following rightward PA. Again, if rightward PA increases activity in right M1 (Magnani et al. 2014), and this increased activity spreads to the right PPC, then this could explain why participants were faster to initiate reaches to left targets following rightward PA (i.e., increased activity in visuomotor regions of the right hemisphere). To put it simply, according to the results of Magnani et al. (2014), PA might influence reach RTs to targets in the direction opposite the prism shift through creating something akin to a “field effect” that is often observed in patients with optic ataxia following damage to the PPC. That is, patients with optic ataxia are slower and less accurate at executing reaches towards targets on the contralesional side with either hand (Perenin and Vighetto 1988; Pisella et al. 2007; Striemer et al. 2007). In contrast, by increasing activity in motor regions (i.e., M1 and/or PPC) in the hemisphere opposite the prism after-effect, it might be possible to create the reverse of this effect, namely faster RTs to initiate reaches to targets in the direction opposite the prism shift.
Ultimately, whether our results can be better explained through prisms altering motor attention mechanisms in the left PPC, or though prisms creating a “field effect” by increasing activity in motor regions in the hemisphere opposite the prism after-effect, remains an open topic for future investigation. Future studies could adopt our behavioral paradigm in concert with measures such as EEG/ERP recordings to examine whether activity in motor regions of the two hemispheres are systematically altered by PA, and whether these changes are related to observed effects in RT to initiate reaches towards targets. Using electrophysiological recordings would also allow a better separation between the effect of prisms on sensory aspects of target detection, from mechanisms related to motor preparation, and reach planning. Finally, another important question for future investigation is whether the effects of PA on reach initiation RT transfer to the un-adapted hand.
It is important to note that the results from the current study could also potentially be extended to the treatment of directional hypokinesia in patients with right brain damage. That is, if PA increases hemispheric activity in the direction opposite the prism after-effect (Magnani et al. 2014), or makes it easier for the left PPC to allocate motor attention to targets in left space, then rightward PA could make it easier for patients with right brain damage to initiate leftward reaches.
Conclusion
In conclusion, the results of the current study demonstrate that PA can speed reach initiation in the direction of the prism-induced after-effect. Specifically, participants were faster to initiate reaches towards right targets following leftward PA and vice versa following rightward PA. Our results could be explained through PA altering brain circuits in the left PPC that are involved in controlling motor attention (i.e., reach planning), or through the creation of a “field effect” via increases in cortical excitability in M1 in the hemisphere opposite the prism after-effect (Magnani et al. 2014). Taken together, the current data add further support to the notion that PA can influence performance in tasks that require a motor response with the adapted hand (Striemer and Danckert 2010a, b; Striemer et al. 2016). Finally, our results may be applied to help treat directional hypokinesia in patients with right hemisphere brain damage.
Notes
For smaller angles 1 prism diopter = 0.57° or 1° = 1.75 prism diopters (Millodot 2009).
In order to determine whether the delayed leftward shift in straight-ahead pointing in our rightward shifting prisms group may have influenced the data in our reaching task, we also compared the RT data from the first half versus the second half of the post-prisms reaching task. To analyze this we conducted a two-way within-subject ANOVA with time (first half, second half) and target (left, right) as factors. No significant effects were observed with this analysis. This suggests that the delayed leftward shift in straight-ahead pointing following rightward prism adaptation did not influence the RT data in the post-prisms reaching task.
