
Abstract
Rationale
Clinical studies have shown that some psychoactive recreational drugs have therapeutic applications in anxiety, depression, and schizophrenia. However, to date, there are few studies on the therapeutic potential efficacy of recreational drugs in compulsive neuropsychiatric disorders.
Objectives
We explored the therapeutic potential of different psychoactive and psychedelic drugs in a preclinical model of compulsive behavior.
Methods
Outbred male Wistar rats were selected as either high (HD) or low (LD) drinkers according to their behavior in schedule-induced polydipsia (SIP). Subsequently, we assessed the effects of acute administration of scopolamine (0.125, 0.25, and 0.5 mg/kg), methamphetamine (0.25, 0.5, 1.25, and 2.5 mg/kg), ketamine (1.25, 2.5, 5, and 10 mg/kg), cannabidiol (1 and 3 mg/kg), WIN21255–2 (0.5, 075, and 1 mg/kg), and AM404 (0.25 and 0.5 mg/kg) on compulsive drinking in SIP.
Results
Scopolamine reduced dose-dependent compulsive drinking in HD compared with LD rats in SIP. Methamphetamine induced a dose-dependent inverted U-curve effect in both groups, in which lower doses increased and higher doses reduced compulsive drinking in SIP. Ketamine, cannabidiol, WIN21255-2, and AM404 did not have any relevant effects in SIP.
Conclusions
These data provide new evidence that low doses of scopolamine and intermediate doses of methamphetamine might therapeutically reduce compulsive behaviors and suggest that there is not a direct participation of the endocannabinoid system in compulsive behavior on SIP. The research in the underlying neurochemical mechanisms of these psychoactive drugs might provide an additional insight on new therapeutic targets in compulsive neuropsychiatric disorders.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Compulsivity is associated with the loss of personal control over a broad range of behaviors that are prone to excess (Hollander et al. 2016). This condition is characterized by the performance of repetitive, undesirable, and dysfunctional acts without adaptive function, such as eating, gambling, and checking, according to rigid rules or to avoid negative consequences (Chamberlain et al. 2006; Fineberg et al. 2014). Compulsive behavior is typically observed in obsessive-compulsive disorder (OCD) and autism (Gillan et al. 2017; Kim et al. 2017) but has also been observed in many other neuropsychiatric disorders, such as schizophrenia (de Haan et al. 2009; Swets et al. 2014), bipolar disorder (Amerio et al. 2014), anxiety (Agarwal et al. 2016), specific phobias (Torres et al. 2014, 2016), depression (Agarwal et al. 2016; Baer et al. 2015; Rickelt et al. 2016), and addiction (Fineberg et al. 2014). Compulsivity has different facets; indeed, some studies have supported a prognostic role for compulsivity in treatment outcome (Blanco et al. 2009; Grant et al. 2010; Ruehle et al. 2012). Reflecting the heterogeneity in compulsive symptoms, the treatment results are highly variable (Gillan et al. 2017). Pharmacological treatments for compulsivity are focused on antipsychotics and selective serotonin reuptake inhibitors (SSRI); however, up to 40% of patients do not respond successfully (Marinova et al. 2017). These facts motivate the exploration of new drugs that may have a therapeutic potential to reduce compulsive behavior.
In recent years, psychoactive drugs commonly used for recreational purposes, such as ketamine, scopolamine, methamphetamine, or cannabinoids, have aroused some interest because of their potential therapeutic applications. Previous studies have investigated the role of glutamatergic drugs in OCD (Marinova et al. 2017), in which ketamine remains an experimental treatment. Ketamine is a N-methyl-d-aspartate (NMDA) receptor antagonist that modulates the glutamatergic system and exerts some effects on AMPA receptors, acetylcholine receptors, GABA receptors, μ-opioid receptors, and ĸ-opioid receptors, as well as inhibits the synaptic reuptake of noradrenaline and serotonin (Bruhn et al. 2017). In clinical studies, ketamine is a drug of interest because of its potential for a rapid onset of action, and administration of 0.5 mg/kg ketamine has shown antidepressant efficacy (Ionescu et al. 2014; Phelps and Brutsche 2009; Sos et al. 2013; Zarate et al. 2006), decreased suicidal ideations (Diazgranados et al. 2010) and anhedonia (Lally et al. 2014), and reduced symptomatology in posttraumatic stress disorder (Feder et al. 2014). Scopolamine is a non-selective muscarinic acetylcholine receptor (MAChR) antagonist that blocks the cholinergic function of the central nervous system by targeting muscarinic M1AchR and M2AchR receptors (Ionita et al. 2017). A single dose of scopolamine rapidly increases the spine number and function in layer V neurons, mTOR signaling, and glutamate release in the prefrontal cortex (Zarate et al. 2013). Scopolamine administration (4 μg/kg) exhibited rapid antidepressant efficacy in patients with treatment-resistant depression (Furey et al. 2013) and induced anxiolytic effects in women (Furey et al. 2010). Methamphetamine (METH) is a psychostimulant drug, similar to d-amphetamine (Drug Enforcement Administration 2013; Wu et al. 2007), and exhibits potent full agonism of trace amine-associated receptor 1 (TAAR1), which increases cyclic adenosine monophosphate (cAMP) production and either completely inhibits or reverses the function of vesicular monoamine transporters for dopamine, norepinephrine, and serotonin (Cruickshank and Dyer 2009). METH also decreases the metabolism of monoamines by inhibiting monoamine oxidase, resulting in prolonged neuronal signaling (Cadet and Krasnova 2007; Krasnova and Cadet 2009). Although METH has become a major drug of abuse worldwide (Rau et al. 2016), it has been used for the treatment of weight control and depression as well as to increase alertness and prevent sleep (Drug Enforcement Administration, 2013). Recent studies have shown the therapeutic role of low doses of acute METH administration for neuroprotection in cognitive and behavioral impairment after severe traumatic brain injury (Rau et al. 2012, 2016).
Cannabinoid drugs exhibit effects via the cannabinoid receptors CB1 and CB2 (Russell 2017). The CB1 receptor is expressed in different brain areas, including those associated with impulsive-compulsive behaviors, such as the prefrontal cortex, striatum, and limbic system (Herkenham et al. 1991; Micale et al. 2009). In preclinical and clinical studies, cannabidiol (CBD) has demonstrated a broad range of potential therapeutic properties, such as antipsychotic, antidepressant, anxiolytic, antiepileptic, sedative, and neuroprotective effects (Bergamaschi et al. 2011; Campos et al. 2016). Human studies on the exogenous cannabinoid AM404, an inhibitor of the fatty-acid amide hydrolase enzyme for endocannabinoid reuptake (Patel and Hillard, 2006), have reported anxiolytic properties and the attenuation of schizophrenia symptoms (Crippa et al. 2010; Leweke et al. 2012; Schubart et al. 2011; Zuardi et al.1995). The full cannabinoid CB1 receptor agonist WIN55212-2 (Komaki et al. 2015) demonstrated neuroprotective effects on Parkinson disease (for a review, see More and Choi, 2011).
However, there are few preclinical and clinical studies on the therapeutic role of these psychoactive and psychedelic drugs on reducing compulsive behaviors in neuropsychiatric disorders. Schedule-induced polydipsia (SIP) model is characterized by the development of an adjunctive behavior of excessive drinking in food-deprived animals exposed to intermittent food-reinforcement schedules (Falk, 1961, 1966). The drinking response in SIP, considered as adaptive in nature, could serve as a coping or stress-reducing mechanism as indicated by reductions in plasma corticosterone levels (Brett and Levine, 1979; Dantzer et al. 1988a). Moreover, different studies have demonstrated relevant individual differences in SIP acquisition (Dantzer et al. 1988a,b; Mittleman et al. 1988). The excessive and persistent drinking response could indicate the development of a maladaptive compulsive behavior in SIP (Platt et al. 2008; Moreno and Flores, 2012). A similar phenomenon, known as psychogenic polydipsia, which includes compulsive non-regulatory fluid consumption, is present in 6–20% of psychiatric patients (de Leon et al. 1994, 2002; Dundas et al. 2007; Evenson et al. 1987; Iftene et al. 2013), and its treatment has shown a limited effectiveness (Greendyke et al. 1998; Delva et al. 2002). Different dopamine and serotonin agents such as antipsychotics and SSRIs efficiently reduce compulsive drinking on SIP (Íbias et al. 2016; for a review, see: Moreno and Flores, 2012; Platt et al. 2008). Moreover, the selection of high drinker (HD) and low drinker (LD) animals by SIP acquisition has shown that there are relevant behavioral and neurochemical differences. HD rats show lack of inhibitory control in 5-choice serial reaction time task (Moreno et al. 2012), latent inhibition deficit and cognitive inflexibility in reversal learning task (Navarro et al. 2016), and increased compulsive behavior in SIP after consuming tryptophan-depleted diet (Merchan et al. 2017) compared to LD rats. The administration of psychostimulant drugs, such as d-amphetamine and cocaine, as well as SSRIs, can efficiently reduce dose-dependent compulsive drinking behavior in HD rats in SIP (López-Grancha et al. 2008; Navarro et al. 2015). Indeed, administration of the serotonin 5-HT2A/C receptor agonist DOI, a psychedelic drug, demonstrated a dose-dependent reduction of compulsive drinking in SIP for HD rats.
