
Abstract
The present study investigates the plasticity of 7-month-old infants’ orienting of attention during their perception of grasping actions. Previous research has shown that when infants observe a grasping hand, they shift their attention in line with the grasping direction, which is indicated by a reliable priming effect in this direction. The mechanisms behind this priming effect are largely unknown, and it is unclear how malleable this priming effect is with respect to a brief exposure to novel action-target contingencies. In a spatial-cueing paradigm, we presented a series of training trials prior to a series of test trials. These training sequences significantly modulated infants’ attention. This suggests that action perception, when assessed through shifts of attention, is not solely based on the infants’ grasping experience but quickly adapts to context-specific observed regularities.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Infants at an early stage of life already interpret observed actions as goal-directed (e.g. Woodward 2009). To a large extent, their ability to interpret the social environment is based on experiences made through everyday interactions. These experiences are acquired either through their own action performance (e.g. von Hofsten 2007) or through observed statistical regularities of actions performed by others (e.g. Wu et al. 2011). The present research examined whether infants’ perception of manual grasping actions is malleable through a brief observational experience.
Manual grasping is one of the most fundamental action skills acquired early in life because grasping provides the first opportunity to manipulate and reshape the environment (Bertenthal and Clifton 1998; Flanagan and Johansson 2002). A few days after birth, infants aim their arm movements towards interesting objects (von Hofsten 1982), and by the age of 5–7 months, grasping has become proficient enough that infants extrapolate object motions on linear paths and adapt their grasping movement accordingly (von Hofsten et al. 1998).
At around the same age, infants develop a remarkable sensitivity to grasping actions performed by others (e.g. Woodward 2009). This sensitivity is assessed using a variety of research designs and dependent measures tapping into a number of different cognitive processes involved in action perception (e.g. Gredebäck and Daum 2015). Some of these processes take place over a few hundred milliseconds (Gredebäck et al. 2010a). For example, overt shifts of attention (e.g. as found in predictive gaze shifts; Falck-Ytter et al. 2006; Gampe et al. 2015) or covert shifts of attention (Gredebäck et al. 2010b; Daum and Gredebäck 2011a) usually unfold within the first few hundred milliseconds after a directional action or gesture is observed (Gredebäck et al. 2010a). Other processes, for example as assessed via the measurement of looking times, extend over several seconds (Woodward 1998; Daum et al. 2012) .
A vast amount of research relied on measuring infants’ looking time in response to action outcomes that are in agreement with, or violate, their expectations—referred to here as action evaluation. Research into infants’ action evaluation has shown that 6-month-olds expect grasping actions to be directed towards goals (Woodward 1998), even if only initial parts of the action are presented (Daum et al. 2008; Brandone and Wellman 2009). Research techniques to measure action evaluation have recently been complemented with measures that target infants’ abilities to assess future states of perceived actions—which is referred to as action prediction (Gredebäck and Daum 2015; Gredebäck and Falck-Ytter 2015). Action prediction studies analyse the latency of predictive gaze shifts and have shown that infants start to predict the goal of observed grasping actions at between 6 and 12 months of age (Falck-Ytter et al. 2006; Hunnius and Bekkering 2010; Kanakogi and Itakura 2011; Cannon and Woodward 2012; Melzer et al. 2012).
In the present study, infants’ action perception was assessed in an action priming paradigm; that is, via the shifts of their attention during the observation of an action. Action priming is a very early component during the processing of observed actions. It occurs as soon as a social agent has been identified, and results in a shift of attention in the direction of the agent’s potential goal—even without an actual goal being visible. In contrast to both action evaluation and prediction, infants have less time and information available to process the relation between agent and goal. Accordingly, while a number of action evaluation (e.g. Woodward 1998) and action prediction studies (Gredebäck et al. 2009; Krogh-Jespersen et al. 2015) have shown that infants interpret grasping actions as being related to particular goals, action priming assesses an earlier processing phase where infants process the observed grasping action as being directed towards a particular location in space, potentially to interact with an object that is located at this position. This modulation of attention in the direction of others’ actions occurs within milliseconds and even before overt responses (such as eye movements) are observable. One way of assessing these early attentional processes caused by goal-directed agents is the application of a spatial-cueing paradigm (as introduced by Posner 1980). The logic behind this paradigm is that if an observer identifies the directionality of a symbolic cue such as an arrow (as used e.g. by Posner 1980) or a more socially relevant cue such as a grasping hand (e.g. Daum and Gredebäck 2011b), she will be faster at detecting a target that appears at a location that is congruent with the cue’s direction than an object at an incongruent location (this difference between reaction times, i.e. faster reaction times in congruent than in incongruent trials, is referred to hereinafter as priming effect). In infancy, priming effects have previously been reported during the perception of gaze shifts (Hood et al. 1998; Senju et al. 2006), grasping gestures (e.g. Daum and Gredebäck 2011b), and pointing gestures (Rohlfing et al. 2012; Daum et al. 2013; Bertenthal et al. 2014).
Previous research has shown that besides their sensitivity to social cues (e.g. gaze, pointing, grasping), infants evince priming effects caused by differences in visual properties of a stimulus (Jakobsen et al. 2013). Jakobsen et al. found that infants already shift their visual attention in the direction of an arrow, resulting in a significant priming effect. However, in this case the priming effect was caused by the perceptual properties of the arrow (i.e. that the head of the arrow is perceptually more salient than its end) and not by the conceptual meaning of the arrow, which is learned much later in life. Priming studies using more socially relevant cues such as pointing or a grasping hand (Daum and Gredebäck 2011a; Bertenthal et al. 2014) showed that the priming effect is not caused by the perceptual properties of the hand but rather by social content and an early step in a long chain of action perception processes (Gredebäck and Daum 2015).
Two priming studies that are of particular relevance for the present purpose have demonstrated a priming effect for grasping actions (Daum and Gredebäck 2011a; Wronski and Daum 2014). In both studies, infants were presented with a grasping hand (cue) followed by a toy (target) that appeared either in the direction of grasping (congruent trials) or in the opposite direction (incongruent trials). When the cue consisted of a static picture of a grasping hand, children at the age of 7 months showed a reliable priming effect (Daum and Gredebäck 2011a). This priming effect was already evident in 5-month-olds when the cue was a naturally moving grasping hand (Wronski and Daum 2014). Together, the two studies demonstrate that at the age of 7 months, infants shift their attention in the direction of the observed grasping action. This suggests that at the age of 7 months, infants perceive a grasping hand as being directed towards a location in space along the extension of the grasping action. Because this action priming occurs without any familiarisation during the experimental procedure, we refer to this robust effect as infants’ prior expectation about grasping hands and to the resulting priming effect as the default priming effect that occurs when 7-month-old infants observe a grasping hand.
