
Abstract
We have recently shown that visuomotor adaptation following reaches with a misaligned cursor not only induces changes in an individual’s motor output, but their proprioceptive sense of hand position as well. Long-term changes are seen in motor adaptation; however, very little is known about the retention of changes in felt hand position. We sought to evaluate whether this recalibration in proprioception, following visuomotor adaptation, is sufficiently robust to be retained the following day (~24 h later), and if so, to determine its extent. Visuomotor adaptation was induced by having subjects perform reaches to visual targets using a cursor representing their unseen hand, which had been gradually rotated 45° counterclockwise. Motor adaptation and proprioceptive recalibration were determined by assessing subjects’ reach aftereffects and changes in hand bias, respectively. We found that subjects adapted their reaches and recalibrated their sense of hand position following training with a misaligned cursor, as shown in Cressman and Henriques (J Neurophysiol 102:3505–3518, 2009). More importantly, subjects who showed proprioceptive recalibration in the direction of motor adaptation on Day 1 did retain changes in felt hand position and motor adaptation on Day 2. These findings suggest that in addition to motor changes, individuals are capable of retaining sensory changes in proprioception up to 24 h later.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The ease with which one can accurately reach to a doorbell can lead one to misconceive the action as simple, whereas in reality it involves an array of complex underlying neural transformations. In order to place the limb at a desired spatial location, goal-directed movements require the central nervous system to interpret sensory information (e.g. vision, proprioception) regarding the target and hand’s position, thereafter transforming this information into the appropriate motor commands (Jeannerod 1988). In most cases, this sensory information is congruent, such that the position one sees and feels their hand overlap. However, under circumstances where this is not the case (e.g. when reaching towards an object under the water surface), the brain will respond by learning a new visuomotor mapping, thus modifying the hand’s subsequent motor commands accordingly. Specifically, when presented with altered visual feedback of their hand, such as while wearing prism goggles or in virtual reality environments, subjects compensate by adjusting their sensorimotor transformations in order to maintain movement accuracy, thereby gradually reducing the error signal experienced when visual and desired movement outcomes are distinct. When visual feedback is removed while performing visual open-loop reaches, individuals will continue to make adapted movements, and these reach errors, termed aftereffects, provide a measure of motor learning. For instance, following training with a rotated cursor, subjects’ open-loop movement trajectories will remain deviated in the direction of adaptation.
Various studies have demonstrated that induced motor changes accompanying visuomotor adaptation remain long after training has ended, and can last from several days up to a year following training. Specifically, after adapting to a perturbation, subjects continue to show reach aftereffects (Yamamoto et al. 2006) or faster relearning when exposed to the same perturbation on a later day (i.e. presence of savings) (Krakauer et al. 1999; Bock et al. 2001; Tong et al. 2002; Caithness et al. 2004; Klassen et al. 2005; Krakauer et al. 2005). For instance, when Klassen et al. (2005) retested subjects 1 day after they had modified their cursor reaches to an abrupt 30° rotation, the initial errors produced at the start of the second day were similar to those at the end of training on the first day, suggesting almost complete retention of motor adaptation. Likewise, Caithness et al. (2004) found that when subjects were retested on a 30° visuomotor rotation 48 h after initial adaptation, initial deviations in reaching were reduced by approximately half when compared to those produced in the very first block 2 days before, once again suggesting substantial retention of the visuomotor adaptation. Bock et al. (2001) also demonstrated retention of motor adaptation in an experiment where a sensorimotor discordance was introduced by reversing cursor movement either left–right or up–down. In this study, subjects tracked a moving visual target by shifting the perturbed cursor with a joystick. They were tested again in a second session on the perturbed tracking task at one of various time points (8, 25 min, 1, 2 h, or 1 month later). Their overall performance (the root mean square error of the distance between the cursor and target) was greatly improved; specifically, when retested up to 1 month later, subjects showed almost complete retention of the adaptation, such that tracking errors were observed to be close to end-of-training levels (baseline). Yamamoto et al. (2006) found that in addition to the commonly observed short-term changes accompanying the process of motor learning, motor memory can be retained for extended periods of time. In this study, subjects learned to use a joystick to move a cursor, which was gradually rotated 40°, onto a target. Following training, when reaching in the absence of a cursor, subjects showed compensation in their movement aftereffects for 63–91 % of the rotation. More importantly, 59–91 % of the compensation was still present 1 year later, relative to the initial post-test (41–77 % of the visuomotor distortion).
Long-lasting effects on motor performance have also been found following adaptation to other perturbations, such as a velocity-dependent force field (Brashers-Krug et al. 1996; Shadmehr and Brashers-Krug 1997). For example, Shadmehr and Brashers-Krug (1997) found that after subjects had adapted their reaches with a velocity-dependent force field, the level of performance achieved on that first day persisted up to 24 h later while reaching in the same force field. Interestingly, this improvement remained constant when retested 5 months later, suggesting the formation of long-term motor memories following this type of learning. Moreover, subjects who participated in prism studies have also shown similar long-lasting aftereffects following training (Hay et al. 1966; Martin et al. 1996; Hatada et al. 2006). For instance, individuals who spent 6 weeks adapting their throwing movements while wearing displacing prisms showed almost perfect retention when retested 9, 18, and 27 months following initial training, such that they accurately threw balls near the centre of the target, on their first throw, right after donning prism goggles (Martin et al. 1996).
