
Abstract
Rationale
Drug-related cues evoke craving and stimulate motivational systems in the brain. The acoustic startle reflex captures activation of these motivational processes and affords a unique measure of reactivity to drug cues.
Objectives
This study examined the effects of cannabis-related cues on subjective and eye blink startle reactivity in the human laboratory and tested whether these effects predicted youth’s cue-elicited cannabis craving in the natural environment.
Methods
Participants were 55 frequent cannabis users, ages 16 to 24 years (M = 19.9, SD = 1.9; 55% male; 56% met criteria for cannabis dependence), who were recruited from a clinical trial to reduce cannabis use. Eye blink electromyographic activity was recorded in response to acoustic probes that elicited startle reactivity while participants viewed pleasant, unpleasant, neutral, and cannabis picture cues. Following the startle assessment, participants completed an ecological momentary assessment protocol that involved repeated assessments of cue-elicited craving in real time in their real-world environments.
Results
Multilevel models included the presence or absence of visible cannabis cues in the natural environment, startle magnitude, and the cross-level interaction of cues by startle to test whether cue-modulated startle reactivity in the laboratory was associated with cue-elicited craving in the natural environment. Analyses showed that cannabis-related stimuli evoked an appetitive startle response pattern in the laboratory, and this effect was associated with increased cue-elicited craving in the natural environment, b = − 0.15, p = .022, 95% CI [− 0.28, − 0.02]. Pleasant stimuli also evoked an appetitive response pattern, but in this case, blunted response was associated with increased cue-elicited craving in the natural environment, b = 0.27, p < .001, 95% CI [0.12, 0.43].
Conclusions
Our findings support cue-modulated startle reactivity as an index of the phenotypic expression of cue-elicited cannabis craving.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Cannabis is the most widely used internationally regulated drug by youth worldwide (Johnston et al. 2013; Substance Abuse and Mental Health Services Administration 2015). Although the perceived harmfulness of cannabis use among youth decreased over the past two decades, there is strong evidence that cannabis use among adolescents is not benign (Keyes et al. 2016). Frequent, long-term, or heavy cannabis use among youth has acute and lasting adverse consequences, including cognitive impairments, disruption of brain functioning and development, and liability for addiction (Volkow et al. 2014). Understanding factors associated with cannabis misuse among youth is essential for understanding the pathogenesis of cannabis use disorder (CUD) and developing treatments for CUD; however, research investigating these mechanisms among youth is still in its infancy.
Craving, or urge to use a drug, is central to contemporary models of addiction (Drummond 2001). Addictive substances, including cannabis, produce a host of pleasurable effects by potentiating dopaminergic activity in reward regions of the brain (Volkow, Koob, & McLellan, Volkow et al. 2016). This increase in dopamine release promotes associative learning that over time triggers activation of the brain’s reward and appetitive motivational systems in response to environmental stimuli paired with substance use. This anticipatory response (i.e., “cue reactivity”) to conditioned stimuli elicits craving and heightens the probability and intensity of subsequent substance use among adolescents and young adults (Ramirez and Miranda 2014). Indeed, subjective and physiological reactivity to drug cues are thought to reflect motivational processes that both maintain substance use and increase the risk for relapse (Sayette 2016; Tiffany and Wray 2012).
Experimentally, researchers capture these motivational processes in the human laboratory by exposing participants to drug cues associated with their substance use across a variety of modalities (pictures, videos, in vivo). This methodology reliably induces changes in subjective craving and physiological and neurobiological reactivity (Carter and Tiffany 1999). In two studies, Gray et al. (2008, 2011) found that teenagers who met criteria for CUD displayed increased skin conductance to cannabis cues, particularly in vivo cues, relative to neutral cues. Neuroimaging research showed increased activation in the limbic and paralimbic systems as well as the visual cortex among cannabis users while viewing cannabis-related images (Charboneau et al. 2013). Among frequent cannabis users, one study found activation in the reward-related areas of the brain (i.e., the orbitofrontal cortex, anterior cingulate cortex, and striatum) differentiated between individuals with high versus low problematic cannabis use severity (Goldman et al. 2013). Henry et al. (2014) also found that increased cannabis use was associated with correspondingly increased event-related potentials, specifically the motivation-based P300 response when exposed to cannabis cues. On the whole, these studies suggest that cannabis users show consistent reactivity to cannabis-related cues across several neurobiological measures, particularly those related to motivation and reward.
An important gap in our understanding of cue reactivity effects, particularly reactivity captured via neurobiological measures, is whether this phenomenon predicts real-world behaviors in the natural environment. Consequently, the clinical implications of laboratory-based cue reactivity methods remain unknown. This study examined the effects of cannabis-related cues on subjective and eye blink startle reactivity in the human laboratory and tested whether these effects predicted youth’s cue-elicited cannabis craving in the natural environment. By pairing responses from the human laboratory with the natural environment, we aimed to provide a more comprehensive and rigorous understanding of the emotional and attentional mechanisms underlying cannabis use and addiction among youth.
As a unique measure of appetitive reactivity to drug cues, the acoustic startle reflex may provide a useful measure of motivation in response to cannabis cues (for a review, see Meehan & Miranda, Meehan and Miranda Jr 2013). Startle reactivity occurs in response to a sudden noise blast that results in an abrupt contraction of skeletal muscles. This response is sensitive to emotion-laden stimuli; the magnitude of the response is attenuated for pleasant (or appetitive) stimuli and potentiated for unpleasant (or aversive) stimuli (Bradley and Lang 2000), thus providing an index of a cue’s emotional valence. Importantly, affective modulation of the startle response contrasts with other physiological measures of cue reactivity (e.g., skin conductance, heart rate) that tend to increase as an individual’s emotional arousal intensifies, regardless of valence. The startle reflex is inhibited by drug pictorial cues compared to other pictorial cues (e.g., unpleasant), which indicates an appetitive startle response to drug cues (e.g., alcohol, Rubio et al. 2011; and tobacco, Cui et al. 2012; Lam et al. 2012). For example, research with patients with alcohol dependence has found that alcohol-cue modulated startle responses were lower than startle responses of negative and neutral cues (i.e., appetitive response; Leménager et al. 2014; Rubio et al. 2011) and they were comparable to positive cues (Rubio et al. 2011). Similar patterns are documented among cigarette smokers: cigarette and positive cues had startle responses that were comparable to each other and both cues induced lower startle responses than negative and neutral cues (Cui et al. 2012). However, limited research has examined these implications among youth and within the context of cannabis craving.