The close temporal relation in development between perception and performance of grasping actions (for a recent study of the neural correlates of this close link between action priming and grasping proficiency, see Bakker et al. 2015) has led to the assumption that an observed action is mapped onto the observer’s motor representation of the action, which allows the observer to generate predictions about the future consequences of the observed action in the same way as for one’s own actions (Jeannerod 2001; e.g. Gallese 2009). The assumption that this mechanism is already functional early in life has received support from findings indicating that grasping perception is correlated with infants’ own productive grasping performance (Daum et al. 2011; Kanakogi and Itakura 2011; Melzer et al. 2012), that infants’ motor experience is causally related to their grasping perception and action priming (Sommerville et al. 2005; Bakker et al. 2016), and that the infant motor cortex is active during the perception and production of grasping actions (Nyström 2008; Southgate et al. 2009; Elsner et al. 2013). Accordingly, infants’ lifetime experience of grasping performance is an important source for their perception of others’ grasping actions.
However, research in social-cognitive development has shown that infants’ action perception is not only based on their action experience but is supported by alternative sources as well (e.g. Hunnius and Bekkering 2014). Infants interpret actions as being goal-directed actions which they are not yet able to perform when, for example, said actions are performed with tools they cannot yet use (Hofer et al. 2005; Biro and Leslie 2007; Boyer et al. 2011; Southgate and Begus 2013), when they are performed by inanimate agents such as balls (Gergely et al. 1995), toy figures (Luo and Baillargeon 2005), or when they are performed by schematic animals (Schlottmann and Ray 2010; Daum et al. 2012). In all cases, the observed behaviour cannot be mapped onto the infants’ motor repertoire and they need to rely on alternative interpretational mechanisms. While some authors explain the early goal attribution by means of inborn principles and biases, another potential candidate for this is learning through observed statistical regularities. This acquisition of knowledge through statistical learning is involved in a variety of developing skills such as categorisation (Mareschal et al. 2002) and word learning (Estes et al. 2007). With respect to the perception of goal-directed actions, previous research suggests that, around their first birthday, infants use statistical regularities for their goal predictions (Henrichs et al. 2013) and they also revise their initial predictions based on observed statistical regularities if they observe unsuccessful actions (Brandone et al. 2014). The implementation of statistical regularities of observed actions therefore plays an important role in infants’ interpretation of observed events.
The goal of the present study was to test the plasticity of infants’ action perception through observed statistical regularities. Previous research has shown that action observation does not substantially alter infants’ subsequent action perception (Sommerville et al. 2012; Cannon et al. 2012) or their action evaluation (Sommerville and Woodward 2005). But so far, no prior research has tested the plasticity of action perception through action priming in infants. When compared to action priming, action evaluation and prediction differ in the important respect that they require not only the identification of the directionality of a cue but also a more in-depth processing of the observed action in relation to an observable goal. In contrast, action priming merely involves the rapid orientation of attention in the direction of an observed action cue without the relation to an observable goal. We therefore tested the assumption that action priming is a more flexible process than action prediction and evaluation. To do so, we used the same laboratory set-up and paradigm as the two previous studies mentioned above that assessed priming effects in grasping actions (Daum and Gredebäck 2011a; Wronski and Daum 2014). In addition, infants were presented with a brief training sequence at the beginning of the experiment. In this training phase, the target consistently appeared either at the congruent or at the incongruent location. In a subsequent test phase, the infants saw a sequence of test trials in which congruent and incongruent trials were presented in a random order and equal distribution.
Based on the previous findings discussed above, two potential outcomes can be expected: first, if action priming is still a flexible process at the age of 7 months, a short training sequence is expected to affect infants’ attention. This effect of training should become evident, in particular, after incongruent training in which the relation between cue and target is novel, so where it is in conflict with the infants’ prior expectation about the directionality of a grasping hand. Second, and in contrast with the hypothesis formulated above, if action priming is largely rooted in the infants’ lifetime experience of grasping (which, by the age of 7 months, is about 4 months), a short sequence of incongruent trials is not expected to affect the infants’ prior expectation and the corresponding action priming. In this case, the different training sequences are not expected to modify the default priming effect.
Finally, in case a short training sequence does have an effect on the subsequent test trials, the question remains to which extent this effect remains is stable or will be affected by the additional information of the random distribution of congruent an incongruent trials during the test phase. To explore the stability of a potential training effect, we conducted an exploratory analysis of potential changes of the infants’ responses over the time course of the experimental test trials.
Experiment 1
In Experiment 1, 7-month-old infants were presented with a spatial-cueing paradigm (adapted from Daum and Gredebäck 2011a). The cue consisted of the static picture of a grasping hand followed by a target at a congruent or incongruent location. The experimental procedure included a training phase and a subsequent test phase. In the training phase, one group of infants received a series of congruent trials; another group of different infants received a series of incongruent trials. During the test phase, congruent and incongruent trials were presented in a random order and equal distribution.
Method
Participants
The final sample consisted of 48 infants aged 7 months (26 girls, 22 boys; mean age: 7 months; 3 days, range: 6;19–7;14). In addition, 26 infants aged 7 months (12 girls, 14 boys) were tested but not included in the final sample due to distress or restlessness (n = 3), technical problems (n = 1), or fewer than 6 valid training or test trials (n = 18). Contact information was obtained from public birth records. All infants were born full term (37- to 42-week gestation) and with normal birth weight (>2500 g). All parents gave informed consent before the study. The study was approved by the local ethics committee and was conducted in accordance with the Declaration of Helsinki.
Test environment, stimuli, and apparatus
The laboratory was unfurnished except for the test equipment. The stimuli were presented, and gaze was measured using a Tobii 1750 near-infrared eye tracker (Tobii Technology AB, Danderyd, Sweden) with an infant add-on (precision: 1°, accuracy: .5°, sampling rate: 50 Hz) and the software ClearView (version 2.7.1). A 9-point infant calibration was used. During calibration, a blue-and-white sphere expanded and contracted (extended diameter = 3.3°) in synchrony with a sound. Viewing distance was approximately 60 cm, and monitor diameter was 17 inch (24.8 × 20.7°). The infants were seated in a car safety seat (Maxi Cosi Cabrio) in front of the eye tracker.
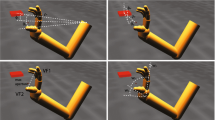
The stimulus presentation followed closely the one used in Daum and Gredebäck (2011a, Experiment 1), see Fig. 1. Each trial started with one of four looming toys (horizontal and vertical dimensions: maximum 4.5°, minimum 2.3°) presented at the centre of the monitor accompanied by a sound. As soon as the infant fixated the central stimulus, a hand (cue, 5.0° × 4.6°) was presented grasping towards either the left or the right for 1000 ms. The cue was followed by a renewed presentation of the initially presented toy (now referred to as target). The target appeared at a congruent location (in the same direction as the grasping hand, see Fig. 1, upper panel) or an incongruent location (in the opposite direction to the grasping hand, see Fig. 1, lower panel). The distance from the nearest edge of the cue to the target was 9.3°. The target remained visible until the infant looked at it for approximately 1000 ms or until 5000 ms had elapsed. Then, a new trial began with another centrally presented looming stimulus.

