
Abstract
Various studies have confirmed that prostaglandins (PG) alter the bladder motor activity and micturition reflex in both human and animals. However, no sufficient data is reported about the effect of cyclooxygenase (COX) inhibitors neither in normal bladder physiology nor in pathological conditions. This study aims to compare the potential effects of some COX inhibitors with varying COX-1/COX-2 selectivities (indomethacin, ketoprofen, and diclofenac) with that of the selective COX-2 inhibitor (DFU) on bladder function. The role played by some PGs and their receptors in controlling detrusor muscle function in normal condition and in cystitis is also studied. Organ bath experiments were performed using isolated rat detrusor muscle. Direct and neurogenic contractions were induced using ACh and electric stimulation (EFS), respectively. A model of hemorrhagic cystitis was induced by single injection of cyclophosphamide (300 mg/kg) in rats, and confirmed by histophathological examination. Results are expressed as mean ± SEM of 5–9 rats. Alprostadil and iloprost (1 nM- 10 µM) concentration-dependently potentiated ACh (100 μM)- and EFS (4 Hz)-induced contraction, with maximum potentiation of 40.01 ± 5.29 and 27.59 ± 6.64%, respectively, in case of ACh contractions. In contrast, ONO-AE1-259 (selective EP2 agonist, 1 nM–10 μM) inhibited muscle contraction. SC51322 (EP1-antagonist, 10 μM) and RO1138452 (IP antagonist, 10 μM) inhibited both direct and neurogenic responses. Hemorrhagic cystitis reduced both ACh and EFS responses as well as the potentiatory effect of iloprost and the inhibitory effect of RO1138452 on ACh contractions. ONO-AE3-237 (DP1 antagonist, 1 μM) significantly potentiated contractions in cystitis but showed no effect in normal bladder. A significant inhibition of contractile response was observed in presence of indomethacin, ketoprofen, and diclofenac at all tested concentrations (20, 50, and 100 μM). Highest effect was induced by diclofenac. The effect of these COX inhibitors on EFS contractions was intensified in case of cystitis, indomethacin being the most potent. Atropine (1 nM) significantly reduced indomethacin effect on ACh contraction only in normal rats. On the other hand, DFU (10−6 M) significantly potentiated the contractile effect of ACh in case of cystitis although it showed no effect in normal rats. EP1 receptors seem to play an important role in rat bladder contractility. DP1 receptors as COX-2, on the other hand, gain an important role only in case of cystitis. The use of non-selective COX inhibitors in cystitis may be associated with bladder hypoactivity; selective COX-2 inhibitors may be a safer option.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Urinary bladder disorders are among the most common disorders of the lower genitourinary tract. They can be categorized into different conditions including storage conditions such as overactive bladder (OAB) and underactive bladder (UAB), obstructive conditions such as bladder outlet obstruction, inflammatory conditions such as interstitial and hemorrhagic cystitis in addition to bladder cancer (Andersson 2016). In 2008, almost 45.2% of the worldwide populations have been reported to be affected by at least one lower urinary tract symptom including 10.7% suffering from OAB (Irwin et al. 2011).
The major neurotransmitters involved in contractility of bladder smooth muscles are acetylcholine (ACh) and adenosine triphosphate (ATP). Muscarinic and purinergic receptors activation causes elevation of intracellular calcium (Ca2+) level and activation of voltage-dependent L-type Ca2+ channels resulting in contraction of the muscle (Ding et al. 2009). The contractile receptors in the detrusor muscles include muscarinic receptors (M2 and M3) which are the dominant ones, and purinergic receptors (P2X1). M3 receptors are the main contractile component. However, M2 receptors are the more expressed ones and are responsible mainly for the recontraction of the detrusor muscles after relaxation (Uchiyama and Chess-Williams 2004; Hegde 2006).
The exact role of endogenous prostaglandins (PGs) in different tissues of lower urinary tract is not well elucidated, but various studies have confirmed that exogenous PGs alter the bladder motor activity and micturition reflex in both human and animals (Maggi 1992; Anderson 1993). The main PGs synthesized in the bladder under physiological and pathological conditions are PGE2, PGF2α, PGI2, and thromboxane A2 (TXA2) (Rahnama'i et al. 2012). The major effective PG in rabbit bladder is PGE2 followed by PGI2, PGF2α, and TXA2 (Leslie et al. 1984). In rat bladder, PGI2 is the major effective PG followed by PGE2 and TXA2 (Jeremy et al. 1984). In human bladder, the main PG is PGI2 followed by PGE2, PGF2α, and TXA2 (Jeremy et al. 1987). EP1 and EP3 receptors are reported to be involved in bladder contraction, while EP2 and EP4 induce relaxation (Coleman et al. 1994). The inhibition of PG synthesis was also shown to decrease the cholinergically mediated contraction of guinea pig urinary bladder (Rahnama'i et al. 2013). PGE2 level was found to be higher in patients with OAB (Tanaka et al. 2011). The effect of PGE2 on detrusor muscles is mainly mediated via EP1 receptors, as its effect was blocked by EP1 receptor antagonist in rats with OAB. Moreover, EP1 receptors are involved in micturition reflex initiation in normal rats and also in case of bladder outlet obstruction (Lee et al. 2007).