In order to evaluate the cannabis cue-modulated startle response and whether cannabis craving in the human laboratory predicts real-world craving among youth, this study paired laboratory and ecological momentary assessment (EMA) methods. EMA provides a real-time assessment of participants’ behaviors and experiences in their natural environment (Shiffman, Stone, & Hufford, Shiffman et al. 2008), which are valuable in investigating the implications of controlled laboratory effects on real-world behavior. Consistent with prior research on startle response and drug cues (Cui et al. 2012; Rubio et al. 2011), we hypothesized that cannabis pictorial cues would produce a more appetitive startle response, similar to a pleasant pictorial cue. Second, we hypothesized that attenuated (more appetitive) startle response to cannabis pictorial cues in the laboratory would predict greater craving in response to cannabis cues in the natural environment.
Materials and methods
Participant selection
Eighty-six eligible adolescents and young adults were recruited from the community to participate in a clinical trial testing the effects of a medication in combination with a psychosocial intervention on cannabis use (Miranda et al. 2017). Inclusion criteria were 15 to 24 years old; cannabis use at least twice weekly in the past 30 days; and able to read simple English. Exclusion criteria were mandated to treatment by the court system or by their parents; axis I psychopathology other than cannabis, alcohol, nicotine or disruptive behavior disorders, as defined by DSM-IV-TR; active suicidality or psychotic symptoms; and medical conditions or medications that contraindicated taking study medication. Female participants were excluded if they were pregnant, nursing, or unwilling to use birth control. A medical history, physical exam, and laboratory tests were used to confirm medical eligibility.
Procedure
This study involves analysis of baseline, pre-randomization data from a larger clinical trial evaluating the effect of a medication to reduce cannabis use (Miranda et al. 2017). The Brown University Institutional Review Board approved all study procedures. Participants were recruited from the community through advertisements and flyers seeking research volunteers who wanted to reduce their cannabis use. Interested volunteers completed a brief telephone screening to determine provisional eligibility criteria. Youth who appeared eligible were invited to complete a comprehensive in-person interview and medical screening to confirm eligibility. Written informed consent was obtained for youths aged 18 to 24 years; minors (< 18 years) provided written assent and their parents provided written informed consent. Data for the present study were collected in the human laboratory and natural environment. An initial baseline laboratory visit, prior to trial randomization and medication administration, preceded an EMA period of approximately 1 week, which was also pre-randomization and pre-medication. Therefore, none of the participants were on medication to reduce cannabis use during the course of the laboratory and EMA procedures included in this report.
Startle response in the laboratory
Participants sat in a comfortable chair 1.5 m from a high-definition 27-in. flat screen color television. Electrodes were attached to participants and signal quality was checked. Participants viewed 60 color photographs, which included 15 cannabis-related stimuli, 15 non-cannabis-related pleasant, 15 unpleasant, and 15 neutral stimuli. Pleasant, unpleasant, and neutral stimuli were selected from the International Affective Picture System (IAPS; Lang et al. 1999). Pictures with pleasant and unpleasant ratings were matched on rated arousal. Images were presented using a microcomputer in 3 blocks of 20; each block included nearly equal numbers of cannabis, pleasant, neutral, and unpleasant themes. Ordering of images within each block was randomly determined and the order of block presentation was counterbalanced across participants.
To elicit startle reactivity, acoustic probes, which consisted of 95 dB (SPL A), 50 ms, white-noise bursts with instantaneous rise time, were delivered biaurally on 15 trials in each block via matched Telefonics TDH 49 headphones (Farmingdale, NY). Acoustic probes were delivered at randomly determined variable points following picture onset (i.e., 2, 4.5, or 5.5 s) and during only 45 out of the 60 pictures to decrease predictability of the acoustic probe. Eye blink electromyographic (EMG) activity (μV) was recorded and scored using a commercial system (Human Startle Version 2.20; Coulbourn Instruments, Allentown, PA) from a bipolar configuration of 3 mm Ag-AgCl surface electrodes filled with Microlyte Gel (Coulbourn, Allentown, PA) and placed in an inferolateral position over the orbicularis oculi of each participant’s left eye with a reference minielectrode placed over the right mastoid process. Raw signals were amplified X 10,000 using a Coulbourn V75-05 Bioamplifier with an 8–150 Hz bandpass, full-wave rectified, and integrated using a Coulbourn V76-23 contour-following integrator with a 10 msec time constant. Our startle procedures were similar to validated procedures and prior research examining the cue-modulated startle response (e.g., Cui et al. 2012; Lam et al. 2012; Leménager et al. 2014; Miranda et al. 2002, 2003; Rubio et al. 2011).
Natural environment
Participants completed an EMA period of approximately 1 week in their usual settings in their natural environments using handheld wireless devices (Omnia; Samsung Electronics, Ridgefield Park, NJ). Following the laboratory procedure described above, participants completed an EMA training session, in which they were oriented to the device and received detailed instructions for study protocols. Participants completed reports upon waking, before and after cannabis use, and in response to device-delivered audible prompts. To focus on cue-elicited craving, the present analyses utilized device-prompted responses occurring outside of cannabis-use episodes. Participants entered their responses into the devices in response to the device-delivered audible prompts at several times each day by tapping directly on the screen. Prompts were delivered at randomly selected times once within each 3-h block (e.g., 3 p.m. to 6 p. m.) and prompts were suspended when participants were sleeping or indicated an incompatible activity (e.g., driving). Response types included visual analogue sliding bars (e.g., degree of current craving), multiple checkboxes when more than one option is appropriate (e.g., who are you with), and categorical checkboxes when only one response was possible (e.g., location). Our EMA procedures were similar to prior research using EMA to examine substance use among adolescents and young adults (Miranda et al. 2014; Ramirez and Miranda 2014) and procedures using the first EMA week prior to medication randomization in other clinical trials (e.g., Ray et al. 2010).