References
Angeli V, Benassi MG, Ladavas E (2004) Recovery of oculo-motor bias in neglect patients after prism adaptation. Neuropsychologia 42(9):1223–1234
Berberovic N, Mattingley JB (2003) Effects of prismatic adaptation on judgements of spatial extent in peripersonal and extrapersonal space. Neuropsychologia 41(4):493–503
Berberovic N, Pisella L, Morris AP, Mattingley JB (2004) Prismatic adaptation reduces biased temporal order judgements in spatial neglect. Neuroreport 15(7):1199–1204
Buxbaum LJ, Ferraro MK, Veramonti T, Farné A, Whyte J, Ladavas E, Coslett HB (2004) Hemispatial neglect: subtypes, neuroanatomy, and disability. Neurology 62(5):749–756
Cherney LR, Halper AS, Kwasnica CM, Harvey RL, Zhang M (2001) Recovery of functional status after right hemisphere stroke: relationship with unilateral neglect. Arch Phys Med Rehabil 82(3):322–328
Clarke S, Crottaz-Herbette S (2016) Modulation of visual attention by prismatic adaptation. Neuropsychologia 92:31–41. doi:10.1016/j.neuropsychologia.2016.06.022
Clower DM, Hoffman JM, Votaw JR, Faber TL, Woods RP, Alexander GE (1996) Role of posterior parietal cortex in the recalibration of visually guided reaching. Nature 383(6601):618–621
Colent C, Pisella L, Bernieri C, Rode G, Rossetti Y (2000) Cognitive bias induced by visuo-motor adaptation to prisms: a simulation of unilateral neglect in normal individuals? Neuroreport 11(9):1899–1902
Corbetta M, Shulman GL (2002) Control of goal-directed and stimulus-driven attention in the brain. Nat Rev Neurosci 3(3):201–215
Corbetta M, Kincade JM, Ollinger JM, McAvoy MP, Shulman GL (2000) Voluntary orienting is dissociated from target detection in human posterior parietal cortex. Nat Neurosci 3(3):292–297
Coulthard E, Parton A, Husain M (2006) Action control in visual neglect. Neuropsychologia 44(13):2717–2733
Danckert J (2014) Spatial neglect: not simply disordered attention. In: The behavioral consequences of stroke. Springer, Berlin, pp 71–94
Danckert J, Ferber S (2006) Revisiting unilateral neglect. Neuropsychologia 44(6):987–1006
Danckert J, Ferber S, Goodale MA (2008) Direct effects of prismatic lenses on visuomotor control: an event-related functional MRI study. Eur J Neurosci 28(8):1696–1704. doi:10.1111/j.1460-9568.2008.06460.x
Di Monaco M, Schintu S, Dotta M, Barba S, Tappero R, Gindri P (2011) Severity of unilateral spatial neglect is an independent predictor of functional outcome after acute inpatient rehabilitation in individuals with right hemispheric stroke. Arch Phys Med Rehabil 92(8):1250–1256. doi:10.1016/j.apmr.2011.03.018
Dijkerman HC, McIntosh RD, Milner AD, Rossetti Y, Tilikete C, Roberts RC (2003) Ocular scanning and perceptual size distortion in hemispatial neglect: effects of prism adaptation and sequential stimulus presentation. Exp Brain Res 153(2):220–230
Ferber S, Murray LJ (2005) Are perceptual judgments dissociated from motor processes?—A prism adaptation study. Cogn Brain Res 23(2–3):453–456
Ferber S, Danckert J, Joanisse M, Goltz HC, Goodale MA (2003) Eye movements tell only half the story. Neurology 60(11):1826–1829
Girardi M, McIntosh RD, Michel C, Vallar G, Rossetti Y (2004) Sensorimotor effects on central space representation: prism adaptation influences haptic and visual representations in normal subjects. Neuropsychologia 42(11):1477–1487
Goedert KM, Leblanc A, Tsai SW, Barrett AM (2010) Asymmetrical effects of adaptation to left- and right-shifting prisms depends on pre-existing attentional biases. J Int Neuropsychol Soc 16(5):795–804. doi:10.1017/S1355617710000597
Heilman KM, Bowers D, Coslett HB, Whelan H, Watson RT (1985) Directional hypokinesia: prolonged reaction times for leftward movements in patients with right hemisphere lesions and neglect. Neurology 35(6):855–859
Herlihey TA, Black SE, Ferber S (2012) Terminal, but not concurrent prism exposure produces perceptual aftereffects in healthy young adults. Neuropsychologia 50(12):2789–2795. doi:10.1016/j.neuropsychologia.2012.08.009
Husain M, Rorden C (2003) Non-spatially lateralized mechanisms in hemispatial neglect. Nat Rev Neurosci 4(1):26–36
Husain M, Mattingley JB, Rorden C, Kennard C, Driver J (2000) Distinguishing sensory and motor biases in parietal and frontal neglect. Brain 123(Pt 8):1643–1659