Cyclooxygenase (COX) inhibitors include non-selective COX inhibitors and selective COX-2 inhibitors. Most of them are competitive inhibitors of COX enzyme at its active site resulting in inhibiting the formation of the pro-inflammatory PGs, so they are mainly used as analgesic and anti-inflammatory medications. Besides, some have antipyretic effect (Simmons et al. 2004). In rat urinary bladder, COX-1 is the constitutive isoform while COX-2 is expressed under inflammatory conditions as in case of hemorrhagic cystitis (Klinger et al. 2007). As reported by Lecci et al. (2000), COX-1 isoenzyme partially modulates the threshold of micturition reflex in normal rats, while COX-2 is involved in the urodynamic changes which result from urinary bladder inflammation. Takagi-Matsumoto et al. (2004) reported that ketoprofen was more effective than indomethacin in increasing bladder volume capacity in normal rats without affecting micturition, while in cystitis, they increased bladder capacity and decreased micturition frequency. Cardozo et al. (1980) reported that flurbiprofen is effective in reducing urinary incontinence in patients with OAB, and suggested that it could be suitable for patients with detrusor instability. However, indomethacin could not inhibit carbachol-induced contraction in isolated porcine bladder (Badawi et al. 2008).
Most of the reported data investigated the effect of COX inhibitors on bladder function in models of OAB. However, little data is reported about the potential negative role of COX inhibitors in bladder physiology neither in humans nor in experimental animals.
Aim of the work
This study has as hypothesis to test whether inhibiting PG synthesis or action would affect bladder contractility, and whether the use of selective COX-2 inhibitor will be associated with less side effects than the conventional non-selective COX inhibitors, in normal and inflamed bladders. In this regard, the study aims to compare the potential effects of some COX inhibitors with varying COX-1/COX-2 selectivities (such as indomethacin, ketoprofen, and diclofenac) with that of the selective COX-2 inhibitor (DFU) on bladder function. The involvement of effective PGs’ receptors in the pathophysiology of cystitis as well as the differential effects of nonsteroidal anti-inflammatory drugs (NSAIDs) with various selectivities in such condition in comparison to normal status will also be investigated. The study is expected to shed some light on the PG-related pathways involved in cyclophosphamide (CYP)-induced hemorrhagic cystitis as well as the consequences of the use of NSAIDs in the management of such condition.
Material and methods
Chemicals
The chemicals used in this study and their respective sources were as follows: acetylcholine chloride (Sigma), alprostadil (Tocris), atropine sulfate (Merck), cyclophosphamide monohydrate (Endoxan® vial, Baxter), 5,5-dimethyl-3-(3-fluorophenyl)-4-(4-methylsulphonyl) phenyl-2(5H)-furanone (DFU, Cayman), diclofenac sodium (Sigma), iloprost (Tocris), indomethacin (Sigma), ketoprofen (European Egyptian Pharm. Co.), (Z)-7-[(1R,2R,3R,5R)-5-chloro-3-hydroxy-2-[(E,4S)-4-hydroxy-4-(1-prop-2-enylcyclobutyl)but-1enyl] cyclopentyl]hept-5-enoic acid (ONO-AE1-259) and 2-[1-[4-[[(2S)-4,6-dimethyl-2,3-dihydro-1,4-benzoxazin-2-yl]methoxy]benzoyl]-2-methylindol-4-yl]acetic acid (ONO-AE3-237) are gift from Ono Pharmaceutical Co.Ltd, Chuo-ku, Osaka, japan, N-[4-[(4-propan-2-yloxyphenyl)methyl]phenyl]-4,5-dihydro-1H-imidazol-2-amine (RO1138452 hydrochloride, Cayman), 3-chloro-N′-[3-(furan-2-ylmethylsulfanyl)propanoyl]-6H-benzo[b][1,5]benzoxazepine-5-carbohydrazide (SC51322, Tocris).
For preparation of 10% (v/v) formalin solution, a stock solution (40% v/v formaldehyde) was diluted 1:9 with distilled water containing phosphate buffer (4 g/L NaH2PO4, 6.5 g/L Na2HPO4) to keep neutral pH.
The isolated rat detrusor smooth muscle
Experiments were carried out on male albino Wistar rats weighing 250–270 g (age; 3–4 months). The animals were obtained from the Alexandria University Faculty of Pharmacy Animal House. The rats were housed in plastic cages, and held under a 12-h dark/light cycle, with free access to water and chow. Study protocols comply with the guidelines for the proper conduct of animal experiments and are approved by the Animal Care and Use Committee (ACUC)—Faculty of Pharmacy—Alexandria University. All applicable international, national, and institutional guidelines for the care and use of animals were followed. Preparation of the muscle was performed as described by Luheshi and Zar (1991). Rats were anesthetized by intraperitoneal injection of thiopentone (50 mg/kg) and then sacrificed by exsanguination. In order to expose and isolate the urinary bladder, the lower abdomen was opened and the bladder separated from associated connective tissues and blood vessels. The bladder was then excised above its trigone to isolate only the detrusor muscles of the bladder away from its neck and urethral sphincter. The bladder was then emptied of any residual urine and washed several times using Krebs solution. The bladder was then fixed in a petri dish and two lateral incisions along its longitudinal axis were made and the bladder was unfolded giving rectangular sheet which was cut longitudinally to give two strips of detrusor muscle 2 × 15 mm. The detrusor muscle was fixed at one end between two parallel platinum electrodes 4 to 5 mm apart and mounted in a 10-mL organ bath containing Krebs solution (composition in mM; NaCl 118, KCl 4.7. CaCl2 2.5, MgSO4·7H2O 1.2, KH2PO4 1.2, NaHCO3 25, and glucose 11, pH = 7.4), kept at 37 °C and continuously aerated with 95% (v/v) O2 and 5% (v/v) CO2. The other end was tied and attached to a force displacement transducer (Grass FT-03) which in turn was connected to a computerized data acquisition system through an MLAC11 Grass adapter cable. Tension studies were performed using Lab Chart-7 pro software (Power Lab 4/35, model ML 866/P; AD Instrument Pty Ltd., Castle Hill, Australia). The tissue was left to equilibrate under a resting tension of 0.5 g for 60 min during which the physiological solution was replaced each 15 min. Meanwhile, time-matched control experiment was conducted over the duration of the experiment.