Stimulus sequences of an incongruent trial (upper panel) and a congruent trial (lower panel) in Experiment 1. Left Attention grabber (looming at 1 Hz with sound) presented until the infants fixated it; then, the trial was started. Middle Cue (grasping hand) is presented for 1000 ms. Right Target (same as attention grabber, looming with 1 Hz with sound) appears at a congruent or an incongruent location
The general paradigm consisted of a training phase and a subsequent test phase. In the training phase, two conditions were presented in a between-subjects design. In the congruent training condition, the targets always appeared at the grasping-congruent location, while in the incongruent training condition, the target always appeared at the grasping-incongruent location. In the test phase, the relation between grasping direction and the location of the target appearance was no longer predictable. The order of the targets as well as the relation between the direction of the grasping hand and the location of the target was pseudo-randomised. In order to avoid adaptation effects that might potentially override the specific manipulation of the training, no more than three repetitions of cue direction, target location, or cue–target relation were allowed. In the training phase, a total of 12 trials were presented to each infant. The maximum number of test trials presented was 64. Infants were randomly assigned to one of the two training conditions and to one of four different orders in the test phase.
Procedure
The infants were tested in the laboratory at a time of day when they were likely to be alert and in a good mood. All infants were tested individually with one parent present. Each participant and his/her parents were first escorted to a reception room. For approximately 10 min, the infant was allowed to explore the room, while the research assistant described the test procedure to the parents, and one of the parents signed a consent form. Then, the infant and one parent were brought to the test room. The research assistant helped the parent to position the infant in the car seat. During stimulus presentation, the parent sat on a chair behind the infant. Parents were instructed not to interact with their children during testing. They were encouraged, however, to put both hands symmetrically close to the child if it appeared necessary to comfort the infant. Once infant and parent seemed comfortable, the research assistant left the room and the stimulus presentation started.
Data analysis
For gaze analysis, three square areas of interest (AOI) were defined on the screen covering the cue (cue AOI; horizontal and vertical dimension: 7.5°) and the two target locations (target AOIs; horizontal and vertical dimension: 4.7°). A trial was considered valid if the infant fixated the central cue for at least 200 ms (Gredebäck et al. 2006) prior to making a gaze shift to the target. The saccadic reaction time (SRT) was defined as the difference in time between the appearance of the target and the arrival of the gaze at the respective target AOI (Gredebäck et al. 2010a). Individual SRTs less than 100 ms and greater than two standard deviations of each individual mean were excluded from analysis. Infants had to provide at least 6 training trials and 6 test trials (with a minimum of three trials per type [congruent, incongruent]) to be included in the final analysis. This criterion was based on previous studies where similar criteria were used as inclusion rates and similar minimum numbers of trials were achieved (Hood et al. 1998; Daum and Gredebäck 2011a; Rohlfing et al. 2012). Since we had no particular hypothesis regarding effects of sex and order of presentation, and preliminary analyses indicated no main effects of these two factors (all ps > .11), the data were collapsed for the remaining analysis over these factors.
The main analysis was performed in three steps: (1) the number of trials performed by each infant in the respective conditions was evaluated. (2) Mean SRTs were analysed separately for training and test trials. In the training trials, the mean SRTs were analysed using a univariate analysis of variance (ANOVA) with training (congruent, incongruent) as between-subjects variable. In the test trials, the mean SRTs in congruent and incongruent trials were analysed using a 2 × 2 (training [congruent, incongruent], trial type [congruent, incongruent]) repeated-measures analysis of variance (ANOVA), with trial as a within-subjects and training as a between-subjects variable. (3) SRTs were then compared for reliable priming effects per condition. To better compare the groups of infants in the two training conditions and to compare the results of the two experiments reported in this manuscript, the proportional SRT was calculated by dividing the SRT in congruent trials by the SRT in incongruent trials. A number smaller than 1 indicate the default priming effect (i.e. shorter SRTs in congruent than in incongruent trials); a number greater than 1 indicate a reversed priming effect (i.e. longer SRTs in congruent than in incongruent trials). The proportional SRTs were analysed using a one-way ANOVA with training as a between-subjects factor. The proportional SRTs of the different conditions were further analysed discretely by separate t tests for each training condition. (4) In an exploratory analysis, potential changes of the SRTs over trials were explored using linear regression analyses. The main results of both Experiments 1 and 2 are displayed in Fig. 2; p values are reported two-tailed throughout.

Proportional SRTs for the congruent (grey bars) and the incongruent (black bars) training conditions in Experiment 1 (left) and Experiment 2 (right). Values below 1 indicate a default priming effect, and values above 1 indicate a reversed priming effect (*p < .05)
Finally, to explore the stability of the potential training effect, changes of SRTs and proportional SRTs over the course of the experimental trials were explored via two additional analyses: first, separate linear regression analyses of the SRTs in congruent trials, incongruent trials, and over all trails, independent of condition, and second, an analysis of the proportional SRTs over the course of the experimental trials. For this analysis, we divided the 64 test trials into four quartiles, calculated the mean proportional SRT for each participant, and analysed these values using a 4 × 2 (quartile [Q1, Q2, Q3, Q4] × experimental condition [congruent training, incongruent training]) ANOVA with quartile as within-subjects and experimental condition as between-subjects factor. We want to emphasise that due to the decreasing number of valid trials over the course of the test phase (see the analysis of the number of trials in “Results” section), this analysis is of a merely exploratory nature and the results need to be considered with care.
Results
Number of trials
In the congruent training condition, infants contributed an average number of 10.4 training trials (SD = 1.8, range 6–12) and 34.5 test trials (SD = 16.2, range 8–60). In the incongruent training condition, infants contributed an average number of 10.2 training trials (SD = 2.1, range 6–12) and 25.5 test trials (SD = 13.2, range 8–55). The number of training trials contributed did not differ between conditions, t(46) = .38, p = .71, but infants performed more test trials in the congruent training condition than in the incongruent training condition, t(46) = 2.1, p = .04.Footnote 1 A linear regression analysis showed that the number of valid data points per trial (combined for congruent and incongruent trials) decreased over the course of the experiment, both during the congruent training condition, β = −.89, t(63) = −15.34, p < .001, from M(first 5 trials) = 18.2 data points to M(last 5 trials) = 5.4 data points, and the incongruent training condition, β = −.94, t(63) = −22.24, p < .001, from M = 17.2 data points to M = 2 data points.
Mean SRTs
No between-subjects effects of training condition were found, neither in the training trials, F < 1, nor in the test trials, F < 1. Furthermore, in the test trials, the ANOVA resulted in a significant interaction between training condition (congruent, incongruent) and trial (congruent, incongruent), F(1, 46) = 5.78, p < .05, η 2 = .11. After congruent training, SRTs were shorter in congruent than in incongruent trials (mean SRTs: congruent trials: M = 441.13 ms, SD = 110.60 ms; incongruent trials: M = 470.50 ms, SD = 123.33 ms), while after incongruent training the pattern was reversed and SRTs were shorter in incongruent than in congruent trials (mean SRTs: congruent trials: M = 468.62 ms, SD = 114.24 ms; incongruent trials: M = 445.68 ms, SD = 71.71 ms). This effect will be explored further in the next section.