In the present study, we investigated the potential therapeutic role of psychoactive and psychedelic drugs that have historically been used for recreational purposes in reducing compulsive drinking in a compulsive phenotype, the HD rats selected by SIP procedure. We explored the dose-response effects on compulsive drinking in two different groups of HD and LD rats. In the first group of HD and LD rats, we explored the effects of scopolamine, METH, and ketamine, and in the second group of HD and LD rats, we assessed the effects of acute administration of cannabinoid drugs, including cannabidiol, WIN55212-2, and AM404, on reducing compulsive drinking in SIP. The results are discussed in terms of the potential therapeutic effects of psychoactive and psychedelic drugs in a compulsive phenotype rat population and their implication as new pharmacological therapies for compulsive neuropsychiatric disorders.
Materials and methods
Subject
A total of 40 male Wistar rats (Janvier Labs, France) weighing approximately 250–350 g at the start of the experiments were used in the present study. The animals were housed four rats per cage (50 × 15 × 25 cm) at 22 °C, with a 12:12-h light-dark cycle (lights off at 08:00 h) and food and water provided ad libitum. Prior to SIP training and after 10 days of habituation, rats were gradually reduced to 85% of their free-feeding body weight through controlled feeding, and their body weights were maintained at this level of deprivation throughout the experiments. Food was provided daily at approximately 30 min after each experimental session. All testing was performed between 9:00 and 15:00 h. All procedures were performed according to the Spanish Royal Decree 1201/2005 on the protection of experimental animals and the European Community Council Directives (86/609/EEC).
SIP procedure
Rats were tested in eight operant SIP chambers (35 × 25 × 34 cm) (MED Associates, St. Albans, VT, USA). A detailed description of the apparatus has previously been provided (López-Grancha et al. 2008; Moreno et al. 2010). The scheduling and recording of experimental events were controlled using a computer and the commercial software Med PC (Cibertec SA, Spain). Over two successive days, a water ingestion test was conducted (baseline). Sixty pellets (Noyes 45-mg dustless reward pellets; TSE Systems, Germany) were placed together, and the amount of water consumed by each rat in 60 min was measured. After 1 day of adaptation, animals were exposed to a fixed time 60-s (FT-60s) schedule of food pellet presentation in 60-min sessions. Water bottles containing 100 ml of fresh water were provided during each session. After 20 daily sessions, animals were separated into two specific populations, LD and HD, according to drinking rates (average for each animal over the last five sessions) above or below the group median, respectively. For the experimental drug studies, LD and HD rats were further divided into two subsequent groups according to their drinking rate: a first experimental group of HD (n = 10) and LD (n = 10) and second experimental group of HD (n = 10) and LD (n = 10). The following measures were recorded for each rat: (a) total amount of water (milliliters) removed from the bottle, (b) total number of licks on the bottle, and (c) total number of entries into the food magazine.
Experimental design
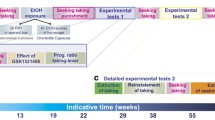
The behavioral effects of acute systemic administration of different drugs were tested in two separated groups of LD and HD rats in SIP. All animals received drugs according to a fully randomized Latin-square design, separated by a minimum of 72 h between drug test sessions and 15 days between different drug experiments (animals continued performing SIP sessions during these days). The experimental sessions were conducted on Tuesdays and Fridays, and baseline testing was performed on Mondays and Thursdays. On Wednesdays, animals performed the task, but the results were not analyzed. The experimental events are summarized in Fig. 1.

Experimental procedure illustrated in a timetable. Drug doses were administered according to a fully randomized Latin-square design
Experiment 1
We examined the effects of scopolamine, METH, and ketamine in LD and HD rats in SIP. The effects of scopolamine (0.125, 0.25, and 0.5 mg/kg), METH (0.25, 0.5, 1.25, and 2.5 mg/kg), and ketamine (1.25, 2.5, 5, and 10 mg /kg) were investigated in Group 1. The drug doses, injection time of approximately 30 min prior to behavioral testing and intraperitoneal (i.p.) administration, were implemented based on previous experiments (de la Peña et al. 2012; Petryshen et al. 2016; Refsgaard et al. 2017; Tizabi et al. 2012; Yamazaki et al. 2015).
Experiment 2
We explored the effects of CBD, WIN55212-2, and AM404 in HD and LD rats in SIP. We assessed the effects of the following drugs on SIP in group 2: CBD (1 and 3 mg/kg), WIN55212-2 (0.5, 075 and 1 mg/kg), and AM404 (0.25 and 0.5 mg/kg). The drug doses, injection time of approximately 30 min prior to behavioral testing, and intraperitoneal (i.p.) administration were selected based on previous experiments (Adamczyk et al. 2008; Campolongo et al. 2012; Espejo-Porras et al. 2013; Komaki et al. 2015; Zanelati et al. 2010).
Drugs
Scopolamine (scopolamine hydrobromide, (S)-3Hydroxy-2-fenylproprionic acid (1R,2R4S7S,9S)-9-methyl-3-oxa-9-azatricyclo[3.3.1.02,4]non-7-yl ester), METH ((+)-methamphetamine hydrochloride), and ketamine were dissolved in 0.9% saline. CBD ((−)-Cannabidiol), WIN55212-2 ((R)-(+)-WIN55212-2 mesylate salt), and AM404 (N-(4-hydroxyphenyl)-arachidonylamide) were suspended in 2% Tween-80 in 0.9% saline. All drugs were purchased from Sigma-Aldrich (Madrid, Spain), except CBD, which was purchased from Tocris Bio-Techne (Madrid, Spain). The injection volumes were 1 ml/kg for all drugs.
Data analysis
Behavioral data on SIP acquisition were analyzed using two-way repeated-measure analysis of variance (ANOVA), with “group” (LD and HD) as the between-subject factor and “sessions” (20 sessions) as the within-subject factor. The effects of the different drugs in LD and HD on SIP were analyzed using two-way repeated-measure ANOVA, with group (LD and HD) as the between-subject factor and “drug” (different doses of drug and vehicle) as the repeated within-subject factor. Post hoc comparisons were performed using the Newman-Keuls test. Statistical significance was set at p < 0.05. All analyses were computed using Statistica software (version 6.0).
Results
LD and HD selected by SIP
The mean water intake in LD and HD during acquisition and maintenance of SIP is shown in Fig. 2. In the experimental phase, the mean water intake over the last 5 days of SIP was 5.9 ± 0.1 and 16.6 ± 1.1 ml for LD and HD, respectively. The number of licks also showed SIP acquisition. The mean total licks averaged across the last 5 days of SIP were 763.4 ± 118.9 and 2296.6 ± 254.2 for LD and HD, respectively (data not shown).

The mean (± SEM) water intake in FT-60s across 20 sessions of SIP. Statistical analyses indicate significant differences between high drinkers (n = 20) and low drinkers (n = 20) from that session onwards (asterisk). Significant differences from session 1 (number sign)
ANOVA revealed significant differences in water intake according to the interaction between SIP acquisition sessions and LD vs. HD (interaction SIP session × group effect: F19, 722 = 14.56, p < 0.001). This difference was also confirmed by the significant interaction observed in the total number of licks (interaction SIP session × group effect: F19, 722 = 6.79, p < 0.001). Post hoc analysis indicated that the FT-60s schedule of food delivery induced different drinking rates across the 20 test sessions in both groups. Differences between the LD and HD animals were evident in the water intake at session 3 (p < 0.05) onwards. Furthermore, animals in the HD group significantly increased their consumption of water from session 4 (p < 0.01) compared to session 1. Differences between the LD and HD groups in the number of total licks at session 4 (p < 0.01) were also observed, and HD rats increased their number of licks from session 4 (p < 0.05) compared to session 1. There were no significant differences between LD and HD animals in the total magazine entries in SIP (data not shown).