The contractile responses to ACh (in g tension) in absence and presence of the tested drug were measured and the percentage change from control was calculated. For electric field stimulation (EFS) experiments, each strip was subjected to increasing frequencies (1, 4, and 16 Hz; voltage 80 V, pulse duration 1 ms, 3 min interval). After incubation with the studied drug, the frequency response curve or the submaximal frequency was repeated. The contractile responses to EFS in absence and presence of the tested drug were measured and the percentage change from control was calculated.
The tested COX inhibitors were selected based on their IC50 ratio (COX-1/COX-2); indomethacin was more COX-1 inhibitor (IC50 ratio = 0.0125) followed by ketoprofen (IC50 ratio = 0.016), while diclofenac was more COX-2 inhibitor (IC50 ratio = 1.97) and DFU was chosen as a highly selective COX-2 inhibitor (IC50 ratio = 74) (Brideau et al. 2001; Rao and Knaus 2008).
Induction of hemorrhagic cystitis
As described by Wada et al. (2013), rats (weighing 250–270 g) were divided into two groups; the first group was injected with saline and served as control, while the second group received a single intraperitoneal (I.P.) injection of CYP (300 mg/kg) to induce hemorrhagic cystitis. Forty-eight hours after injection, rats were sacrificed, the urinary bladder was isolated, and half of the bladder was fixed in formalin and prepared for histopathological examination, while the other half was used for tension studies experiments where detrusor muscle contractility was measured and compared to the control group in response to ACh and EFS-induced contraction.
Histopathological examination of rat urinary bladder
To confirm the induction of CYP-induced hemorrhagic cystitis, the isolated bladder was fixed in 10% (v/v) buffered formalin for 24 h. Tissue taken from the detrusor smooth muscle region was then embedded in paraffin and stained with hematoxylin-eosin (H & E). A pathologist blinded to the study scored the tissue for the mean histological damages including inflammation, hemorrhage, ulceration, and edema on a scale from 1 to 4 (1 for normal and 4 for severe changes) as described by Gray et al. (1986) and Oter et al. (2004) and shown in Table 1.
Data analysis and statistics
All the obtained results are expressed as mean ± SEM. Values between parentheses (n) represent the number of animals in each group. For analysis of unpaired data, the Student’s t test was used, while for multiple comparison, analysis of variance (ANOVA) was used followed by Dunnett’s post-test. Statistical significance was considered at the level of P < 0.05. Analysis was performed using computer software program GraphPad Prism 6.01.
Results
Effect of selected prostanoids and prostanoids receptors antagonists on rat detrusor muscle
Low concentrations of ACh (10−9, 10−8 M) showed no effect on rat detrusor muscles, while at concentrations (10−7–10−3 M), ACh induced a concentration-dependent contraction of rat detrusor muscle with a maximum of 2.22 ± 0.23 g (at 10−3 M). EFS (1–16 Hz) also induced a frequency-dependent contraction of rat detrusor muscle with a maximum attained contraction of 4.12 ± 0.36 g (at 16 Hz) (Supplement 1). Effect of alprostadil (synthetic PGE1 10−9–10−5 M), iloprost (PGI2 analogue, 10−9–10−5 M), or ONO-AE1-259 (selective EP2 agonist, 10−9–10−5 M) was tested on the contractile effect of a submaximal ACh concentration of 10−4 M and a submaximal frequency of 4 Hz on rat detrusor muscles and compared to time-matched control over the experiment duration. Alprostadil caused a concentration-dependent potentiation of both ACh and EFS-induced contraction with a maximum attained potentiation of 40.01 ± 5.29 and 25.32 ± 4.61%, respectively. Iloprost caused a significant potentiation of ACh and EFS-induced contraction only at high concentrations (10−6, 10−5 M) with a maximum of 27.59 ± 6.64 and 19.52 ± 2.83%, respectively. On the other hand, ONO-AE1-259 caused a concentration-dependent inhibition of both ACh and EFS-induced contraction. A significant inhibition of ACh-induced contraction was observed at concentrations (10−7–10−5 M) of ONO-AE1-259 with a maximum attained inhibition of 16.37 ± 1.20%. In case of EFS, a significant inhibition was observed at concentrations (10−6, 10−5 M) of ONO-AE1-259 with a maximum of 14.07 ± 4.18% (Fig. 1).

Effect of alprostadil (10−9–10−5 M) iloprost (10−9–10−5 M) and ONO-AE1-259 (10−9–10−5 M) on ACh (10−4 M, left panel)- and electric field stimulation (4 Hz, EFS, right panel)-induced contraction of isolated rat detrusor muscles. Responses are expressed as mean ± SEM. * denotes significant difference from control (p < 0.05). Values between parentheses indicate the number of animals
Cyclophosphamide-induced hemorrhagic cystitis was confirmed by the histopathological changes shown in the urinary bladder of CYP-injected rats compared to control as shown in Table 2 and Fig. 2. As shown in Fig. 3, a downward shift and significant inhibition of the concentration response curve of ACh as well as the frequency response curve were observed in CYP-injected rats compared to control. The maximum attained contraction induced by ACh (at 10−3 M) was 0.83 ± 0.07 g in CYP-injected rats compared to 2.14 ± 0.17 g in control (Fig. 3a). Similarly, the contractile effect induced by low (1 Hz) and high frequencies (16 Hz) was significantly lower in CYP-injected rats amounting to 0.18 ± 0.02 and 0.81 ± 0.07 g, respectively, compared to 0.65 ± 0.09 and 3.83 ± 0.50 g in control (Fig. 3b). The potentiatory effect of alprostadil on ACh-induced contraction of detrusor muscle isolated from CYP-injected rats was not significantly different from that of control, percentages potentiation were 32.51 ± 5.37 and 40.01 ± 5.29%, respectively. However, the potentiatory effect of iloprost at 10−4 M ACh was significantly decreased to 10.15 ± 3.00% in CYP-injected rats compared to 27.59 ± 6.64% in control (Fig. 4a). On the other hand, the potentiatory effects of both alprostadil and iloprost on EFS-induced contraction of isolated detrusor muscles of CYP-injected rats were not significantly changed compared to control (Fig. 4b).

