
Abstract
This study investigated the extent to which specific interacting constraints of performance might increase or decrease the emergent complexity in a movement system, and whether this could affect the relationship between observed movement variability and the central nervous system’s capacity to adapt to perturbations during balancing. Fifty-two healthy volunteers performed eight trials where different performance constraints were manipulated: task difficulty (three levels) and visual biofeedback conditions (with and without the center of pressure (COP) displacement and a target displayed). Balance performance was assessed using COP-based measures: mean velocity magnitude (MVM) and bivariate variable error (BVE). To assess the complexity of COP, fuzzy entropy (FE) and detrended fluctuation analysis (DFA) were computed. ANOVAs showed that MVM and BVE increased when task difficulty increased. During biofeedback conditions, individuals showed higher MVM but lower BVE at the easiest level of task difficulty. Overall, higher FE and lower DFA values were observed when biofeedback was available. On the other hand, FE reduced and DFA increased as difficulty level increased, in the presence of biofeedback. However, when biofeedback was not available, the opposite trend in FE and DFA values was observed. Regardless of changes to task constraints and the variable investigated, balance performance was positively related to complexity in every condition. Data revealed how specificity of task constraints can result in an increase or decrease in complexity emerging in a neurobiological system during balance performance.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In humans, conceptualized as complex adaptive systems (Riley et al. 2012), movement variability is omnipresent due to the distinct constraints that shape each individual’s goal-directed behaviors (Davids et al. 2003). Movement variability has been studied as the natural variations that occur in motor performance across multiple repetitions of a task, reflecting changes in both space and time (Newell and Slifkin 1998; Stergiou et al. 2006).
In dynamical system theory, these variations have a functional role to drive adaptive behaviors in movement systems, allowing the central nervous system (CNS) to exploit the high dimensionality offered by the abundance of motor system degrees of freedom (DOF) (Davids et al. 2003). Adaptive behavior refers to a form of learning characterized by gradual improvement in performance in response to altered conditions (Krakauer and Mazzoni 2011). The relationship between variability and adaptive behavior will change depending on task constraints faced by each individual. Several studies have related movement variability to the capacity of the CNS to adapt behaviors to environmental changes (Davids et al. 2003, 2006; Renart and Machens 2014; Riley and Turvey 2002).
In order to observe motor behavior changes during adaptation, several studies have examined changes in the neuromuscular system analyzing postural control dynamics and their relationship with physiological complexity (Manor et al. 2010; Manor and Lipsitz 2013). This is because during postural control, the CNS regulates the activities of many neuromuscular components acting together in a complementary manner (Manor et al. 2010; Riley and Turvey 2002).
Previous analyses of the relationship between postural control and variability in movement coordination have examined two different global dimensions: the magnitude of observed variability and the structural dynamics of variability, addressed by analyzing its complexity (Stergiou et al. 2006). Complexity has been defined as the number of system components and coupling interactions among them (Newell and Vaillancourt 2001). Some researchers have indicated that complexity in different physiological processes can be observed through nonrandom fluctuations on multiple timescales in physiological dynamics (Costa et al. 2002; Lipsitz and Goldberger 1992; Manor et al. 2010). This second dimension provides additional information about properties of the dynamics of observed variability on multiples scales, which reveals important information on strategies used by the CNS during task performance (Caballero et al. 2014).
The complexity of center of pressure (COP) has been a prominent measure used for assessing the relationship between the complexity shown in a biological signal, and a neurobiological system’s capacity to adapt to perturbations in motor tasks such as postural control and balance (Decker et al. 2010; Goldberger et al. 2002b; Menayo et al. 2014).
This methodological prominence has emerged because it has been considered a collective variable, responsible for capturing postural organization and balance in individuals (Riley and Turvey 2002).
Data on balance performance have suggested that complexity in a biological signal may be related to the CNS’s capacity to reorganize degrees of freedom to adapt to perturbations (Barbado et al. 2012; Goldberger et al. 2002b). Adaptive movement responses have also been considered to exemplify functional exploratory behaviors, which reveal useful sources of information to perform and learn new skills (Stergiou et al. 2006). In this regard, less complexity in COP dynamics has been associated with less capacity to adapt (Barbado et al. 2012; Manor et al. 2010). Moreover, in some cases, the loss of complexity in COP dynamics has been related to disorders in the CNS (Cattaneo et al. 2015; Schmit et al. 2006).
However, the direction of this relationship remains somewhat unclear. Other studies of performance in balance tasks have reported data which do not support the aforementioned relationship, reporting greater complexity in fluctuations of COP associated with worse task performance (Duarte and Sternad 2008; Vaillancourt and Newell 2002). For example, in Duarte and Sternad’s (2008) study comparing young and elderly people, they found a higher degree of complexity in older people over an extended time (30 min) during performance in a standing balance task. This finding indicates that high levels of complexity could reflect a decreased adaptive capacity of CNS over longer timescales. Vaillancourt and Newell (2002, 2003) suggested that increases or decreases in the complexity of CNS behaviors can be functional, but may be dependent on the nature of both the intrinsic dynamics of the system and the task constraints that need to be satisfied. Due to specific performance constraints encountered, there may be a reduction in the number of configurations available to a dynamical system through a restructuring of the state space of all possible configurations available (Davids et al. 2003; Newell and Vaillancourt 2001). Here, we sought to understand the extent to which specific interacting constraints of performance might lead to an increase or decrease in emergent complexity in a movement system, during task performance.