Jonikaitis D, Deubel H (2011) Independent allocation of attention to eye and hand targets in coordinated eye-hand movements. Psychol Sci 22(3):339–347. doi:10.1177/0956797610397666
Karnath HO, Niemeier M, Dichgans J (1998) Space exploration in neglect. Brain 121(Pt 12):2357–2367
Kimura D (1993) Neuromotor mechanisms in human communication. Oxford University Press, Oxford
Loftus GR, Masson MEJ (1994) Using confidence intervals in within-subject designs. Psychon Bull Rev 1:476–490
Luaute J, Halligan P, Rode G, Rossetti Y, Boisson D (2006) Visuo-spatial neglect: a systematic review of current interventions and their effectiveness. Neurosci Biobehav Rev 30(7):961–982
Luaute J, Schwartz S, Rossetti Y, Spiridon M, Rode G, Boisson D, Vuilleumier P (2009) Dynamic changes in brain activity during prism adaptation. J Neurosci 29(1):169–178. doi:10.1523/JNEUROSCI.3054-08.2009
Magnani B, Caltagirone C, Oliveri M (2014) Prismatic adaptation as a novel tool to directionally modulate motor cortex excitability: evidence from paired-pulse TMS. Brain Stimul 7(4):573–579. doi:10.1016/j.brs.2014.03.005
Maravita A, McNeil J, Malhotra P, Greenwood R, Husain M, Driver J (2003) Prism adaptation can improve contralesional tactile perception in neglect. Neurology 60(11):1829–1831
Martin-Arevalo E, Laube I, Koun E, Farne A, Reilly KT, Pisella L (2016) Prism adaptation alters electrophysiological markers of attentional processes in the healthy brain. J Neurosci 36(3):1019–1030. doi:10.1523/JNEUROSCI.1153-15.2016
Mattingley JB, Bradshaw JL, Phillips JG (1992) Impairments of movement initiation and execution in unilateral neglect. Directional hypokinesia and bradykinesia. Brain 115(Pt 6):1849–1874
Mattingley JB, Husain M, Rorden C, Kennard C, Driver J (1998) Motor role of human inferior parietal lobe revealed in unilateral neglect patients. Nature 392(6672):179–182
Michel C (2006) Simulating unilateral neglect in normals: myth or reality? Restor Neurol Neurosci 24:419–430
Michel C (2015) Beyond the sensorimotor plasticity: cognitive expansion of prism adaptation in healthy individuals. Front Psychol 6:1979. doi:10.3389/fpsyg.2015.01979
Michel C, Cruz R (2015) Prism adaptation power on spatial cognition: adaptation to different optical deviations in healthy individuals. Neurosci Lett 590:145–149. doi:10.1016/j.neulet.2015.02.001
Michel C, Pisella L, Halligan PW, Luaute J, Rode G, Boisson D, Rossetti Y (2003a) Simulating unilateral neglect in normals using prism adaptation: implications for theory. Neuropsychologia 41(1):25–39
Michel C, Rossetti Y, Rode G, Tilikete C (2003b) After-effects of visuo-manual adaptation to prisms on body posture in normal subjects. Exp Brain Res 148(2):219–226
Millodot M (2009) Dictionary of optometry and visual science, 7th edn. Butterworth Heinemann-Elsevier, United Kingdom
Morris AP, Kritikos A, Berberovic N, Pisella L, Chambers CD, Mattingley JB (2004) Prism adaptation and spatial attention: a study of visual search in normals and patients with unilateral neglect. Cortex 40(4–5):703–721
Nijboer TC, McIntosh RD, Nys GM, Dijkerman HC, Milner AD (2008) Prism adaptation improves voluntary but not automatic orienting in neglect. Neuroreport 19(3):293–298
Perenin MT, Vighetto A (1988) Optic ataxia: a specific disruption in visuomotor mechanisms. I. Different aspects of the deficit in reaching for objects. Brain 111(3):643–674
Pisella L, Rode G, Farné A, Tilikete C, Rossetti Y (2006) Prism adaptation in the rehabilitation of patients with visuo-spatial cognitive disorders. Curr Opin Neurol 19(6):534–542
Pisella L, Striemer C, Blangero A, Gaveau V, Revol P, Salemme R, Rossetti Y (2007) Perceptual deficits in optic ataxia? In: Haggard P, Rossetti Y, Kawato M (eds) Sensorimotor foundations of higher cognition: attention and performance XXII. Oxford University Press, Oxford, pp 47–72
Redding GM, Wallace B (2006) Prism adaptation and unilateral neglect: review and analysis. Neuropsychologia 44(1):1–20
Rode G, Rossetti Y, Boisson D (2001) Prism adaptation improves representational neglect. Neuropsychologia 39(11):1250–1254
Rossetti Y, Rode G, Pisella L, Farné A, Li L, Boisson D, Perenin MT (1998) Prism adaptation to a rightward optical deviation rehabilitates left hemispatial neglect. Nature 395(6698):166–169