A Paraffin cross-section photomicrograph of control rat urinary bladder showing low degree of inflammation, hemorrhage and edema, absence of ulceration, and normal urothelial architecture. H & E. approx. ×10, scale bar = 200 μm (panels a and b); approx. ×40, scale bar = 50 μm (panel c). B Paraffin cross-section photomicrograph of CYP-injected rat urinary bladder showing (filled triangle) high degree of inflammation and hemorrhage, (filled square) mild ulceration, (empty triangle) severe edema, and (downward arrow) proliferated urothelium. H & E. approx. ×10, scale bar = 200 μm (panels a and b); approx. ×40, scale bar = 50 μm (panel c)

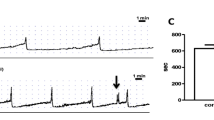
Representative tracings showing the effect of cyclophosphamide (CYP, 300 mg/kg) injection on ACh (panel a) or electric field stimulation (EFS, panel b)-induced contraction of isolated rat detrusor muscles

a Left: effect of alprostadil (10−5 M) and iloprost (10−5 M) on ACh-induced contraction of isolated detrusor muscles of cyclophosphamide (CYP)-injected rats. Right: comparison of potentiation induced by alprostadil or iloprost on ACh (10−4 M)-induced contraction in control and CYP-induced cystitis. Responses are expressed as mean ± SEM.* denotes significant difference from control (p < 0.05). Values between parentheses indicate the number of animals. b Left: effect of alprostadil (10−5 M) and iloprost (10−5 M) on EFS-induced contraction of isolated detrusor muscles of cyclophosphamide (CYP)-injected rats. Right: comparison of potentiation induced by alprostadil or iloprost on EFS (4 Hz)-induced contraction in control and CYP-induced cystitis. Responses are expressed as mean ± SEM.* denotes significant difference from control (p < 0.05). Values between parentheses indicate the number of animals
Effect of SC51322 (selective EP1 antagonist), RO1138452 (selective IP antagonist), and ONO-AE3-237 (selective DP1 antagonist) on ACh and EFS-induced contraction
Both SC51322 and RO1138452 (10−5 M, Orie and Clapp 2011; Yan et al. 2013) produced a significant rightward shift of both ACh concentration and frequency response curves. Effect of SC51322 was compared to its vehicle (0.95% (v/v) ethanol), it significantly decreased the maximum attained contraction of ACh (at 10−3 M) from 1.25 ± 0.09 g (as control) to 0.53 ± 0.10 g, while RO1138452 decreased the maximum attained ACh contraction from 2.11 ± 0.18 to 1.53 ± 0.15 g. The effect of RO1138452 was more prominent at low ACh concentration (10−6 M) while at high ACh concentration (10−3 M), the inhibitory effect of SC51322 became more pronounced (Supplement 2). Same patterns of effects were observed in case of EFS (Supplement 3). In CYP-induced cystitis, and in a similar pattern to normal rats (i.e., rats injected with saline rather than CYP), RO1138452 was more effective than SC51322 at low ACh concentration (10−6 M) as it caused 66.53 ± 5.81% inhibition of ACh-induced contraction compared to 38.33 ± 13.57% in case of SC51322. While at high ACh concentration (10−3 M), the inhibitory effect of SC51322 became more pronounced with a percentage inhibition of 52.93 ± 4.75% compared to 9.82 ± 2.57% for RO1138452. Compared to control normal rats, the inhibitory effect of RO1138452 on ACh-induced contraction was significantly decreased in CYP-injected rats, while the effect of SC51322 was not significantly changed. In case of EFS; the inhibitory effects of both antagonists in CYP-injected rats were almost similar at low and intermediate frequencies (1 and 4 Hz). However, at high frequency (16 Hz), the inhibitory effect of SC51322 was more pronounced with a percentage inhibition of 68.68 ± 4.32% compared to 48.55 ± 3.71% in case of RO1138452 (Fig. 5).

Comparative effect of SC53122 (10−5 M, panel a), RO1138452 (10−5 M, panel b), and ONO-AE3-237 (10−6 M, panel c) on ACh (10−4 M, left panel) and electric stimulation (EFS, 4 Hz, right panel)-induced contraction of isolated detrusor muscle in normal and cyclophosphamide (CYP)-induced cystitis-injected rats. * denotes significant difference from control (p < 0.05). Values between parentheses indicate the number of animals
On the other hand, ONO-AE3-237 (10−6 M, Guan et al. 2015) produced no significant effect on ACh-induced contraction in normal rats; however, in CYP-injected rats, it produced a significant potentiation of ACh-induced contraction mainly at higher ACh concentrations (10−4 and 10−3 M). At 10−4 M ACh, ONO-AE3-237 potentiated ACh-induced contraction by 23.51 ± 6.20% in CYP-injected rats, while in control, it inhibited the contraction by 3.67 ± 4.11%. At frequency of 4 Hz, ONO-AE3-237 also potentiated EFS-induced contraction by 33.51 ± 9.33% in CYP-injected rats compared to 2.21 ± 6.03% in control (Fig. 5). It is to be mentioned that ONO-AE3-237 is dissolved in 9.5% v/v ethanol which gave a final bath concentration of 0.095% v/v and showed no significant effect on ACh as well as EFS-induced contraction.