Another important question concerns whether the “controversy” surrounding the relationship between observed movement variability and the capacity to adapt to unexpected perturbations may actually be due to the specific experimental procedures of analysis selected to address complexity (Goldberger et al. 2002b; Stergiou et al. 2006). For instance, it has been suggested that entropy measures which analyze the regularity of a signal do not measure the complexity of system dynamics (Goldberger et al. 2002b). These studies did not consider whether signal regularity was clearly related to the complexity of system dynamics. Instead, it may be more appropriate to use fractal measures or long-range autocorrelation analysis, such as detrended fluctuation analysis (DFA), to investigate complexity in complex adaptive systems. Regardless, several studies have shown the utility of entropy measures in interpreting the randomness in experimental data from physiological systems in relation to postural control (Barbado et al. 2012; Donker et al. 2007; Menayo et al. 2014), heart rate (Lake et al. 2002; Wilkins et al. 2009), neuromotor control of movements early in life (Smith et al. 2011), mental fatigue (Liu et al. 2010), intracranial pressure (Hornero et al. 2005) or local muscle fatigue (Xie et al. 2010).
Up to now, the literature seems to support the view that motor variability is related to adaptive capacity, but the direction of the relationship seems to be unclear, possibly for different reasons, including: (1) the role that specific task constraints may play in shaping emergent behaviors; and (2) the difficulty in choosing the most appropriate tool to measure and address complexity in motor behavior. Addressing possible reasons for this methodological controversy behind the relationship between movement variability and adaptive capacity, we sought to understand whether manipulation of task constraints would result in a modification of participant performance strategies, due to the emergence of novel exploratory behaviors captured by the reorganization of motor system degrees of freedom to adapt to challenging performance situations. In this regard, we analyzed emergent movement adaptations under varying task constraints. We also used different nonlinear tools to measure the complexity of observed system variability. We hypothesized that increases or decreases in the complexity of a behavior depend on the nature of the task constraints to be satisfied. In particular, we expected that increasing difficulty and availability of biofeedback would lead to a reduction in the number of configurations available in the motor system, causing a loss of complexity and performance decrements.
Methods
Participants
Fifty-two healthy volunteers (13 women) took part in this study [age = 25.5 (6.01) years, height = 1.70 (0.25) m, mass = 70.66 (10.33) kg]. They had no previous experience in the balance task used in this study.
Written informed consent was obtained from each participant prior to testing. The experimental procedures used in this study were in accordance with the Declaration of Helsinki and were approved by a University Office for Research Ethics.
Experimental procedure and data collection
To assess COP fluctuation, ground reaction forces were recorded at 1000 Hz on a Kistler 9287BA force platform.
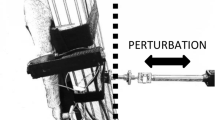
The task required the participants to stand on a wooden platform (0.50 m × 0.50 m) and perform eight trials of 70 s each, with 1-min rest periods between trials. Standing stability and availability of visual biofeedback were manipulated. The decision to manipulate these two different task constraints was taken because both are heavily used in the literature to analyze and train postural control. In particular, the use of biofeedback was chosen to control “error sensitivity.” According to Herzfeld and Shadmehr (2014, p. 149) “when we make a movement and experience an error, on the next attempt our brain updates motor commands to compensate for some fraction of the error,” and this error sensitivity term varies substantially from individual to individual and from task to task. Thus, error sensitivity remains constant for all participants. Two of the eight trials were performed on a solid floor (stable condition or SC). The other six were performed on an unstable platform (unstable condition or UC). All trials were performed under four different levels of difficulty, defined by the stability of the base of support. To achieve this aim, a wooden platform (0.02 m thick) was affixed to the flat surface of three polyester resin hemispheres with the same height (0.1 m) and different diameters: UC1 = 0.50 m of diameter; UC2 = 0.40 m of diameter; and UC3 = 0.30 m of diameter (Fig. 1). Each condition was experienced under two different visual biofeedback conditions: (a) without visual biofeedback, where the representation of COP displacement was not displayed. Here, the instruction to participants was to stay “as still as possible” (Duarte and Sternad 2008); and (b) with visual biofeedback, where COP displacement, besides a static center target (0.003 m of diameter on the base of support and 0.05 m projected on the wall in front of the participant; scale displays: 16.6–1), was displayed in real time. Participants were instructed to keep their COP on the target (Fig. 1).