In addition to motor changes, visuomotor adaptation has been shown to result in short-term sensory changes. Previous studies examining reaching with altered feedback of the hand have suggested that visuomotor adaptation is partially due to the recalibration of proprioception (felt joint or limb position in space) to match its visual representation (Hay et al. 1965; Redding and Wallace 2004; Bernier et al. 2005; Redding et al. 2005; Simani et al. 2007). In these previous studies, subjects used their adapted hand to perform proprioceptive estimation tasks, thus making it difficult to discern whether recalibration to such targets arose due to proprioceptive recalibration or motor adaptation. However, changes in felt hand position have been observed using paradigms which do not require subjects to make goal-directed movements, hence eliminating the possibility of a motor confound (Cressman and Henriques 2009, 2010b; Ostry et al. 2010). Specifically, Cressman and Henriques (2009) had participants move a robot manipulandum along a constrained pathway to a designated location. Once it reached its final position, a visual or body midline (a non-visual reference point) reference marker appeared and subjects made a forced-choice judgment of the position of their hand relative to the marker (left or right). When tested following visuomotor adaptation, subjects displayed a significant change in felt hand position in the expected direction of motor adaptation. Results from our laboratory have shown that this recalibration of hand position occurs under a multitude of contexts, including when the hand is moved passively or guided by the robot during felt hand position estimation tasks (Cressman and Henriques 2009) and both in healthy young and older adults (Cressman et al. 2010). Proprioceptive recalibration also arises following adaptation to both rotated and translated cursor distortions (Cressman and Henriques 2009), in both the left and right hands of right-handed individuals (Salomonczyk et al. 2012; Mostafa et al. 2014), and even after exposure to a sensory discrepancy in which no goal-directed movements were made during training (Cressman and Henriques 2010b; Salomonczyk et al. 2013).
Still unknown is whether these changes in hand proprioception persist beyond training, following visuomotor adaptation, the way reach adaptation or motor memory does. Retention of the change in sense of felt hand position was investigated following training with a velocity-dependent force field perturbation and has been shown to persist at least 24 h after adaptation (Ostry et al. 2010). This change in felt limb position was measured by having subjects report the direction their hand had been deflected (left or right) during their reaches; this was measured at various intervals throughout the experiment, including before, after, and 24 h following training. The authors observed a perceptual shift of approximately 11 % (compared with peak deviations in aftereffects) of the estimated magnitude of learning following training, and interestingly, these sensory changes were also observed to a similar extent 24 h later. The latter finding suggests that persistent changes in the sensory system are present following adaptation to a velocity-dependent force field. How these somatosensory changes compare with motor changes, such as aftereffects or savings, is not known, since changes in movement were not re-measured the following day. Moreover, it is unclear whether the persistence of proprioceptive changes in individuals would occur following training with a visuomotor rotation, since normal vision of the hand in the intervening 24 h would presumably lead to a return of the usual visual–proprioceptive alignment.
As previously mentioned, a multitude of studies delve into the long-term changes in motor systems; however, literature exploring the persistence of proprioceptive changes following visuomotor adaptation is left unexplored. Our study aims to investigate whether proprioceptive recalibration and motor adaptation are retained after a 24-h period following training, and if found, the magnitude of this effect. These results will provide insight into the contributions of sensory memory to visuomotor adaptation and the relationship between the processes underlying motor and sensory adaptation.
Methods
Subjects
Sixty right-handed subjects were recruited from York University (mean age = 23.5 year, SD = 5.5 year, 43 women), 39 of which were enrolled in a first-year university psychology course and participated for course credit. All subjects were free from neurological impairments as well as pre-screened for self-reported right-handedness and corrected to normal vision. Data from 12 of our 60 subjects were excluded from analyses due to inconsistent performance in the Proprioceptive Estimates (mentioned below) following aligned reach training (baseline performance). Specifically, the two-alternative forced-choice (2AFC) responses were too erratic to properly fit a logistic function, and therefore, we were unable to get a reliable estimate of proprioceptive bias for these 12 subjects in their baseline task. Without such a baseline estimate of bias, we could not address the question of whether proprioceptive biases change following visuomotor adaptation, and so these subjects were excluded from the dataset. An additional subject was also removed since the changes in their proprioceptive bias, although in the expected direction of motor adaptation, fell outside 3 SD of the norm (a shift in proprioceptive bias of nearly 30° rightwards). Thus, results from 47 subjects are presented below. Informed consent was given, and the experiment was performed in accordance with the ethical guidelines set by the York Human Participants Review Sub-Committee, York University’s Ethics Review Board and the standards of the Canadian Tri-Council Research Ethics guidelines.
General experimental set-up
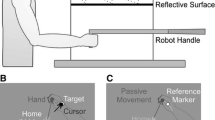
Subjects were seated in a chair adjustable in both height and distance, ensuring that they could comfortably see and reach to all target positions presented on an opaque and reflective surface; a view of the set-up is presented in Fig. 1a. Once the chair was adjusted, the position was fixed and remained consistent for all experimental sessions. Subjects were instructed to grasp the vertical handle of a two-joint robot manipulandum (Interactive Motion Technologies) with their right hand. Visual stimuli were projected from a monitor (SyncMaster model 510N; refresh rate: 72 Hz; Samsung, Brisbane, CA) installed 17 cm above the robot and viewed by subjects as a reflected image. The position of the manipulandum’s gripped-handle was recorded throughout all reaching trials at a sampling rate of 50 Hz and a spatial accuracy of .1 mm. The room lights were dimmed and the reflective surface was positioned so that images displayed on the monitor appeared to lie in the same horizontal plane as that of the robot handle. Subjects’ view of their right hand was obstructed by the reflective surface and a black cloth draped between the experimental set-up and their shoulders.