Effect of indomethacin, ketoprofen, diclofenac, and DFU on ACh and EFS-induced contraction
A concentration response curve of ACh (10−9–10−3 M) was conducted on isolated rat detrusor muscle in absence and presence of indomethacin, ketoprofen, diclofenac (20, 50, and 100 μM), or DFU (selective COX-2 inhibitor; 10−7–10−5 M). In normal rats, a significant inhibition of ACh-induced contraction and a downward shift of the concentration response curve were observed in presence of indomethacin, ketoprofen, and diclofenac at all tested concentrations (20, 50, and 100 μM). As for EFS-induced contraction, a downward shift of the frequency response curve was observed in presence of high concentrations of indomethacin, diclofenac, and ketoprofen (at doses 50 and 100 μM). Only diclofenac succeeded to inhibit EFS-induced contraction at low concentration (20 μM) and at low frequency (Supplement 4). On the other hand, DFU produced no significant effect on neither ACh nor EFS-induced contraction compared to vehicle.
In CYP-injected rats with confirmed cystitis, a similar downward shift of ACh concentration response curve was observed in presence of indomethacin, ketoprofen, and diclofenac but only at high ACh concentrations (10−5 and 10−4 M) with no significant effect on lower doses (Supplement 5). At 10−4 M ACh, the percentage inhibition induced by indomethacin was decreased from 49.79 ± 5.13 to 27.13 ± 5.45% in control and CYP-injected rats, respectively. In case of ketoprofen, the percentage inhibition was decreased from 43.37 ± 4.50 to 14.46 ± 3.74%, and in case of diclofenac, it was decreased from 51.55 ± 3.97 to 22.42 ± 4.46%. In contrast to normal rats, DFU (10−6 M) in case of cystitis significantly potentiated the contractile effect of ACh at high concentrations (10−5–10−3 M) compared to DMSO (vehicle). It significantly increased the maximum attained contraction from 1.04 ± 0.11 to 1.39 ± 0.20 g with a percentage potentiation of 30.64 ± 6.63%.
When comparing the effect of the non-selective COX inhibitors on EFS-induced contraction, they all produced more inhibition in case of cystitis almost at all frequencies tested. At 4 Hz, the percentages inhibition induced by indomethacin, ketoprofen, and diclofenac was increased from 25.32 ± 6.42% in normal rats to 63.45 ± 8.09% in CYP-injected rats, from 15.32 ± 5.72 to 44.60 ± 4.26% and from 29.79 ± 2.95 to 52.88 ± 6.84%, respectively. In CYP-injected rats, indomethacin produced the highest inhibition of EFS-induced contraction followed by diclofenac then ketoprofen. On the other hand, DFU produced no significant effect on EFS-induced contraction in CYP-injected rats (Fig. 6).

Comparative effect induced by indomethacin (50 μM, panel a), ketoprofen (50 μM, panel b), diclofenac (50 μM, panel c), or DFU (10−6 M, panel d) on ACh-induced contraction (left) and on electric field stimulation (EFS)-induced contraction (right) of isolated rat detrusor muscles in control and cyclophosphamide (CYP)-induced cystitis. Responses are expressed as mean ± SEM.* denotes significant difference from control (p < 0.05). Values between parentheses indicate the number of animals
Effect of indomethacin on ACh-induced contraction in presence of atropine
In presence of atropine (1 nM), indomethacin (50 μM) was still able to produce a significant inhibition of ACh-induced contraction; however, its inhibitory effect was significantly lowered. At 10−6 M of ACh, the percentage inhibition induced by indomethacin in presence of atropine was decreased to 39.31 ± 6.22% compared to 78.55 ± 5.28% in absence of atropine.
In case of cystitis, the percentage inhibition induced by atropine was decreased compared to control. For example, at 10−4 M ACh, atropine inhibited ACh-induced contraction in CYP-injected rats by 16.69 ± 3.30% compared to 34.94 ± 5.11% in control. In contrast, at 10−3 M ACh, atropine unexpectedly potentiated the maximum attained contraction from 1.34 ± 0.1 to 1.53 ± 0.12 g in CYP-injected rats with a percentage potentiation of 19.14 ± 2.05% while this effect was absent in control. In presence of atropine (1 nM), indomethacin (50 μM) was still able to cause a significant inhibition of ACh-induced contraction at (10−5 and 10−4 M ACh) and, in contrast to control, the inhibitory effect induced by indomethacin was not significantly decreased in presence of atropine in CYP-injected rats (Fig. 7).

a Effect of atropine (1 nM) and indomethacin (50 μM) in presence of atropine (1 nM) on ACh-induced contraction of isolated rat detrusor muscles in normal (left) and cyclophosphamide (CYP, right)-induced cystitis. * denotes significant difference from control. # denotes significant difference from atropine (p < 0.05). Values between parentheses indicate the number of animals. b Percentage inhibition induced by indomethacin (50 μM) on ACh-induced contraction of isolated rat detrusor muscles in absence and presence of atropine (1 nM) in normal (left) and cyclophosphamide (CYP, right)-induced cystitis. Responses are expressed as mean ± SEM. + denotes significant difference from indomethacin group (p < 0.05). Values between parentheses indicate the number of animals
Discussion
Cyclophosphamide is an antineoplastic agent used for the treatment of different types of tumors and as immunosuppressive agent (Aschan et al. 1999). One of its major side effects is hemorrhagic cystitis which involves the release of different inflammatory mediators and production of reactive oxygen species and other mediators (Cox 1979).