Schematic illustration of the protocol distribution and the different surfaces used: a stable platform, b UC1: unstable platform with 50 cm of diameter, c UC2: unstable platform with 40 cm of diameter, d UC3: unstable platform with 30 cm of diameter
Data analysis and reduction
An application under Labview 2009 (Mathworks, Natick MA, USA), developed in our laboratory, was used to perform the data analysis. COP time series were previously downsampled from 1000 to 20 Hz due to: (1) there being little of physiological significance above 10 Hz in the COP signal (Borg and Laxåback 2010), and suggestions to use sampling frequencies close to COP dynamics (Caballero et al. 2013); and (2) signal oversampling possibly leading to artificial colinearities, affecting the variability data (Rhea et al. 2011). The first 5 s and last 5 s of each trial were discarded to avoid nonstationarity related to trial initiation (van Dieën et al. 2010). Time series length was 1200 data points. It has to be taking in account that one time series were shorter than 1200 data points (590 data points) due to the fact that two participants were unbalanced before 70 s. We computed the time series data before these failures. That result was included in the analysis because it did not show outlier values in any of the assessed variables. Two filtering processes were used to analyze different postural control behaviors that are related to two different components of COP displacement: rambling and trembling (Zatsiorsky and Duarte 1999). The first is defined as the motion of a moving reference point with respect to which the body’s equilibrium is instantly maintained and characterized by large amplitudes at low frequencies. This component could be related to central control (Tahayori et al. 2012). Thus, we used a low-pass filter (fourth-order, zero-phase-lag, Butterworth, 5 Hz cutoff frequency) (Lin et al. 2008) to assess it. The trembling component is defined as the oscillation of COP around a reference point trajectory, being characterized by short amplitudes at high frequencies (Zatsiorsky and Duarte 1999). This component could be related to peripheral control (Tahayori et al. 2012). Hence, we used a high-pass filter (fourth-order, zero-phase-lag, Butterworth, 10 Hz cutoff frequency), similar to that used by Manor et al. (2010).
Postural sway was assessed using traditional bivariate COP-based measures combining the anterior–posterior (AP) and medial–lateral (ML) displacement trajectories: bivariate variable error (BVE) and mean velocity magnitude (MVM). These variables were used to assess task performance and were calculated over the signal, filtered using a low-pass filter. We used just the filtered signal using a low-pass filter because static balance is characterized by small amounts of postural sway which is analyzed at low frequencies.
BVE was measured as the average value of the absolute distance to each participant’s own midpoint (Eq. 1) (Hancock et al. 1995; Prieto et al. 1996).
where N is the number of data points in the COP displacement time series and i is each successive data point.
MVM was measured as the average velocity of COP (Eq. 2) (Prieto et al. 1996).
where T is the trial duration (60 s).
The variables used to assess the complexity of COP were fuzzy entropy (FE) and detrended fluctuation analysis (DFA). These variables were calculated after both were filtered and processed (low-pass and high-pass filters). The variables were calculated over the resultant distance (RD) COP time series (Fig. 2), instead of the AP and ML time series, due to the fact that the orientation of the base of support is only approximately aligned with the axes of the force platform, especially in unstable situations (Prieto et al. 1996). Thus, measures based on the AP time series probably reflect some ML movements of the participant, and vice versa, while the RD vector is not sensitive to the orientation of the base of support with respect to the force platform (Prieto et al. 1996; Roerdink et al. 2011). RD is the vector distance from the center of the posturogram to each pair of points in the AP and ML time series (Eq. 3).

An example of the COP resultant magnitude time series over 60 s for a participant. SC, stable condition; UC1, unstable platform with 50 cm of diameter; UC2, unstable platform with 40 cm of diameter; UC3, unstable platform with 30 cm of diameter; α, autocorrelation values obtained by DFA
FE typically returns values that indicate the degree of irregularity in the signal. This measure computes the repeatability of vectors of length m and m + 1 that repeat within a tolerance range of r of the standard deviation of the time series (equations from 4 to 12). Higher values of FE thus represent lower repeatability of vectors of length m to that of m + 1, marking a greater irregularity in the time domain of the signal. Lower values represent a greater repeatability of vectors of length m + 1 and are, thus, a marker of lower irregularity in signal output. To calculate this measure, we used the following parameter values: vector length, m = 2; tolerance window, r = 0.2 × SD; and gradient, n = 2. In previous research, these parameter values have shown high levels of consistency, which underlies their frequent use (Chen et al. 2007). FE was calculated according to the procedures of Chen et al. (2007). We also conducted analyses of other related complexity measures, such as Sample entropyFootnote 1. However, we chose FE because it displays some advantages, such as a stronger relative consistency, less dependency on data length, free parameter selection and more robustness to noise (Chen et al. 2009; Xie et al. 2010).
DFA represents a modification of classic root-mean-square analysis with random walk to evaluate the presence of long-term correlations within a time series using a parameter referred to as the scaling index, α (Bashan et al. 2008; Peng et al. 1995). The scaling index α corresponds to a statistical dependence between fluctuations at one timescale and those over multiple timescales (Decker et al. 2010). This procedure estimates the fractal scaling properties of a time series (Duarte and Sternad 2008) and has also been used to describe the complexity of a process (Goldberger et al. 2002a).