Experimental set-up and design. a Side view of the experimental set-up. b, c Top view of experimental surface visible to subjects. b Visuomotor distortion introduced in the misaligned Reach Training task. The 1-cm green cursor representing the hand was gradually rotated 45° CCW relative to the subject’s actual hand location (black ring). Reach targets (yellow ring) 1 cm in size were located 10 cm from the home position (black circle) at 30° CW, 0°, or 30° CCW relative to the home position. c In the Proprioceptive Estimates, subjects actively pushed their hand out 10 cm along a constrained linear path (depicted in red) from the home position and judged the position of their hand with respect to a reference marker (yellow rings). Reference markers were either visual (30° CW, 0°, or 30° CCW relative to the home position), or proprioceptive, prompted by an auditory cue, wherein subjects judged the position of their hand relative to their perceived midline (color figure online)
General procedure
The experiment consisted of two separate testing sessions, each conducted on two consecutive days, 24 h apart. The first session consisted of two blocks (Fig. 2, first 2 rows), which included three types of tasks per block. In Block 1, subjects completed reach training trials with an aligned cursor (Fig. 2, top row, box 1), during which the visual cursor was accurately aligned with the subject’s hand, followed by open-loop reaches made without visual feedback (No-Cursor task; Fig. 2, top row, boxes 2 and 4) and estimates of felt hand position (Proprioceptive Estimates; Fig. 2, top row, box 3). The latter was followed immediately by Block 2, which was similar to Block 1, except reach training trials involved a cursor that was rotated 45° counterclockwise (CCW) relative to the hand (Fig. 2, middle row, box 1). The rotation was introduced gradually during the reach training trials, increasing by .75° per trial, such that by trial 60 the 45° distortion was achieved. On the next day, subjects were retested on the No-Cursor reaches, to measure persisting aftereffects (Fig. 2, bottom row, box 1), as well as on the Proprioceptive Estimates (Fig. 2, bottom row, box 2), to measure whether changes in felt hand position from the previous day had been retained.

Schematic showing the order in which tasks were completed and the number of trials completed per task. Tasks completed during Block 1 provided baseline measures of performance, wherein the subject’s unseen hand was aligned with the cursor during reach training (top row, box 1). In Block 2, the cursor was gradually rotated 45° CCW relative to the subject’s actual hand location during reach training (middle row, boxes 1 and 5). On Day 2, subjects were only tested on open-loop No-Cursor reaches and Proprioceptive Estimates without any reach training
Reach training
During this task, subjects made reaching movements to a yellow target circle (1 cm in diameter) displayed on the reflective surface. The three visual targets were at 30° CW, 0°, or 30° CCW relative to the home position and located 10 cm radially from the home position. The order of the targets was pseudo-randomized, such that subjects reached once to each of the three targets before any target was repeated. The position of the subject’s unseen hand was represented by a green circular cursor, also 1 cm in diameter, which was either aligned (Block 1) or rotated (Block 2) (Fig. 1b) relative to the hand’s actual location. During Block 1, subjects made 60 reaches (Fig. 2, top row, box 1) to the visual target, while in Block 2, they made 150 reaches before Proprioceptive Estimates and 30 reaches afterwards (Fig. 2, middle row, boxes 1 and 5).
Trials began with the subject’s hand at the home position 40 cm from their body midline, and after 300 ms, a yellow target appeared. Subjects were then asked to reach towards the target while moving the robot handle as quickly and accurately as possible, so as to align both circles. For each trial, the cursor became visible only after the handle was moved 4 cm away from the home position so that subjects had likely reached peak velocity prior to the onset of visual feedback.
The reach was completed once the cursor’s centre was positioned within .5 cm of the target’s centre. At that point, both circles disappeared and subjects moved their hand back to the home position via a robot-generated grooved pathway. If any attempt to move outside the established path was made, a resistance force (proportional to the depth of penetration with a stiffness of 2 N/mm and a viscous damping of 5 N/(mm/s)) was generated perpendicular to the grooved wall (Henriques and Soechting 2003).
No-Cursor reaches
The No-Cursor task was similar to the Reach Training task, except subjects were instructed to reach towards the target upon presentation without cursor feedback of their unseen hand. The task consisted of 15 trials and was administered before and after the Proprioceptive Estimates, in each block, on the first day (Fig. 2, first and middle row, boxes 2 and 4 in each row). In addition, No-Cursor reaches were measured once more before the proprioceptive task on Day 2 as a final measure of reach adaptation (Fig. 2, bottom row, box 1). Thus, throughout the experiment, aftereffects were measured on five separate occasions. After subjects’ endpoint position was maintained for 500 ms, the visual target disappeared, subjects moved their hand back to the home position via a grooved pathway, and the trial was considered complete.
Proprioceptive Estimates
The Proprioceptive Estimates was used to measure subjects’ felt hand location. A trial began with the illumination of the home position indicated by a green 1-cm circle for 500 ms. Subjects were instructed to wait until the green circle disappeared to push their hand outwards along a constrained, robot-generated linear pathway described earlier (Fig. 1c). Once their hand arrived at the end of the path, one of three visual markers, represented by a yellow circle (1 cm in size) located at 30° CW, 0°, or 30° CCW relative to the home position, appeared. Alternatively, on one-quarter of the trials, we used a proprioceptive marker, such that rather than a visual marker an auditory cue was heard at the end of the pathway, which indicated to subjects that they were to estimate their hand position with respect to their perceived body midline. Without time constraints, subjects then made a two-alternative forced-choice (2AFC) judgment about the position of their hand (left or right) relative to the reference marker using a keyboard with their free left hand. To begin the next trial, they then moved the robot directly back to the home position along the robot-generated linear route. Subjects completed 200 trials per task, administered once in each block on Day 1 (Fig. 2, top and middle row, box 3) and once on Day 2 (Fig. 2, bottom row, box 2). The position of the hand with respect to each reference marker was adjusted over trials using an adaptive staircase algorithm (Kesten 1958; Treutwein 1995), as described by Cressman and Henriques (2009).