Experiment 1. Effects of scopolamine, methamphetamine, and ketamine on SIP
The effects of scopolamine on water intake, total licks, and total magazine entries in SIP are shown in Fig. 3a and Table 1. Scopolamine significantly reduced compulsive water intake in HD rats compared to LD rats (group × drug interaction, F3, 54 = 6.24, p < 0.01; group, F1, 18 = 12.71, p < 0.01; drug, F3, 54 = 11.67, p < 0.001). Post hoc analyses revealed that scopolamine reduced dose-dependent water intake in HD rats at the following doses: 0.125 (p < 0.05), 0.25 (p < 0.01), and 0.5 mg/kg (p < 0.001) compared with vehicle in the same group. Scopolamine did not affect water intake in LD rats. The comparison between LD and HD revealed a dose-dependent reduction of the significant differences in water intake (vehicle, p < 0.001; 0.5 mg/kg, p < 0.05). Moreover, scopolamine also significantly reduced the total licks in HD rats compared with the LD group (group × drug interaction, F3, 54 = 7.19, p < 0.001; group, F1, 18 = 7.02, p < 0.05; drug, F3, 54 = 7.72, p < 0.001). Post hoc comparison confirmed a decrease in the total licks in the HD group at the highest dose used 0.5 mg/kg (p < 0.01) compared with vehicle in the same group. Differences between LD and HD remained significant at all doses tested. Scopolamine administration increased magazine entries in both groups of rats (group × drug interaction, F3, 54 = 2.55, p = 0.07; group, F1, 18 = 2.15, p = 0.16; drug, F3, 54 = 3.94, p < 0.05). Post hoc analyses revealed an increase in magazine entries in both groups only at the highest dose tested 0.5 mg/kg (p < 0.05) compared with vehicle.

Effects of a scopolamine, b methamphetamine METH, and c ketamine on the water intake of high drinkers (HD, n = 10) and low drinkers (LD, n = 10) rats in SIP. Data are expressed as the means ± SEM. *p < 0.05; **p < 0.01; ***p < 0.001 indicate significant differences versus vehicle administration in the same group of rats
The effects of METH on water intake, total licks, and magazine entries on SIP are shown in Fig. 3b and Table 1. METH significantly changed the water intake on SIP (group × drug interaction, F4, 72 = 4.35, p < 0.01; group, F1, 18 = 16.85, p < 0.001; drug, F4, 72 = 54.78, p < 0.001). Post hoc analysis revealed that METH produced a dose-dependent U-inverted curve effect. The lower doses, 0.25 and 0.5 mg/kg, significantly increased water intake in both group of rats, LD (p < 0.05 and p < 0.01) and HD (p < 0.01 and p < 0.05), compared with vehicle in the same group. However, a higher dose of 1.25 mg/kg METH induced a different effect between HD and LD groups, whereas reduced water intake (p < 0.01) was observed in HD rats, and METH treatment did not affect the LD group (p = 0.23) compared with vehicle in the same group. This decrement in water intake in the HD group also revealed a reduction of the significant differences between HD and LD rats in water intake (vehicle, p < 0.001; 1.25 mg/kg, p < 0.05). At the highest dose used, 2.5 mg/kg METH significantly reduced water intake in both groups: LD (p < 0.01) and HD (p < 0.001) compared with vehicle in the same group. METH significantly altered the total licks in SIP (group × drug interaction, F4, 72 = 3.4, p < 0.05; group, F1, 18 = 7.28, p < 0.05; drug, F4, 72 = 51.02, p < 0.001). Post hoc analyses showed that the lowest doses used (0.25 and 0.5 mg/kg) increased the total licks in the LD group (p < 0.05, p < 0.01, respectively) and a dose of 0.5 mg/kg increased the total licks in the HD group (p < 0.01) compared to vehicle in the same group. A dose of 1.25 mg/kg reduced the total licks only in the HD group (p < 0.01) compared with vehicle in the same group. The highest dose of METH reduced the total licks in the HD group (p < 0.001) and showed a decreasing trend in LD animals (p = 0.06) compared with vehicle in the same group. The comparison between the LD and HD groups showed that the dose of 1.25 mg/kg reduced the significant differences by decreasing the total licks response in the HD group (vehicle, p < 0.05; 1.25 mg/kg, p = 0.24). A drug effect via METH administration was observed in magazine entries in both groups (group × drug interaction, F4, 72 = 0.21, p = 0.9; group, F1, 18 = 1.93, p = 0.18; drug, F4, 72 = 6.35, p < 0.001). The post hoc comparison revealed a reduction in magazine entries in both groups only at the highest dose used 2.5 mg/kg (p < 0.01) compared with vehicle.
The effects of ketamine on water intake, total licks, and magazine entries in SIP are shown in Fig. 3c and Table 1. ANOVA showed that ketamine induced significant differences in water intake (group × drug interaction, F4, 72 = 1.09, p = 0.37; group, F1, 18 = 11.64, p < 0.01; drug, F4, 72 = 5.22, p < 0.005), total licks (group × drug interaction, F4, 72 = 1.69, p = 0.16; group, F1, 18 = 4.8, p < 0.05 drug, F4, 72 = 3.28, p < 0.05), and magazine entries (group × drug interaction, F4, 72 = 2.56, p = 0.69; group, F1, 18 = 0.88, p = 0.37; drug, F4, 72 = 2.57, p = 0.05). Post hoc comparison revealed that only the 5-mg/kg dose reduced water intake (p = 0.007) and total magazine entries (p = 0.04) in both groups of rats compared with vehicle, while differences in total licks were only observed between drug doses in both groups but not between any doses compared to vehicle.
Experiment 2. Effects of cannabinoids on SIP
The effects of CBD on water intake and other behavioral measures in SIP are shown in Fig. 4a and Table 2. CBD induced non-selective slight reduction in water intake in both group of rats in SIP (group × drug interaction, F2, 36 = 0.98, p = 0.98; group, F1, 18 = 13.34, p < 0.01; drug, F2, 36 = 3.26, p = 0.05). Post hoc analysis showed that only the 1 mg/kg dose reduced the water intake (p = 0.04) in both group of rats. The measure of total licks showed a non-significant trend in response to CBD administration in SIP (group × drug interaction, F2, 36 = 0.34, p = 0.71; group, F1, 18 = 13.21, p < 0.01; drug, F2, 36 = 2.98, p = 0.06). The significant differences in water intake and total licks were maintained between HD and LD rats at all doses tested. There was also a drug effect following CBD administration in magazine entries in both groups of rats (group × drug interaction, F2, 36 = 1.62, p = 0.21; group, F1, 18 = 0.66, p = 0.42; drug, F2, 36 = 4.03, p < 0.05). Post hoc comparison revealed differences between CBD doses, but not compared with vehicle.

Effects of a cannabidiol CBD, b WIN55212-2, and c AM404 on the water intake of high drinkers (HD, n = 10) and low drinkers (LD, n = 10) in SIP. Data are expressed as the means ± SEM
The effects of WIN-55212 on water intake, total licks, and total magazine entries in SIP are shown in Fig. 4b and Table 2. WIN-55212 did not produce significant effects on water intake in SIP (group × drug interaction, F3, 54 = 0.19, p = 0.90; group, F1, 18 = 7.6, p < 0.05; drug, F3, 54 = 1.26, p = 0.30). No significant effect was observed on the total licks in SIP (group × drug interaction, F3, 54 = 0.05, p = 0.98; group, F1, 18 = 6.35, p < 0.05; drug, F3, 54 = 0.67, p = 0.57). The significant differences in water intake and total licks were maintained between HD and LD rats. No significant differences were observed in the magazine entries in SIP (group × drug interaction, F3, 54 = 0.17, p = 0.92; group, F1, 18 = 0.32, p = 0.32; drug, F3, 54 = 1.54, p = 0.22).
The effects of AM404 on water intake, total licks, and total magazine entries in SIP are shown in Fig. 4c and Table 2. AM404 did not induce significant effects on water intake in SIP (group × drug interaction, F2, 36 = 0.73, p = 0.49; group, F1, 18 = 6.78, p < 0.05; drug, F2, 36 = 0.3, p = 0.74). No significant effects were observed on the total licks in SIP (group × drug interaction, F2, 36 = 1.49, p = 0.24; group, F1, 18 = 8.44, p < 0.01; drug, F2, 36 = 0.66, p = 0.52). AM404 did not affect magazine entries in SIP (group × drug interaction, F2, 36 = 0.3, p = 0.74; group, F1, 18 = 1.64, p = 0.22; drug, F2, 36 = 0.48, p = 0.62).
Discussion
The present study investigated the potential therapeutic role of recreational psychoactive and psychedelic drugs in an animal model of compulsivity. The findings showed that scopolamine and METH administration altered compulsive drinking in SIP. In HD rats, which were characterized by excessive and persistent compulsive drinking in SIP, systemic administration of scopolamine reduced this behavior in a dose-dependent manner. Moreover, METH administration revealed an inverted U-curve effect via an increase at lower doses and decrease at higher doses of compulsive drinking in both groups of rats in SIP. Although 1.25 mg/kg METH revealed a decrease in compulsive water intake in HD rats, this treatment did not affect LD behavior. However, neither ketamine nor cannabinoid drugs administration induced selective effects on compulsive drinking in SIP as LD and HD maintained significant differences at all doses tested.