In the current study, CYP-induced hemorrhagic cystitis was confirmed by histopathological examination which is consistent with that previously reported by Oter et al. (2004). Both ACh and EFS-induced contractions of detrusor muscles were significantly diminished in rats with cystitis which is consistent with the reported lowering of carbachol and potassium chloride-induced contraction of rat detrusor muscles in CYP-injected rats (Giglio et al. 2005). Moreover, responses to methacholine, ATP, and adenosine are reported to be diminished in rats with hemorrhagic cystitis (Aronsson et al. 2014). Detrusor muscles hypoactivity may be attributed to the reported downregulation of purinergic receptors (A1) which are important for detrusor muscles contraction (Vesela et al. 2011), the increased expression of inducible nitric oxide synthase (iNOS) and endothelial NOS (eNOS) which in turn results in increasing the synthesis of nitric oxide (NO) (Oter et al. 2004; Giglio et al. 2005) and as a result, inhibiting ACh-induced release of PGE2 in urothelium (Nile and Gillespie 2012). Hypoactivity may also be attributed to the increased expression of M5 receptors in detrusor smooth muscles which in turn may stimulate NO production (Giglio et al. 2005).
Prostaglandins are suggested to play an important role as key mediators in hemorrhagic cystitis; upregulation of COX-2 expression, and prostaglandin (PGE2 and PGD2) production were demonstrated in the urinary bladder in acute and chronic CYP-induced cystitis. In addition, bladder function was improved with administration of a specific COX-2 inhibitor, DFU (Hu et al. 2003). In the current study, alprostadil potentiated both ACh and EFS-induced contraction in cystitis as well as in control bladder and its potentiatory effect in cystitis was not significantly different than control. The results obtained from normal rats are consistent with those previously reported by Palea et al. (1998) who showed that PGE1 induces contraction of human detrusor muscles. Boie et al. (1997) have reported that alprostadil’s effect involves the activation of all PGs receptors of the E-series (EP4 > EP3 > EP2 > EP1). It may also have some affinity to IP receptors. However, Coleman et al. (1994) stated that, in detrusor muscles, EP1 and EP3 receptors are the ones involved in contraction; therefore, we can suggest that they may be the major contributors to the observed potentiation of ACh and EFS-induced contraction induced by alprostadil. It can be concluded that alprostadil may have a potential clinical importance in case of bladder hypoactivity.
Unexpectedly, iloprost—PGI2 analogue—also potentiated detrusor muscle contractions, but to a lesser extent than alprostadil. IP receptors in smooth muscles are expected to mediate relaxation (Narumiya et al. 1999); however, iloprost is reported to activate EP1 first followed by IP receptors (Whittle et al. 2012) which may explain the observed effect. Boie et al. (1997) reported that iloprost acts as a partial agonist at EP1 and a full agonist at EP3 receptors in rats. Both receptors are Gq-coupled, they activate phospholipase C and elevate the intracellular calcium level. In CYP-induced hemorrhagic cystitis, the potentiatory effect of iloprost, but not that of alprostadil, on ACh-induced contraction was significantly lowered compared to normal rats. This could be attributed to the fact that inflammation triggers the production of more PGI2 which succeed—through IP receptor activation—to counteract the weak potentiatory effect induced by iloprost rather than the strong potentiatory effect induced by activating EP1 and EP3 receptors by alprostadil. It is to be mentioned that the reduction in iloprost action was only observed in case of ACh but not when contraction was induced by EFS: This may be attributed to the reduction of M2-mediated release of the contractile PGs in urothelium, the major part affected by cystitis (Giglio et al. 2005; Nile and Gillespie 2012). EFS-induced contraction, on the other hand, may induce the release of not only ACh but also other contractile mediators as co-transmitters such as ATP (Andersson and Arner 2004) which may mask the alteration in PG actions. Further measurements of the tested PGs levels and PGs receptors expression in detrusor muscles in normal and inflammatory condition are needed to confirm this hypothesis.
Both EP1 antagonist (SC51322) and IP antagonist (RO1138452) caused a downward shift of both ACh concentration and frequency response curves. IP antagonist was the most effective in decreasing the contractile effect of ACh and its effect on ACh-induced contraction was more pronounced than on EFS-induced contraction. In fact, the effect of IP antagonist can be attributed to muscarinic receptor blockade as previously shown by Bley et al. (2006). The current study also showed that the inhibitory effect of EP1 receptor antagonist on detrusor muscles contractility was not significantly changed in cystitis compared to control. However, the inhibitory effect of IP antagonist on ACh but not on EFS-induced contraction was significantly reduced in cystitis. This may be attributed to the reduction of M2-mediated release of the contractile PGs in urothelium as the major part affected by cystitis as a result of increased NO synthesis (Giglio et al. 2005; Nile and Gillespie 2012). It is to be mentioned that ethanol (the solvent used for SC51322) inhibited both cholinergic and EFS-induced contraction of normal detrusor muscles, possibly via inhibiting calcium influx through calcium channels and decreasing the intracellular calcium level (Ohmura et al. 1997). This effect—on the other hand—was absent in case of cystitis. This can be explained by the fact that eNOS, iNOS, and NO expression is reported to be increased in case of cystitis (Oter et al. 2004), which may in turn inhibit calcium influx and mask ethanol’s action.
The current study also showed that, in normal rats, DP1 receptor antagonist (ONO-AE3-237) caused no significant effect on neither cholinergic nor neuronally mediated contraction of rat detrusor muscles. This suggests that PGD2 may not be released under normal physiological conditions. However, Guan et al. (2015) have reported that exogenous PGD2 has a relaxant effect on detrusor smooth muscles via activation of DP1 receptors. In contrast, in CYP-induced cystitis, DP1 receptor antagonist showed significant potentiation of both cholinergic and neuronally mediated contraction. This may be attributed to inducing the production of PGD2 in hemorrhagic cystitis or upregulation of the DP1 receptors which may favor the effect of the inhibitory PGs resulting in detrusor muscles hypoactivity. More studies are needed with measurement of PG synthesis and receptor expression to exactly explain this observation.
