
Abstract
Rationale
The adverse consequences of human addictive drug use could be the result of either addictive drug consumption resulting in punishment (e.g., incarceration) or failure to engage in negative-reinforced behaviors that might compete with drug-maintained behaviors (e.g., contingency management strategies that reset payment amounts for drug free urines).
Objective
The goal of the present study was to establish a discrete-trial cocaine-vs-negative reinforcer (SNR) choice procedure where rats were presented with a simplified model of this conflict: choose negative reinforcement (i.e., escape or avoid foot shock) or choose an intravenous (IV) cocaine infusion followed by an inescapable shock.
Methods
Responding was maintained in male and female rats by IV cocaine infusions (0.32–1.8 mg/kg/inf) and a SNR (0.1–0.7 mA shock) under a discrete-trial concurrent “choice” schedule during daily sessions. Following parametric reinforcer magnitude and response requirement experiments, the effects of 12 h extended access cocaine self-administration and acute diazepam (0.32–10 mg/kg, IP) pretreatment were determined on cocaine-vs-SNR choice.
Results
Negative reinforcement was chosen over all cocaine doses. Lowering shock magnitude or increasing SNR response requirement failed to promote behavioral reallocation towards cocaine. Extended access cocaine self-administration sessions resulted in high daily cocaine intakes but failed to significantly increase cocaine choice in all (19) but one rat. Acute diazepam pretreatment also did not alter choice behavior up to doses that produced behavioral depression.
Conclusions
These results suggest that SNRs may be a source of reinforcement that effectively compete with and mitigate maladaptive addictive drug-maintained behaviors in the general population.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Continued drug use despite adverse consequences is one hallmark characteristic of addiction embedded into multiple clinical diagnostic criteria for substance use disorder. Adverse consequences of cocaine use could arise from a variety of different sources ranging from social stigma to cardiovascular pain or death. Cocaine-related overdose deaths have increased every year since 2010, earning cocaine use disorder the title of the “twin” or “silent” epidemic in relation to the current opioid crisis (Jones et al. 2017; Lipari and Park-Lee 2020; Fischer et al. 2021). Encounters with the adverse consequences of cocaine use may also motivate some individuals to seek treatment for cocaine use disorder (Fortney et al. 2010). To improve our fundamental knowledge of the basic mechanisms of continued cocaine use despite adverse consequences towards the development of candidate treatments, research efforts are increasingly utilizing preclinical procedures that model aspects of this clinical situation.
Addictive drug use despite adverse consequences in nonhumans has commonly been studied by pairing a self-administered intravenous (IV) drug infusion with a putative aversive stimulus, such as electric shock. Studies in both rodents and nonhuman primates consistently report that electric shock functions as a positive punisher and decreases cocaine self-administration under a broad range of experimental conditions (Grove and Schuster 1974; Bergman et al. 1981; Fontes and Shahan 2022; Durand et al. 2022). Furthermore, multiple studies report decreased sensitivity of cocaine self-administration to punishment after a history of extended cocaine self-administration, suggesting that extended access cocaine might reveal an “addiction phenotype” of continued drug use despite adverse consequences (Deroche-Gamonet et al. 2004; Pelloux et al. 2007; Kawa et al. 2019). However, these studies used single-operant drug self-administration procedures where the primary dependent measure is the rate of behavior and the environmental context included access to only a single reinforcer (i.e., IV drug infusion). The use of single-operant drug self-administration procedures poses interpretive complications that can be addressed with preclinical drug choice procedures (for review, see Negus and Banks 2021).
In a typical preclinical drug choice procedure, monkeys or rats have concurrent access to a drug reinforcer and a nondrug positive reinforcer (e.g., food or social interaction) (Johnson et al. 2016; Lile et al. 2016; Townsend et al. 2021; Venniro et al. 2021). In drug choice procedures, pairing drug-taking behavior with a positive punisher decreases drug self-administration and promotes behavioral reallocation towards the unpunished positive reinforcer (Johanson 1977; Negus 2005; Woolverton et al. 2012; Pelloux et al. 2015). In addition to these positive reinforcers, negative reinforcers are also available in our natural environment and may also compete with behavior maintained by addictive drugs. In contrast to a positive reinforcer which is operationally defined a stimulus whose presentation increases the likelihood of the operant response that preceded it (i.e., cocaine, food), a negative reinforcer (SNR) is defined as a stimulus whose removal increases the likelihood of the operant response that preceded it (Skinner 1938). An example of negative reinforcement in humans would be using a sterile syringe to prevent a secondary infection, where infection is the SNR. An example in rodents and nonhuman primates would be operant responding to escape or avoid the SNR of an electric foot shock (e.g., Sidman 1953; Berger and Robert Brush 1975; Babbini et al. 1979). In contrast to the extensive literature on drug-vs-positive reinforcer choice, behavioral allocation between IV drug infusions and a SNR has not been examined to the best of our knowledge.
The aim of the current study was to develop a novel discrete-trial cocaine-vs-SNR choice procedure to model a clinical situation in which failing to emit a negative-reinforced behavior results in presentation of the adverse consequence. An example of this type of conflict would be meeting your parole officer as scheduled to avoid jail time versus using cocaine, missing your parole officer meeting, and serving jail time. In the cocaine-vs-SNR choice procedure, rats were presented with a choice between responding to escape/avoid electric shock or responding for an IV cocaine infusion followed by an inescapable electric shock. There were three main experimental goals. One goal was to determine cocaine-vs-SNR choice sensitivity to independent variables such as reinforcer magnitude and response requirement that are effective in reallocating behavior under cocaine-vs-positive reinforcer choice conditions (Iglauer et al. 1976; Nader and Woolverton 1992; Negus 2003; Thomsen et al. 2013; Chow et al. 2022). A second goal was to determine the effects of extended access cocaine self-administration on cocaine-vs-SNR choice to test the hypothesis that extended cocaine access would increase cocaine choice followed by electric shock and decrease SNR choice. The final goal was to determine the effects of acute diazepam pretreatment on cocaine-vs-SNR choice to extend the literature on benzodiazepine effects on punished responding to include drug-choice conditions.
Materials and methods
Subjects
A total of 26 Sprague-Dawley rats (13M, 13F; Envigo, Frederick, MD) weighing 230–300 g upon arrival were used. Animals were singly housed in a temperature and humidity-controlled vivarium and maintained on a 12-h light/dark cycle (lights off at 6:00 pm). Water and food (Teklad Rat Diet, Envigo) were provided ad libitum in the home cage. Behavioral sessions were conducted 5 days per week from approximately 11 am–1 pm. Animal maintenance and research were conducted in accordance with the 2011 Guidelines of the National Institutes of Health Committee on Laboratory Animal Resources. Both enrichment and research protocols were approved by the Virginia Commonwealth University Institutional Animal Care and Use Committee.