Scopolamine on compulsive drinking in SIP
The muscarinic acetylcholine MAChR antagonist scopolamine reduced dose-dependent compulsive drinking in HD rats via a reduction in water intake and total licks in SIP. Only the highest dose used reduced the number of magazine entries, indicating a possible modulation of food motivation (Pratt and Kelley, 2004). Our results are in agreement with those obtained by Sanger in 1976, where 1 mg/kg reduced SIP drinking in rats. However, the present results highlight the relevance of the effects of scopolamine at lower doses 0.125, 0.25, and 0.5 mg/k, reducing SIP drinking in a predisposed compulsive population, the HD compared the LD rats, selected by SIP. The present findings support a role for MAChR in the mechanisms underlying compulsive behavior in SIP. Scopolamine also reduces other compulsive behaviors, such as the perseverative behavior displayed in the marble-burying test (Broekkamp et al. 1986) and prevention of stereotypic augmentation (Ohmori et al. 1995). Other muscarinic MAChR antagonists, such as dicyclomine and tropicamide, reduce the activity of the muscarinic M1 receptor and modulate perseverative behavior by decreasing the number of marbles buried in the marble-burying test without producing sedative effects in wild-type animals and in a Fragile X syndrome mouse model (FXS), which is characterized by a wide spectrum of behavioral abnormalities (Veeraragavan et al. 2011a,b). Interestingly, cholinergic supersensitivity has been observed in patients with OCD (Lucey et al. 1993), and patients with fragile X syndrome frequently present symptoms of OCD (Feinstien and Reiss 1998; Hagerman 2002). Moreover, scopolamine can significantly increase the activity of acetylcholinesterase (AChE) levels in the cortex and hippocampus; and curiously in our laboratory, the acute exposure to chlorpyrifos (CPF), a common organophosphate (OP) insecticide whose primary mechanism of neurotoxic action is AChE inhibition, increased compulsive drinking on SIP (Cardona et al. 2006, 2011). Furthermore, scopolamine reduced the stress-induced corticosterone response in an animal model of depression (Katz and Hersh, 1981). Since corticosterone response is implicated in SIP (Dantzer et al. 1988a), this might be another important factor in the observed scopolamine dose-dependent reduction of compulsive drinking in HD animals on SIP. However, other studies have suggested that the muscarinic MAChR agonist oxotremorine methiodide is also effective in reducing compulsive behaviors in marble burying and self-grooming (0.001 and 0.01 mg) in a mouse model of autism (Amodeo et al. 2014). Therefore, altering cholinergic signaling through muscarinic receptors may provide a new therapeutic target for compulsive spectrum disorders and should be extensively investigated.
METH on compulsive drinking in SIP
The administration of METH, a monoamine transmission facilitator that inhibits the dopamine transporter involved in its reuptake, produces an inverted U-shaped dose-dependent effect on compulsive drinking in HD and LD rats in SIP. Thus, low doses (0.25 and 0.5 mg/kg) produced a significant increment, and the highest dose (2.5 mg/kg) reduced compulsive drinking in both groups compared with vehicle. The intermediate dose of 1.25 mg/kg only reduced compulsive water intake in HD rats, reducing the significant differences between HD and LD rats in SIP. In a previous study, we showed a decrease in compulsive drinking after administration of the psychoactive drugs d-amphetamine (0.5 and 1 mg/kg) and cocaine (10 and 20 mg/kg) in SIP (Lopez-Grancha, et al. 2008). METH and d-amphetamine have a similar functional mechanism (Drug Enforcement Administration 2013; Wu et al. 2007). The present results of METH provide additional evidence to previous results of dopaminergic agents in reducing compulsive drinking on SIP (Íbias et al. 2016; for review see Moreno and Flores 2012). Consistent with these results, d-amphetamine (100 ml/kg) decreased the compulsive response of mice in the marble-burying test (Jimenez-Gomez et al. 2011). Previous studies have shown that METH administration (0.5, 1, and 2 mg/kg) reduced impulsive decision-making in a rat model, where rats choose between a delayed fixed (large) amount of water and immediate adjusted (small) amount of water (Richards et al. 1999). Other studies have also demonstrated the anxiolytic-like effects of METH administration (3 and 5 mg/kg) on increasing the time spent and distance traveled by rats in the open arm in the elevated plus maze (Tamaki et al. 2008; Xu et al. 2016). Recent evidence from preclinical and clinical studies indicate that METH under certain circumstances and correct dosing can produce a neuroprotective effect on cognition and neurogenesis after acute brain injury (for review see Rau et al. 2016).
Ketamine on compulsive drinking in SIP
We did not observe a selective effect via ketamine administration at any dose tested (1.25, 2.5, 5, or 10 mg/kg) on HD or LD rats in SIP, suggesting that NMDA receptors might not play a direct role in modulating compulsive behavior. Previous studies have demonstrated that ketamine administration induces dissociative dose-dependent effects; thus, rats receiving ketamine at 20 mg/kg/h showed dissociative behaviors (increased circling, reduced rearing, increased head weave, increased ataxia, and reduced grooming), while doses of 5 and 10 mg/kg/h reduced rearing and grooming (Radford et al. 2017). Acute (0.5–5.0 and 10 mg/kg i.p.) and chronic administration (0.5–2.5 mg/kg daily for 10 days) of ketamine resulted in a dose-dependent and prolonged decrease in immobility in the forced swimming test in rats, confirming the antidepressant-like effects of this drug (Refsgaard et al. 2017; Tizabi et al. 2012). Moreover, other studies have suggested the potential of ketamine to treat posttraumatic stress disorder, showing a decrease in freezing and anxious behaviors in rats and normalization of time spent in the aversive context after chronic ketamine administration (Zhang et al. 2015). In contrast, subchronic MK-801 administration, a potent NMDA antagonist, increased compulsive drinking in SIP (Hawken et al. 2011).
The role of cannabinoids on compulsive drinking in SIP
No significant effects on compulsive drinking were observed in HD and LD rats in response to acute AM404 and WIN55212-2 administration in SIP. CBD, at the dose of 1 mg/kg, induced a non-selective effect by a slight reduction of water intake and magazine entries in both groups in SIP. The results of the present study contrast with previous finding in which CBD, AM404, and WIN55212-2 have demonstrated therapeutic potential as antidepressive and anxiolytic drugs that reduce compulsive behavior in marble burying in rodent models. Different studies have demonstrated that acute administration of 15, 30, 60, and 120 mg/kg CBD (Casarotto et al. 2010; Deiana et al. 2012; Nardo et al. 2014) and 1 and 3 mg/kg WIN55212-2 and AM404 (Gomes et al. 2011) and intracerebroventricular injections of 0.05 μg/mouse (Umathe et al. 2011) reduced the compulsive behavior displayed in the marble-burying test in rodents. Cannabinoids also show an antipsychotic-like profile without inducing extrapyramidal-like effects, and 1 mg/kg WIN55212-2 increased the percentage of pre-pulse inhibition in spontaneously hypertensive rats, a model of schizophrenia (Levin et al. 2014). Self-administration of WIN55212-2 (290.5 ± 0.7 μg/kg) attenuated the psychotomimetic effects on phencyclidine-induced schizotypal symptoms in adult rats, such as hypermotility and the anxiety state (Spano et al. 2013; Umathe et al. 2012). At 15–60 mg/kg, CBD improved schizophrenic symptoms in rodents, inhibiting hyperlocomotion in the circular arena induced by psychotomimetic drugs (Moreira and Guimarães 2005). Moreover, at 1 and 3 mg/kg, WIN55212-2 and AM404 showed anxiolytic-like effects by increasing the time spent in the open arm of the elevated plus maze (Komaki et al. 2015; Patel and Hillard 2006). Furthermore, WIN55212-2 3 mg/kg administration improved decision choice strategies, increasing preferences for advantageous choices and decreasing disadvantageous choices in rats (Gueye et al. 2016). At 1 mg/kg, CBD produced beneficial effects in reversing the contextual fear-conditioning deficit displayed by spontaneously hypertensive rats (Levin et al. 2012). Acute administration of 30 mg/kg CBD; 0.2 mg/kg WIN55212-2; and 0.1, 0.3, 1, and 3 mg/kg AM404 induced therapeutic antidepressant-like effects in rats in the forced swim test (Adamczyk et al. 2008; Bambico et al. 2007; Biojone et al. 2011). Notably, although the interaction between the endocannabinoid and serotoninergic systems plays a primordial role in the regulation of depressive and anxiety behaviors (Umathe et al. 2011), the cannabinoid drugs at the present doses did not induce a selective effect on compulsive drinking behavior displayed by HD rats in SIP. The present study could present a limitation in the doses used; however, they were chosen according to the literature (Adamczyk et al. 2008; Campolongo et al. 2012; Espejo-Porras et al. 2013; Komaki et al. 2015; Zanelati et al. 2010). Further studies should explore if any other doses or treatments with cannabinoid drugs could induce a selective change in compulsive drinking behavior on SIP.