Proportional SRTs
The one-way ANOVA on the proportional SRTs revealed a significant effect of training, F(1, 46) = 6.26, p < .05, η 2 = .12. The proportional SRTs differed significantly between the two groups, which indicates that the training sequence had a significant influence on the priming effect (see Fig. 2). As expected, infants in the congruent training condition showed a default priming effect, indicated by a proportional SRT that was significantly smaller than 1 (M = .94, SD = .12), t(23) = 2.35, p < .05. In contrast, infants in the incongruent training condition showed a proportional SRT that was numerically greater but not statistically different from 1 (M = 1.05, SD = .17), t(21) = 1.42, p = .17.
Exploratory analysis of SRTs over time
We first calculated the mean SRT per trial (for both conditions combined and separately for congruent and incongruent trials) and conducted a linear regression analysis over the course of the experiments (Bonferroni corrected for multiple (n = 6) comparisons, adjusted alpha level: p = .008). In the congruent training condition, the SRTs did not change over the course of the test trials (all βs < .09, all ts < .61, all ps > .54). In the incongruent training condition, we found that SRTs overall, β = .32, t(39) = 2.11, p = .04, tended to increase over trials, but this tendency did not survive the correction for multiple comparisons. Both, SRTs in congruent trials, β = .24, t(39) = 1.55, p = .13, and incongruent trials, β = .06, t(39) = .40, p = .69, did not change over the course of the trials.
Next, we analysed whether the proportional SRTs changed over the course of the test trials. This analysis revealed no significant main effect or interaction, all Fs < 1.95, all ps > .13. Because the number of missing trials was high, in particular in the third and fourth quartile, we additionally ran the same analysis with a set of data in which we replaced the missing values with the grand mean value calculated over both experimental conditions of Experiment 1, M(grand) = 1.00 (sic!). This analysis resulted in a trend towards a main effect of experimental condition, F(1, 46) = 3.43, p = .07, η 2 = .07, primarily reflecting the above-reported tendency that the proportional SRTs were larger in the incongruent training condition (Q1: M = 1.02, Q2: M = 1.03, Q3: M = 1.03; Q4: M = 1.03) than in the congruent training condition (Q1: M = .94, Q2: M = .95, Q3: M = 1.02; Q4: M = .98). No main effect of quartile was present, F < 1, nor was there an interaction between quartile and experimental condition, F < 1. In all, the difference between the two experimental conditions was stable across the course of the test trials of Experiment 1.
Discussion
Previous research has shown that with no additional training phase, 7-month-old infants shift their gaze earlier towards a target that appears in the direction of an observed grasping action (Daum and Gredebäck 2011a). The results of Experiment 1 demonstrate that this default priming effect, in which the children were assumed to show prior to the current experimental session, can be modified by a short sequence of incongruent training trials. From the observed statistical regularities during the training phase, the infants rapidly altered their prior expectation about the relation between cue and target. This modification of their expectation had a substantial impact on the priming effect, and this impact was maintained over the course of the experimental trials. In contrast, when presented with a series of congruent training trials, the infants continued to show the previously reported default priming effect (as reported in Daum and Gredebäck 2011a). We will come back to this aspect in “General discussion” section.
Two aspects of these results need further attention: the first aspect concerns the strength of the infants’ representation of the observed grasping action and the resulting expectation about the grasping direction. In Experiment 1, the stimulus material consisted of the static picture of a grasping hand. Research with adults has shown that static images of actions indeed result in premotor and motor cortex activity (e.g. Proverbio et al. 2009). However, it was also shown that dynamic actions trigger the respective motor representation more strongly (e.g. Buccino et al. 2001). Accordingly, the observation of a static hand might only have weakly triggered the infants’ motor representation of grasping. This could have resulted in a rather fragile default priming effect, which is strong enough to become evident without prior training but which can easily be modified by observed statistical regularities. The motor representation might be more strongly triggered by the observation of a more naturalistic grasping action. This stronger activation might then, in turn, hamper the rapid learning through observed statistical regularities, resulting in a smaller (or no) effect of the incongruent training on the infants’ prior expectations than in Experiment 1. In contrast, and as discussed in the introduction, learning is facilitated by the consistently observed violations of the infants’ expectation (Stahl and Feigenson 2015). In the present study, the rapid observational learning might have been facilitated by the consistently observed violations of the infants’ expectation during incongruent training. However, when presented with the picture of a static hand, the expectation about the grasping direction might be less strong when triggered by a static compared to a dynamic hand. Hence, this expectation might be more strongly violated when a more natural dynamic grasping action is observed than when only static action pictures are shown. In this case, a short incongruent training phase would enlarge the effect of the incongruent training effect as observed in Experiment 1.
The second aspect concerns the fact that in Experiment 1, the training resulted in a significant modification of the default priming effectFootnote 2 but did not result in a reversed priming effect. Hence, the incongruent training could just have puzzled the infant, resulting in a disappearance of the default priming effect, which could be caused by random gaze behaviour, but not in the appearance of a reversed priming effect. To replicate and extend the findings of Experiment 1, Experiment 2 tested the effect of training on infants’ perception of dynamic actions.
Experiment 2
To further test the plasticity of infants’ action perception, two groups of 7-month-old infants were presented with the same experimental paradigm as in Experiment 1 but with a dynamic grasping action serving as cue (adapted from Wronski and Daum 2014).
Method
Participants
The final sample consisted of 44 infants aged 7 months (21 girls, 23 boys; mean age: 7;1, range: 6;17–7;14). In addition, 21 infants (13 girls, 8 boys) were tested but not included in the final sample due to distress or restlessness (n = 2), technical problems (n = 1), or fewer than 6 valid training or test trials (n = 17). Recruitment was identical to Experiment 1.
Stimulus material and procedure
The procedure of Experiment 2 was identical to Experiment 1. The stimulus material (adapted from Wronski and Daum 2014, see Fig. 3) differed with respect to the following modifications. Instead of a static picture, the cue consisted of a video clip of a human hand performing a horizontal grasping movement. At the start of the trial, the hand was closed to a fist (4.5 × 3.9 visual degrees). It then performed a grasping movement to its maximum grasping aperture (6.1 × 5.4 visual degrees) from the starting position at the periphery of the screen towards the centre of the screen. No other parts of the wrist or arm were shown, to ensure equal distribution of visual information on the screen at the final position of the hand. Total duration of cue presentation was 2500 ms (500-ms still frame at the start and 2000-ms movement). After disappearance of the cue, a black screen was shown for 320 ms. This gap was included to decrease the likelihood that SRTs are biased by ongoing overt eye movements caused by the dynamic cue (e.g. Mitrani and Dimitrov 1978). Following this gap, the initial attention-grabbing stimulus measuring 3.2 × 4.3 visual degrees (referred to as target) appeared at a horizontal peripheral location on the screen at a distance of 11.9 visual degrees from the last position of the cue at the centre of the screen (Fig. 3).