Data analyses
No-Cursor reaches
We investigated whether subjects adapted their reaches after training with a rotated cursor and if this adaptation was maintained 24 h later by looking at No-Cursor reaches (i.e. aftereffects). Reach endpoint errors were defined as the angular deviation between a movement vector (from the home position to the reach endpoint) and a reference vector (from the home position to the target). Similarly, reach errors at peak velocity were defined as the angular difference between a movement vector at peak velocity and a reference vector. We conducted a two-way repeated measure analysis of variance (RM-ANOVA) based on the No-Cursor reaches (Average Aligned, Rotated Time 1, Rotated Time 2, and Retention) and three targets (0°, 30° CW, and 30° CCW). This was followed by our pre-planned comparisons between the average of the two sets of No-Cursor reaches, following aligned reach training, that served as baseline (Fig. 2, top row, boxes 2 and 4) with the set of reaches produced immediately after training with the rotated cursor (Rotated Time 1; Fig. 2, middle row, box 2) and then again with the set that followed approximately 25–30 min later, after performing the Proprioceptive Estimates (Rotated time 2; Fig. 2, middle row, box 4). These comparisons measured whether reach adaptation had occurred, as well as the extent of this adaptation. To test for retention, we compared subjects’ performance at Rotated Time 2 with the No-Cursor reaches measured 24 h later (Retention; Fig. 2, bottom row, box 1).
Proprioceptive Estimates
The position at which subjects perceived their hand to be aligned with the reference marker was measured by fitting their responses to a logistic function for each of the four reference marker locations. Biases, which are the measure of accuracy of a subject’s proprioceptive sense of hand position, were then calculated. It is also understood as the probability of reporting left or right equally often (50 %). A subject’s bias for a particular marker was excluded if they surpassed +3 SD (12 out of 552 marker values were replaced with their respective mean).
Proprioceptive bias was analysed comparing those measured after aligned and misaligned training on Day 1 and again on Day 2 (Day-1 Aligned, Day-1 Rotated, Day-2 Retention) and marker location (30° CW, 0°, 30° CCW, or proprioceptive midline) using a RM-ANOVA. A generalized estimating equation (GEE) was also performed to determine whether replacing missing data with group means influenced our results. We found that the results of the GEE analysis were similar to those obtained using mean substitution, such that it did not produce any changes in the means nor in the significance of our results, cross-validating the results we had obtained using the RM-ANOVA. Therefore, only the results from the RM-ANOVA, based on mean substitution, are reported here.
A Bonferroni correction was applied to all post hoc pair-wise comparisons, and all ANOVA results are reported with Greenhouse–Geisser corrected p values to compensate for violations of sphericity.
Results
Reach adaptation and retention
Mean reach endpoint errors per trial, from the No-Cursor tasks, are displayed in Fig. 3a. In order to measure the retention of visuomotor adaptation, we primarily determined whether reach adaptation had occurred. This was done by comparing mean reach endpoint errors, from the No-Cursor tasks, following training with an aligned cursor (Fig. 3a, average of panels 1 and 2), with those produced immediately after training with a rotated cursor (panel 3) and those produced after the Proprioceptive Estimates (panel 4), all on Day 1. We found that mean reach endpoint errors did significantly change by an average of 11.8° in the expected direction of adaptation (~26 % of the distortion), when measured immediately following training with a rotated cursor (Fig. 3b, dark grey bar). When we measured mean reach endpoint error on the same day as training, but after 150 trials of the Proprioceptive Estimates, the reach aftereffects were reduced to approximately 4.7° (~10 % of the distortion) (Fig. 3b, light grey bar). Although smaller than the changes produced immediately after rotated reach training, these reach aftereffects were still significantly deviated in the direction of adaptation, compared with baseline measures (p < .001). Interestingly, rather than observing an abrupt decay in reach error from panel 3 to panel 4 (Fig. 3a), there was a gradual decline in the magnitude of endpoint error across trials that flanked the Proprioceptive Estimates on Day 1.

a Mean endpoint error, per trial, for each No-Cursor task. Panels 1 and 2 reflect the No-Cursor reaches performed after reach training with an aligned cursor, while panels 3 and 4 reflect those after training with a rotated cursor, all on Day 1. Panels 2 and 4 were conducted after the Proprioceptive Estimates, however, each following the different reach training sessions. Panel 5 shows No-Cursor reaches measured 24 h following reach training. b Mean reach aftereffects, seen as changes in endpoint error, immediately after (Day 1) and 24 h after (Day 2) training with misaligned feedback of the hand, relative to baseline No-Cursor reaches, averaged across panels 1 and 2. The dark grey bar shows reach aftereffects immediately after reach training with a misaligned cursor (a, panel 3 relative to baseline), while the light grey bar shows aftereffects following subsequent completion of the Proprioceptive Estimates (a, panel 4 relative to baseline). The white bar shows reach aftereffects when tested 24 h later (a, panel 5 relative to baseline). Corresponding errors at peak velocity are shown as diamonds. Error bars reflect SEM
Following the confirmation of initial reach adaptation on Day 1, we then determined whether these aftereffects persisted 24 h later. To do so, we compared the subjects’ last set of No-Cursor reaches on Day 1 (Fig. 3a, panel 4) with those measured immediately upon their return to the laboratory 24 h later (i.e. Day 2). These retained reach aftereffects were not different from those in the last set of No-Cursor reaches on Day 1 (Fig. 3b, light grey vs. white bar) (p > .99). Aftereffects from Day 2 relative to the baseline results were also significantly different (p < .001).