This measure was computed according to the procedures of Peng et al. (1995). In this study, the slope α was obtained from the window range 4 ≤ n ≤ N/10 to maximize the long-range correlations and reduce errors incurred by estimating α (Chen et al. 2002). Different values of α indicate the following: α > 0.5 implies persistence in position (the trajectory tends to remain in its current direction); α < 0.5 implies anti-persistence in position (the trajectory tends to return from where it came) (Roerdink et al. 2006).
Statistical analysis
Normality of the variables was evaluated using the Kolmogorov–Smirnov test with the Lilliefors correction. Mixed repeated-measures ANOVA with two intra-individual factors, task difficulty level and biofeedback availability, was used to assess effects of both factors on performance outcome measures and complexity variables. Outcomes of the ANOVAs were considered to be statistically significant when there was a <5 % chance of making a type I error (p < 0.05). Bonferroni adjustment for multiple comparisons was performed to ascertain differences between task performances under different constraints according to each intra-individual factor. Partial eta squared \((\eta_{\varvec{p}}^{2} )\) was calculated as a measure of effect size and to provide a proportion of the overall variance that is attributable to the factor. Values of effect size ≥0.64 were considered strong, around 0.25 were considered moderate and ≤0.04 were considered small (Ferguson 2009).
Finally, the Pearson product moment correlation coefficients were calculated to assess relationships between performance variables (BVE and VMM) and complexity measures (FE and DFA).
Results
Mean values obtained under each balance condition and pairwise comparisons between difficulty conditions and biofeedback conditions are displayed in Table 1.
MVM showed higher values in biofeedback condition (F 1, 51 = 74.876; p < 0.001; \(\eta_{\varvec{p}}^{2}\) = 0.595). In contrast, despite BVE not revealing overall differences between biofeedback availability conditions (F 1, 51 = 2.637; p = 0.111; \(\eta_{\varvec{p}}^{2}\) = 0.049), at lower levels of difficulty, lower values of BVE were observed in the biofeedback condition (Fig. 3). BVE differences observed between biofeedback conditions did decrease as task difficulty level increased, and even disappeared at the most difficult performance levels. Additionally, both performance variables displayed higher values when task difficulty increased, being significantly different between conditions (BVE: F 1.83, 93.36 = 374.305; p < 0.001; \(\eta_{\varvec{p}}^{2}\) = 0.880; MVM: F 1.89, 96.6 = 491.241; p < 0.001; \(\eta_{\varvec{p}}^{2}\) = 0.906) (Fig. 3).

Pairwise comparisons between difficulty levels and biofeedback conditions in performance variables. a, significant differences between biofeedback conditions; 1, significant differences according to SC; 2, significant differences according to UC1; 3, significant differences according to UC2; 4, significant differences according to UC3
With regard to complexity variables, in the low-pass filtered signal, higher FE (F 1, 51 = 77.660; p < 0.001; \(\eta_{\varvec{p}}^{2}\) = 0.604) and lower DFA values (F 1, 51 = 65.392; p < 0.001; \(\eta_{\varvec{p}}^{2}\) = 0.562) were observed when biofeedback was available. However, differences in these dependent measures decreased as task difficult level were increased (Fig. 4). Regarding the high-pass filtered signal, the presence of biofeedback did not display effects on any complexity variable (FE: F 1, 51 = 3.949; p = 0.052; \(\eta_{\varvec{p}}^{2}\) = 0.072; DFA: F 1, 51 = 1.744; p = −192; \(\eta_{\varvec{p}}^{2}\) = 0.033).

Pairwise comparisons between difficulty levels and biofeedback conditions in complexity variables. a, significant differences between biofeedback conditions; 1, significant differences according to SC; 2, significant differences according to UC1; 3, significant differences according to UC2; 4, significant differences according to UC3
Complexity values at different task difficulty levels varied according to the filter used, the biofeedback condition and the variable recorded (Fig. 4). When variables were calculated over the low-pass filtered signal, in the presence of biofeedback, FE values were significantly different between SC and UC3 and between UC3 and UC1, decreasing as difficulty increased. However, without biofeedback, FE increased with task difficulty, displaying significant differences in the value between SC and every UC condition. Regarding DFA in the conditions with biofeedback, significant differences were observed between UC1 and UC3 and between UC2 and UC3, reaching the highest values at the most difficult task level. Without biofeedback, DFA values decreased from SC to UC2 and UC3, and from UC1 to UC2, attaining the highest values at the least difficult task level.
On the other hand, when complexity variables were calculated with the high-pass filtered signal, FE decreased and DFA increased as task difficulty increased regardless of the availability of biofeedback. So, in most of the conditions, dependent variables showed significant differences between levels of task difficulty, but differences between biofeedback conditions were only found with low-pass filtered signals.