Apparatus and catheter maintenance
Eight modular operant chambers located in sound-attenuating cubicles (Med Associates, St. Albans, VT) were equipped with electric grid floors (ENV-412 C and ENV-413C) and two retractable levers on the right chamber wall. A set of three LED lights (red, yellow, green) were mounted above the right, drug-associated lever. A white stimulus light was mounted above the left, negative reinforcement-associated lever. Rats were surgically implanted with a custom-made jugular catheter and vascular access port using previously described methods (Townsend et al. 2021). Intravenous (IV) cocaine was delivered by activation of a syringe pump (PHM-100, Med Associates) located inside the sound-attenuating cubicle. Liquid food-maintained responding training sessions occurred in operant chambers equipped with a retractable “dipper” cup (0.1 ml) positioned between the two levers. After each behavioral session, catheters were flushed with gentamicin (0.4 mg), followed by 0.1 ml of heparinized saline (10 U/ml). Catheter patency was verified at the end of each experiment by instantaneous muscle tone loss following IV methohexital (0.5 mg) administration.
Single-operant training
Behavioral history can influence operant responding maintained by both drug and non-drug reinforcers (Mckearney 1968; Barrett and Spealman 1978; Spealman 1979a). To address the potential influence of operant training history, the training order of negative reinforcement and cocaine self-administration was counterbalanced between rats. Twelve rats (6 M, 6 F) were initially trained on negative reinforcement (i.e., foot shock escape and avoidance) and 10 rats (5 M, 5 F) were initially trained on cocaine self-administration. To investigate potential effects of non-drug positive reinforcement training on acquisition of negative reinforced responding, a small group of four (2 M, 2 F) rats were initially trained on food-maintained responding, then negative reinforcement, and finally, cocaine self-administration. Final sample sizes are reported for each experiment.
Negative reinforcement training
Rats were initially trained by hand using successive approximation to lever-press to escape electric foot shock during daily 30-min sessions consisting of 60 trials. In each trial, a 3-s foot shock (0.4 mA) was presented along with the left lever and the associated white stimulus light above the lever. Responding was under a fixed-ratio (FR) 1 schedule of reinforcement such that a single response immediately terminated the shock, retracted the lever, and extinguished the stimulus light. The white house light was illuminated throughout all negative reinforcement training sessions. Acquisition criteria was defined as successful escape of ≥ 80% of the trials for 3 consecutive days. Once rats met escape acquisition criteria, rats were transitioned to an avoidance-training procedure that also consisted of 60 trials. In this avoidance procedure, shock presentation was preceded by a 30-s avoidance period during which the left, SNR-associated lever was extended, and white stimulus light was on. A single response (FR1) during the avoidance period canceled the upcoming shock for that trial, retracted the left lever, and extinguished the SNR stimulus light. If the rat failed to emit a response during the avoidance period, a 3-s shock (0.7 mA) was presented. During shock presentation, the left lever remained extended and the white SNR stimulus light remained illuminated, signaling the availability of an escape response. The shock intensity was increased to 0.7 mA during avoidance training because pilot studies suggested that this shock intensity resulted in the highest rate of acquiring avoidance behavior. Rats were trained on the avoidance procedure for a total of 5 consecutive days, and the number of escape and avoidance trails completed each day were recorded.
Cocaine self-administration training
Rats were trained to lever-press for an IV infusion of 0.32 mg/kg cocaine on the right lever under an initial FR1/20-s time out schedule of reinforcement during daily 2-hr sessions as previously described (Townsend et al. 2021). Each session began with a non-contingent cocaine infusion followed by a 60-s time out. The response period was signaled by extension of the right lever and illumination of the associated tricolor stimulus light above the lever. Following each response-requirement completion, the lever retracted, the stimulus light was extinguished, and an IV cocaine infusion was administered. Once rats earned ≥ 30 cocaine infusions during the 2-h session, the FR requirement was increased to FR3. Acquisition criteria was defined as ≥ 30 cocaine infusions under an FR3 schedule of reinforcement for 3 days. There was no upper limit to the number of cocaine infusions the rat could earn during the training sessions.
Food-maintained responding training
Rats were trained to lever press for a 5-s presentation of liquid food (32% chocolate-flavored Ensure™ diluted in water; Abbott Laboratories, Chicago, IL) on the right lever under an initial FR1/20-s time-out schedule of reinforcement during daily 2-h sessions as previously described (Townsend et al. 2021). Each session began with non-contingent food presentation followed by a 60-s time out. Liquid food availability was signaled by the illumination of the tricolor stimulus light above the right lever. After earning ≥ 30 food reinforcers during a 2-h session, the FR requirement was increased to FR3. Acquisition criteria was defined as ≥ 30 food reinforcers under an FR3 schedule of reinforcement for 3 days.
Cocaine-vs-SNR choice procedure
Following successful training of both SNR- and cocaine-maintained responding alone, rats were trained in the terminal discrete-trial cocaine-vs-SNR choice procedure. Daily behavioral sessions consisted of two forced trials followed by 9 discrete choice trials. The first forced trial was a cocaine-only trial during which the right, cocaine-associated lever and tricolor stimulus lights were presented. Response requirement (FR3) completion resulted in an IV infusion of the cocaine dose available during the subsequent choice trials. No shock was administered during the cocaine-only forced trial. The cocaine forced choice trial incorporated a 30-min limited hold, meaning if the response requirement was not met within 30-min, an IV cocaine infusion was administered non-contingently. Following the cocaine-only forced trial, there was a 4 min and 27 s time out during which all stimulus lights were extinguished, and all levers were retracted. Next, a negative reinforcement forced trial was initiated during which the left, SNR-associated lever was extended, and stimulus light was illuminated for 30-s. Response requirement (FR1) completion on the left lever during this 30-s period resulted in an avoidance response and canceled the upcoming shock. In the absence of an avoidance response, a 3-s shock stimulus (0.7 mA) was presented, and response requirement completion during the shock resulted in an escape response which immediately terminated the shock stimulus. Following completion of both forced trials, choice trials were initiated. During each of the nine choice trials, both the cocaine- and SNR-associated levers were extended, and the respective stimulus lights were illuminated for 30-s. During this 30-s response period, the rat could complete the response requirement on the cocaine-associated lever (FR3) for a cocaine infusion, immediately followed by a 3-s inescapable foot shock (0.7 mA), or the SNR-associated lever (FR1) to cancel the upcoming shock stimulus (i.e., avoidance response). Response requirement completion on either lever resulted in retraction of both levers, extinction of all stimulus lights, and initiation of a 4 min and 27 s time out period. If the response requirement was not met on either lever during the 30-s avoidance period, a 3-s electric shock (0.7 mA) was presented while both levers remained extended and stimulus lights remained on. During the 3-s electric shock, response requirement completion on the cocaine-associated lever resulted in a cocaine infusion with no shock termination (e.g., full 3-s shock), whereas response requirement completion on the SNR-associated lever resulted in immediate shock termination (i.e., escape response, < 3-s shock). If the response requirement was not completed on either lever after 3 s, all stimuli including shock were terminated, levers retracted, the trial was recorded as an omission, and a 4 min and 27 s time out period was initiated. Response requirement completion on the cocaine-associated lever was counted as a cocaine trial, and response requirement completion on the SNR-associated lever was counted as a SNR trial. Rats were tested in the choice procedure 5 days/week (Mon–Fri).