Measures
Human laboratory
During their initial baseline visit, participants completed structured interviews and self-report measures.
Demographics
Participants reported their age, sex, and race/ethnicity.
Psychopathology
Participants were assessed for psychiatric diagnoses, including CUDs, with the Kiddie Schedule for Affective Disorders for School-Age Children, a semi-structured interview based on DSM-IV-TR criteria (Kaufman et al. 1997). Case consensus based on participant responses was used to determine diagnoses.
Cannabis use
Participants reported their cannabis use over the past 90 days using a timeline follow-back (TLFB) interview (Sobell and Sobell 1992). The TLFB is a reliable measure of substance use that correlates highly with plasma tetrahydrocannabinol levels (THC; Hjorthøj et al. 2012).
Picture ratings
At the end of the startle procedure, participants viewed each picture a second time and rated it for valence (0 = unhappy to 8 = happy) and arousal (0 = calm; 8 = excited) using the Self-Assessment Manikin (SAM; Lang et al. 1990). Participants also rated their cannabis craving for each photograph (0 = no urge to 10 = strongest ever).
Natural environment
Participants’ craving/urge to use cannabis as well as the presence of visible cannabis cues were assessed at device-delivered prompts during the one-week ecological momentary assessment period.
Cannabis craving/urge
Visual analog sliding bars assessed urge to smoke pot. The prompt stated “How strong is your urge to smoke pot right now?” Response options ranged from 0 = “no urge” to 10 = “strongest ever.”
Visible cannabis cues
Participants indicated the visible presence of cannabis use using the following prompt: “Is pot visible?” Response options were 0 = “no”; 1 = “yes, indirectly (e.g., High Times, photos)”; and 2 = “yes, directly”. They were coded to indicate whether visible cannabis cues were not present (no = 0) or present (indirectly/directly = 1).
Analytic plan
Startle data reduction procedures
Startle paradigm slide condition was a within-participant, repeated variable with four categories of pictorial cues: cannabis, pleasant, neutral, and unpleasant. A larger EMG startle response indicated a more aversive response to the pictorial cue, and a smaller EMG startle response indicated a more appetitive response to the pictorial cue. Consistent with prior work (Miranda et al. 2002, 2003), integrated startle EMG activity to each auditory probe was computer scored and reviewed. Eye-blink reflex magnitudes were calculated as the difference between the integrated EMG during the 20 msec before probe onset (baseline) and the maximum integrated EMG response between 21 and 120 msec after the acoustic probe. Trials were rejected if (a) the onset of the startle reflex did not occur in the 21- to 120-msec post-startle period, (b) EMG activity within the 20-msec baseline period was 12 μV or more (excessive noise), or (c) the change from baseline to the peak of activity during the 21 to 120 msec after probe onset was less than 2 μV (adapted from Grillon et al. 1998). Of the 86 participants enrolled in the study, four were excluded from analyses due to equipment malfunctioning during the startle reactivity task. To ensure that an adequate number of trials across all trials were included in each participant’s data, a criterion of at least 24.4% valid trials across blocks was used; as such, we excluded 22 participants. An additional 5 participants were excluded because of an insufficient number (< 3) of valid startle EMG responses to the cannabis slides. Given our sample is comprised of cannabis users who are youth and the sensitivity of eye-blink electrodes, we used stringent criteria for including participants to ensure a rigorous examination of the startle procedure and obtain robust findings. Prior studies often do not provide detailed data reduction criteria; however, of the ones we identified that explain their criteria, some use less stringent criteria to reduce their data (e.g., use outlier responses of >3 SDs; Cui et al. 2012; Gantiva et al. 2015; Lam et al. 2012). Nonetheless, our rates of excluded data (i.e., non-responders) are comparable to other work (e.g., Cui et al. 2012), and non-responders are common in clinical populations and among startle paradigms using auditory startle prompts (Blumenthal et al., 2005).
Because of large individual differences in this measure, EMG blink magnitudes are expressed in the standardized t-score metric (M, 50; SD, 10) by using the individual mean and SD from each participant across all four valence categories. The data were then sorted by slide valence category. Each participant’s response pattern was not altered by this standardization (t-score) procedure, in that the relative magnitude of each participant’s response to the four slide valence categories was maintained. Use of this transformation is consistent with previous investigations (e.g., Miranda et al. 2002, 2003). Analyses were conducted with both raw and standardized scores for affective modulation of startle.
Laboratory cue-elicited responses
Initial descriptive analyses of startle reactivity to pictorial cues in the human laboratory explored whether the expected linear trend in average startle magnitude was produced (i.e., t scores of pleasant < neutral < unpleasant) and whether the same pattern was produced for cannabis slides (i.e., t scores of cannabis < neutral < unpleasant). Average subjective ratings of valence and arousal across slide conditions were also compared. Multilevel growth models (MLGMs) provided a formal, statistical analysis of the linear trend in startle magnitude across slide conditions. The MLGM approach accounted for the within-participant correlation associated with repeated slide presentations and varying numbers of usable startle observations across participants (Gibbons et al. 2010; Raudenbush and Bryk 2002; Singer and Willett 2003). Paired-samples t tests also statistically compared startle magnitude in cannabis, pleasant, neutral slide conditions.
Natural environment cue-elicited responses
MLGM was again used to account for repeated observations of craving within subjects and variable numbers of reports of craving for each subject. Subjective ratings of craving in the natural environment (level 1) were nested within participants (level 2). Initial analyses tested whether the presence of visible cannabis cues in the natural environment (0 = cues not present; 1 = cues present) elicited craving in the natural environment. Next, several contextual covariates of cue-elicited craving were evaluated (i.e., time of day, weekend, presence of peers, location).