Performance variables (BVE and MVM) were positively correlated, but showed an inverse correlation with complexity variables. Furthermore, the degree of dependence between them varied according to the filter used and biofeedback availability. When the low-pass filtered signal was used (Table 2), and in conditions without biofeedback, BVE was negatively correlated with FE and positively correlated with DFA. Nevertheless, in conditions with biofeedback, this correlation was only found at the highest task difficulty level. MVM showed positive correlation with FE and negative correlation with DFA despite the availability of biofeedback. Additionally, FE and DFA variables displayed an inverse relationship in every condition.
When the high-pass filter was used (Table 3), BVE was negatively correlated with FE, only in the most difficult task condition regardless of the availability of biofeedback. A positive correlation between BVE and DFA was found when biofeedback was available, only at the lowest and highest task difficulty levels, but no correlation between them was found in conditions without biofeedback. With regard to MVM, this variable was negatively correlated with FE in all of the unstable conditions (with or without biofeedback). MVM was positively correlated with DFA only in the stable condition when the biofeedback was available. In the condition without biofeedback, this correlation was observed in UC1 and UC2.
Discussion
Recently, it has been argued that an increase or decrease in the complexity of a behavioral or physiological system depends on interactions between system intrinsic dynamics and performance task constraints (Vaillancourt and Newell 2002, 2003). In this experiment, we investigated the complexity of movement system variability during performance of different balance tasks, observing that participants modified their postural control dynamics according to task difficulty and availability of biofeedback. In addition, regardless of these changes to task constraints, performance was positively related to complexity.
Performance decreased when balance task difficulty was increased as reported in previous research (Barbado et al. 2012; Borg and Laxåback 2010). Values in performance measures, both in BVE and MVM, increased as task difficulty level increased (Fig. 3). However, availability of biofeedback had different effects on BVE and MVM values. With biofeedback, BVE values decreased significantly, but only at lower task difficulty levels. However, as difficulty level was increased, biofeedback availability did not influence the amount of variability observed in COP measures. In stable or less challenging unstable task conditions, different locations of the COP on the surface of support allowed a participant to maintain stability (Caballero et al. 2014). However, increasing task difficulty limited the region of stability, signifying that in the difficult balancing conditions, there were a limited number of COP locations where system stability could be maintained (Lee and Granata 2008). Under more stable balancing conditions, visual biofeedback was used to maintain COP location on the target. Under more challenging postural control conditions, visual biofeedback information might have been redundant, because participants did not have many COP locations where they could maintain system stability. They only had possible outcome solution: the same as displayed by the available biofeedback signal. From a dynamical systems’ viewpoint, differences between biofeedback conditions could be interpreted as the existence of different types of attractors in a performance landscape. It seems that participants used a behavior similar to a fixed-point attractor when biofeedback was available, characterized by a fixed point in state space where no movement is observed (van Emmerik and van Wegen 2000). Nevertheless, participants explored the oscillatory COP dynamics (Vaillancourt and Newell 2003) without biofeedback in the least challenging conditions. Availability of biofeedback seemed to change postural control strategies by decreasing the number of configurations available to a dynamical movement system (Davids et al. 2003). In this regard, available information seemed to constrain the system to one area of the attractor landscape in this task.
On the other hand, MVM values displayed an increase in biofeedback conditions compared to when biofeedback was not available. Although there are a greater number of COP locations where stability can be maintained, this increase in MVM could be due to the fact that under the less challenging task constraints, visual biofeedback drives the system to one specific location. Without biofeedback, participants focused on avoiding falling. In the conditions with biofeedback, they tried to adjust their COP to the target, performing a greater number of adjustments. The increased values of MVM in biofeedback situations can also be related to an increased error sensitivity of the individuals regulated by the CNS (Herzfeld and Shadmehr 2014). In this sense, MVM could be an index of the amount of corrections needed to adjust the COP location, increasing neuromuscular effort and resulting from participant exploratory behaviors. Higher COP velocity would be an index of exploratory behaviors in discovering stable performance solutions under relatively novel task constraints (Davids et al. 1999).
According to previous studies, COP analysis has revealed two different postural control mechanisms: rambling and trembling (Mochizuki et al. 2006; Tahayori et al. 2012). These two processes may reflect changes in the body reference configuration and changes in the properties of the mechanical and neural structures implementing the supraspinal control signals (Danna-Dos-Santos et al. 2008). Observed variability of low-pass filtered COP, related to volitional control (rambling component), showed a higher degree of irregularity and less long-range autocorrelation when biofeedback was available. The changes in these variables, influenced by biofeedback, might indicate that the existence or not of this task constraint drives the system to different kinds of behaviors. The system would transit to a state space, displaying lower values of complexity without biofeedback (similar to oscillatory dynamic), and a behavior related to a fixed-point attractor in conditions with feedback, revealing more complexity in COP behaviors (van Emmerik and van Wegen 2000). Taking into account the effect of difficulty level, when biofeedback was available, the degree of irregularity of low-pass filtered COP decreased as task difficulty increased, whereas the long-range autocorrelation values increased. However, under task constraints when biofeedback was not available, the trend for FE and DFA values was inverted. Moreover, as task difficulty levels increased, clearly the difference between biofeedback conditions was reduced. This finding reflects again the redundancy of biofeedback in these more challenging conditions, where COP locations compatible with maintaining system stability are reduced. Unlike the findings of Manor et al. (2010) which support the role of complexity of fluctuations related to peripheral adjustments in postural control when standing, our results seem to indicate that complexity is more related to volitional changes in COP dynamics, reflecting a search strategy in participants to cope with task constraints which do not necessarily require an involvement of a greater number of DOF. According to Danna-Dos-Santos et al. (2008), this search strategy could be reflected by the rambling component. These findings are supported by Newell and Vaillancourt (2001) who suggested that the increase or the decrease in complexity can be independent of the number of component mechanical degrees of freedom being harnessed as a system, but the direction of the changes in complexity is driven by task constraints.