Conclusions
The present study is one of the first to assess the therapeutic potential of recreational psychoactive and psychedelic drugs in compulsive drinking behavior on SIP. The results of the present study showed the efficacy of the muscarinic acetylcholine receptor antagonist scopolamine to dose-dependently and selectively reduce compulsive drinking in HD rats in SIP. Blockade of muscarinic acetylcholine receptors could be of therapeutic interest according to previous data on reducing compulsive behaviors in an animal model of FXS. Administration of METH induced an inverted U-curve effect in compulsive drinking in both groups of rats in SIP. Therefore, the present results support the implication of muscarinic cholinergic system, like the dopamine and serotonin system, in the mechanisms of compulsive behavior in SIP. However, cannabinoid drugs did not show any therapeutic potential in the reduction of compulsive drinking in SIP. The lack of results from cannabinoid administration could reflect the different facets of compulsivity, in which the same treatment might show different results, highlighting different neurochemical mechanisms in the heterogeneity of compulsive behaviors. Future studies on the compulsive phenotype of HD rats in SIP could further identify new therapeutic pharmacological targets for compulsive neuropsychiatric disorders.
References
Adamczyk P, Mccreary AC, Filip M (2008) Activation of endocannabinoid transmission induces antidepressant-like effects in rats. J Physiol Pharmacol 940:217–228
Agarwal V, Yaduvanshi R, Arya A, Gupta PK, Sitholey P (2016) A study of phenomenology, psychiatric co-morbidities, social and adaptive functioning in children and adolescents with OCD. Asian J Psychiatr 22:69–73. https://doi.org/10.1016/j.ajp.2016.04.005
Amerio A, Odone A, Liapis CC, Ghaemi SN (2014) Diagnostic validity of comorbid bipolar disorder and obsessive-compulsive disorder: a systematic review. Acta Psychiatr Scand 129(5):343–358. https://doi.org/10.1111/acps.12250
Amodeo DA, Yi J, Sweeney JA, Ragozzino ME (2014) Oxotremorine treatment reduces repetitive behaviors in BTBR T+ tf/J mice. Front Synaptic Neurosci 6:17. https://doi.org/10.3389/fnsyn.2014.00017
Baer L, Trivedi MH, Huz I, Rush AJ, Wisniewski SR, Fava M (2015) Prevalence and impact of obsessive-compulsive symptoms in depression. J Clin Psychiatry 76(12):1668–1674. https://doi.org/10.4088/JCP.14m09670
Bambico FR, Katz N, Debonnel G, Gobbi G (2007) Cannabinoids elicit antidepressant-like behavior and activate serotonergic neurons through the medial prefrontal cortex. J Neurosci 27(43):11700–11711. https://doi.org/10.1523/JNEUROSCI.1636-07.2007
Bergamaschi MM, Queiroz RH, Chagas MH, de Oliveira DC, De Martinis BS, Kapczinski F, Quevedo J, Roesler R, Schröder N, Nardi AE, Martín-Santos R, Hallak JE, Zuardi AW, Crippa JA (2011) Cannabidiol reduces the anxiety induced by simulated public speaking in treatment-naïve social phobia patients. Neuropsychopharmacology 36(6):1219–1226. https://doi.org/10.1038/npp.2011.6
Biojone C, Casarotto PC, Resstel LB, Zangrossi H Jr, Guimarães FS, Moreira FA (2011) Anti-aversive effects of the atypical antipsychotic, aripiprazole, in animal models of anxiety. J Psychopharmacol 25(6):801–807. doi: https://doi.org/10.1177/0269881110376690.
Blanco C, Potenza MN, Kim SW, Ibáñez A, Zaninelli R, Saiz-Ruiz J, Grant JE (2009) A pilot study of impulsivity and compulsivity in pathological gambling. Psychiatry Res 167(1–2):161–168. https://doi.org/10.1016/j.psychres.2008.04.023
Brett LP, Levine S (1979) Schedule-induced polydipsia suppresses pituitary-adrenal activity in rats. J Comp Physiol Psychol 93(5):946–956. https://doi.org/10.1037/h0077619
Broekkamp CL, Rijk HW, Joly-Gelouin D, Lloyd KL (1986) Major tranquillizers can be distinguished from minor tranquillizers on the basis of effects on marble burying and swim-induced grooming in mice. Eur J Pharmacol 126(3):223–229. https://doi.org/10.1016/0014-2999(86)90051-8
Bruhn J, Scheffer GJ, van Geffen GJ (2017) Clinical application of perioperative multimodal analgesia. Curr Opin Support Palliat Care 11(2):106–111. https://doi.org/10.1097/SPC.
Cadet JL, Krasnova IN (2007) Interactions of HIV and methamphetamine: cellular and molecular mechanisms of toxicity potentiation. Neurotox Res 12(3):181–204. https://doi.org/10.1007/BF03033915
Campolongo P, Ratano P, Manduca A, Scattoni ML, Palmery M, Trezza V, Cuomo V (2012) The endocannabinoid transport inhibitor AM404 differentially modulates recognition memory in rats depending on environmental aversiveness. Front Behav Neurosci 6:11. https://doi.org/10.3389/fnbeh.2012.00011
Campos AC, Fogaça MV, Sonego AB, Guimarães FS (2016) Cannabidiol, neuroprotection and neuropsychiatric disorders. Pharmacol Res 112:119–127. https://doi.org/10.1016/j.phrs.2016.01.033
Cardona D, López-Grancha M, Lopez-Crespo G, Nieto-Escamez F, Sanchez-Santed F, Flores P (2006) Vulnerability of long-term neurotoxicity of chlorpyrifos: effect on schedule-induced polydipsia and a delay discounting task. Psychopharmacology 189(1):47–57. https://doi.org/10.1007/s00213-006-0547-4
Cardona D, Lopez-Crespo G, Sanchez-Amate MC, Flores P, Sanchez-Santed F (2011) Impulsivity as long-term sequelae after chlorpyrifos intoxication: time course and individual differences. Neurotox Res 19(1):128–137. https://doi.org/10.1007/s12640-009-9149-3
Casarotto PC, Gomes FV, Resstel LB, Guimarães FS (2010) Cannabidiol inhibitory effect on marble-burying behaviour: involvement of CB1 receptors. Behav Pharmacol 21(4):353–358. https://doi.org/10.1097/FBP.0b013e32833b33c5
Chamberlain SR, Fineberg NA, Blackwell A, Robbins TW, Sahakian BJ (2006) Motor inhibition and cognitive flexibility in obsessive- compulsive disorder and trichotillomania. Psychiatry Interpers Biol Process 163(7):1282–1284
Crippa JAS, Zuardi AW, Hallak JEC (2010) Therapeutical use of the cannabinoids in psychiatry. Rev Bras Psiquiatr 32(Suppl 1):S56–S66
Cruickshank CC, Dyer KR (2009) A review of the clinical pharmacology of methamphetamine. Addiction 104(7):1085–1099. https://doi.org/10.1111/j.1360-0443.2009.02564.x
Dantzer R, Terlouw C, Mormède P, Le Moal M (1988a) Schedule-induced polydipsia experience decreases plasma corticosterone levels but increases plasma prolactin levels. Physiol Behav 43(3):275–279. https://doi.org/10.1016/0031-9384(88)90187-4
Dantzer R, Terlouw C, Tazi A, Koolhaas JM, Koob GF, Le Moal M (1988b) The propensity for schedule-induced polydipsia is related to differences in conditioned avoidance behaviour and in defense reactions in a defeat test. Physiol Behav 43(3):269–273. https://doi.org/10.1016/0031-9384(88)90186-2