In sum, these results confirm that motor adaptation on Day 1 was retained the following day, such that changes in open-loop reaches following rotated reach training on Day 1 and those measured 24 h later, on Day 2, were significantly deviated compared with baseline reaches (p < .001). Reach aftereffects at peak velocity displayed similar results to that of endpoint reach errors (Fig. 3b, diamonds).
Proprioceptive recalibration and retention
Estimates of hand position following visuomotor adaptation were analysed to investigate proprioceptive recalibration both immediately following training and 24 h later. Figure 4a depicts the positions at which subjects perceived their hand to be aligned with each reference marker following aligned reach training (diamonds), rotated reach training (triangles), and 24 h later (squares). Baseline measures of sense of felt hand position revealed a slight leftward bias, which has been seen in our previous studies and has been suggested to arise due to a systematic hand bias (Jones et al. 2010; Salomonczyk et al. 2012, 2013). Mean bias estimates differed significantly when comparing those measured after aligned and misaligned training on Day 1 and again on Day 2 (F(2,92) = 5.2, p = .007). Post hoc analysis revealed that on average, subjects showed a significant 2.6° shift rightwards in sense of felt hand position following rotated reach training, as compared to following aligned reach training (p = .033) (Fig. 4c, first white bar). When these subjects were retested on the Proprioceptive Estimates the following day, we observed a significant 2.6° shift once again in hand bias relative to baseline measures (p = .0013) (Fig. 4c, first grey bar), suggesting that the change in bias found immediately after rotated reach training was perfectly retained 24 h later. However, this 2.6° shift observed immediately following reach training is lower than usually observed in previous experiments, using similar methods, where subjects trained with a misaligned cursor (30° distortion) (Cressman and Henriques 2009; Salomonczyk et al. 2012). Upon further inspection, we noticed a number of subjects (n = 15, ~32 %) whose change in hand bias was not in the expected direction following training with a rotated cursor (Fig. 4d, grey circles). Since testing for retention requires an initial change that can be retained, we performed a re-analysis on a subgroup of subjects (n = 32), that did show a positive change in bias of at least .001° rightwards, the expected direction of adaptation, when compared to baseline performance (Fig. 4d, hollow circles). Figure 4b depicts the positions at which subjects in the subgroup perceived their hand to be aligned with each reference marker following aligned reach training (diamonds), rotated reach training (triangles), and 24 h later (squares). This subgroup’s mean bias estimates also differed significantly when comparing bias measured after aligned and misaligned training on Day 1 and again on Day 2 (F(2,62) = 13.4, p < .001). These changes in Proprioceptive Estimates of hand position were comparable across all visual and proprioceptive (body midline) reference marker locations for this subgroup (F(1.2,39.1) = 2.1, p = .15). Post hoc analysis revealed that on average, these subjects showed a significant 4.5° shift rightwards in their sense of felt hand position following rotated reach training, as compared to following aligned reach training (Fig. 4c, second white bar), (p < .001). Moreover, proprioceptive recalibration for Day 2 was approximately 3.2°, a change in bias that was significantly different than baseline measures (p = .01), and represents a 72 % retention of the 4.5° change in bias seen immediately following misaligned reach training. While this change in proprioceptive bias 1 day after training appears smaller than those produced immediately after reach training, this reduction did not reach significance (p = .51). Interestingly, for 11 out of these 32 subjects, their changes in bias on Day 2 were greater than changes in felt hand position measured immediately after reaching with a rotated cursor relative to baseline performance. These results suggest that changes in sense of felt hand position following visuomotor adaptation can be partially retained up to 24 h later, similar to motor adaptation.

a Mean two-dimensional (2-D) proprioceptive biases after all subjects were exposed to aligned (diamonds) and rotated (triangle) reach training (n = 47), for each reference marker location. Mean change in sense of felt hand position measured the following day is represented by squares, and the circles represent actual reference marker positions; for clarity, biases for the proprioceptive marker are shown as hollow shapes displaced slightly above the biases for the visual reference markers. b 2-D proprioceptive biases for our subgroup (n = 32). c Mean change in bias following training with a misaligned cursor (white bars) as well as 24 h later (grey bars). The first pair of bars is the measures for all our subjects, whereas the second pair is for the subgroup. Error bars reflect SEM. d The mean change in Proprioceptive Estimates, for each subject, on Day 2 is plotted as a function of the change on Day 1. Approximately 32 % of the 47 subjects did not show any change in sense of felt hand position in the direction of adaptation (grey circles) possibly due to lack of interleaved reach training. Those who showed a change of at least .001° in the positive direction were represented as hollow circles
A one-way ANOVA revealed that our subgroup did not differ from the subjects that did not show a positive bias (grey circles in Fig. 4d) with respect to their slope (i.e. uncertainty range) fitted to the Proprioceptive Estimates (F(1,140) = .476, p = .49). Notably, when re-analysed, this subgroup also showed similar reach aftereffects as all of the subjects (mentioned above).
We also applied a regression procedure and observed that retained mean change in aftereffects for all subjects (β = .195, p = .189) as well as the subgroup (β = −.0922, p = 922) were not a significant predictor of retained mean change in proprioceptive bias.