These contrasting results could have emerged for different reasons. First, it is possible that the balance task constraints used in both studies were different. Thus, the type of control requirements for keeping balance could have differed. Another reason could be due the populations studied. Manor et al. (2010) studied COP complexity in people with risk factors for falls for whom peripheral control could be a key factor in avoiding falls, while the participants of our study were healthy people with little risk of falling. Nevertheless, it is difficult to compare the results of the two studies because Manor et al. (2010) did not analyze low-pass COP signals. In future studies, it would be interesting to assess both kind of components of COP displacement and changes in COP complexity in relation to distinct task constraints and with different populations.
Regarding the high-pass filtered COP signal, the availability of biofeedback did not affect system complexity, but task difficulty did, showing a decrease in irregularity and an increase in long-range autocorrelation as task difficulty increased. Taking into account that this filter procedure could reflect peripheral postural control (trembling component), this lack of effect of the biofeedback condition could be due to the fact that the fluctuations of the trembling component represent an involuntary adjustment of COP (Danna-Dos-Santos et al. 2008; Tahayori et al. 2012). On the other hand, the fact that the most difficult conditions revealed less irregularity and greater long-range autocorrelation of the COP signal could indicate that, in these situations, individuals reduced the number of involuntary adjustments due to the difficulty in correcting COP displacement because of the increase in inertia.
Regarding correlational analysis, a direct relationship between BVE and complexity was found in both low-pass and (to lesser extent) high-pass filtered COP signals. These results seem to indicate that participants who showed lower balance performance exhibit a lower number of postural adjustments. Conversely, MVM was directly related to complexity in the low-pass filtered COP signal and, inversely, to complexity in the high-pass filtered COP signal. This finding could mean that individuals who displayed low COP velocities showed a higher number of peripheral postural adjustments and a low number of volitional corrections. Additionally, when participants showed higher COP velocities, it could mean that the peripheral system could not control stability and more volitional postural corrections were needed to maintain balance.
The fact that the relationships between balance performance variables and complexity were stronger in the low-pass filtered COP, revealed the prevalence of volitional adjustments in postural control to maintain balance. Peripheral adjustments played a less relevant role in the postural control strategy during the balance tasks analyzed in this study.
Our results indicated that a specific relationship that emerges between system complexity and performance is dependent on task constraints (Newell and Vaillancourt 2001; Vaillancourt and Newell 2002, 2003; Vaillancourt et al. 2004). It seems that each performance variable varied according to different task constraints encountered by participants, revealing different trends. These findings signified that when researchers wish to assess the relationship between an individual’s capacity to adapt and system complexity when learning or under different performance constraints, contradictory results may be observed due to the influence of distinct task constraints designed into experiments. Furthermore, this is a very important point to take into account when the system complexity is related to system constraints of aging, illness or damage.
To conclude, in this study, we provided some support for the idea that specific task constraints can lead to an increase or decrease in complexity emerging in a neurobiological system during performance. Informational constraints, such as availability of biofeedback and level of task difficulty, shaped emergent strategies of movement coordination, due to participants searching for different attractors to functionally regulate their behaviors.
Notes
Sample entropy was also calculated as another entropy measure to assess the degree of irregularity of CoP values. To calculate this measure, we used the following parameter values: vector length, m = 2; tolerance window, r = 0.2 * SD (Pincus, 1991). The results were very similar to the FE results, in both the effect of the different constraints and the correlation between performance and complexity.