Deiana S, Watanabe A, Yamasaki Y, Amada N, Arthur M, Fleming S, Woodcock H, Dorward P, Pigliacampo B, Close S, Platt B, Riedel G (2012) Plasma and brain pharmacokinetic profile of cannabidiol (CBD), cannabidivarine (CBDV), Δ9-tetrahydrocannabivarin (THCV) and cannabigerol (CBG) in rats and mice following oral and intraperitoneal administration and CBD action on obsessive-compulsive behaviour. Psychopharmacology 219(3):859–873. https://doi.org/10.1007/s00213-011-2415-0
Delva NJ, Chang A, Hawken ER, Lawson JS, Owen JA (2002) Effects of clonidine in schizophrenic patients with primary polydipsia: three single case studies. Prog Neuro-Psychopharmacol Biol Psychiatry 26(2):387–392. https://doi.org/10.1016/S0278-5846(01)00246-9
Diazgranados N, Ibrahim L, Brutsche N, Ameli R, Henter I, Luckenbaurgh D, Machado-Vieira R, Zarate C (2010) Rapid resolution of suicidal ideation after a single infusion of an NMDA antagonist in patients with treatment-resistent major depressive disorder. J Clin Psychiatry 71(12):1605–1611. https://doi.org/10.4088/JCP.09m05327blu.Rapid
Drug Enforcement Administration (2013). Methamphetamine. https://www.deadiversion.usdoj.gov/drug_chem_info/meth.pdf (accessed May 26 2017)
Dundas B, Harris M, Narasimhan M (2007) Psychogenic polydipsia review: etiology, differential, and treatment. Curr Psychiatry Rep 9(3):236–241. https://doi.org/10.1007/s11920-007-0025-7
Espejo-Porras F, Fernández-Ruiz J, Pertwee RG, Mechoulam R, Gracía C (2013) Motor effects of the non-psychotropic phytocannabinoid cannabidiol that are mediated by 5-HT1A receptors. Neuropharmacology 75:155–163. https://doi.org/10.1016/j.neuropharm.2013.07.024
Evenson RC, Jos CJ, Mallya AR (1987) Prevalence of polydipsia among public psychiatric patients. Psychol Rep 60(3):803–807. https://doi.org/10.2466/pr0.1987.60.3.803
Falk JL (1961) Production of polydipsia in normal rats by an intermittent food schedule. Science 133(3447):195–196. https://doi.org/10.1126/science.133.3447.195
Falk JL (1966) Schedule-induced polydipsia as a function of fixed interval length1. J Exp Anal Behav 9(1):37–39. https://doi.org/10.1901/jeab.1966.9-37
Feder A, Parides MK, Murrough JW, Perez AM, Morgan JE, Saxena S, Kirkwood K, aan het Rot M, Lapidus KAB, Wan L, Iosifescu D, Charney DS (2014) Efficacy of intravenous ketamine for treatment of chronic posttraumatic stress disorder. JAMA Psychiatry 71(6):681–188. https://doi.org/10.1001/jamapsychiatry.2014.62
Feinstein C, Reiss AL (1998) Autism: the point of view from fragile X studies. J Autism Dev Disord 28(5):393–405. https://doi.org/10.1023/A:1026000404855
Fineberg NA, Chamberlain SR, Goudriaan AE, Stein DJ, Vanderschuren LKMJ, Gillan CM, Shekar S, Gorwood PAPM, Voon V, Morein-Zamir S, Denys D, Sahakian BJ, Moeller FG, Robbins TW, Potenza MN (2014) New developments in human neurocognition: clinical, genetic, and brain imaging correlates of impulsivity and compulsivity. CNS Spectr 19(01):69–89. https://doi.org/10.1017/S1092852913000801
Furey ML, Khanna A, Hoffman EM, Drevets WC (2010) Scopolamine produces larger antidepressant and antianxiety effects in women than in men. Neuropsychopharmacology 35(12):2479–2488. https://doi.org/10.1038/npp.2010.131
Furey ML, Drevets WC, Hoffman EM, Frankel E, Speer AM, Zarate CA (2013) Potential of pretreatment neural activity in the visual cortex during emotional processing to predict treatment response to scopolamine in major depressive disorder. JAMA psychiatry 70(3):280–290. https://doi.org/10.1001/2013.jamapsychiatry.60
Gillan CM, Fineberg NA, Robbins TW (2017) A trans-diagnostic perspective on obsessive-compulsive disorder 1–21. https://doi.org/10.1017/S0033291716002786
Gomes FV, Resstel LB, Guimarães FS (2011) The anxiolytic-like effects of cannabidiol injected into the bed nucleus of the stria terminalis are mediated by 5-HT1A receptors. Psychopharmacology 213(2–3):465–473. https://doi.org/10.1007/s00213-010-2036-z
Grant JE, Chamberlain SR, Odlaug BL, Potenza MN, Kim SW (2010) Memantine shows promise in reducing gambling severity and cognitive inflexibility in pathological gambling: a pilot study. Psychopharmacology 212(4):603–612. https://doi.org/10.1007/s00213-010-1994-5
Greendyke RM, Bernhardt AJ, Tasbas HE, Lewandowski KS (1998) Polydipsia in chronic psychiatric patients: therapeutic trials of clonidine and enalapril. Neuropsychopharmacology 18(4):272–281. https://doi.org/10.1016/S0893-133X(97)00159-0
Gueye AB, Trigo JM, Vemuri KV, Makriyannis A, Le Foll B (2016) Effects of various cannabinoid ligands on choice behaviour in a rat model of gambling. Behav Pharmacol 27(2-3 spec issue):258-69. https://doi.org/10.1097/FBP
de Haan L, Dudek-Hodge C, Verhoeven Y, Denys D (2009) Prevalence of psychotic disorders in patients with obsessive-compulsive disorder. CNS Spectr 14(8):415–417. https://doi.org/10.1017/S1092852900020381
de Leon J, Verghese C, Tracy JI, Josiassen RC, Simpson GM (1994) Polydipsia and water intoxication in psychiatric patients: a review of the epidemiological literature. Biol Psychiatry 35(6):408–419. https://doi.org/10.1016/0006-3223(94)90008-6
de Leon J, Tracy J, McCann E, McGrory A (2002) Polydipsia and schizophrenia in a psychiatric hospital: a replication study. Schizophr Res 57(2-3):293–301. https://doi.org/10.1016/S0920-9964(01)00292-4
dela Peña I, Lee JC, Lee HL, Woo TS, Lee HC, Sohn AR, Cheong JH (2012) Differential behavioral responses of the spontaneously hypertensive rat to methylphenidate and methamphetamine: lack of a rewarding effect of repeated methylphenidate treatment. Neurosci Lett 514(2):189–193. https://doi.org/10.1016/j.neulet.2012.02.090
Hagerman RJ, Hagerman PJ (2002) The fragile X premutation: into the phenotypic fold. Curr Opin Genet Dev 12(3):278–283. https://doi.org/10.1016/S0959-437X(02)00299-X
Hawken ER, Delva NJ, Reynolds JN, Beninger RJ (2011) Increased schedule-induced polydipsia in the rat following subchronic treatment with MK-801. Schizophr Res 125(1):93–98. https://doi.org/10.1016/j.schres.2010.07.022.
Herkenham M, Lynn AB, Johnson MR, Melvin LS, de Costa BR, Rice KC (1991) Characterization and localization of cannabinoid receptors in rat brain: a quantitative in vitro autoradiographic study. J Neurosci 11(2):563–583
Hollander E, Doernberg E, Shavitt R, Waterman RJ, Soreni N, Veltman DJ, Sahakian BJ, Fineberg NA (2016) The cost and impact of compulsivity: a research perspective. Eur Neuropsychopharmacol 26(5):800–809. https://doi.org/10.1016/j.euroneuro.2016.02.006
Íbias J, Miguéns M, Pellón R (2016) Effects of dopamine agents on a schedule-induced polydipsia procedure in the spontaneously hypertensive rat and in Wistar control rats. J Psychopharmacol 30(9):856–866. https://doi.org/10.1177/0269881116652598
Iftene F, Bowie C, Milev R, Hawken E, Talikowska-Szymczak E, Potopsingh D, Hanna S, Mulroy J, Groll D, Millson R (2013) Identification of primary polydipsia in a severe and persistent mental illness outpatient population: a prospective observational study. Psychiatry Res 210(3):679–683. https://doi.org/10.1016/j.psychres.