Experiment 1: Effect of cocaine dose on cocaine-vs-SNR choice
In the first experiment, three cocaine doses were tested to determine a cocaine-vs-SNR choice dose-effect function. Cocaine dose (0.32, 1.0, 1.8 mg/kg/inf) was varied by changing the infusion duration (e.g., 300g rat; 5, 15, 27 s of pump activation, respectively), and cocaine dose presentations were counterbalanced between rats. Based on the observed low number of cocaine trials completed across all three doses examined, a saline condition was not tested. Each cocaine dose was evaluated over a 6-day period. During the first 2 days, rats were tested at the given cocaine dose vs. SNR (0.7 mA). On the final 4 days, the shock stimulus was removed (i.e., no shock condition). The number of days tested under shock and no shock conditions were determined from pilot studies (see Figure S4). Results from the final 2 days of testing under each condition were averaged and used for data analysis.
Experiment 2: Effect of shock magnitude on cocaine-vs-SNR choice
The second experiment systematically determined effects of different shock magnitudes on cocaine-vs-SNR choice. Based on the results of experiment 1, 1.8 mg/kg/inf cocaine was used in the choice procedure for experiments 2–5. Shock magnitude was incrementally reduced and then increased every other day (0.7, 0.5, 0.3, 0.1, 0.3, 0.5, and 0.7 mA) across 14 test days. Shock intensity was manipulated through custom MedPC programming and verified by daily voltmeter measurements. Results from the second day of testing at each shock magnitude are reported and were used for data analysis.
Experiment 3: Effect of response requirement on cocaine-vs-SNR choice
Experiment 3 systematically manipulated the response requirement (1, 2, 4, 8, and 16) for the SNR using a between-day progressive-ratio (PR) schedule for 5 consecutive days. Shock magnitude (0.7 mA), cocaine dose (1.8 mg/kg/inf), and cocaine response requirement (FR3) parameters were held constant.
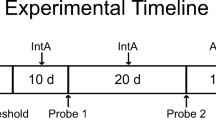
Experiment 4: Effect of extended cocaine access on cocaine-vs-SNR choice
Experiment 4 determined the effects of 12-hr extended access cocaine self-administration on cocaine-vs-SNR choice. Two separate cohorts of rats were used in this experiment. One cohort was tested in the cocaine-vs-SNR choice procedure at the 0.7 mA shock intensity, and the other was tested at a 0.3 mA shock intensity. Shock intensities were selected based on the results of experiment 2. Following baseline cocaine-vs-SNR choice, 12-h extended access cocaine self-administration sessions were introduced Sunday–Thursday for 2 consecutive weeks in addition to the daily cocaine-vs-SNR sessions conducted Monday–Friday. A timeline of this experiment is shown in Fig. 4A. Rats were placed in the operant chambers at 6 pm and could respond for 0.32 mg/kg/inf cocaine under a FR3/10-s time-out schedule of reinforcement with no consequent electric shock. The cocaine dose and session duration used were based on the literature (Wee et al. 2007). At approximately 6 am, rats were removed from the operant chambers and returned to their home cages. Body weights were assessed daily immediately prior to the choice session. After 2 weeks of extended cocaine access, a 1-week “washout” period occurred wherein only daily cocaine-vs-SNR choice sessions continued. Data collected on the Friday of this week served as the “post-extended access” data point for subsequent analyses.
Experiment 5: Effect of acute diazepam treatment on cocaine-vs-SNR choice
Following a week of baseline cocaine-vs-SNR choice after experiment 4, acute diazepam (vehicle, 0.32, 1.0, 3.2, and 10 mg/kg) treatment effects were determined. Presentation order of diazepam doses and vehicle were counterbalanced between rats and administered intraperitoneally 10 min before the choice session. A 1 day “washout” session was incorporated between each vehicle or diazepam dose in which no injections were administered but cocaine-vs-SNR choice sessions still occurred (data not shown). The same rats used in experiment 4 were used in this experiment to determine acute diazepam effects on cocaine-vs-SNR choice at either the 0.7 or 0.3 mA shock intensity, respectively.
Data analysis
The primary dependent measures in the discrete-trial cocaine-vs-SNR choice procedure were (1) cocaine trials completed, (2) SNR (both avoidance and escape responses) trials completed, and (3) omitted trials. These measures were plotted as a function of cocaine dose or independent variable manipulation. Other dependent measures included the latency to earn a cocaine infusion during the cocaine-only forced trial and the number of cocaine infusions earned during extended-access sessions. Data were analyzed using repeated-measures one- or two-way analysis of variance, or mixed-effects analysis as appropriate. In experiment 1, shock condition and cocaine dose were the main factors. In experiment 2, reinforcer trials completed (cocaine or SNR) and shock magnitude were the main factors. Omitted trials were analyzed separately. In experiment 3, reinforcer trials completed (cocaine or SNR) and response requirement were the main factors. Omitted trials were analyzed separately. In experiment 4, shock intensity group and experimental day were the main factors. In experiment 5, shock intensity group and diazepam dose were the main factors. Sphericity violations were corrected using the Greenhouse-Geisser epsilon. Significant main effects or interactions were followed by planned post hoc tests that corrected for multiple comparisons. The criterion for significance was set a priori at the 95% level of confidence (p < 0.05), and all analyses were conducted using GraphPad Prism (v 9.4.1, La Jolla, CA).
Drugs
(-)-Cocaine HCl was provided by the National Institute on Drug Abuse Drug Supply Program (Bethesda, MD, USA). Cocaine was dissolved in sterile saline for injection and passed through a 0.22-micron sterile filter before IV administration. Diazepam HCl solution was purchased from a commercial vendor (DASH Pharmaceuticals, Saddle River, NJ) and administered intraperitoneally 10 min before the session. Cocaine and diazepam doses are expressed as the salt form listed above.