Discussion
The goal of this study was to examine whether induced changes in subjects’ sense of felt hand position following visuomotor adaptation were robust enough to be maintained into the following day, and if so, to determine their magnitude. We assessed motor adaptation (seen as reach aftereffects) and proprioceptive recalibration of hand bias (seen as a change in felt hand position) immediately after training with a rotated cursor and 24 h later. We found that, immediately following reach training with distorted visual feedback, subjects adapted their mean open-loop reaching movements by 11.8°, or 26 % of the distortion, compared with baseline reaches. Following the Proprioceptive Estimates, subjects continued to show reach aftereffects, such that they had deviated mean reaching trajectories of 4.7°, or 10.4 % of the distortion, compared with baseline. When retested 24 h later, subjects showed retention of these adapted hand movements from Day 1 by 5.4°, approximately 46 % of mean aftereffects produced in the first set of No-Cursor reaches following reach training, and complete retention of aftereffects produced during the last set of No-Cursor reaches. For subjects that displayed proprioceptive recalibration in the expected direction, sense of felt hand position shifted by 4.5° compared with baseline results and did indeed retain 72 % of this change the following day. In addition, their changes in reach aftereffects were similar to those reported for the entire group. These results suggest that following visuomotor adaptation, changes affecting the sensory system, specifically proprioception, can persist beyond initial training, up to at least 24 h.
Proprioceptive and motor changes following visuomotor adaptation
Although both proprioceptive and motor changes occur following visuomotor adaptation (Cressman and Henriques 2009), the nature of their relationship still remains unclear. In our laboratories’ previous work, changes in subjects’ sense of felt hand position were not significantly related to changes in their aftereffects after free-reach training with a misaligned cursor (Cressman and Henriques 2009, 2010b; Salomonczyk et al. 2011, 2012). However, the aforementioned relationship had not yet been investigated following an extended lapse of time. We found that proprioceptive recalibration of hand bias following visuomotor adaptation is still not significantly related to changes in aftereffects when measured approximately 24 h after training with a misaligned cursor, suggesting that the two processes may occur concurrently through distinctive processes. This idea is strongly supported in studies where specific generalization patterns occur. For instance, Mostafa et al. (2014) found that after training with a misaligned cursor, motor adaptation had transferred from subjects’ right (dominant) hand to their left (non-dominant) hand; however, proprioceptive changes did not transfer at all between hands and were only found in the arm exposed to the distortion. Generalization patterns have also been explored by Cressman and Henriques (2010a) wherein subjects’ hand movements and proprioceptive recalibration to novel targets were studied following visuomotor adaptation. They found that subjects’ reaches confirmed previous studies’ observations of localized and narrow generalization patterns in motor adaptation (Krakauer et al. 2000; Wang and Sainburg 2005). Interestingly, proprioceptive recalibration generalized quite broadly in comparison, such that the magnitude of recalibration observed for the trained target direction was found at similar levels up to 90° CCW from the trained direction. They also found that after training with small cursor gains, subjects’ reaches had been adapted, whereas their sense of felt hand position was not recalibrated. They suggest that these generalization patterns in sensory and motor plasticity may be the result of two respective error signals processed in different areas of the brain: one that arises from a discrepancy between desired and actual movement, mainly responsible for motor changes, and one from a discrepancy between visual and Proprioceptive Estimates of hand position, mainly responsible for sensory changes.
Retention of motor adaptation
Consistent with previous studies, subjects’ reaches following visuomotor adaptation remained deviated in the direction of adaptation up to 1 day later. Although most studies measure retention through facilitation, in the form of savings or increased re-adaptation rates, we wished to compare changes in sensory estimates with those changes observed in motor endpoint error. Mean reach aftereffects observed only 24 h later compared with the mean of those immediately following rotated reach training were smaller, with only 46 % retained, in comparison with those observed up to an entire year later as found by Yamamoto et al. (2006), which showed up to 91 % retention relative to the initial post-test. One explanation for the latter’s high to near-perfect retention of aftereffects after an extended period of time may have been due to the joystick paradigm itself, whereas similar results may be less likely for reaches which are less restrained to a particular tool. Specifically, certain perturbations (i.e. those associated with an uncommon everyday tool) may provide a sufficiently distinct context, such that everyday movements would not interfere with learning and retention following adaptation (Brashers-Krug et al. 1996; Wolpert et al. 1998; Krakauer et al. 1999).
Visual open-loop reaches were performed to measure aftereffects following training with a gradually introduced rotated cursor, both before (Fig. 3a, panel 3) and immediately after (Fig. 3a, panel 4) the Proprioceptive Estimates, which took approximately 25–30 min to complete. The smooth continuous decrease in reach aftereffects across trials suggests that this estimation task between the these two No-Cursor tasks did not cause nor accelerate this decay and that the smaller aftereffects seen in the last set of No-Cursor reaches are likely due to a gradual washout. In some of our other studies on this topic, we try to maintain the same level of reach adaptation by interleaving rotated cursor training with Proprioceptive Estimate trials, a technique that others have also used for similar purposes (Kesten 1958; Simani et al. 2007; Synofzik et al. 2006; Ostry et al. 2010). Nonetheless, this continuous decrease in reach aftereffects appears to saturate at approximately 5° on Day 1 (Fig. 3a, panel 4) and seems to be maintained into the following day at a similar level (Fig. 3a, panel 5). Specifically, when measured 24 h later, aftereffects from the last set of open-loop reaches on Day 1 were almost completely preserved on Day 2. Smith et al. (2006) described two adaptive processes that result from learning: a fast-learning process and a slow-learning process. Since the fast-learning process is thought to decay much earlier than its counterpart, its gradual waning would reveal the slow-learning process that is thought to be responsible for savings and anterograde interference (Shadmehr et al. 2010). It is possible that the aftereffects observed immediately after training (Fig. 3a, panel 3) may account for a combination of the two processes including this fast-learning process of adaptation highly involved in initial learning, whereas those observed after the gradual decay are the resulting aftereffects from the slow-learning process. Furthermore, it is these robust aftereffects resulting from the slow-learning process, accounting for 42 % of the first set of aftereffects, which are retained the next day. In a study by Joiner and Smith (2008), a force field adaptation task was used to study the relationship between initial learning and long-term retention; retention measured 24 h later was observed to have a very similar shape as the model-predicted slow-learning process (r = .99), suggesting that the slow process contributes strongly to retention of motor memories, whereas the fast-process does not.