Stimulus sequences of a congruent trial (upper panel) and an incongruent trial (lower panel) in Experiment 2. Left Attention grabber (looming at 1 Hz with sound) presented until the infants fixated it; then, the trial was started. Middle three: a closed fist is presented for 500 ms followed by a grasping movement towards the centre for 2000 ms and an empty screen for 320 ms. Right Target (same as attention grabber, looming with 1 Hz with sound) appears at a congruent or an incongruent location
Data analysis
Data analysis was identical to Experiment 1 except that the criterion for the validity of a trial was adjusted. A cue-start AOI was defined (11.7° × 11.6°) that extended from the starting position of the cue to the position of the cue after 1000 ms of cue presentation with a tolerance of 50 pixels around the maximal cue extension. The cue-end AOI (11.7° × 12.8°) was adjusted to be in line with Experiment 1 and was defined as the area around the last frame of the cue before disappearance with an additional surrounding tolerance of 50 pixels in each dimension.Footnote 3 A trial was considered valid when the infant had looked at the cue-start AOI for at least 200 ms during the first 1000 ms of the cue presentation and at the cue-end AOI for at least 200 ms during the last 500 ms of cue presentation. The main results of Experiment 2 are displayed in Fig. 2, and p values are reported two-tailed throughout.
Results
Number of trials
In the congruent training condition, infants contributed an average number of 7.3 training trials (SD = 1.2, range 6–9) and 13.8 test trials (SD = 9.4, range 6–37). In the incongruent training condition, infants contributed an average number of 8.6 training trials (SD = 2.1, range 6–12) and 19.4 test trials (SD = 9.6, range 6–37). The numbers of training trials contributed differed between conditions, t(42) = 2.48, p < .05, with more training trials contributed in the incongruent training condition. There was a marginal difference in the number of test trials contributed, t(42) = 1.97, p = .06, indicating that on average, the infants tended to perform fewer test trials in the congruent than in the incongruent training condition. A linear regression analysis showed that the number of valid data points per trial (combined for congruent and incongruent trials) decreased over the course of the experiment, both during the congruent training condition, β = −.83, t(63) = −11.47, p < .001, from M = 12.0 data points to M = 1.4 data points, and the incongruent training condition, β = −.91, t(63) = −16.51, p < .001, from M = 15.2 data points to M = .6 data points.
Mean SRTs
In the training trials, no between-subjects effect of training was found, F < 1. In the test trials, and in contrast to the training trials of Experiment 2 and in contrast to Experiment 1, we found a significant difference in the SRTs between the two training conditions, F(1, 42) = 15.35, p < .001, η 2 = .27. The mean SRTs in the congruent training condition (M = 512.0 ms, SD = 199.7 ms) were longer than in the incongruent training condition (M = 326.7 ms, SD = 64.8 ms). In light of the unexpected difference of the SRTs between the two training conditions, it is important to note that this difference was not yet present during training, where infants of both training groups provided similar SRTs (congruent training condition: M = 392.01 ms, SD = 177.22 ms; incongruent training condition: M = 349.95 ms, SD = 110.37 ms). Furthermore, and paralleling the findings of Experiment 1, the interaction between training condition and test trials was significant, F(1, 42) = 9.44, p < .01, η 2 = .18. As in Experiment 1, SRTs were shorter in congruent than in incongruent trials (mean SRTs: congruent trials: M = 423.07 ms, SD = 191.88 ms; incongruent trials: M = 514.87 ms, SD = 237.43 ms), while this was reversed after incongruent training, where SRTs were shorter in incongruent than in congruent trials (mean SRTs: congruent trials: M = 339.07 ms, SD = 65.16 ms; incongruent trials: M = 314.00 ms, SD = 69.46 ms). This effect will be explored further in the next section.
Proportional SRTs
The one-way ANOVA revealed a significant effect of training, F(1, 42) = 9.45, p < .01, η 2 = .18 (see Fig. 2). Further analyses revealed that infants in the congruent training condition showed a default priming effect, indicated by a proportional SRT smaller than 1 (M = .88, SD = .25), t(21) = 2.27, p < .05. In contrast, infants in the incongruent training condition showed a proportional SRT significantly greater than 1 (M = 1.08, SD = .19), t(21) = 2.10, p < .05, reflecting a reversed priming effect.
Exploratory analysis of SRTs over time
As in Experiment 1, we tested the extent to which SRTs changed over the course of the trials using linear regressions (Bonferroni corrected for multiple comparisons (n = 6), adjusted alpha level: p = .008). This analysis showed no change over trials in any of the conditions (all βs < .26, all ts < 1.64, all ps > .11).
The analysis of the proportional SRTs revealed no significant main effect or interaction, all Fs < 1.40, all ps > .27. To account for the large number of missing trials, we additionally ran the same analysis with a set of data in which we replaced the missing values with the grand mean value calculated over both experimental conditions of Experiment 2, M(grand) = 1.07. This analysis resulted in a significant main effect of experimental condition, F(1, 41) = 5.06, p < .05, η 2 = .11, reflecting the above-reported effect that the proportional SRTs were larger in the incongruent training condition (Q1: M = 1.14, Q2: M = 1.04, Q3: M = 1.23; Q4: M = 1.16) than in the congruent training condition (Q1: M = .90, Q2: M = 1.18, Q3: M = .90; Q4: M = 1.02). No main effect of quartile was present, F < 1. However, the interaction between quartile and experimental condition was significant, F(3, 126) = 3.22, p < .05, η 2 = .07. This reflects the finding that in contrast to the quartiles 1, 3, and 4, SRTs in quartile 2 were larger in the congruent training condition than in the incongruent training condition (see means above). In all, the exploratory analysis revealed that the difference between the two experimental conditions induced by the different training conditions was relatively stable across the course of the trials of Experiment 2.
Discussion
Experiment 2 replicates and extends the findings of Experiment 1 with more ecologically valid cues. Without any training, 7-month-old infants’ attention is modulated in the direction of an observed dynamic grasping action (Wronski and Daum 2014).Footnote 4 In the present study, a short incongruent training phase had a significant effect on infants’ attention, resulting in a reversed priming effect in the subsequent test phase. This further supports the assumption that at this age, infants rapidly learn relations between cues and targets from observed statistical regularities, and that these learned regularities can overwrite existing priming effects. The exploratory analysis of the time course of this effect shows that, similar to Experiment 1, the effect of the training was maintained over the course of the experimental trials.
In contrast to Experiment 1, we found a difference in the mean reaction times between the two training conditions. We will discuss this effect further in “General discussion”. Furthermore, infants watched more trials in the incongruent than in the congruent training condition, both in the training phase and in the test phase. This might reflect the effect that the infants in the incongruent training condition were generally more attentive. This may have been a factor for the reversed priming effect. However, the potential lower attention of the infants in the congruent training condition did not generally affect the infants’ attentional processes as indicated by the significant priming effect in the expected direction.