Rossit S, Malhotra P, Muir K, Reeves I, Duncan G, Livingstone K, Harvey M (2009) No neglect-specific deficits in reaching tasks. Cereb Cortex 19(11):2616–2624. doi:10.1093/cercor/bhp016
Rothi L, Heilman KM (eds) (1997) Apraxia: the neuropsychology of action. Psychology Press, UK
Rushworth MF, Nixon PD, Renowden S, Wade DT, Passingham RE (1997) The left parietal cortex and motor attention. Neuropsychologia 35(9):1261–1273
Rushworth MF, Ellison A, Walsh V (2001a) Complementary localization and lateralization of orienting and motor attention. Nat Neurosci 4(6):656–661. doi:10.1038/88492
Rushworth MF, Krams M, Passingham RE (2001b) The attentional role of the left parietal cortex: the distinct lateralization and localization of motor attention in the human brain. J Cogn Neurosci 13(5):698–710. doi:10.1162/089892901750363244
Sapir A, Kaplan JB, He BJ, Corbetta M (2007) Anatomical correlates of directional hypokinesia in patients with hemispatial neglect. J Neurosci 27(15):4045–4051
Sarri M, Greenwood R, Kalra L, Driver J (2010) Prism adaptation does not change the rightward spatial preference bias found with ambiguous stimuli in unilateral neglect. Cortex 47(3):353–366. doi:10.1016/j.cortex.2010.01.006
Schindler I, McIntosh RD, Cassidy TP, Birchall D, Benson V, Ietswaart M, Milner AD (2009) The disengage deficit in hemispatial neglect is restricted to between-object shifts and is abolished by prism adaptation. Exp Brain Res 192(3):499–510. doi:10.1007/s00221-008-1585-4
Schintu S, Pisella L, Jacobs S, Salemme R, Reilly KT, Farne A (2014) Prism adaptation in the healthy brain: the shift in line bisection judgments is long lasting and fluctuates. Neuropsychologia 53:165–170. doi:10.1016/j.neuropsychologia.2013.11.013
Serino A, Angeli V, Frassinetti F, Ladavas E (2006) Mechanisms underlying neglect recovery after prism adaptation. Neuropsychologia 44(7):1068–1078
Striemer CL, Danckert J (2007) Prism adaptation reduces the disengage deficit in right brain damage patients. Neuroreport 18(1):99–103
Striemer CL, Danckert J (2010a) Dissociating perceptual and motor effects of prism adaptation in spatial neglect. Neuroreport 21(6):436–441. doi:10.1097/WNR.0b013e328338592f
Striemer CL, Danckert J (2010b) Through a prism darkly: re-evaluating prisms and neglect. Trends Cogn Sci 14(7):308–316. doi:10.1016/j.tics.2010.04.001
Striemer CL, Danckert J (2013) The influence of prism adaptation on perceptual and motor components of neglect: a reply to saevarsson and kristjansson. Front Hum Neurosci 7:255. doi:10.3389/fnhum.2013.00255
Striemer CL, Sablatnig J, Danckert J (2006) Differential influences of prism adaptation on reflexive and voluntary covert attention. J Int Neuropsychol Soc 12(3):337–349
Striemer CL, Blangero A, Rossetti Y, Boisson D, Rode G, Vighetto A, Danckert J (2007) Deficits in peripheral visual attention in patients with optic ataxia. Neuroreport 18(11):1171–1175
Striemer CL, Locklin J, Blangero A, Rossetti Y, Pisella L, Danckert J (2009) Attention for action? Examining the link between attention and visuomotor control deficits in a patient with optic ataxia. Neuropsychologia 47(6):1491–1499. doi:10.1016/j.neuropsychologia.2008.12.021
Striemer CL, Ferber S, Danckert J (2013) Spatial working memory deficits represent a core challenge for rehabilitating neglect. Front Hum Neurosci 7:334. doi:10.3389/fnhum.2013.00334
Striemer CL, Russell K, Nath P (2016) Prism adaptation magnitude has differential influences on perceptual versus manual responses. Exp Brain Res 234(10):2761–2772. doi:10.1007/s00221-016-4678-5
Tilikete C, Rode G, Rossetti Y, Pichon J, Li L, Boisson D (2001) Prism adaptation to rightward optical deviation improves postural imbalance in left-hemiparetic patients. Curr Biol 11(7):524–528
Acknowledgements
This worked was funded through Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery, and Glenrose Rehabilitation Hospital Clinical Research Grants awarded to C.S. The authors would like to thank two anonymous reviewers for their helpful comments on an earlier version of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Striemer, C.L., Borza, C.A. Prism adaptation speeds reach initiation in the direction of the prism after-effect. Exp Brain Res 235, 3193–3206 (2017). https://doi.org/10.1007/s00221-017-5038-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-017-5038-9