Laboratory and natural environment
Focal analyses tested whether startle magnitude to pictorial cues in the human laboratory predicted cue-elicited craving in the natural environment. A series of multilevel models (MLMs) tested whether EMG startle reactivity to pictorial cues in the human laboratory predicted craving in the natural environment. MLMs included the presence of visible cues, startle magnitude to each slide condition, and the cross-level interaction of cues × startle to test whether EMG startle reactivity in the human laboratory was associated with cue-elicited craving in the natural environment. Pseudo-R2 were calculated from the residual variance of nested models to identify the percentage of cue-elicited craving variability accounted for by EMG startle reactivity. Final models accounted for contextual covariates and potential person-level influences (i.e., age, gender, and number of CUD symptoms). MLM and MLGM analyses were implemented with SAS/STAT software, version 9.4 [2002–2012] (SAS Institute Inc. n.d), with slide condition (level 1) nested within participant (level 2). Models included random intercept terms and unstructured covariance matrices.
Results
The analyzed sample included a total of 55 participants ages 16 to 24 years (M = 19.87, SD = 1.94; 54.5% male; 47.3% White; 36.4% Black; 21.8% Hispanic). A large number of participants (56.4%) met criteria for current cannabis dependence, and 9 (16.4%) met criteria for current cannabis abuse; thus, 15 (27.3%) did not meet criteria for a current CUD diagnosis. The mean age of onset for cannabis dependence was 17.35 years (SD = 1.89, range = 14 to 22). Participants used cannabis on 70.81% of the past 90 days (SD = 27.97) with an average of 0.73 g of marijuana (SD = 0.61) per use day. Analyses comparing participants in the analytic sample to participants who were not included due to equipment malfunctioning or non-response indicated that they did not significantly differ in age, t(84) = −.95, p = .346, sex, χ2 (1) = 2.1, p = .148, or the number of CUD symptoms, t(84) = 1.25, p = .216.
Laboratory cue-elicited responses
As a preliminary step, participants’ ratings of valence, arousal, and craving in response to pictorial cues in the human laboratory were compared (see Table 1). Higher valence ratings were expected for cannabis and pleasant slides, relative to neutral and unpleasant. Higher arousal ratings were expected for unpleasant, pleasant, and cannabis slides, relative to neutral. Finally, higher craving ratings were expected for cannabis slides relative to pleasant, neutral, and unpleasant. The specified pattern of slide ratings was supported (see Fig. 1). With regard to valence, the expected inverse association of ratings for pleasant and unpleasant slides was shown, r = − .50, p < .001, and a similar inverse association of ratings for cannabis and unpleasant slides was shown, r = − .56, p < .001. Overall, ratings of valence, arousal, and craving were highest for cannabis slides, with subjective craving almost three points higher for cannabis slides than pleasant slides.

Comparisons of mean subjective ratings of valence, arousal, and craving in response to pictorial cues in the human laboratory. Error bars represent standard error of the mean. Valence ratings were higher for cannabis and pleasant slides, relative to neutral and unpleasant. Arousal ratings were higher for unpleasant, pleasant, and cannabis slides, relative to neutral. Finally, craving ratings were highest for cannabis slides relative to pleasant, neutral, and unpleasant
Paired-samples t-tests compared valence, arousal, and craving ratings across cue types. All pairwise differences were statistically significant, ps < .035. Valence. All pairwise differences for valence ratings were statistically significant, ps < .001. Arousal. The smallest magnitude, yet still statistically significant, difference of arousal was for cannabis and pleasant slides, t (54) = 2.55, p = .014. All other pairwise differences were also statistically significant, p < .001. Craving. The smallest magnitude, yet still statistically significant, difference of craving ratings was for neutral and unpleasant slides, t (54) = 2.16, p = .035. All other pairwise differences were also statistically significant, p < .001.
As a manipulation check of the startle laboratory paradigm, a linear slope evaluated the change in EMG reactivity across unpleasant (Time = 0), neutral (Time = 1), and pleasant (Time = 2) slide conditions using MLGMs. Similarly, a linear slope evaluated the change in EMG reactivity across unpleasant (Time = 0), neutral (Time = 1), and cannabis (Time = 2) slide conditions. In both cases, the slope effect was significant in a negative direction (pleasant: slope = − 2.02, p < .001, 95%CI [− 2.75, − 1.30]; cannabis: slope = − 1.56, p < .001, 95%CI [− 2.28, − 0.84]), providing statistical support for the expected linear trends in startle reactivity.
Next the overall magnitude of EMG reactivity was compared across slide types (Table 2). As expected, the magnitude of EMG startle response (presented as a t score) was highest for unpleasant pictorial cues, indicating a more aversive response, and lowest for cannabis and pleasant cues, indicating a more appetitive response (Table 2). There were statistically significant differences in startle magnitude for unpleasant cues compared to neutral cues, t(42) = −3.65, p = .001, pleasant cues, t(47) = −4.79, p < .001, and cannabis cues, t(50) = −4.33, p < .001. Pleasant cues were not statistically different from neutral cues, t(44) = −1.60, p = .116. Cannabis cues were also not statistically different from neutral cues, t(46) = −0.19, p = .853, or pleasant cues, t(50) = 1.41, p = .164.
Natural environment cue-elicited responses
Device timestamps were used to identify random prompts that occurred after use of cannabis or alcohol. All random prompts completed after alcohol or cannabis use on a given day were excluded to avoid confounding cue-elicited responses with acute subjective effects of drug use and satiation. Of 1164 reports completed, 307 (26.4%) followed drug use and were excluded. An additional 5 reports were missing data for visible cannabis cues; albeit minimal, cases of missing data occurred when the device malfunctioned and prematurely exited a report. Table 3 compares several contextual variables in the presence and absence of visible cannabis cues in the natural environment. Descriptive comparisons of counts and percentages within cue categories are presented in the left columns. A formal, logistic MLM test predicting the presence of visible cannabis cues simultaneously from several contextual variables is presented in the right columns. On the whole, cannabis cues were present for 207 (24.3%) random prompt reports. Visible cues were twice as likely to be present when youth were in the context of other peers, OR = 2.07, p = .002. In contrast, cues were less likely in school, public places, or in the car, relative to at youths’ own homes, ps from .002 to .046 (see Table 3). Next, an MLM predicting craving from cues showed that visible cannabis cues in the natural environment elicited craving, b = 1.22, p < .001, 95% CI [0.75, 1.68].