General discussion
In the present study, we applied a spatial-cueing paradigm to test the plasticity of 7-month-old infants’ perception of goal-directed grasping actions through action priming. Previous research showed that infants at this age shift their attention in the direction of an observed grasping action (Daum and Gredebäck 2011a; Bakker et al. 2015; Wronski and Daum 2014). This suggests that by this age they have developed a sensitivity to the goal-directedness of grasping actions and expect graspable objects to appear along the direction of the grasp. In the present study, we applied the same paradigm of these two mentioned studies and added a short training sequence at the beginning of the experimental session, in which the relation between cue and target was consistently either congruent or incongruent. This training differentially affected the infants’ action priming in the subsequent test phase. After congruent training, the infants continued to show the default priming effect. After incongruent training, by contrast, the priming effect was significantly reduced when the cue consisted of the static picture of a grasping hand (Experiment 1) and even reversed when the cue consisted of a dynamic grasping action (Experiment 2). An exploratory analysis of the infants’ reactions over the course of the experimental session additionally revealed that the difference in the proportional SRTs induced during the training phase of the different conditions did not decrease within the first few test trials, but remained stable across the experimental test phase. This suggests that infants’ action perception as measured through action priming is not solely grounded in their lifetime experience of their own grasping. In contrast, around halfway through their first year of life, infants’ perception of grasping actions is still flexible and malleable through short sequences of observations.
Previous studies investigating the impact of observational experience on action perception have shown that action perception seems to be relatively robust against observational experience. In particular, repeated exposure to the presentation of claws and nonfunctional hand–object interactions does not appear to alter infants’ action evaluation (Sommerville and Woodward 2005; Sommerville et al. 2012) or action prediction (Cannon et al. 2012), whereas repeated action experience substantially did. These two measures differ from action priming with respect to a significant aspect. Action evaluation and prediction require not only the identification of the directionality of a cue but a more in-depth processing of the observed action in relation to an observable goal. In contrast, action priming merely involves the rapid orientation of attention in the direction of an observed action without an observable goal to be processed.
In combination with previous studies, the present findings demonstrate that two aspects are involved in action perception. On the one hand, it is of great importance to quickly adjust one’s own perceptual and attentional system to contextual factors. The present findings show that this is already the case early in life. As a default, an observed grasping action primes the infant’s attention in the direction of grasping. However, at the age of 7 months, this action priming is still flexible and quickly adapts to observed regularities. On the other hand, it is important that our interpretation of the social world is not dramatically and rapidly modified by a few instances that are in contrast to core assumptions about others’ goal-directed actions. This becomes evident in the fact that infants rely on the profound and permanent knowledge of previous experience when they have to predict the actual goal of an observed action or to evaluate the observed goal with respect to their expectations. This profoundness becomes evident, for example, in the fact that infants need almost 200 days of experience of being fed before they predict the goal of a feeding action (Gredebäck and Melinder 2010). Furthermore, with respect to grasping, a 7-month-old’s experience is substantial; at around this age, alert infants spontaneously perform 100–250 hand movements every 10 min (e.g. Wallace and Whishaw 2003). The influence of this extensive grasping experience on their grasping perception is certainly not altered with a few instances of counter-intuitive observations.
One question that remains in the present study is how long the effect of the incongruent training on infants’ action expectations persists. Given the relative flexibility of infants’ attention, the statistical information about the distribution of congruent and incongruent trials during the test phase could be expected to reduce the influence of the training phase over time, at least in the incongruent training condition. The analysis over trials suggests that the change in the infants’ action expectations is subject to some stability because at least over the course of the current experiment, that is around 5–10 min, no significant reduction of the training effect was observed. However, this finding needs to be interpreted with great caution because the number of trials also significantly decreased over the course of the experimental session, which might have resulted in an increase of variance in the data towards the end that blurred potential changes of effect over time. Further research is needed to explore these temporal aspects in more detail.
To conclude, in combination with previous findings, the present findings suggest that social cognition and the way we perceive the social world need stability, which is provided by the infants’ lifetime action experience that serves as the basis for the prediction and the evaluation of actions. At the same time, social perception needs flexibility to quickly adapt to regularities of a current situation, which is provided by the plasticity of infants’ attentional processes for observed actions and the flexibility and quick adaptability of action priming. Thus, both stable high-level social-cognitive processes and flexible low-level social-perceptual processes govern infants’ perception and interpretation of social events.
Differential effect of training
At first glance, the training applied at the beginning of the experimental session could be expected to cause a modification of the infants’ attention independent of condition, resulting in a reversed priming effect after incongruent training and an even stronger default priming effect after congruent training. However, after congruent training, the priming effect was similar to that reported in previous studies without any training. Why was this the case? The two training conditions differed with respect to the matching of the prior expectation with the observation during the training. During the congruent training, infants’ observation was consistent with their prior expectation: a target appeared in the direction of a grasping hand. In contrast, during the incongruent training, the observation consistently violated this prior expectation. In adults, unexpected and or surprising events lead to an enhanced prediction error that in turn draws attention to allow reaction and learning (e.g. Friston et al. 2006). In infants, violations of expectations typically result in increased attention towards observed events (e.g. Baillargeon et al. 1985). Recent evidence even shows that like adults, infants also use “unexpectedness” as a cue for learning (Stahl and Feigenson 2015). In this particular study, infants were presented with events that did or did not correspond to their expectations. During subsequent presentation of the object involved in the violation of expectation, infants showed more effective learning (i.e. increased exploration of the object). The authors conclude that “early in life, expectancy violations offer a wedge into the problem of what to learn” (p. 91). Accordingly, the repeated observation of incongruent (i.e. unexpected) cue–target relations resulted in an increase of attention compared to the observation of a congruent cue–target relation. This increase of attention might have caused a more in-depth processing of the incongruent training events and a respective modification of the default priming effect in the incongruent training condition. The repeated violation of expectation indicated that something unusual was happening and conveyed the message that this novel information might be a relevant learning opportunity.
Differences in the SRTs between conditions in Experiment 2
The second aspect that needs further attention is the difference in the overall SRTs between the two conditions of Experiment 2. During the training phase of Experiment 2, the SRTs were comparable in both conditions. In contrast, the SRTs in the test phase were shorter after incongruent than after congruent training. A simple explanation of this effect is that it is caused by a simple sampling effect; infants in the congruent training condition were just generally slower than infants in the incongruent training condition. However, this explanation can be ruled out by the fact that the SRTs did not differ between conditions in the training phase. We therefore prefer an alternative explanation that is based on the violation of infants’ prior expectation in the incongruent training condition. This increase of attention mentioned above that was caused by the repeatedly observed incongruence during training might have persisted into the test phase, resulting in shorter reactions overall than after congruent training. In adults, increased levels of attention result in shorter reaction times (for an overview, see Einöther and Giesbrecht 2013) and in enhanced learning (e.g. Bernstein 1969). The assumption of increased attention in the incongruent training condition is further supported by the larger number of training and test trials performed here than in the congruent training condition in Experiment 2. Accordingly, during incongruent training, the repeated violation of expectation resulted in an increase of the infants’ attention, which persisted into the test phase and resulted in a decrease of attention that occurred at a later point in time in the incongruent than in the congruent training condition. In Experiment 1, this difference in overall SRTs was not found. It is likely that the dynamic grasping action in Experiment 2 was more naturalistic and triggered the infants’ prior expectation more strongly. As a result, the series of incongruent dynamic trials presented in Experiment 2 resulted in a stronger violation of expectation than the series of incongruent static trials in Experiment 1. Further research will elucidate whether this level of increased attention is merely based on a nonspecific behavioural arousal that increases the activity of the whole processing systems or whether the infants’ attention is specifically increased for the processing of the stimuli noticed during training.