Retention of proprioceptive recalibration
Our main interest for this study was the degree of retention with regard to changes in proprioceptive hand position following visuomotor adaptation, which had not been investigated to date. When we analysed our entire group of subjects, we found that, on average, individuals significantly recalibrated their sense of hand position to be 2.6° more to the right (the direction of motor adaptation) compared with baseline measures. When tested the following day, subjects showed perfect retention (99.6 % retained) of this shift in sense of felt hand position. However, given the smaller magnitude following adaptation compared with our previous papers (~6° change with a 30° distortion) (Cressman and Henriques 2009, 2010b; Cressman et al. 2010; Salomonczyk et al. 2011), we investigated further and noticed a group of subjects who did not show a shift in the expected direction following training with a visuomotor discrepancy (Fig. 4d, grey circles), despite having equivalent slopes (or uncertainty ranges) as the others, confirming correct performance of the task. These subjects may not have shown expected recalibration due to a lack of interleaved reach training, which would have helped maintain proprioceptive recalibration throughout the experiment. For instance, in our previous studies that had weaved reach training trials throughout Proprioceptive Estimates, approximately 83–100 % of subjects showed the expected shift in sense of hand position (Cressman and Henriques 2009, 2010b; Cressman et al. 2010; Salomonczyk et al. 2011, 2012; Clayton et al. 2013), whereas in those that had not (Salomonczyk et al. 2013; Mostafa et al. 2014), only 72–91 % of subjects showed this recalibration. However, interleaving rotated reach training with the Proprioceptive Estimates in this study would have proven to be problematic, since any type of re-training on Day 2 had to be avoided in order to ensure we were testing for retention of motor and proprioceptive changes.
So that we may draw clear conclusions on the persistence of proprioceptive recalibration, we then only looked at subjects who showed at least a .001° proprioceptive shift in bias in the direction of the distortion (32 in total, or the remaining 68 %). This allowed us to see whether subjects who showed at least some changes in felt hand position continued to do so the following day. On average, the shift in the sense of felt hand position for these 32 subjects increased to approximately 4.5° and their estimates of hand position were significantly shifted by 3.2°, compared with baseline, 24 h later. These results are similar to those found by Ostry et al. (2010) where subjects retained a shift of 2 mm in perceived in limb position following 24 h after adaptation to a force field. While the size of the retention of proprioceptive recalibration in this study appears rather small, it is similar, if not larger, than the changes in felt hand motion found by Ostry et al., such that the somatosensory changes they observed were about 10 % of the size of initial reach aftereffects on Day 1 (~20 mm) compared with 22 % for all our subjects and 27 % for our subgroup. Interestingly, 7 out of the 15 subjects who did not show a change in hand position in the expected direction of adaptation immediately following visuomotor adaptation did so 24 h later by an average of 4.9° rightwards (Fig. 4d, top left quadrant). It may be possible that for a small number of people, this change in proprioception benefits by offline gains, an improvement in performance following a time period deprived of training.
Our results, both the initial change in hand proprioception following reach training and its retention a day later, are also consistent with Berniker and Kording’s (2008) source-estimation model, which is based on how a person’s nervous system attributes observed movement errors, such as when adapting to a perturbation. Specifically, the model asks whether one principally assigns the source of errors to changes in world-based or body-based properties. Berniker and Kording (2011) have applied their model to successfully simulate the change in proprioceptive bias observed in our original study (Cressman and Henriques 2009), and those of others (Synofzik et al. 2006; Ostry et al. 2010), attributing these perceptual changes to a body-based source of error. Likewise, in this later paper, the authors describe the subsequent savings or retention by parcelling out the contribution of world-based and body-based estimates. Their model suggests that we, like Ostry et al. (2010), should find a change in this proprioceptive (body-based) estimate of hand position that persists across time, even after a washout, which is seen in the current study.
In summary, subjects expectedly showed motor adaptation and proprioceptive recalibration following visuomotor adaptation. As shown in previous studies, their adapted reach movements were retained up to the next day following training with a rotated cursor. More importantly, subjects’ induced sensory changes, specifically proprioceptive recalibration, persisted up to 24 h later as well. These results suggest that sensory memory may play a significant role alongside motor memory with regard to the long-term changes, an individual may experience following visuomotor adaptation, and further investigation may provide insight into the relationship between both of these processes.