Ionescu DF, Luckenbaugh DA, Niciu MJ, Richards EM, Slonena EE, Vande Voort JL, Brutsche NE, Zarate CA (2014) Effect of baseline anxious depression on initial and sustained antidepressant response to ketamine. J Clin Psychiatry 75(09):e932–e938. https://doi.org/10.4088/JCP.14m09049
Ionita R, Postu PA, Beppe GJ, Mihasan M, Petre BA, Hancianu M, Cioanca O, Hritcu L (2017) Cognitive-enhancing and antioxidant activities of the aqueous extract from Markhamia tomentosa (Benth.) K. Schum. stem bark in a rat model of scopolamine. Behav Brain Funct 13(1):5. https://doi.org/10.1186/s12993-017-0123-6
Jimenez-Gomez C, Osentoski A, Woods JH (2011) Pharmacological evaluation of the adequacy of marble burying as an animal model of compulsion and/or anxiety. Behav Pharmacol 22(7):711–713. https://doi.org/10.1097/FBP.0b013e32834afebe
Katz RJ, Hersh S (1981) Amitriptyline and scopolamine in an animal model of depression. Neurosci Biobehav Rev 5(2):265–271. https://doi.org/10.1016/0149-7634(81)90008-7
Kim M, Lee TH, Choi JS, Kwak YB, Hwang WJ, Kim T, Lee JY, Lim JA, Park M, Kim YJ, Kim SN, Kim DJ, Kwon JS (2017) Neurophysiological correlates of altered response inhibition in internet gaming disorder and obsessive-compulsive disorder: perspectives from impulsivity and compulsivity. Sci Rep 7:41742. https://doi.org/10.1038/srep41742
Komaki A, Hashemi-firouzi N, Shojaei S, Souri Z, Heidari S, Shahidi S (2015) Study the effect of endocannabinoid system on rat behavior in elevated plus-maze. Basic Clin Neurosci 6(3):147–153
Krasnova IN, Cadet JL (2009) Methamphetamine toxicity and messengers of death. Brain Res Rev 60(2):379–407. https://doi.org/10.1016/j.brainresrev.2009.03.002
Lally N, Nugent AC, Luckenbaugh DA, Ameli R, Roiser JP, Zarate CA (2014) Anti-anhedonic effect of ketamine and its neural correlates in treatment-resistant bipolar depression. Transl Psychiatry 4(10):e469. https://doi.org/10.1038/tp.2014.105
Levin R, Almeida V, Peres FF, Calzavara MB, da Silva ND, Suiama MA, Niigaki ST, Zuardi AW, Hallak JE, Crippa JA, Abílio VC (2012) Antipsychotic profile of cannabidiol and rimonabant in an animal model of emotional context processing in schizophrenia. Curr Pharm Des 18(32):4960–4965. https://doi.org/10.2174/138161212802884735
Levin R, Peres FF, Almeida V, Calzavara MB, Zuardi AW, Hallak JE, Crippa JA, Abílio VC (2014) Effects of cannabinoid drugs on the deficit of prepulse inhibition of startle in an animal model of schizophrenia: the SHR strain. Front Pharmacol 5:10. https://doi.org/10.3389/fphar.2014.00010
Leweke FM, Piomelli D, Pahlisch F, Muhl D, Gerth CW, Hoyer C, Klosterkötter J, Hellmich M, Koethe D (2012) Cannabidiol enhances anandamide signaling and alleviates psychotic symptoms of schizophrenia. Transl Psychiatry 2(3):e94. https://doi.org/10.1038/tp.2012.15
López-Grancha M, Lopez-Crespo G, Sanchez-Amate MC, Flores P (2008) Individual differences in schedule-induced polydipsia and the role of GABAergic and dopaminergic systems. Psychopharmacology 197(3):487–498. https://doi.org/10.1007/s00213-007-1059-6
Lucey JV, Butcher G, Clare AW, Dinan TG (1993) The anterior pituitary responds normally to protirelin in obsessive-compulsive disorder: evidence to support a neuroendocrine serotonergic deficit. Acta Psychiatr Scand 87(6):384–388. https://doi.org/10.1111/j.1600-0447.1993.tb03392.x
Marinova Z, Chuang D-M, Fineberg N (2017) Glutamate-modulating drugs as a potential therapeutic strategy in obsessive-compulsive disorder. Curr Neuropharmacol doi 15(7):977–995. https://doi.org/10.2174/1570159X15666170320104237
Merchán A, Navarro SV, Klein AB, Aznar S, Campa L, Suñol C, Moreno M, Flores P (2017) Tryptophan depletion affects compulsive behaviour in rats: strain dependent effects and associated neuromechanisms. Psychopharmacology 234(8):1223–1236. https://doi.org/10.1007/s00213-017-4561-5
Micale V, Cristino L, Tamburella A, Petrosino S, Leggio GM, Drago F, Di Marzo V (2009) Altered responses of dopamine D3 receptor null mice to excitotoxic or anxiogenic stimuli: possible involvement of the endocannabinoid and endovanilloid systems. Neurobiol Dis 36(1):70–80. https://doi.org/10.1016/j.nbd.2009.06.015
Mittleman G, Jones GH, Robbins TW (1988) The relationship between schedule-induced polydipsia and pituitary-adrenal activity: pharmacological and behavioral manupulations. Behav Brain Res 28(3):315–324. https://doi.org/10.1016/0166-4328(88)90134-9
More SV, Choi D-K (2011) Promising cannabinoid-based therapies for Parkinson’s disease: motor symptoms to neuroprotection 10:17. doi: https://doi.org/10.1186/s13024-015-0012-0, Promising cannabinoid-based therapies for Parkinson’s disease: motor symptoms to neuroprotection, 1
Moreira FA, Guimarães FS (2005) Cannabidiol inhibits the hyperlocomotion induced by psychotomimetic drugs in mice. Eur J Pharmacol 512(2-3):199–205. https://doi.org/10.1016/j.ejphar.2005.02.040
Moreno M, Flores P (2012) Schedule-induced polydipsia as a model of compulsive behavior: Neuropharmacological and neuroendocrine bases. Psychopharmacology 219(2):647–659. https://doi.org/10.1007/s00213-011-2570-3
Moreno M, Cardona D, Gómez MJ, Sánchez-Santed F, Tobeña A, Fernández-Teruel A, Campa L, Suñol C, Escarabajal MD, Torres C, Flores P (2010) Impulsivity characterization in the Roman high- and low-avoidance rat strains: behavioral and neurochemical differences. Neuropsychopharmacology 35(5):1198–1208. https://doi.org/10.1038/npp.2009.224
Moreno M, Gutiérrez-Ferre VE, Ruedas L, Campa L, Suñol C, Flores P (2012) Poor inhibitory control and neurochemical differences in high compulsive drinker rats selected by schedule-induced polydipsia. Psychopharmacology 219(2):661–672. https://doi.org/10.1007/s00213-011-2575-y
Nardo M, Casarotto PC, Gomes FV, Guimarães FS (2014) Cannabidiol reverses the mCPP-induced increase in marble-burying behavior. Fundam Clin Pharmacol 28(5):544–550. https://doi.org/10.1111/fcp.12051
Navarro SV, Gutiérrez-Ferre V, Flores P, Moreno M (2015) Activation of serotonin 5-HT2 A receptors inhibits high compulsive drinking on schedule-induced polydipsia. Psychopharmacology 232(4):683–697. https://doi.org/10.1007/s00213-014-3699-7
Navarro SV, Alvarez R, Colomina MT, Sanchez-Santed F, Flores P, Moreno M (2016) Behavioral biomarkers of schizophrenia in high drinker rats: a potential Endophenotype of compulsive neuropsychiatric disorders. Schizophr bull pii: sbw141
Ohmori T, Abekawa T, Koyama T (1995) Scopolamine prevents augmentation of stereotypy induced by chronic methamphetamine treatment. Psychopharmacology 121(2):158–163. https://doi.org/10.1007/BF02245625
Patel S, Hillard CJ (2006) Pharmacological evaluation of cannabinoid receptor ligands in a mouse model of anxiety: further evidence for an anxiolytic role for endogenous cannabinoid signaling. J Pharmacol Exp Ther 318(1):304–311. https://doi.org/10.1124/jpet.106.101287
Petryshen TL, Lewis MC, Dennehy KA, Garza JC, Fava M (2016) Antidepressant-like effect of low dose ketamine and scopolamine co-treatment in mice. Neurosci Lett 620:70–73. https://doi.org/10.1016/j.neulet.2016.03.051
Phelps L, Brutsche N (2009) Family history of alcohol dependence and initial antidepressant response to an NMDA antagonist. Biol Psychiatry 65(2):181–184. https://doi.org/10.1016/j.biopsych.2008.09.029
Platt B, Beyer CE, Schechter LE, Rosenzweig-Lipson S (2008) Schedule-induced polydipsia: a rat model of obsessive-compulsive disorder Curr Protoc Neurosci Chapter 9:Unit 9.27. doi: https://doi.org/10.1002/0471142301.ns0927s43
Pratt WE, Kelley AE (2004) Nucleus accumbens acetylcholine regulates appetitive learning and motivation for food via activation of muscarinic receptors. Behav Neurosci 118(4):730–739. https://doi.org/10.1037/0735-7044.118.4.730
Radford KD, Park TY, Lee BH, Moran S, Osborne LA, Choi KH (2017) Dose-response characteristics of intravenous ketamine on dissociative stereotypy, locomotion, sensorimotor gating, and nociception in male Sprague-Dawley rats. Pharmacol Biochem Behav 153:130–140. https://doi.org/10.1016/j.pbb.2016.12.014
Rau TF, Kothiwal AS, Rova AR, Brooks DM, Poulsen DJ (2012) Treatment with low-dose methamphetamine improves behavioral and cognitive function after severe traumatic brain injury. J Trauma Acute Care Surg 73(2 Suppl 1):S165–S172. https://doi.org/10.1097/TA.0b013e318260896a
Rau T, Ziemniak J, Poulsen D (2016) The neuroprotective potential of low-dose methamphetamine in preclinical models of stroke and traumatic brain injury. Prog Neuro-Psychopharmacol Biol Psychiatry 64:231–236. https://doi.org/10.1016/j.pnpbp.2015.02.013
Refsgaard LK, Pickering DS, Andreasen JT (2017) Investigation of antidepressant-like and anxiolytic-like actions and cognitive and motor side effects of four N -methyl- D-aspartate receptor antagonists in mice. Behav Pharmacol 28(1):37–47. https://doi.org/10.1097/FBP.