Results
Cocaine and SNR training
A total of 21 rats (11 M, 10 F) completed both cocaine and SNR training and participated in the cocaine-vs-SNR choice experiments. No sex differences were observed during training. Nearly all rats (25/26) acquired escape responding (Table S1), escaping between 80 and 100% of the shock trials (Figure S1B). Consistent with the prior literature (Coyle et al. 1973; Kuribara and Tadokoro 1984), only a small subset (6/21) of rats acquired avoidance responding (Figure S2); however, because both escape and avoidance are SNR-maintained responses, subjects who acquired either escape or avoidance and cocaine self-administration participated in the cocaine-vs-SNR choice experiments. The avoidance contingency was maintained during the cocaine-vs-SNR choice experiments to allow for detection of any environmental or pharmacological manipulations that may alter avoidance behavior. There was a trend towards rats trained on cocaine first emitting more avoidance responses during training (Figure S2); however, avoidance behavior was variable throughout the five experiments (Table S2), making effects of training history on avoidance behavior difficult to determine. There was no significant effect of training history on acquisition of cocaine- or SNR-maintained responding as shown in Supplemental Table S1 and Figure S1.
Experiment 1: Effects of cocaine dose
Figure 1 shows choice trials completed for cocaine (0.32–1.8 mg/kg/inf), negative reinforcement, and trials omitted in the cocaine-vs-SNR choice procedure under both 0.7 mA shock (i.e., shock) and 0 mA shock (i.e., no shock) conditions. Panels A–C show results from the subset of rats classified as “Avoiders.” Rats were categorized as Avoiders if the subject emitted an avoidance response on at least four trials at each cocaine dose under shock conditions. Rats that emitted an avoidance response on three or fewer trials and instead emitted an escape response were classified as “Escapers,” and their results are shown in panels D–F. Under shock conditions, both avoider and escaper rats completed significantly more SNR trials than cocaine regardless of cocaine dose (Avoiders trial type: F(1, 3) = 68.6, p = 0.004; Escapers trial type: F(1.3, 21.3) = 28.1, p < 0.001). In the absence of electric shock, there was no significant change in the number of SNR or cocaine trials completed, nor omissions among Avoiders. In contrast, Escapers were sensitive to removal of electric shock such that cocaine trials increased (shock condition: F(1, 16) = 14.9, p = 0.0014) and SNR trials decreased (shock condition: F(1, 16) = 83.8, p < 0.0001). Omitted trials also increased in Escapers during shock removal (shock condition: F(1, 16) = 37.5, p < 0.0001). Cocaine dose-effect curves under shock and no shock conditions were also analyzed according to training history group (Figure S3), but less robust effects were observed.

Trials completed for cocaine (FR3), negative reinforcement (foot shock escape or avoidance, FR1), or omitted in a 9 discrete-trial cocaine-vs-negative reinforcement choice procedure as a function of cocaine dose. A–C Trials completed by “Avoider” (n = 4, 3F/1M) rats. D–F Trials completed by “Escaper” (n = 17, 7F/10M) rats. Filled symbols denote shock (0.7 mA) condition; open symbols denote no shock condition. All points represent mean ± SEM. Brackets represent significant main effect of shock condition. “Avoider/Escaper” classification: under shock conditions, “avoider” rats avoided at least 4 of the 9 trials at each cocaine dose tested. “Escaper” rats avoided 3 or fewer trials at each cocaine dose
Experiment 2: Effects of shock magnitude
Figure 2 shows the effect of manipulating shock magnitude on behavioral allocation between 1.8 mg/kg/inf cocaine and negative reinforcement. Rats were classified as Avoiders if at least four trials were avoided at each shock amplitude tested and classified as Escapers if three or fewer trials were avoided, and escape responses were emitted. Results from Avoiders revealed a significant main effect of reinforcer trial type (F(1, 5) = 10,588, p < 0.0001). The number of omitted trials remained low across all shock intensities in Avoiders. In contrast, reinforcer trials completed in Escapers was sensitive to shock magnitude (shock magnitude: F(3, 32.8) = 4.4, p = 0.01; reinforcer trial type: F(1, 11) = 24.7, p = 0.0004; reinforcer trial type × shock magnitude: F(3.1, 29.5) = 13.8, p < 0.0001; Fig. 2B). During initial testing, shock magnitudes ≥ 0.3 mA resulted in significantly more SNR trials completed than cocaine trials. There were never significantly more cocaine trials completed than SNR trials completed at any shock amplitude. Omitted trials did change as a function of shock magnitude in Escapers (F(3, 31) = 5.9, p = 0.003), however Dunnett’s post hoc test correcting for multiple comparisons did not detect any differences in omitted trials from the initial 0.7 mA testing. No sex differences were observed.

Effect of shock magnitude on cocaine-vs-negative reinforcer choice. Abscissae: shock magnitude. Ordinates: number of trials completed for cocaine, negative reinforcement, or omitted in the 9 discrete-trial choice procedure. All points represent mean ± SEM from the second test day. A Trials completed by “Avoider” (n = 6, 4F/2M) rats; brackets represent significant (p < 0.05) main effect of reinforcer trial type; B trials completed by “Escaper” (n = 12, 4F/8M) rats; *significant (p < 0.05) Sidak’s post hoc comparison between SNR and cocaine trials completed within a shock amplitude. “Avoider/Escaper” classification: under shock conditions, “avoider” rats avoided at least 4 of the 9 trials at each shock intensity tested. “Escaper” rats avoided 3 or fewer trials at each shock intensity
Experiment 3: Effects of response requirement
Figure 3 shows the effects of increasing the response requirement for the SNR on cocaine-vs-SNR choice. Due to a lack of differences in cocaine-vs-SNR choice behavior between avoider and escaper rats in experiments 3–5, the results for both phenotypes are shown together in Figs. 3, 4, and 5. Increasing the SNR response requirement resulted in decreased SNR trials completed (reinforcer type: F(2.7, 24.3) = 8.7, p = 0.0006; response requirement × reinforcer type: F(2.4, 20.3) = 14.2, p <0.0001) and an increase in omitted trials (F(2.7, 23.6) = 9.8, p = 0.0003). Cocaine trials completed were unaltered by manipulating the SNR response requirement. As an additional experiment, choice behavior was determined when the response requirement for both the SNR and 1.8 mg/kg/inf cocaine were equal at FR1 for 5 consecutive days. Figure S5 shows that trials completed for cocaine, negative reinforcement, or omitted were not different when the cocaine response requirement was either FR3 or FR1. No sex differences were observed.