Laboratory and natural environment
Focal analyses tested whether subjective responses to cannabis cues, EMG startle reactivity to cannabis cues, or both predicted cue-elicited craving in the natural environment. Subjective craving while viewing cannabis pictures in the laboratory was associated with greater craving in the natural environment, in general, b = 0.56, p < .001, 95% CI [0.38, 0.74], but did not predict craving specifically in the presence of cues, p = .296. Conversely, greater EMG startle reactivity to cannabis slides in the laboratory (less appetitive response) was not significantly associated with craving in the natural environment in general, b = − 0.16, p = .159, 95% CI [− 0.39, 0.06], but was significantly associated with decreased craving when youth were in the presence of visible cannabis cues, b = − 0.15, p = .022, 95% CI [− 0.28, − 0.02]. In other words, an appetitive startle response to cannabis cues in the laboratory was associated with increased craving in the natural environment when cues were present. These results are nearly identical when sex, age, and CUD symptom count are accounted for in the models. The pattern of significant results is also the same when contextual covariates are included and when both subjective craving and EMG startle reactivity to cannabis cues are included simultaneously in a single model predicting cue-elicited craving (see Supplemental Table 1).
To verify specificity of this finding to cannabis-related stimuli, we also explored the effects of EMG reactivity to pleasant, neutral, and unpleasant pictorial stimuli in the laboratory on cue-elicited craving in the natural environment. In contrast to findings for cannabis slides, greater EMG startle reactivity to pleasant slides in the laboratory (less appetitive response) was significantly associated with increased craving when youth were in the presence of visible cannabis cues, b = 0.27, p < .001, 95% CI [0.12, 0.43]. Put differently, a blunted appetitive startle response to pleasant cues in the laboratory was associated with increased craving in the natural environment when cues were present. This effect remains significant when participant and contextual covariates are included (see Supplemental Table 2). Interactive effects were not significant for neutral or unpleasant stimuli, ps = .961 and .357, respectively.
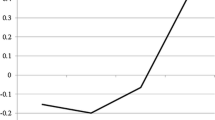
The cross-level interactive effects of EMG startle reactivity to cannabis and pleasant pictorial cues in the human laboratory predicting craving in response to visible cannabis cues in the natural environment are depicted graphically in Fig. 2 (Preacher et al. 2006). Lines represent the simple slopes at ±1 standard deviation of startle t scores for pictorial cues (gray and black lines), as well as the average startle magnitude (dashed line). For cannabis slides, the simple slope representing cue-elicited craving among youth with low startle reactivity was significant, slope = 1.80, SE = 0.34, p < .001 (gray line), whereas the simple slope among youth with high startle reactivity remained significant, but was attenuated, slope = 0.68, SE = 0.33, p = .037 (black line). For pleasant slides, the simple slope representing cue-elicited craving among youth with low startle reactivity was not significant, p = .210 (gray line), whereas the simple slope among youth with high startle reactivity was significant, slope = 2.16, SE = 0.38, p < .001 (black line). To put our findings into context, pseudo-R2 values suggested that 3.00% of the variance in cannabis craving was due to visible cannabis cues in the environment. Additionally, 7.20% of the variability in real-world cannabis-cue induced craving was accounted for by startle response pleasant cues, and 0.58% of the variability in real-world cannabis-cue induced craving was accounted for by cannabis cues.

EMG startle reactivity to cannabis pictorial cues (left panel) and pleasant pictorial cues (right panel) in the human laboratory predicting craving in response to visible cannabis cues in the natural environment. Lines represent the simple slopes at ± 1 standard deviation of startle t scores for pictorial cues (gray and black lines), as well as the average startle magnitude (dashed line)
Discussion
This study is the first to examine cue-modulated startle reactivity and its relationship with cannabis craving in the laboratory and natural environment. Cannabis-related stimuli evoked an appetitive startle response pattern in the laboratory, and a cannabis-modulated appetitive startle response in the laboratory was associated with increased cue-elicited craving in the natural environment. Pleasant stimuli also evoked an appetitive response pattern, but by contrast, an appetitive startle response to pleasant cues was associated with decreased cue-elicited craving in the natural environment. On the whole, these findings support cue-modulated startle reactivity as an index of the phenotypic expression of cue-elicited cannabis craving in the laboratory and natural environment.
Our findings extend prior research in two important ways. First, this study advances our understanding of the phenomenology of craving in the context of both cannabis misuse and adolescents and emerging adults. Drug craving plays a key role in all contemporary theories of addiction and cannabis users show consistent reactivity to cannabis-related cues across several neurobiological measures (Charboneau et al. 2013; Gray et al. 2011; Henry et al. 2014), underscoring the importance of the brain’s reward and motivation regions (Drummond 2001). Cue-modulated startle response is well examined in research across several substances such as alcohol, tobacco, and cocaine (Franken et al. 2004; Geier et al. 2000; Miranda et al. 2002; Rubio et al. 2011). However, this study is the first to provide support for the cue-modulated eye blink startle response procedure as a laboratory paradigm within the context of cannabis and specifically among youth. Initial analyses provided support for this paradigm, as demonstrated by higher self-report ratings of valence, arousal, and craving for cannabis slides compared to the other slides, including higher subjective craving for cannabis slides than pleasant slides. Demonstrating a linear trend in startle reactivity suggesting a greater appetitive response to cannabis cues relative to neutral and unpleasant cues also further supported these findings. Thus, the cue-modulated startle response paradigm has promise for future laboratory research of youth’s substance craving. In addition, this cue-elicited craving response may be especially important during adolescence, as youth have exaggerated reward-based reactivity as compared to adults, and these tendencies are associated with heightened risk for substance use (Doremus-Fitzwater and Spear 2016; Spear 2011).
The second way our findings extend prior research is by demonstrating how subjective and psychophysiological assessments captured in controlled laboratory settings predict clinically relevant real-world behavior. Unlike previous work that did not document associations between self-reported cue-elicited craving in the laboratory and natural environment craving (e.g., tobacco craving; Shiffman et al. 2015), we found a significant relationship between the appetitive startle response to cannabis cues in the human laboratory and cannabis craving in the natural environment. Specifically, we found that an appetitive startle response to cannabis in the laboratory was associated with increased craving when youth were in the presence of visible cannabis cues in their natural environment. Our results may differ from prior work because of differences in our method of assessing cues in the laboratory (psychophysiological versus subjective) and the natural environment (indirect and direct visible cues versus only direct cues), sample age (youth rather than middle aged adults), and substance of focus (cannabis versus tobacco).