Little and controversial data is reported in literature about the potential negative role of COX inhibitors on normal bladder physiology in humans and experimental animals. In the present work, all the tested non-selective COX inhibitors (indomethacin, ketoprofen, and diclofenac) lowered the detrusor muscle activity. Similarly, indomethacin and other COX inhibitors were reported to inhibit the cholinergic-mediated contraction of guinea pig detrusor muscles (Rahnama'i et al. 2013) and were found to be effective in reducing bladder overactivity in experimental animals (Dobrek et al. 2014). However, Badawi et al. (2008) reported that, in porcine detrusor muscles, indomethacin did not affect cholinergic activity at low concentration. Such discrepancy may be due to species variation. In the current study, diclofenac—which is more selective COX-2 inhibitor—was the most effective in lowering the contractile effect of ACh on rat detrusor muscles followed by indomethacin then ketoprofen. Previous reports have revealed that diclofenac may act by other COX-independent mechanisms (Kothari et al. 1987; Maggi et al. 1991; Papworth et al. 1997; Palea et al. 1998; Ortiz et al. 2003; Selg et al. 2007).
In attempt to explain the inhibitory effect of COX inhibitors on detrusor, the involvement of muscarinic activity in the effect of indomethacin was investigated. The current study showed that, in presence of atropine, the inhibitory effect of indomethacin on ACh-induced contraction was decreased, suggesting that indomethacin—as a part of its action—may interfere with muscarinergic signaling in urothelium. It may inhibit the synthesis of PGE2—the major contractile PG in bladder—released from urothelium in response to ACh-mediated activation of M2 receptors as reported by Nile and Gillespie (2012), resulting in inhibiting ACh-induced contraction. So, upon blocking muscarinic receptors, part of the inhibitory effect of indomethacin may be lost. Other mechanisms that may contribute to the inhibitory effect of indomethacin may include the following: first, inhibition of the synthesis of PGE2 in urothelium may affect the release of ATP which contributes to detrusor muscles contractility (Nile et al. 2010). Secondly, Nile and Gillespie (2012) reported that PGE2 may induce the release of ACh. Therefore, it can be suggested that inhibition of PGE2 synthesis may be associated with inhibition of ACh release. Finally, COX inhibitors may increase the activity of acetylcholinesterase due to inhibition of PGF2α synthesis (Borda et al. 1982). On ACh in CYP-induced cystitis, the effect of the non-selective COX inhibitors was significantly reduced compared to normal rats. Moreover, the inhibitory effect of indomethacin did not significantly change in case of muscarinic blockade. These findings may be attributed to the reduction of M2-mediated release of PGE2 in urothelium which got seriously affected in cystitis (Giglio et al. 2005; Nile and Gillespie 2012), meaning that part of the inhibitory effect of atropine as well as indomethacin may be lost. Figure 8 summarizes the proposed potential mechanisms underlying hypoactivity of detrusor muscle induced by hemorrhagic cystitis in respect to COX, PGs, and their respective receptors, in comparison to normal bladder.

Schematic representation of the proposed potential mechanisms involved in the pathophysiology of cyclophosphamide-induced hemorrhagic cystitis is rats and the possible effect of prostanoids modulators on the proposed mechanisms. COX = cyclooxygenase enzyme, DP1 = prostaglandin D2 receptor, EP = prostaglandin E receptor, M = muscarinic receptor, IP = prostacyclin receptor, NO = nitric oxide. (+) = mediates contraction, (−) = mediates relaxation of rat detrusor muscle, and (x)= blockade and (x)= blockade
Concerning the neuronally mediated detrusor muscles contractility, the current study showed that the non-selective COX inhibitors inhibited EFS-induced contraction in cystitis; however, the degree of inhibition was significantly increased in cystitis compared to control, and indomethacin showed the highest degree of inhibition, indicating that this effect may be proportional to the increased selectivity on COX-1. This effect may be attributed to the increased expression of PGE2 in rat bladder in CYP-induced cystitis compared to control as reported by Linares-Fernandez and Alfieri (2007), so preventing its synthesis may cause more inhibition of EFS-induced contraction in cystitis. This suggestion is further supported by the previous studies which revealed that neuronal stimulation of urinary bladder induces the release of different contractile mediators including ACh, ATP, and contractile PGs (Andersson and Arner 2004). In cystitis, the effect of ACh is decreased as a result of increased M5 receptors and NO expression (Giglio et al. 2005), and the effect of ATP may be also decreased due to the downregulation of purinergic receptors (Vesela et al. 2011). Therefore, the neuronal release of contractile PGs may have the upper hand in the contractile effect of EFS in cystitis.
On the other hand, regarding the effect of the selective COX-2 inhibitor, DFU; although it did not affect detrusor muscles contractility in normal bladder, it significantly potentiated ACh-induced contraction of detrusor muscles in cystitis, but with no significant effect on the neuronally mediated contraction. This is in accordance with Klinger et al. (2007) who reported that COX-2 isoenzyme is not expressed in normal bladder, while in cystitis, COX-2 becomes highly expressed and may participate to the net deterioration appearing in detrusor muscles function, so COX-2 inhibitors may have a beneficial effect on the muscle activity and could be useful for the management of such condition as reported by Lecci et al. (2000). It could be also suggested that, in cystitis, COX-2 isoenzyme may interact with the cholinergic activity of detrusor muscles to a greater extent than neuronally mediated pathways which involves other contractile mediators. In contrast, non-selective COX inhibitors may cause further deterioration of detrusor muscles contractility in cystitis. This contradicts the report of Macedo et al. (2008) who suggested that COX inhibitors could be added to 2-mercaptoethanesulphonic acid (MESNA) in order to reduce the degree of inflammation accompanied with hemorrhagic cystitis and improve bladder function.