Effect of negative reinforcer response requirement on cocaine-vs-negative reinforcer choice. All points represent the mean ± SEM. Symbols denote significant (p < 0.05) Dunnett’s post hoc comparisons within a trial type: *difference from FR1 SNR trials; #difference from FR1 omitted trials; n = 10 (6F/4M)

Effect of extended cocaine access on overnight cocaine intake, bodyweight, and choice trials completed for cocaine, negative reinforcer, or omitted. Abscissae: experimental day. A Schematic of experimental design; B number of cocaine infusions earned (FR3/TO10, 0.32 mg/kg/inf) during the overnight session; C latency in seconds to complete the initial cocaine-only trial. D–F Trials completed for cocaine, negative reinforcer, or omitted during the choice session. Filled and open circles denote rats tested in the cocaine-vs-negative reinforcer choice procedure under 0.7 mA (n = 12, 6F/6M), or 0.3 mA (n = 8, 4F/4M) shock condition, respectively. Baseline (BL) is the Friday prior to initiating extended access (EA) cocaine on the following Sunday. Post-EA is 7 days after terminating extended access sessions. All points represent the mean ± SEM. Brackets represent significant main effect of shock condition. Symbols denote significant (p < 0.05) Dunnett’s post hoc comparisons *different from day 1 in both shock intensity groups; #different from baseline only in the 0.7 mA shock group

Effect of diazepam pretreatment (0.32–10 mg/kg, ip) on cocaine-vs-negative reinforcer choice. Abscissae: diazepam dose in milligrams per kilogram administered intraperitoneally 10 min before the session. A Cocaine trials completed, B negative reinforcer trials completed, C omitted trials, and D latency to respond for the first cocaine infusion. Filled and open symbols denote rats tested at the 0.7 mA (n = 7, 4F/3M) and 0.3 mA (n = 5, 2F/3M) shock condition, respectively. All points represent the mean ± SEM. Symbols represent significant (p < 0.05) Dunnett’s post hoc comparisons: #different from vehicle only in the 0.7 mA shock group; *different from vehicle in both shock intensity groups
Experiment 4: Effects of extended cocaine access
Figure 4A shows the effects of 12-h extended access cocaine (0.32 mg/kg/inf, FR3/10-s time out) self-administration on cocaine-vs-SNR choice in two cohorts that differed in shock magnitude (0.7 vs. 0.3 mA). Cocaine daily intake during the extended access sessions was approximately 96 mg/kg/day (Fig. 4B), and only a significant decrease in cocaine infusions was observed on day 3 compared to day 1 in the 0.7 mA shock intensity group (F(3.4, 32.5) = 9.3, p < 0.0001; Fig. 4B). Bodyweight also decreased from baseline in both shock intensity groups (0.7 mA shock condition: F(2.9, 28.3) = 13.5, p < 0.0001; 0.3 mA shock condition: F(1.9, 13.5) = 5.9, p = 0.01, data not shown). Figure 4C shows the latency to complete the FR3 response requirement for cocaine during the first forced trial in each shock intensity group. Latency significantly increased during the extended-access period in the 0.7 mA shock intensity group (F(3.6, 33.3) = 4.4, p = 0.007). Extended cocaine access did not significantly alter cocaine trials completed under either the 0.7 or 0.3 mA shock condition (Fig. 4D). More SNR trials were completed under the 0.7 mA shock condition compared to the 0.3 mA shock condition, and there was no significant effect of extended cocaine access (shock condition: F(1, 11) = 12, p = 0.005; Fig. 4E). There were more omitted trials in the 0.3 mA shock condition compared to the 0.7 mA shock condition (shock condition: F(1, 11) = 15.6, p = 0.002; Fig. 4F). Although there was a main effect of experimental day on omitted trials (experimental day: F(4, 43.9) = 2.9, p = 0.033), post hoc analysis correcting for multiple comparisons did not detect significant changes from baseline omissions in either shock intensity group. In addition, no sex differences were observed (Figures S6–7). Individual subject analysis reveals that of the 20 rats tested, only one rat in the 0.7 mA shock condition increased cocaine choice following extended cocaine access (Figure S8), increasing from zero to eight cocaine trials completed.
Experiment 5: Effects of acute diazepam treatment
Figure 5 shows acute diazepam effects on cocaine-vs-SNR choice under both 0.7 and 0.3 mA shock conditions. More cocaine trials were completed under 0.3 than 0.7 mA shock conditions (shock condition: F(1, 6) = 8.1, p = 0.029; Fig. 5A). Although there was a main effect of diazepam dose (dose: F(1.8, 10.7) = 6.1, p = 0.02), diazepam did not alter cocaine trials completed upon post hoc analysis that corrected for multiple comparisons. More SNR trials were completed under the 0.7 mA shock condition than the 0.3 mA shock condition (shock condition: F(1, 6) = 13.3, p = 0.011; Fig. 5B). There was a main effect of diazepam dose (dose: F(1.4, 8.3) = 18.9, p = 0.001) and 10 mg/kg diazepam decreased SNR trials completed under 0.7mA shock condition (F(2.2, 12.7) = 26.4, p <0.0001; Fig. 5B). More trials were omitted in the 0.3 than 0.7 mA shock condition (shock condition: F(1, 6) = 8.9, p = 0.025; Fig. 5C). There was a main effect of diazepam dose (dose: F(1.9, 11.5) = 108.6, p < 0.0001), and 10 mg/kg diazepam increased omissions under both shock conditions (0.7mA shock condition: F(1.4, 7.8) = 115.2, p < 0.0001; 0.3 mA shock condition: F(2.2, 9.0) = 29.75, p < 0.0001). Furthermore, there was a main effect of diazepam dose on latency to complete the FR3 response requirement for a cocaine infusion during the initial forced trial component (dose: F(2.6, 15.9) = 25.9, p < 0.001) such that the start latency was significantly increased relative to vehicle after 3.2 and 10 mg/kg diazepam in the 0.7 mA shock cohort (F(1.7, 11.7) = 69.9, p < 0.001; Fig. 5D). All rats reached the 30-min limited hold following 10 mg/kg diazepam. No sex differences were observed.