Although human laboratory studies are the standard for understanding mechanisms that underlie addiction, pairing EMA methods with laboratory paradigms can provide important information not obtainable from laboratory paradigms alone. For example, laboratory paradigms typically cannot inform the temporal sequence of putative mechanisms on substance use in real time and naturalistic settings. Therefore, even when laboratory studies identify putative mechanisms (e.g., craving) that promote the development or maintenance of pathological drug use, such findings cannot directly address whether these factors predict key features of drug use in everyday life. By pairing a laboratory paradigm with EMA we showed direct associations between how participants react to cannabis and other affectively charged cues in the human laboratory and how they respond to cannabis cues in daily life. This finding supports the ecological validity of the cue-modulated startle response laboratory paradigm. Our results provide support for the clinical utility of the appetitive startle response to cannabis cues in the human laboratory as an intermediary biobehavioral endophenotype underlying cannabis craving in the real world.
Examining both drug-related and non-drug-related motivationally relevant stimuli in cue reactivity research is critical to comprehensively understand and discern the motivational influences of drug-related cues (Versace et al. 2017). In addition to examining cannabis-cue reactivity, our study demonstrated unique results for non-cannabis pleasant-cue reactivity and no significant effects for neutral or negative cues. A blunted response to non-drug related pleasant cues is a key feature of addiction (Drummond 2001). We found that a blunted appetitive response to pleasant cues in the laboratory was associated with increased cannabis-cue induced craving in the natural environment. These results may be explained by individual differences in cue-modulated startle responses to reward-related cues (i.e., cannabis or pleasant cues), such as motivational or reward sensitivities in the brain, that are often overlooked in the literature (Versace et al. 2017). Future psychophysiological research is needed to examine individual differences in modulated brain responses to better inform our findings.
It is important to highlight the study’s findings within the context of its limitations. The generalizability of our results is limited to the youth in our sample who were between the ages of 16 and 24 years and interested in receiving a psychosocial intervention combined with a novel medication (or placebo) that may help them reduce their cannabis use, many of whom met criteria for cannabis dependence. Future research is needed to examine the generalizability of our findings to youth who are younger and use cannabis but have not yet developed dependence. Replication in a sample with greater variability of cannabis use, especially among adolescents who use cannabis but do not develop CUD, using longitudinal designs across adolescence is necessary to explore the relation of startle-reflex patterns to dependence progression. This is important as it may inform our understanding of the trajectories involved in developing an appetitive startle response to cannabis. Additionally, because we used data from a clinical trial, our EMA period was limited to the one-week prior to randomization and medication administration. This may have limited the range of cannabis craving usually experienced in varying contexts in the natural environment. Future research would benefit from examining the startle reflex effects in the laboratory on cannabis craving over a more extended period of time in the natural environment to better estimate the robustness of our findings. Our eligibility criteria for cannabis use were based on participants’ self-report; future research should include biomarkers to confirm participants’ eligibility and cannabis use. Although we found temporally sequenced associations between cannabis cue exposure and craving in the natural environment, the correlational nature of EMA data still limits interpretation somewhat. For example, it is possible that participants were more likely to experience cannabis cues on days when they planned to use, which might call to question the cue reactivity effect. This limitation is mitigated, however, by the fact that our findings were upheld when we re-tested all models while controlling for whether days were cannabis use or non-use days. Even so, additional research is needed to directly examine the effects of cannabis use intentions on cue reactivity captured in real-world settings.
Moreover, in order to rigorously test our effects, we set stringent criteria and eliminated many participants due to an insufficient number of valid startle responses (i.e., non-responders); however, we still found robust results. Although non-responders are more common in clinical populations and among startle paradigms using auditory startle prompts (Blumenthal et al. 2005), future researchers using this method should describe in detail their data reduction procedures to better discern an appropriate range of non-responders among youth. Furthermore, by nature, the startle response is a defensive or aversive response. Future research should consider directly testing psychophysiological measures of appetitive motivation (e.g., postauricular reflex; Quevedo et al. 2009). Lastly, our study only focused on cannabis craving; additional research is needed to understand the generalizability of these effects for craving substances other than cannabis.
Despite these limitations, this study has important implications for future research. Future studies may use the startle response to identify youth at greatest risk for cue-elicited cannabis craving in the natural environment. Studies show that cue-modulated startle response predicts substance use treatment success (Loeber et al. 2007), and thus leveraging this paradigm to understand how and why certain individuals respond better to certain treatments would advance treatment options. In addition, individuals’ motivation to change their substance use is related to their startle reflex patterns (Garfield et al. 2015). Thus, more research is needed to better understand the clinical implications of the startle response as a mechanism related to cannabis craving to better aid treatment interventions for youth.
In summary, this study’s findings from the laboratory and natural environment provide support for cue-modulated eye blink startle response as an index of the phenotypic expression of cue-elicited cannabis craving among youth. Emotion modulation of startle reactivity is universal across species. Thus, it is a unique translational tool for bridging preclinical and clinical research and for understanding the neurobiology of emotional and attention processes that underlie addiction.
Change history
07 July 2018
The name of Hayley Treloar Padovano was incorrectly tagged in the original version of this article. Instead of Padovano as family name and Hayley Treloar as given name, it should be Hayley as given name and Treloar Padovano as family name. Correct presentation of author name when online should be Treloar Padovano H.