To conclude, when 7-month-old infants observe a grasping action, they have a prior expectation about the direction of the perceived grasping action indicated by a reliable action priming effect. The findings of the present study demonstrate that at this age, action priming is still flexible and can be modified via a short sequence of observational training trials. This suggests that action perception is not solely grounded in the infants’ lifetime action experience but that action priming quickly adapts to observed regularities.
Notes
We tried hard to come up with an idea about the reason why infants completed more trials in the congruent than in the incongruent training condition. The fact that in Experiment 2, the number of completed trials was in an exact opposite pattern compared to Experiment 1 (i.e. more trials in the incongruent than in the congruent training condition) makes it difficult to even come up with a consistent speculation.
This assumption is supported by the comparison of the proportional SRT in the test phase of the incongruent training condition in the present study with the results of the 7-month-olds in Experiment 1 of Daum and Gredebäck (2011a), which yielded a significant difference between the proportional SRTs, t(40) = 3.20, p < .01. An additional analysis between studies indicated that the proportional SRTs differed neither between the test phase of the congruent training condition of the present Experiment 1 nor the results of Experiment 1 in Daum and Gredebäck (2011a), t(40) = 1.05, p = .30.
Note that the AOIs around the cue were larger in Experiment 2 than in Experiment 1. The reason for this was that in Experiment 2 the cue was moving and thus covered a larger area within a similar time window.
As in Experiment 1, we conducted a comparison between the proportional SRT of the test phase in the incongruent training condition of the present study and the results of the 7-month-olds in Experiment 1 by Wronski and Daum (2014). This analysis yielded a significant difference between the two experiments, t(38) = 2.70, p < .01. An additional analysis between studies indicated that the proportional SRTs did not differ between the test phase of the congruent training condition of the present Experiment 1 in the present study and the respective condition in Experiment 1 by Wronski and Daum (2014), t(38) = .14, p = 89.
References
Baillargeon R, Spelke ES, Wasserman S (1985) Object permanence in five-month-old infants. Cognition 20:191–208
Bakker M, Daum MM, Handl A, Gredebäck G (2015) Neural correlates of action perception at the onset of functional grasping. Soc Cogn Affect Neurosci 10(6):769–776. doi:10.1093/scan/nsu119
Bakker M, Sommerville JA, Gredebäck G (2016) Enhanced neural processing of goal-directed actions after active training in 4-month-old infants. J Cogn Neurosci 28:472–482. doi:10.1162/jocn_a_00909
Bernstein AS (1969) To what does the orienting response respond? Psychophysiology 6:338–350
Bertenthal BI, Clifton RK (1998) Perception and action. In: Kuhn D, Siegler RS (eds) Handbook of child psychology: Vol. 2. Cognition, perception, and language, 5th edn. Wiley, New York, pp 51–102
Bertenthal BI, Boyer TW, Harding S (2014) When do infants begin to follow a point? Dev Psychol 50:2036–2048. doi:10.1037/a0037152
Biro S, Leslie AM (2007) Infants’ perception of goal-directed actions: development through cue-based bootstrapping. Dev Sci 10:379–398. doi:10.1111/j.1467-7687.2006.00544.x
Boyer TW, Pan JS, Bertenthal BI (2011) Infants’ understanding of actions performed by mechanical devices. Cognition 121:1–11. doi:10.1016/j.cognition.2011.05.012
Brandone AC, Wellman HM (2009) You can’t always get what you want: infants understand failed goal-directed actions. Psychol Sci 20:85–91. doi:10.1111/j.1467-9280.2008.02246.x
Brandone AC, Horwitz SR, Aslin RN, Wellman HM (2014) Infants’ goal anticipation during failed and successful reaching actions. Dev Sci 17:23–34. doi:10.1111/desc.12095
Buccino G, Binkofski F, Fink GR et al (2001) Action observation activates premotor and parietal areas in a somatotopic manner: an fMRI study. Eur J Neurosci 13:400–404. doi:10.1111/j.1460-9568.2001.01385.x
Cannon EN, Woodward AL (2012) Infants generate goal-based action predictions. Dev Sci 15:292–298. doi:10.1111/j.1467-7687.2011.01127.x
Cannon EN, Woodward AL, Gredebäck G et al (2012) Action production influences 12-month-old infants’ attention to others’ actions. Dev Sci 15:35–42. doi:10.1111/j.1467-7687.2011.01095.x
Daum MM, Gredebäck G (2011a) The development of grasping comprehension in infancy: covert shifts of attention caused by referential actions. Exp Brain Res 208:297–307
Daum MM, Gredebäck G (2011b) Spatial cueing by referential human gestures, arrows and mechanical devices. Int J Mind Brain Cogn 2:113–126
Daum MM, Prinz W, Aschersleben G (2008) Encoding the goal of an object-directed but uncompleted reaching action in 6- and 9-month-old infants. Dev Sci 11:607–619
Daum MM, Prinz W, Aschersleben G (2011) Perception and production of object-related grasping in 6-month-olds. J Exp Child Psychol 108:810–818
Daum MM, Attig M, Gunawan R et al (2012) Action seen through babies’ eyes: a dissociation between looking time and predictive gaze. Front Cogn 3:370
Daum MM, Ulber J, Gredebäck G (2013) The development of pointing perception in infancy: effects of communicative verbal cues on covert shifts of attention. Dev Psychol 49:1898–1908. doi:10.1037/a0031111
Einöther SJL, Giesbrecht T (2013) Caffeine as an attention enhancer: reviewing existing assumptions. Psychopharmacology 225:251–274. doi:10.1007/s00213-012-2917-4
Elsner C, D’Ausilio A, Gredebäck G et al (2013) The motor cortex is causally related to predictive eye movements during action observation. Neuropsychologia 51:488–492
Estes K, Evans J, Alibali M, Saffran J (2007) Can infants map meaning to newly segmented words? Statistical segmentation and word learning. Psychol Sci 18:254–260
Falck-Ytter T, Gredebäck G, von Hofsten C (2006) Infants predict other people’s action goals. Nat Neurosci 9:878–879
Flanagan JR, Johansson RS (2002) Hand movements. In: Ramachandran VS (ed) Encyclopedia of the human brain. Academic Press, San Deigo, pp 399–414
Friston K, Kilner J, Harrison L (2006) A free energy principle for the brain. J Physiol-Paris 100:70–87. doi:10.1016/j.jphysparis.2006.10.001
Gallese V (2009) Motor abstraction: a neuroscientific account of how action goals and intentions are mapped and understood. Psychol Res 73:486–498. doi:10.1007/s00426-009-0232-4
Gampe A, Keitel A, Daum MM (2015) Intra-individual variability and continuity of action and perception measures in infants. Front Psychol Dev Psychol. doi:10.3389/fpsyg.2015.00327
Gergely G, Nadasdy Z, Csibra G, Biro S (1995) Taking the intentional stance at 12 months of age. Cognition 56:165–193