Discussion
The present study established a discrete-trial cocaine-vs-SNR choice procedure in male and female rats to test hypotheses related to drug taking despite adverse consequences. We then determined the sensitivity of cocaine-vs-SNR choice to environmental and pharmacological manipulations previously shown to impact cocaine-vs-positive reinforcer (e.g., food or social interaction) choice. There were three main findings. First, rats completed more SNR trials than cocaine trials under a broad range of experimental conditions. The lack of behavioral reallocation away from the SNR and towards cocaine in response to manipulations of reinforcer magnitude and response requirement suggests cocaine and a SNR may be economic independents rather than economic substitutes. Second, extended cocaine self-administration sessions resulted in high cocaine intake but failed to promote behavioral reallocation towards cocaine and away from the SNR. These results do not support the hypothesis that extended cocaine self-administration leads to an addiction-like phenotype of increased cocaine-taking behavior despite adverse consequences in a choice context (Deroche-Gamonet et al. 2004). Finally, acute diazepam pretreatment failed to increase cocaine-vs-SNR choice up to behavioral-depressant doses. These findings stand in contrast to acute benzodiazepine effects on punished responding under single-operant conditions (Spealman 1979b; Hymowitz and Abramson 1983; Dworkin et al. 1989; Howard 1990; Panlilio et al. 2005), and suggest that drug-choice studies provide additional and complementary information on reinforcement processes compared to single-operant drug self-administration. Overall, the results suggest that SNRs in the clinical environment may be a source of reinforcement for the general population that competes with and mitigates maladaptive addictive drug-maintained behaviors.
A concurrently available SNR resulted in low levels of cocaine self-administration
A concurrently available SNR (i.e., shock) resulted in very few cocaine choices. Previous studies have demonstrated that concurrent availability of a non-drug positive reinforcer such as social interaction (Venniro et al. 2021) or saccharin (Lenoir et al. 2007) can exert a protective effect on cocaine self-administration within a discrete-trial choice context, and the current study extends these finding to include a SNR. In addition, behavioral allocation between cocaine and non-drug positive reinforcers has shown sensitivity to positive punishment with either electric shock (Johanson 1977) or IV histamine (Negus 2005; Woolverton et al. 2012) such that behavior was reallocated away from the reinforcer paired with the punisher and towards the alternative, unpunished reinforcer. Consistent with prior punishment studies, rats allocated the majority of their behavior to the unpunished SNR rather than the punished cocaine infusion. The present study observed minimal behavioral allocation between the cocaine and SNR across a range of cocaine doses and shock amplitudes. Surprisingly, even at small shock amplitudes or high SNR response requirements, rats omitted trials despite receiving electric shock rather than reallocating their behavior to the cocaine-associated lever. Given previous cocaine choice studies in both nonhuman primates and rats which demonstrate behavioral reallocation between cocaine and another positive reinforcer in response to parametric manipulations of reinforcer magnitude and response requirement (Iglauer et al. 1976; Nader and Woolverton 1992; Negus 2003; Thomsen et al. 2013; Chow et al. 2022), the lack of behavioral reallocation observed in the present study was unexpected. There are several potential explanations for this result, two of which will be discussed.
One potential explanation is the extension of both the cocaine and SNR -associated levers together during the choice trials resulted in the levers becoming conditioned aversive stimuli. Freezing is an established rodent defense reaction to conditioned aversive stimuli (Bolles 1970), and the increase in omitted trials in the present study could be due to freezing responses. Several lines of evidence in the present results argue against this explanation. First, each rat was individually trained to lever-press in response to the shock stimulus and this training resulted in high success rates of operant escape responding (Supplemental Table 1, Figure S1). Additionally, SNR-maintained responding was high except under conditions of low shock magnitude or high SNR response requirement. Furthermore, the completion of the forced trials for both the cocaine and SNR at the beginning of the choice session under most experimental conditions provide additional evidence that at least single lever presentation was still associated with an operant response and reinforcer presentation.
A more likely explanation for the lack of behavioral reallocation between the cocaine and SNR in response to parametric manipulations is rooted in behavioral economics and the degree to which commodities, including drugs as reinforcers, function as substitutes, complements, or independents (for review, see Bickel et al. 2014). Numerous studies have shown that increasing the magnitude of the food reinforcer promotes behavioral allocation towards food and away from cocaine. Inversely, increasing the available cocaine dose promotes behavioral allocation towards cocaine choice (Nader and Woolverton 1991; Negus 2003; Thomsen et al. 2013). This same relationship also holds for response requirement manipulations (Nader and Woolverton 1992; Thomsen et al. 2013). In behavioral economic terms, food and cocaine when concurrently available as positive reinforcers function as economic substitutes (Bickel et al. 2014). In the current study, behavioral allocation between cocaine and a SNR was minimally sensitive to reinforcer magnitude and response requirement manipulations, suggesting that cocaine and the SNR were not economic substitutes. Rather, cocaine and the SNR in this choice context appeared to be economic independents (Bickel et al. 2014).
Overall, the present results suggest that although negative reinforcement may result in low-levels of cocaine self-administration, it is not an economic substitute for cocaine. Considering this, treatment strategies that utilize a combination of both nondrug positive and negative reinforcement contingencies might be most effective for treatment of cocaine use disorder. Additionally, there is currently debate in the literature regarding whether the distinction between positive and negative reinforcement is empirically founded and involve distinctly different processes (Michael 1975; Perone 2003; Baron and Galizio 2005; Nevin and Mandell 2017). The present choice results support a distinction between positive and negative reinforcement and provide empirical evidence to support future research on both the quantitative and qualitative differences between positive and negative reinforcers.
Extended cocaine access failed to increase cocaine-vs-SNR choice
Increased drug availability through extended access drug self-administration conditions is one common preclinical method to achieve high levels of cocaine intake (Ahmed 2011). Cocaine self-administration under extended access conditions is hypothesized to be associated with the transition to persistent drug-taking behavior despite adverse consequences (Vanderschuren and Everitt 2004; Ahmed 2011), as preclinical studies have demonstrated that extended cocaine access can lead to a decreased sensitivity to shock-associated punishment of cocaine reinforcement in a subset of rats (Deroche-Gamonet et al. 2004; Pelloux et al. 2007; Belin et al. 2009). The present results show that male and female rats achieved high levels of cocaine intake during extended access sessions; however, in contrast to these previous findings and despite the use of similar shock magnitudes in previous studies (Deroche-Gamonet et al. 2004), extended cocaine access failed to increase cocaine-vs-SNR choice. Additionally, out of 20 total rats, only a single rat completed more cocaine trials during the choice session after the extended cocaine access sessions. These results do not support the hypothesis that extended cocaine access leads to increased or persistent cocaine self-administration despite adverse consequences when the adverse consequence is also a SNR. Furthermore, the latency to complete the cocaine response requirement during the initial forced trial component of the choice procedure increased during some extended access conditions (Fig. 4C). If cocaine withdrawal served as an internal SNR such that cocaine self-administration would alleviate this internal SNR (Koob, 2015), this hypothesis would predict unpunished cocaine self-administration latencies to decrease during extended access conditions. Our results indicate the opposite effect, and additionally, rats maintained high levels of SNR trials completed during the extended access period. These results suggest that the exogenous SNR of shock was a more effective reinforcer than cocaine self-administration to alleviate a hypothesized negative internal state. Moreover, the present results are consistent with and extend previous cocaine-vs-food choice studies in rhesus monkeys demonstrating extended access cocaine self-administration failed to increase cocaine choice (Banks and Negus 2010). Overall, these data suggest that persistent cocaine self-administration despite adverse consequences may be driven more by a lack of alternative reinforcers in the environment (Ahmed et al. 2011).