References
Blumenthal TD, Cuthbert BN, Filion DL, Hackley S Lipp OV, Van Boxtel A (2005) Committee report: Guidelines for human startle eyeblink electromyographic studies. Psychophysiology 42(1):1–15. https://doi.org/10.1111/j.1469-8986.2005.00271.x
Bradley, M. M., & Lang, P. J. (2000). Measuring emotion: behavior, feeling, and physiology. In R. D. Lane & L. Nadel (Eds.), Cognitive neuroscience of emotion (pp. 242–276). New York, NY
Carter B, Tiffany S (1999) Meta-analysis of cue-reactivity in addiction research. Addiction 94(3):327–340
Charboneau EJ, Dietrich MS, Park S, Cao A, Watkins TJ, Blackford JU et al (2013) Cannabis cue-induced brain activation correlates with drug craving in limbic and visual salience regions: preliminary results. Psychiatry Res Neuroimaging 214(2):122–131. https://doi.org/10.1016/j.pscychresns.2013.06.005
Cui Y, Robinson JD, Versace F, Lam CY, Minnix JA, Karam-Hage M, Cinciripini PM (2012) Differential cigarette-related startle cue reactivity among light, moderate, and heavy smokers. Addict Behav 37(8):885–889. https://doi.org/10.1016/j.addbeh.2012.02.003
Doremus-Fitzwater TL, Spear LP (2016) Reward-centricity and attenuated aversions: an adolescent phenotype emerging from studies in laboratory animals. Neurosci Biobehav Rev 70:121–134. https://doi.org/10.1016/j.neubiorev.2016.08.015
Drummond DC (2001) Theories of drug craving, ancient and modern. Addiction 96:33–46. https://doi.org/10.1080/09652140020016941
Franken IHA, Hulstijn KP, Stam CSJ, Hendriks VM, Brink WVD (2004) Two new neurophysiological indices of cocaine craving: evoked brain potentials and cue modulated startle reflex. J Psychopharmacol 18(4):544–552. https://doi.org/10.1177/026988110401800412
Gantiva C, Guerra P, Vila J (2015) From appetitive to aversive: motivational interviewing reverses the modulation of the startle reflex by tobacco cues in smokers not ready to quit. Behav Res Ther 66:43–48. https://doi.org/10.1016/j.brat.2015.01.006
Garfield JB, Allen NB, Cheetham A, Simmons JG, Lubman DI (2015) Attention to pleasant stimuli in early adolescence predicts alcohol-related problems in mid-adolescence. Biol Psychol 108:43–50
Geier A, Mucha RF, Pauli P (2000) Appetitive nature of drug cues confirmed with physiological measures in a model using pictures of smoking. Psychopharmacology 150:283–291. https://doi.org/10.1007/s002130000404
Gibbons RD, Hedeker D, DuToit S (2010) Advances in analysis of longitudinal data. Annu Rev Clin Psychol 6:79–107. https://doi.org/10.1146/annurev.clinpsy.032408.153550
Goldman M, Szucs-Reed RP, Jagannathan K, Ehrman RN, Wang Z, Li Y et al (2013) Reward-related brain response and craving correlates of marijuana cue exposure. J Addict Med 7(1):8–16. https://doi.org/10.1097/ADM.0b013e318273863a
Gray KM, LaRowe SD, Upadhyaya HP (2008) Cue reactivity in young marijuana smokers: a preliminary investigation. Psychol Addict Behav 22(4):582–586. https://doi.org/10.1037/a0012985
Gray KM, LaRowe SD, Watson NL, Carpenter MJ (2011) Reactivity to in vivo marijuana cues among cannabis-dependent adolescents. Addict Behav 36(1–2):140–143. https://doi.org/10.1016/j.addbeh.2010.08.021
Grillon C, Dierker L, Merikangas KR (1998) Fear-potentiated startle in adolescent offspring of parents with anxiety disorders. Biol Psychiatry 44(10):990–997. https://doi.org/10.1016/S0006-3223(98)00188-7
Henry EA, Kaye JT, Bryan AD, Hutchison KE, Ito TA (2014) Cannabis cue reactivity and craving among never, infrequent and heavy cannabis users. Neuropsychopharmacology 39(5):1214–1221. https://doi.org/10.1038/npp.2013.324
Hjorthøj CR, Fohlmann A, Larsen A-M, Arendt M, Nordentoft M (2012) Correlations and agreement between delta-9-tetrahydrocannabinol (THC) in blood plasma and timeline follow-back (TLFB)-assisted self-reported use of cannabis of patients with cannabis use disorder and psychotic illness attending the CapOpus randomized clinical trial. Addiction 107(6):1123–1131. https://doi.org/10.1111/j.1360-0443.2011.03757.x
Johnston, L.D., O’Malley, P.M., Bachman, J.G., Schulenberg, J.E., 2013. Monitoring the future national results on drug use: 2012 Overview, Key Findings on Adolescent Drug Use. Institute for Social Research. The University of Michigan, Ann Arbor
Kaufman J, Birmaher B, Brent D, Rao U, Flynn C, Moreci P et al (1997) Schedule for affective disorders and schizophrenia for school-age children-present and lifetime version (K-SADS-PL): initial reliability and validity data. J Am Acad Child Adolesc Psychiatry 36(7):980–988. https://doi.org/10.1097/00004583-199707000-00021
Keyes KM, Wall M, Cerdá M, Schulenberg J, O'Malley PM, Galea S et al (2016) How does state marijuana policy affect US youth? Medical marijuana laws, marijuana use and perceived harmfulness: 1991–2014. Addiction 111(12):2187–2195
Lam CY, Robinson JD, Versace F, Minnix JA, Cui Y, Carter BL et al (2012) Affective reactivity during smoking cessation of never-quitters as compared with that of abstainers, relapsers, and continuing smokers. Exp Clin Psychopharmacol 20(2):139–150
Lang PJ, Bradley MM, Cuthbert BN (1990) Emotion, attention, and startle reflex. Psychol Rev 97:377–395
Lang, P. J., Bradley, M. M., & Cuthbert, B. N. (1999). International affective picture system (IAPS): instruction manual and affective ratings. The center for research in psychophysiology, University of Florida
Leménager T, Hill H, Reinhard I, Hoffmann S, Zimmermann US, Hermann D et al (2014) Association between alcohol-cue modulated startle reactions and drinking behaviour in alcohol dependent patients—results of the PREDICT study. Int J Psychophysiol 94(3):263–271. https://doi.org/10.1016/j.ijpsycho.2014.09.009