References
Barbado D, Sabido R, Vera-Garcia FJ, Gusi N, Moreno FJ (2012) Effect of increasing difficulty in standing balance tasks with visual feedback on postural sway and EMG: complexity and performance. Hum Mov Sci 31:1224–1237
Bashan A, Bartsch R, Kantelhardt JW, Havlin S (2008) Comparison of detrending methods for fluctuation analysis. Phys A 387:5080–5090
Borg FG, Laxaback G (2010) Entropy of balance: some recent results. J NeuroEng Rehabil 7:38. doi:10.1186/1743-0003-7-38
Caballero C, Barbado D, Moreno FJ (2013) El procesado del desplazamiento del centro de presiones para el estudio de la relación complejidad/rendimiento observada en el control postural en bipedestación. Rev Andal Med Deporte 6:101–107
Caballero C, Barbado D, Moreno FJ (2014) Non-linear tools and methodological concerns measuring human movement variability: an overview. Eur J Hum Mov 32:61–81
Cattaneo D, Carpinella I, Aprile I, Prosperini L, Montesano A, Jonsdottir J (2015) Comparison of upright balance in stroke, Parkinson and multiple sclerosis. Acta Neurol Scand. doi:10.1111/ane.12466
Chen Z, Ivanov PC, Hu K, Stanley HE (2002) Effect of nonstationarities on detrended fluctuation analysis. Phys Rev E 65:041107
Chen W, Wang Z, Xie H, Yu W (2007) Characterization of surface EMG signal based on fuzzy entropy. IEEE Trans Neural Syst Rehabil Eng 15:266–272
Chen W, Zhuang J, Yu W, Wang Z (2009) Measuring complexity using FuzzyEn, ApEn, and SampEn. Med Eng Phys 31:61–68. doi:10.1016/j.medengphy.2008.04.005
Costa M, Goldberger AL, Peng C-K (2002) Multiscale entropy analysis of complex physiologic time series. Phys Rev Lett 89:068102
Danna-Dos-Santos A, Degani AM, Zatsiorsky VM, Latash ML (2008) Is voluntary control of natural postural sway possible? J Mot Behav 40:179–185
Davids K, Kingsbury D, George K, O’Connell M, Stock D (1999) Interacting constraints and the emergence of postural behavior in ACL-deficient subjects. J Mot Behav 31:358–366
Davids K, Glazier P, Araujo D, Bartlett R (2003) Movement systems as dynamical systems: the functional role of variability and its implications for sports medicine. Sports Med 33:245–260
Davids K, Bennett S, Newell KM (2006) Movement system variability. Human kinetics, Champaign, USA
Decker LM, Cignetti F, Stergiou N (2010) Complexity and human gait. Rev Andal deMedicina del Deporte 3:2–12
Donker SF, Roerdink M, Greven AJ, Beek PJ (2007) Regularity of center-of-pressure trajectories depends on the amount of attention invested in postural control. Exp Brain Res 181:1–11
Duarte M, Sternad D (2008) Complexity of human postural control in young and older adults during prolonged standing. Exp Brain Res 191:265–276
Ferguson CJ (2009) An effect size primer: a guide for clinicians and researchers. Prof Psychol Res Pract 40:532–538
Goldberger AL, Amaral LAN, Hausdorff JM, Ivanov PC, Peng C-K, Stanley HE (2002a) Fractal dynamics in physiology: alterations with disease and aging. Proc Natl Acad Sci 99:2466–2472
Goldberger AL, Peng C-K, Lipsitz LA (2002b) What is physiologic complexity and how does it change with aging and disease? Neurobiol Aging 23:23–26
Hancock GR, Butler MS, Fischman MG (1995) On the problem of two-dimensional error scores: measures and analyses of accuracy, bias, and consistency. J Mot Behav 27:241–250
Herzfeld DJ, Shadmehr R (2014) Motor variability is not noise, but grist for the learning mill. Nat Neurosci 17:149–150. doi:10.1038/nn.3633
Hornero R, Aboy M, Abásolo D, McNames J, Goldstein B (2005) Interpretation of approximate entropy: analysis of intracranial pressure approximate entropy during acute intracranial hypertension. IEEE Trans Biomed Eng 52:1671–1680
Krakauer JW, Mazzoni P (2011) Human sensorimotor learning: adaptation, skill, and beyond. Curr Opin Neurobiol 21:636–644
Lake DE, Richman JS, Griffin MP, Moorman JR (2002) Sample entropy analysis of neonatal heart rate variability. Am J Physiol Regul Integr Comp Physiol 283:R789–R797
Lee H, Granata KP (2008) Process stationarity and reliability of trunk postural stability. Clin Biomech 23:735–742
Lin D, Seol H, Nussbaum MA, Madigan ML (2008) Reliability of COP-based postural sway measures and age-related differences. Gait Posture 28:337–342. doi:10.1016/j.gaitpost.2008.01.005
Lipsitz LA, Goldberger AL (1992) Loss of ‘complexity’ and aging. Potential applications of fractals and chaos theory to senescence. JAMA 267:1806–1809
Liu J, Zhang C, Zheng C (2010) EEG-based estimation of mental fatigue by using KPCA–HMM and complexity parameters. Biomed Signal Process Control 5:124–130
Manor B, Lipsitz LA (2013) Physiologic complexity and aging: implications for physical function and rehabilitation. Prog Neuropsychopharmacol Biol Psychiatry 45:287–293. doi:10.1016/j.pnpbp.2012.08.020
Manor B et al (2010) Physiological complexity and system adaptability: evidence from postural control dynamics of older adults. J Appl Physiol 109:1786–1791