Acute diazepam did not increase cocaine-vs-SNR choice
Under single-operant behavioral conditions, benzodiazepine administration increases punished responding maintained by multiple reinforcer types (Spealman 1979b; Hymowitz and Abramson 1983; Dworkin et al. 1989; Howard 1990) including addictive drugs (Panlilio et al. 2005). Therefore, we hypothesized that acute diazepam pretreatment would increase cocaine choice in the cocaine-vs-negative reinforcer choice procedure. However, diazepam pretreatment failed to increase cocaine-vs-SNR choice, suggesting that benzodiazepines may only increase punished responding under single-operant contingencies. Diazepam administration before a cocaine-vs-food choice session in rats has been shown to decrease cocaine choice and increase food choice in the absence of any punishment contingencies (Augier et al. 2012). Overall, these results add to the current literature highlighting the importance of incorporating alternative reinforcers into drug self-administration models, and that results from single-operant drug self-administration procedures do not always translate to drug self-administration procedures that include concurrent availability of another reinforcer. Future experiments should evaluate diazepam pretreatment effects on punished drug-vs-alternative positive reinforcer choice procedures to improve our understanding of these differential benzodiazepine effects.
References
Ahmed SH (2011) Animal models of drug addiction. In: Neuromethods, vol 53. Humana Press
Augier E, Vouillac C, Ahmed SH (2012) Diazepam promotes choice of abstinence in cocaine self-administering rats. Addict Biol 17:378–391. https://doi.org/10.1111/j.1369-1600.2011.00368.x
Babbini M, Gaiardi M, Bartoletti M (1979) Stimulus-response relationships in a quickly learned escape from shock: effects of morphine. Pharmacol Biochem Behav 11:155–158
Banks ML, Negus SS (2010) Effects of extended cocaine access and cocaine withdrawal on choice between cocaine and food in rhesus monkeys. Neuropsychopharmacology 35:493–504. https://doi.org/10.1038/npp.2009.154
Baron A, Galizio M (2005) Positive and negative reinforcement: should the distinction be preserved? Behav Anal 28:85–98
Barrett JE, Spealman RD (1978) Behavior simultaneously maintained by both presentation and termination of noxious stimuli. J Exp Anal Behav 3:375–383
Belin D, Balado E, Piazza PV, Deroche-Gamonet V (2009) Pattern of intake and drug craving predict the development of cocaine addiction-like behavior in rats. Biol Psychiatry 65:863–868. https://doi.org/10.1016/j.biopsych.2008.05.031
Berger DF, Robert Brush F (1975) Rapid acquisition of discrete-trial lever-press avoidance: effects of signal-shock interval. J Exp Anal Behav 24:227–239
Bergman J, Johanson CE, Bergman J, Johanson CE (1981) The effects of electric shock on responding maintained by cocaine in rhesus monkeys. Pharmacol Biochem Behav 14:423–426
Bickel WK, Johnson MW, Koffarnus MN et al (2014) The behavioral economics of substance use disorders: reinforcement pathologies and their repair. Annu Rev Clin Psychol 10:641–677
Bolles RC (1970) Species-specific defense reactions and avoidance learning. Psychol Rev 77:32–48
Chow JJ, Beacher NJ, Chabot JM et al (2022) Characterization of operant social interaction in rats: effects of access duration, effort, peer familiarity, housing conditions, and choice between social interaction vs. food or remifentanil. Psychopharmacology (Berl) 239:2093–2108. https://doi.org/10.1007/s00213-022-06064-1
Coyle JT, Wender P, Lipsky A (1973) Avoidance conditioning in different strains of rats: neurochemical correlates. Psychopharmacologia (Berl) 31:25–34
Deroche-Gamonet V, Belin D, Piazza PV (2004) Evidence for addiction-like behavior in the rat. Science 305:1014–1017
Durand A, Girardeau P, Freese L, Ahmed SH (2022) Increased responsiveness to punishment of cocaine self-administration after experience with high punishment. Neuropsychopharmacology 47:444–453. https://doi.org/10.1038/s41386-021-01159-3
Dworkin SI, Bimle C, Miyauchi T (1989) Differential effects of pentobarbital and cocaine on punished and nonpunished responding. J Exp Anal Behav 51:173–184
Fischer B, O’Keefe-Markman C, Lee A, Daldegan-Bueno D (2021) ‘Resurgent’, ‘twin’ or ‘silent’ epidemic? A select data overview and observations on increasing psycho-stimulant use and harms in North America. Subst Abuse Treat Prev Policy 16:1–11. https://doi.org/10.1186/s13011-021-00350-5
Fontes RM, Shahan TA (2022) Effects of repeated exposure to escalating versus constant punishment intensity on response allocation. J Exp Anal Behav 118(1):59–82. https://doi.org/10.1002/jeab.766
Fortney JC, Tripathi SP, Walton MA et al (2010) Patterns of substance abuse treatment seeking following cocaine-related emergency department visits. J Behav Health Serv Res 38:221–233
Grove RN, Schuster CR (1974) Suppression of cocaine self-administration by extinction and punishment. Pharmacol Biochem Behav 2:199–208
Howard JL (1990) Effects of drugs on punished behavior: pre-clinical test for anxiolytics. Pharmacol Ther 45:403–424
Hymowitz N, Abramson M (1983) Effects of diazepam on responding suppressed by response-dependent and independent electric-shock delivery. Pharmacol Biochem Behav 18:769–776
Iglauer C, Llewellyn ME, Woods JH (1976) Concurrent schedules of cocaine injection in rhesus monkeys: dose variations under independent and non-independent variable-interval procedures. Pharmacol Rev 27:367–383
Johanson CE (1977) The effects of electric shock on responding maintained by cocaine injections in a choice procedure in the rhesus monkey. Psychopharmacology (Berl) 53:277–282
Johnson AR, Banks ML, Blough BE et al (2016) Development of a translational model to screen medications for cocaine use disorder I: choice between cocaine and food in rhesus monkeys. Drug Alcohol Depend 165:103–110. https://doi.org/10.1016/j.drugalcdep.2016.05.021
Jones CMC, Baldwin GT, Compton WM (2017) Recent increases in cocaine-related overdose deaths and the role of opioids. Am J Public Health 107:430–432
Kawa AB, Allain F, Robinson TE, Samaha AN (2019) The transition to cocaine addiction: the importance of pharmacokinetics for preclinical models. Psychopharmacology (Berl) 236:1145–1157. https://doi.org/10.1007/s00213-019-5164-0