Loeber S, Croissant B, Nakovics H, Zimmer A, Georgi A, Klein S, Diener C, Heinz A, Mann K, Flor H (2007) The startle reflex in alcohol-dependent patients: changes after cognitive-behavioral therapy and predictive validity for drinking behavior—a pilot study. Psychother Psychosom 76(6):385–390. https://doi.org/10.1159/000107567
Meehan JC, Miranda R Jr (2013) Startle reflex and psychophysiology. Wiley-Blackwell Handb Addict Psychopharmacol:367–598
Miranda R, Meyerson LA, Buchanan TW, Lovallo WR (2002) Altered emotion-modulated startle in young adults with a family history of alcoholism. Alcohol Clin Exp Res 26(4):441–448
Miranda R, Meyerson LA, Myers RR, Lovallo WR (2003) Altered affective modulation of the startle reflex in alcoholics with antisocial personality disorder. Alcohol Clin Exp Res 27(12):1901–1911
Miranda R, Ray L, Blanchard A, Reynolds EK, Monti PM, Chun T, Justus A, Swift RM, Tidey J, Gwaltney CJ, Ramirez J (2014) Effects of naltrexone on adolescent alcohol cue reactivity and sensitivity: an initial randomized trial. Addict Biol 19(5):941–954. https://doi.org/10.1111/adb.12050
Miranda R, Treloar H, Blanchard A, Justus A, Monti PM, Chun T et al (2017) Topiramate and motivational enhancement therapy for cannabis use among youth: a randomized placebo-controlled pilot study. Addict Biol 22(3):779–790. https://doi.org/10.1111/adb.12350
Preacher KJ, Curran PJ, Bauer DJ (2006) Computational tools for probing interactions in multiple linear regression, multilevel modeling, and latent curve analysis. J Educ Behav Stat 31(4):437–448. https://doi.org/10.3102/10769986031004437
Quevedo K, Benning SD, Gunnar MR, Dahl RE (2009) The onset of puberty: effects on the psychophysiology of defensive and appetitive motivation. Dev Psychopathol 21(1):27–45. https://doi.org/10.1017/S0954579409000030
Ramirez J, Miranda R (2014) Alcohol craving in adolescents: bridging the laboratory and natural environment. Psychopharmacology 231:1841–1851. https://doi.org/10.1007/s00213-013-3372-6
Raudenbush SW, Bryk AS (2002) Hierarchical linear models: applications and data analysis methods, 2nd edn. Sage Publications, Inc., Thousand Oaks, CA
Ray LA, Miranda R, Tidey JW, McGeary JE, MacKillop J, Gwaltney CJ et al (2010) Polymorphisms of the μ-opioid receptor and dopamine D(4) receptor genes and subjective responses to alcohol in the natural environment. J Abnorm Psychol 119(1):115–125. https://doi.org/10.1037/a0017550
Rubio G, Borrell J, Jiménez M, Jurado R, Grüsser SM, Heinz A (2011) Variables involved in the cue modulation of the startle reflex in alcohol-dependent patients. Addict Biol 18(1):170–180
SAS Institute Inc. (n.d.). SAS/STAT software. Cary, NC, USA.
Sayette MA (2016) The role of craving in substance use disorders: theoretical and methodological issues. Ann Rev Clin Psychol 12(November 2015):407–433. https://doi.org/10.1146/annurev-clinpsy-021815-093351
Shiffman S, Stone AA, Hufford MR (2008) Ecological momentary assessment. Ann Rev Clin Psychol 4(1):1–32. https://doi.org/10.1146/annurev.clinpsy.3.022806.091415
Shiffman S, Li X, Dunbar MS, Tindle HA, Scholl SM, Ferguson SG (2015) Does laboratory cue reactivity correlate with real-world craving and smoking responses to cues? Drug Alcohol Depend 155:163–169. https://doi.org/10.1016/j.drugalcdep.2015.07.673
Singer JD, Willett JB (2003) Applied longitudinal data analysis. Oxford University Press, Inc., New York, NY
Sobell LC, Sobell MB (1992) Timeline follow-back: a technique for assessing self-reported alcohol consumption. In: Litten R, Allen K (eds) Measuring alcohol consumption. Human Press, Clifton, NJ, pp 41–65
Spear LP (2011) Rewards, aversions and affect in adolescence: emerging convergences across laboratory animal and human data. Dev Cogn Neurosci 1(4):390–403. https://doi.org/10.1016/j.dcn.2011.08.001
Substance Abuse and Mental Health Services Administration. (2015). Behavioral health trends in the United States: results from the 2014 National Survey on Drug Use and Health. (HHS Pulication No. SMA 15–4927, NSDUH Series H-50.
Tiffany ST, Wray JM (2012) The clinical significance of drug craving. Ann N Y Acad Sci 1248:1–17. https://doi.org/10.1111/j.1749-6632.2011.06298.x
Versace F, Engelmann JM, Deweese MM, Robinson JD, Green CE, Lam CY et al (2017) Beyond cue reactivity: non-drug-related motivationally relevant stimuli are necessary to understand reactivity to drug-related cues. Nicotine Tob Res 19(6):663–669. https://doi.org/10.1093/ntr/ntx002
Volkow ND, Baler RD, Compton WM, Weiss SR (2014) Adverse health effects of marijuana use. N Engl J Med 370(23):2219–2227
Volkow ND, Koob GF, McLellan AT (2016) Neurobiologic Advances from the Brain Disease Model of Addiction. N Engl J Med 374(4):363–371. https://doi.org/10.1056/NEJMra1511480
Acknowledgements
The National Institute on Drug Abuse (R01 DA026778) and the National Institute on Alcohol Abuse and Alcoholism (R01 AA007850, K08 AA025011, K23 AA024808) at the National Institutes of Health supported this work. The authors wish to thank Alexander Blanchard for his contribution to the data collection and database management supporting this work.
Author information
Authors and Affiliations
Corresponding author
Additional information
Ethan H. Mereish and Hayley Treloar Padovano were equal first-author contributors.
Rights and permissions
About this article

Cite this article
Mereish, E.H., Padovano, H.T., Wemm, S. et al. Appetitive startle modulation in the human laboratory predicts Cannabis craving in the natural environment. Psychopharmacology 235, 1933–1943 (2018). https://doi.org/10.1007/s00213-018-4890-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-018-4890-z