Gredebäck G, Falck-Ytter T (2015) Eye movements during action observation. Perspect Cogn Sci 10(5):591–598. doi:10.1177/1745691615589103
Gredebäck G, Daum MM (2015) The microstructure of action perception in infancy: decomposing the temporal structure of social information processing. Child Dev Perspect 9:79–83. doi:10.1111/cdep.12109
Gredebäck G, Melinder AMD (2010) Infants’ understanding of everyday social interactions: a dual process account. Cognition 114:197–206
Gredebäck G, Örnkloo H, von Hofsten C (2006) The development of reactive saccade latencies. Exp Brain Res 173:159–164
Gredebäck G, Stasiewicz D, Falck-Ytter T et al (2009) Action type and goal type modulate goal-directed gaze shifts in 14-month-old infants. Dev Psychol 45:1190–1194. doi:10.1037/a0015667
Gredebäck G, Johnson SP, von Hofsten C (2010a) Eye tracking in infancy research. Dev Neuropsychol 35:1–19
Gredebäck G, Melinder AMD, Daum MM (2010b) The neural basis and development of pointing comprehension. Soc Neurosci 5:441–450. doi:10.1080/17470910903523327
Henrichs I, Elsner C, Elsner B et al (2013) Goal certainty modulates infants’ goal-directed gaze shifts. Dev Psychol. doi:10.1037/a0032664
Hofer T, Hauf P, Aschersleben G (2005) Infants’ perception of goal-directed actions performed by a mechanical device. Infant Behav Dev 28:466–480
Hood BM, Willen JD, Driver J (1998) Adult’s eyes trigger shifts of visual attention in human infants. Psychol Sci 9:131–134
Hunnius S, Bekkering H (2010) The early development of object knowledge: a study of infants’ visual anticipations during action observation. Dev Psychol 46:446–454. doi:10.1037/a0016543
Hunnius S, Bekkering H (2014) What are you doing? How active and observational experience shape infants’ action understanding. Philos Trans R Soc Lond B Biol Sci 369:20130490. doi:10.1098/rstb.2013.0490
Jakobsen KV, Frick JE, Simpson EA (2013) Look here! The development of attentional orienting to symbolic cues. J Cogn Dev 14:229–249. doi:10.1080/15248372.2012.666772
Jeannerod M (2001) Neural simulation of action: a unifying mechanism for motor cognition. NeuroImage 14:S103–S109
Kanakogi Y, Itakura S (2011) Developmental correspondence between action prediction and motor ability in early infancy. Nat Commun 2:341. doi:10.1038/ncomms1342
Krogh-Jespersen S, Liberman Z, Woodward AL (2015) Think fast! The relationship between goal prediction speed and social competence in infants. Dev Sci 18:815–823. doi:10.1111/desc.12249
Luo Y, Baillargeon R (2005) Can a self-propelled box have a goal? Psychological reasoning in 5-month-old infants. Psychol Sci 16:601–608
Mareschal D, Quinn PC, French RM (2002) Asymmetric interference in 3- to 4-month-olds’ sequential category learning. Cogn Sci 26:377–389. doi:10.1016/S0364-0213(02)00062-9
Melzer A, Prinz W, Daum MM (2012) Production and observation of contralateral reaching: a close link by 12 months of age. Infant Behav Dev 35:570–579
Mitrani L, Dimitrov G (1978) Pursuit eye movements of a disappearing moving target. Vis Res 18:537–539
Nyström P (2008) The infant mirror neuron system studied with high density EEG. Soc Neurosci 3:334–347
Posner MI (1980) Orienting of attention. Q J Exp Psychol 32:3–25
Proverbio AM, Riva F, Zani A (2009) Observation of static pictures of dynamic actions enhances the activity of movement-related brain areas. PLoS ONE 4:e5389. doi:10.1371/journal.pone.0005389
Rohlfing KJ, Longo MR, Bertenthal BI (2012) Dynamic pointing triggers shifts of visual attention in young infants. Dev Sci 15:426–435. doi:10.1111/j.1467-7687.2012.01139.x
Schlottmann A, Ray E (2010) Goal attribution to schematic animals: do 6-month-olds perceive biological motion as animate? Dev Sci 13:1–10. doi:10.1111/j.1467-7687.2009.00854.x
Senju A, Johnson MH, Csibra G (2006) The development and neural basis of referential gaze perception. Soc Neurosci 1:220–234. doi:10.1080/1747091060098979
Sommerville JA, Woodward AL (2005) Pulling out the intentional structure of action: the relation between action processing and action production in infancy. Cognition 95:1–30
Sommerville JA, Woodward AL, Needham A (2005) Action experience alters 3-month-old infants’ perception of others’ actions. Cognition 96:B1–B11
Sommerville JA, Blumenthal EJ, Venema K, Sage KD (2012) The body in action: the impact of self-produced action on infants’ action perception and understanding. In: Slaughter V, Brownell C (eds) Early development of body representations. Cambridge University Press, Cambridge, pp 247–266
Southgate V, Begus K (2013) Motor activation during the prediction of nonexecutable actions in infants. Psychol Sci 24:828–835. doi:10.1177/0956797612459766
Southgate V, Johnson MH, Osborne T, Csibra G (2009) Predictive motor activation during action observation in human infants. Biol Lett 5:769–772
Stahl AE, Feigenson L (2015) Observing the unexpected enhances infants’ learning and exploration. Science 348:91–94. doi:10.1126/science.aaa3799
von Hofsten C (1982) Eye-hand coordination in newborns. Dev Psychol 18:450–461
von Hofsten C (2007) Action in development. Dev Sci 10:54–60
von Hofsten C, Vishton P, Spelke ES et al (1998) Predictive action in infancy: tracking and reaching for moving objects. Cognition 67:255–285
Wallace PS, Whishaw IQ (2003) Independent digit movements and precision grip patterns in 1–5-month-old human infants: hand-babbling, including vacuous then self-directed hand and digit movements, precedes targeted reaching. Neuropsychologia 41:1912–1918. doi:10.1016/S0028-3932(03)00128-3
Woodward AL (1998) Infants selectively encode the goal object of an actor’s reach. Cognition 69:1–34
Woodward AL (2009) Infants’ grasp of others’ intentions. Curr Dir Psychol Sci 18:53–57
Wronski C, Daum MM (2014) Spatial orienting following directional cues in infancy. Dev Psychol 50:2020–2029. doi:10.1037/a0037155
Wu R, Gopnik A, Richardson DC, Kirkham NZ (2011) Infants learn about objects from statistics and people. Dev Psychol 47:1220–1229. doi:10.1037/a0024023
Acknowledgments
We wish to thank Caterina Böttcher and Kerstin Träger for the acquisition of the participants. We owe special thanks to the infants and parents who participated in this study. This research was supported by ERC Starting Grant 312292 and the Knut and Alice Wallenberg Foundation (2012-0120) awarded to Gustaf Gredebäck.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Declaration of Helsinki and its later amendments or comparable ethical standards.
Rights and permissions
About this article

Cite this article
Daum, M.M., Wronski, C., Harms, A. et al. Action perception in infancy: the plasticity of 7-month-olds’ attention to grasping actions. Exp Brain Res 234, 2465–2478 (2016). https://doi.org/10.1007/s00221-016-4651-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-016-4651-3