Koob GF (2015) The dark side of emotion: the addiction perspective. Eur J Pharmacol 753:73–87
Kuribara H, Tadokoro S (1984) Conditioned lever-press avoidance response in mice: acquisition processes and effects of diazepam. Psychopharmacology (Berl) 82:36–40
Lenoir M, Serre F, Cantin L, Ahmed SH (2007) Intense sweetness surpasses cocaine reward. PLoS One. 2(8):e698. https://doi.org/10.1371/journal.pone.0000698
Lile JA, Stoops WW, Rush CR et al (2016) Development of a translational model to screen medications for cocaine use disorder II: choice between intravenous cocaine and money in humans. Drug Alcohol Depend 165:111–119. https://doi.org/10.1016/j.drugalcdep.2016.05.022
Lipari RN, Park-Lee E (2020) Key substance use and mental health indicators in the United States: results from the 2019 National Survey on Drug Use and Health
Mckearney JW (1968) Maintenance of responding under a fixed-interval schedule of electric shock-presentation. Science 160:1249–1251
Michael J (1975) Positive and negative reinforcement, a distinction that is no longer necessary; or a better way to talk about bad things. Behaviorism 3:33–44
Nader MA, Woolverton WL (1991) Effects of increasing the magnitude of an alternative reinforcer on drug choice in a discrete-trials choice procedure. Psychopharmacology (Berl) 105:169–174. https://doi.org/10.1007/BF02244304
Nader MA, Woolverton WL (1992) Effects of increasing response requirement on choice between cocaine and food in rhesus monkeys. Psychopharmacology (Berl) 108:295–300. https://doi.org/10.1007/BF02245115
Negus SS (2003) Rapid assessment of choice between cocaine and food in rhesus monkeys: effects of environmental manipulations and treatment with d-amphetamine and flupenthixol. Neuropsychopharmacology 28:919–931. https://doi.org/10.1038/sj.npp.1300096
Negus SS (2005) Effects of punishment on choice between cocaine and food in rhesus monkeys. Psychopharmacology (Berl) 181:244–252. https://doi.org/10.1007/s00213-005-2266-7
Negus SS, Banks ML (2021) Confronting the challenge of failed translation in medications development for substance use disorders. Pharmacol Biochem Behav 210:173264. https://doi.org/10.1016/j.pbb.2021.173264
Nevin JA, Mandell C (2017) Comparing positive and negative reinforcement: a fantasy experiment. J Exp Anal Behav 107:34–38. https://doi.org/10.1002/jeab.237
Panlilio LV, Thorndike EB, Schindler CW (2005) Lorazepam reinstates punishment-suppressed remifentanil self-administration in rats. Psychopharmacology (Berl) 179:374–382. https://doi.org/10.1007/s00213-004-2040-2
Pelloux Y, Everitt BJ, Dickinson A (2007) Compulsive drug seeking by rats under punishment: effects of drug taking history. Psychopharmacology (Berl) 194:127–137. https://doi.org/10.1007/s00213-007-0805-0
Pelloux Y, Murray JE, Everitt BJ (2015) Differential vulnerability to the punishment of cocaine related behaviours: effects of locus of punishment, cocaine taking history and alternative reinforcer availability. Psychopharmacology (Berl) 232:125–134. https://doi.org/10.1007/s00213-014-3648-5
Perone M (2003) Negative effects of positive reinforcement. Behav Anal 26:1–14
Sidman M (1953) Avoidance conditioning with brief shock and no exteroceptive warning signal. Science 118:157–158
Skinner BF (1938) The behavior of organisms: an experimental analysis. Appleton-Century-Crofts, New York
Spealman RD (1979a) Behavior maintained by termination of a schedule of self-administered cocaine. Science 204:1231–1233
Spealman RD (1979b) Comparison of drug effects on responding punished by pressurized air or electric shock delivery in squirrel monkeys: pentobarbital, chlordiazepoxide, d-amphetamine and cocaine. J Pharmacol Exp Ther 209:309–315
Thomsen M, Barrett AC, Negus SS, Caine SB (2013) Cocaine versus food choice procedure in rats: environmental manipulations and effects of amphetamine. J Exp Anal Behav 99:211–233. https://doi.org/10.1002/jeab.15
Townsend SKL, Robinson HL et al (2021) A drug-vs-food “choice” self-administration procedure in rats to investigate pharmacological and environmental mechanisms of substance use disorders. J Neurosci Methods 354:109110. https://doi.org/10.1016/j.jneumeth.2021.109110
Vanderschuren LJMJ, Everitt BJ (2004) Drug seeking becomes compulsive after prolonged cocaine self-administration. Science 305:1017–1019. https://doi.org/10.1126/science.1098975
Venniro M, Panlilio LV, Epstein DH, Shaham Y (2021) The protective effect of operant social reward on cocaine self-administration, choice, and relapse is dependent on delay and effort for the social reward. Neuropsychopharmacology 46:2350–2357. https://doi.org/10.1038/s41386-021-01148-6
Wee S, Specio SE, Koob GF (2007) Effects of dose and session duration on cocaine self-administration in rats. J Pharmacol Exp Ther 320:1134–1143. https://doi.org/10.1124/jpet.106.113340
Woolverton WL, Freeman KB, Myerson J, Green L (2012) Suppression of cocaine self-administration in monkeys: effects of delayed punishment. Psychopharmacology (Berl) 220:509–517. https://doi.org/10.1007/s00213-011-2501-3
Acknowledgements
We appreciate the technical assistance of Michelle Arriaga during the training procedures.
Funding
Research reported in this publication was supported by the National Institute on Drug Abuse (NIDA) under award numbers T32DA007027 and R21DA053820. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIDA. The funding source had no role in the experimental design, interpretation, or decision to publish the results.
Author information
Authors and Affiliations
Contributions
Madison Marcus: writing—original draft, writing—review and editing, methodology, software, formal analysis, investigation, data curation, and data visualization. Matthew Banks: formal analysis, writing—review and editing, visualization, conceptualization, supervision, and funding acquisition.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
ESM 1
Supplemental materials include additional data tables and figures. (DOCX 464 kb)
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Marcus, M.M., Banks, M.L. Effects of environmental and pharmacological manipulations on cocaine-vs-negative reinforcer choice in male and female rats. Psychopharmacology 240, 1677–1689 (2023). https://doi.org/10.1007/s00213-023-06404-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-023-06404-9