Richards JB, Sabol KE, de Wit H (1999) Effects of methamphetamine on the adjusting amount procedure, a model of impulsive behavior in rats. Psychopharmacology 146(4):432–439. https://doi.org/10.1007/PL00005488
Rickelt J, Viechtbauer W, Lieverse R, Overbeek T, van Balkom AJ, Marcelis M, Eikelenboom M, Tibi L, Schruers KRJ (2016) The relation between depressive and obsessive-compulsive symptoms in obsessive-compulsive disorder: results from a large, naturalistic follow-up study. J Affect Disord 203:241–247. https://doi.org/10.1016/j.jad.2016.06.009
Ruehle S, Rey AA, Remmers F, Lutz B (2012) The endocannabinoid system in anxiety, fear memory and habituation. J Psychopharmacol 26(1):23–39. https://doi.org/10.1177/0269881111408958
Russell VA (2017) Antihyperalgesic Activities of Endocannabinoids in a Mouse Model of Antiretroviral-Induced Neuropathic 8:1–11. doi: https://doi.org/10.3389/fphar.2017.00136, Antihyperalgesic activities of endocannabinoids in a mouse model of antiretroviral-induced neuropathic pain
Sanger DJ (1976) Scopolamine and adjunctive drinking in rats. Psychopharmacology 48(3):307–309. https://doi.org/10.1007/BF00496867
Schubart CD, Sommer IEC, van Gastel WA, Goetgbuer RL, Kahn RS, Boks MPM (2011) Cannabis with high cannabidiol content is associated with fewer psychotic experiences. Schizophr Res 130(1-3):216–221. https://doi.org/10.1016/j.schres.2011.04.017
Sos P, Klirova M, Novak T, Kohutova B, Horacek J, Palenicek T (2013) Relationship of ketamine’s antidepressant and psychotomimetic effects in unipolar depression. Neuro Endocrinol Lett 34(4):287–293
Spano MS, Fattore L, Cadeddu F, Fratta W, Fadda P (2013) Chronic cannabinoid exposure reduces phencyclidine-induced schizophrenia-like positive symptoms in adult rats. Psychopharmacology 225(3):531–542. https://doi.org/10.1007/s00213-012-2839-1
Swets M, Dekker J, van Emmerik-van Oortmerssen K, Smid GE, Smit F, de Haan L, Schoevers RA (2014) The obsessive compulsive spectrum in schizophrenia, a meta-analysis and meta-regression exploring prevalence rates. Schizophr Res 152(2–3):458–468. https://doi.org/10.1016/j.schres.2013.10.033
Tamaki R, Yoshikawa M, Shinomiya T, Hashimoto A, Kawaguchi M, Byrne DW, Kobayashi H (2008) Acute administration of methamphetamine decreases the mRNA expression of diazepam binding inhibitor in rat brain. Tokai J Exp Clin Med 33(1):51–56
Tizabi Y, Bhatti BH, Manaye KF, Das JR, Akinfiresoye L (2012) Antidepressant-like effects of low ketamine dose is associated with increased hippocampal AMPA/NMDA receptor density ratio in female Wistar-Kyoto rats. Neuroscience 213:72–80. https://doi.org/10.1016/j.neuroscience.2012.03.052
Torres AR, Ferrão YA, Shavitt RG, Diniz JB, Costa DL, do Rosário MC, Miguel EC, Fontenelle LF (2014) Panic disorder and agoraphobia in OCD patients: clinical profile and possible treatment implications. Compr Psychiatry 55(3):588–597. https://doi.org/10.1016/j.comppsych.2013.11.017
Torres AR, Fontenelle LF, Shavitt RG, Ferrão YA, do Rosário MC, Storch EA, Miguel EC (2016) Comorbidity variation in patients with obsessive-compulsive disorder according to symptom dimensions: results from a large multicentre clinical sample. J Affect Disord 190:508–516. https://doi.org/10.1016/j.jad.2015.10.051
Umathe SN, Manna SSS, Jain NS (2011) Involvement of endocannabinoids in antidepressant and anti-compulsive effect of fluoxetine in mice. Behav Brain Res 223(1):125–134. https://doi.org/10.1016/j.bbr.2011.04.031
Umathe SN, Manna SS, Jain NS (2012) Endocannabinoid analogues exacerbate marble-burying behavior in mice via TRPV1 receptor. Neuropharmacology Apr 62(5–6):2024–2033. https://doi.org/10.1016/j.neuropharm.2011.12.030
Veeraragavan S, Bui N, Perkins JR, Yuva-Paylor LA, Carpenter RL, Paylor R (2011a) Modulation of behavioral phenotypes by a muscarinic M1 antagonist in a mouse model of fragile X syndrome. Psychopharmacology 217(1):143–151. https://doi.org/10.1007/s00213-011-2276-6
Veeraragavan S, Bui N, Perkins JR, Yuva-Paylor LA, Paylor R (2011b) The modulation of fragile X behaviors by the muscarinic M4 antagonist, tropicamide. Behav Neurosci 125(5):783–790. https://doi.org/10.1037/a0025202
Wu L-T, Pilowsky DJ, Schlenger WE, Galvin DM (2007) Misuse of methamphetamine and prescription stimulants among youths and young adults in the community. Drug Alcohol Depend 89(2-3):195–205. https://doi.org/10.1016/j.drugalcdep.2006.12.020
Xu P, Qiu Y, Zhang Y, Bai Y, Xu P, Liu Y, Kim JH, Shen HW (2016) The effects of 4-methylethcathinone on conditioned place preference, locomotor sensitization, and anxiety-like behavior: a comparison with methamphetamine. Int J Neuropsychopharmacol 19(4):1–7. https://doi.org/10.1093/ijnp/pyv120
Yamazaki M, Okabe M, Yamamoto N, Yarimizu J, Harada K (2015) Novel 5-HT5A receptor antagonists ameliorate scopolamine-induced working memory deficit in mice and reference memory impairment in aged rats. J Pharmacol Sci 127(3):362–369. https://doi.org/10.1016/j.jphs.2015.02.006
Zanelati TV, Biojone C, Moreira FA, Guimarães FS, Joca SRL (2010) Antidepressant-like effects of cannabidiol in mice: possible involvement of 5-HT1A receptors. Br J Pharmacol 159(1):122–128. https://doi.org/10.1111/j.1476-5381.2009.00521.x
Zarate CA, Singh JB, Carlson PJ, Brutsche NE, Ameli R, Luckenbaugh DA, Charney DS, Manji HK (2006) A randomized trial of an N-methyl-D-aspartate antagonist in treatment-resistant major depression. Arch Gen Psychiatry 63(8):856–864. https://doi.org/10.1001/archpsyc.63.8.856
Zarate C, Duman RS, Liu G, Sartori S, Quiroz J, Murck H (2013) New paradigms for treatment-resistant depression. Ann N Y Acad Sci 1292(1):21–31. https://doi.org/10.1111/nyas.12223
Zhang L-M, Zhou W-W, Ji Y-J, Li Y, Zhao N, Chen H, Xue R, Mei X, Zhang Y, Wang H, Li Y (2015) Anxiolytic effects of ketamine in animal models of posttraumatic stress disorder. Psychopharmacology 232(4):663–672. https://doi.org/10.1007/s00213-014-3697-9
Zuardi AW, Morais SL, Guimarães FS, Mechoulam R (1995) Antipsychotic effect of cannabidiol. J Clin Psychiatry 56(10):485–486
Acknowledgements
This work was supported by grants from the Ministerio de Economía y Competitividad, Spanish Government (grant number PSI2015-70037-R MINECO-FEDER).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declared that they have no conflicts of interest.
Rights and permissions
About this article

Cite this article
Martín-González, E., Prados-Pardo, Á., Mora, S. et al. Do psychoactive drugs have a therapeutic role in compulsivity? Studies on schedule-induced polydipsia. Psychopharmacology 235, 419–432 (2018). https://doi.org/10.1007/s00213-017-4819-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-017-4819-y