Menayo R, Encarnación A, Gea G, Marcos P (2014) Sample entropy-based analysis of differential and traditional training effects on dynamic balance in healthy people. J Mot Behav 46:73–82
Mochizuki L, Duarte M, Amadio AC, Zatsiorsky VM, Latash ML (2006) Changes in postural sway and its fractions in conditions of postural instability. J Appl Biomech 22:51
Newell KM, Slifkin AB (1998) The nature of movement variability. In: Piek JP (ed) Motor behavior and human skill: a multidisciplinary perspective. Human Kinetics, Champaign, pp 143–160
Newell KM, Vaillancourt DE (2001) Dimensional change in motor learning. Hum Mov Sci 20:695–715
Peng CK, Havlin S, Stanley HE, Goldberger AL (1995) Quantification of scaling exponents and crossover phenomena in nonstationary heartbeat time series. Chaos Interdiscip J Nonlinear Sci 5:82–87
Prieto TE, Myklebust JB, Hoffmann RG, Lovett EG, Myklebust BM (1996) Measures of postural steadiness: differences between healthy young and elderly adults. IEEE Trans Biomed Eng 43:956–966
Renart A, Machens CK (2014) Variability in neural activity and behavior. Curr Opin Neurobiol 25:211–220. doi:10.1016/j.conb.2014.02.013
Rhea CK, Silver TA, Hong SL, Ryu JH, Studenka BE, Hughes CML, Haddad JM (2011) Noise and complexity in human postural control: interpreting the different estimations of entropy. PLoS One 6:e17696
Riley MA, Turvey MT (2002) Variability and determinism in motor behavior. J Mot Behav 34:99–125
Riley MA, Shockley K, Van Orden G (2012) Learning from the body about the mind. Top Cogn Sci 4:21–34. doi:10.1111/j.1756-8765.2011.01163.x
Roerdink M, De Haart M, Daffertshofer A, Donker S, Geurts A, Beek P (2006) Dynamical structure of center-of-pressure trajectories in patients recovering from stroke. Exp Brain Res 174:256–269
Roerdink M, Hlavackova P, Vuillerme N (2011) Center-of-pressure regularity as a marker for attentional investment in postural control: a comparison between sitting and standing postures. Hum Mov Sci 30:203–212. doi:10.1016/j.humov.2010.04.005
Schmit JM, Riley MA, Dalvi A, Sahay A, Shear PK, Shockley KD, Pun RY (2006) Deterministic center of pressure patterns characterize postural instability in Parkinson’s disease. Exp Brain Res 168:357–367
Smith BA, Teulier C, Sansom J, Stergiou N, Ulrich BD (2011) Approximate entropy values demonstrate impaired neuromotor control of spontaneous leg activity in infants with myelomeningocele. Pediatric Phys Ther 23:241
Stergiou N, Harbourne RT, Cavanaugh JT (2006) Optimal movement variability: a new theoretical perspective for neurologic physical therapy. J Neurol Phys Ther 30:120–129
Tahayori B, Riley ZA, Mahmoudian A, Koceja DM, Hong SL (2012) Rambling and trembling in response to body loading. Mot Control 16:144–157
Vaillancourt DE, Newell KM (2002) Changing complexity in human behavior and physiology through aging and disease. Neurobiol Aging 23:1–11
Vaillancourt DE, Newell KM (2003) Aging and the time and frequency structure of force output variability. J Appl Physiol 94:903–912
Vaillancourt DE, Sosnoff JJ, Newell KM (2004) Age-related changes in complexity depend on task dynamics. J Appl Physiol (1985) 97:454–455. doi:10.1152/japplphysiol.00244.2004
van Dieën JH, Koppes LLJ, Twisk JWR (2010) Postural sway parameters in seated balancing; their reliability and relationship with balancing performance. Gait Posture 31:42–46
van Emmerik REA, van Wegen EEH (2000) On variability and stability in human movement. J Appl Biomech 16:394–406
Wilkins BA, Komanduri R, Bukkapatnam S, Yang H, Warta G, Benjamin BA (2009) Recurrence quantification analysis (RQA) used for detection of ST segment deviation. FASEB J 23:LB89
Xie H-B, Guo J-Y, Zheng Y-P (2010) Fuzzy approximate entropy analysis of chaotic and natural complex systems: detecting muscle fatigue using electromyography signals. Ann Biomed Eng 38:1483–1496
Zatsiorsky VM, Duarte M (1999) Instant equilibrium point and its migration in standing tasks: rambling and trembling components of the stabilogram. Mot Control 3:28–38
Acknowledgments
This study was made possible by financial support from Science and Innovation Ministry of Spain, Project Cod. DEP2010-19420 and Project Cod. FPU12/00659. Spanish Government.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
This is to inform you that there are no conflicts of interest that could inappropriately influence (bias) this work.
Rights and permissions
About this article

Cite this article
Caballero Sánchez, C., Barbado Murillo, D., Davids, K. et al. Variations in task constraints shape emergent performance outcomes and complexity levels in balancing. Exp Brain Res 234, 1611–1622 (2016). https://doi.org/10.1007/s00221-016-4563-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-016-4563-2