References
Bernier PM, Chua R, Franks IM (2005) Is proprioception calibrated during visually guided movements? Exp Brain Res 167:292–296
Berniker M, Kording K (2008) Estimating the sources of motor errors for adaptation and generalization. Nat Neurosci 11:1454–1461
Berniker M, Kording KP (2011) Estimating the relevance of world disturbances to explain savings, interference and long-term motor adaptation effects. PLoS Comput Biol 7:e1002210
Bock O, Schneider S, Bloomberg J (2001) Conditions for interference versus facilitation during sequential sensorimotor adaptation. Exp Brain Res 138:359–365
Brashers-Krug T, Shadmehr R, Bizzi E (1996) Consolidation in human motor memory. Nature 382:252–255
Caithness G, Osu R, Bays P, Chase H, Klassen J, Kawato M, Wolpert DM, Flanagan JR (2004) Failure to consolidate the consolidation theory of learning for sensorimotor adaptation tasks. J Neurosci 24:8662–8671
Clayton HA, Cressman EK, Henriques DY (2013) Proprioceptive sensitivity in Ehlers-Danlos syndrome patients. Exp Brain Res 230:311–321
Cressman EK, Henriques DY (2009) Sensory recalibration of hand position following visuomotor adaptation. J Neurophysiol 102:3505–3518
Cressman EK, Henriques DY (2010a) Generalization patterns for motor and sensory plasticity differ following visuomotor learning. In: Society for neuroscience. San Diego, California
Cressman EK, Henriques DY (2010b) Reach adaptation and proprioceptive recalibration following exposure to misaligned sensory input. J Neurophysiol 103:1888–1895
Cressman EK, Salomonczyk D, Henriques DY (2010) Visuomotor adaptation and proprioceptive recalibration in older adults. Exp Brain Res 205:533–544
Hatada Y, Miall RC, Rossetti Y (2006) Two waves of a long-lasting aftereffect of prism adaptation measured over 7 days. Exp Brain Res 169:417–426
Hay JC, Pick HLJ, Ikeda K (1965) Visual capture produced by prism spectacles. Psychon Sci 2:215–216
Hay JC, Pick Jr, Herbert L (1966) Visual and proprioceptive adaptation to optical displacement of the visual stimulus. J Exp Psychol 71:150–158
Henriques DY, Soechting JF (2003) Bias and sensitivity in the haptic perception of geometry. Exp Brain Res 150:95–108
Jeannerod M (1988) The neural and behavioural organization of goal-directed movements. Oxford University Press, Oxford
Joiner WM, Smith MA (2008) Long-term retention explained by a model of short-term learning in the adaptive control of reaching. J Neurophysiol 100:2948–2955
Jones SA, Cressman EK, Henriques DY (2010) Proprioceptive localization of the left and right hands. Exp Brain Res 204:373–383
Kesten H (1958) Accelerated stochastic approximation. Ann Math Stat 29:41–59
Klassen J, Tong C, Flanagan JR (2005) Learning and recall of incremental kinematic and dynamic sensorimotor transformations. Exp Brain Res 164:250–259
Krakauer JW, Ghilardi MF, Ghez C (1999) Independent learning of internal models for kinematic and dynamic control of reaching. Nat Neurosci 2:1026–1031
Krakauer JW, Pine ZM, Ghilardi MF, Ghez C (2000) Learning of visuomotor transformations for vectorial planning of reaching trajectories. J Neurosci 20:8916–8924
Krakauer JW, Ghez C, Ghilardi MF (2005) Adaptation to visuomotor transformations: consolidation, interference, and forgetting. J Neurosci 25:473–478
Martin TA, Keating JG, Goodkin HP, Bastian AJ, Thach WT (1996) Throwing while looking through prisms. II. Specificity and storage of multiple gaze-throw calibrations. Brain 119(Pt 4):1199–1211
Mostafa AA, Salomonczyk D, Cressman EK, Henriques DY (2014) Intermanual transfer and proprioceptive recalibration following training with translated visual feedback of the hand. Exp Brain Res 232:1639–1651
Ostry DJ, Darainy M, Mattar AA, Wong J, Gribble PL (2010) Somatosensory plasticity and motor learning. J Neurosci 30:5384–5393
Redding GM, Wallace B (2004) First-trial adaptation to prism exposure: artifact of visual capture. J Mot Behav 36:291–304
Redding GM, Rossetti Y, Wallace B (2005) Applications of prism adaptation: a tutorial in theory and method. Neurosci Biobehav Rev 29:431–444
Salomonczyk D, Cressman EK, Henriques DY (2011) Proprioceptive recalibration following prolonged training and increasing distortions in visuomotor adaptation. Neuropsychologia 49:3053–3062
Salomonczyk D, Henriques DY, Cressman EK (2012) Proprioceptive recalibration in the right and left hands following abrupt visuomotor adaptation. Exp Brain Res 217:187–196
Salomonczyk D, Cressman EK, Henriques DY (2013) The role of the cross-sensory error signal in visuomotor adaptation. Exp Brain Res 228:313–325
Shadmehr R, Brashers-Krug T (1997) Functional stages in the formation of human long-term motor memory. J Neurosci 17:409–419
Shadmehr R, Smith MA, Krakauer JW (2010) Error correction, sensory prediction, and adaptation in motor control. Annu Rev Neurosci 33:89–108
Simani MC, McGuire LM, Sabes PN (2007) Visual-shift adaptation is composed of separable sensory and task-dependent effects. J Neurophysiol 98:2827–2841
Smith MA, Ghazizadeh A, Shadmehr R (2006) Interacting adaptive processes with different timescales underlie short-term motor learning. PLoS Biol 4:e179
Synofzik M, Thier P, Lindner A (2006) Internalizing agency of self-action: perception of one’s own hand movements depends on an adaptable prediction about the sensory action outcome. J Neurophysiol 96:1592–1601
Tong C, Wolpert DM, Flanagan JR (2002) Kinematics and dynamics are not represented independently in motor working memory: evidence from an interference study. J Neurosci 22:1108–1113
Treutwein B (1995) Adaptive psychophysical procedures. Vision Res 35:2503–2522
Wang J, Sainburg RL (2005) Adaptation to visuomotor rotations remaps movement vectors, not final positions. J Neurosci 25:4024–4030
Wolpert DM, Miall RC, Kawato M (1998) Internal models in the cerebellum. Trends Cogn Sci 2:338–347
Yamamoto K, Hoffman DS, Strick PL (2006) Rapid and long-lasting plasticity of input-output mapping. J Neurophysiol 96:2797–2801
Acknowledgments
We wish to thank Rozbeh Kamran-Disfani for his assistance with data collection. This research was supported by an NSERC Discovery Grant to DYPH and EKC.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Nourouzpour, N., Salomonczyk, D., Cressman, E.K. et al. Retention of proprioceptive recalibration following visuomotor adaptation. Exp Brain Res 233, 1019–1029 (2015). https://doi.org/10.1007/s00221-014-4176-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-014-4176-6