Lack of measurements for the expression of the studied PGs receptors (EP1, IP, and DP1) in detrusor muscle of normal as well as CYP-injected rats, as well as the measurement of the effective level of the respective PGs produced in each condition remain a limitation of the current study and needs future investigation. Equally, studying the differential effect of the tested selective and non-selective COX inhibitors on intravesical pressure by in vivo cystometry in hemorrhagic cystitis will be needed to confirm the current results.
Conclusions
EP1 receptors seem to play an important role in rat bladder contractility. Alprostadil seems promising at increasing detrusor muscle activity in normal conditions and to a lesser extend in cystitis. Cyclophosphamide-induced cystitis was not only associated with histopathological damage of urinary bladder, but also with hypoactivity of detrusor muscle. DP1 receptors may be involved in this pathogenicity, so blocking these receptors may have a potential therapeutic benefit that worth further studies.
Non-selective COX inhibitors seem to be unsuitable as anti-inflammatory agent in patients with cystitis, as they may reduce detrusor muscle contractility, while the selective COX-2 inhibitors may be a better option. In normal detrusor muscles, the inhibitory effect of indomethacin on contractility was less than diclofenac and greater than ketoprofen. In contrast, in cystitis, indomethacin was shown to be the worst, reflecting the potential harmful effect induced by inhibiting COX-1 activity in the muscle. Biochemical measurement of PGs production, as well as COX and PG receptors expression remains essential to confirm the results of the current study.
References
Anderson KE (1993) Pharmacology of lower urinary tract smooth muscles and penile erectile tissues. Pharmacol Rev 45(3):253–308
Andersson KE (2016) Potential future pharmacological treatment of bladder dysfunction. Basic Clin Pharmacol Toxicol 119(Suppl 3):75–85
Andersson KE, Arner A (2004) Urinary bladder contraction and relaxation: physiology and pathophysiology. Physiol Rev 84(3):935–986
Aronsson P, Vesela R, Johnsson M (2014) Inhibition of nitric oxide synthase prevents muscarinic and purinergic functional changes and development of cyclophosphamide-induced cystitis in the rat. Biomed Res Int 2014:359179
Aschan J, Carlens S, Hagglund H, Klaesson S, Mattsson J, Remberger M (1999) Improved survival after bone marrow transplantation for early leukemia using busulfan-cyclophosphamide and individualized prophylaxis against graft-versus-host disease: a long-term follow-up. Clin Transpl 13(6):512–519
Badawi JK, Seja T, Bross S (2008) In vitro effects of the cyclooxygenase inhibitor indomethacin and of the phospholipase-C inhibitor U-73122 on carbachol-induced contractions of porcine detrusor muscle. Fundam Clin Pharmacol 22(6):667–672
Bley KR, Bhattacharya A, Daniels DV, Gever J, Jahangir A, O'Yang C, Smith S, Srinivasan D, Ford AP, Jett MF (2006) RO1138452 and RO3244794: characterization of structurally distinct, potent and selective IP (prostacyclin) receptor antagonists. Br J Pharmacol 147(3):335–345
Boie Y, Stocco R, Sawyer N, Slipetz DM, Ungrin MD, Neuschafer-Rube F, Puschel GP, Metters KM, Abramovitz M (1997) Molecular cloning and characterization of the four rat prostaglandin E2 prostanoid receptor subtypes. Eur J Pharmacol 340(2–3):227–241
Borda E, Contreras-Ortiz N, Gutnisky R, Gimeno MF (1982) In vitro effect of acetylcholine and bethanechol on the contractions of the human detrusor muscle. Influence of prostaglandins. Arch Int Pharmacodyn Ther 259(1):31–39
Brideau C, Van Staden C, Chan CC (2001) In vitro effects of cyclooxygenase inhibitors in whole blood of horses, dogs, and cats. Am J Vet Res 62(11):1755–1760
Cardozo LD, Stanton SL, Robinson H, Hole D (1980) Evaluation of flurbiprofen in detrusor instability. Br Med J 280(6210):281–282
Coleman RA, Smith WL, Narumiya S (1994) International Union of Pharmacology classification of prostanoid receptors: properties, distribution, and structure of the receptors and their subtypes. Pharmacol Rev 46(2):205–229
Cox PJ (1979) Cyclophosphamide cystitis—identification of acrolein as the causative agent. Biochem Pharmacol 28(13):2045–2049
Ding HL, Ryder JW, Stull JT, Kamm KE (2009) Signaling processes for initiating smooth muscle contraction upon neural stimulation. J Biol Chem 284(23):15541–15548
Dobrek L, Baranowska A, Skowron B, Thor PJ (2014) The influence of piroxicam, a non-selective cyclooxygenase inhibitor, on autonomic nervous system activity in experimental cyclophosphamide-induced hemorrhagic cystitis and bladder outlet obstruction in rats. Acta Pol Pharm 71(3):497–507
Giglio D, Ryberg AT, To K, Delbro DS, Tobin G (2005) Altered muscarinic receptor subtype expression and functional responses in cyclophosphamide induced cystitis in rats. Auton Neurosci 122(1–2):9–20
Gray KJ, Engelmann UH, Johnson EH, Fishman IJ (1986) Evaluation of misoprostol cytoprotection of the bladder with cyclophosphamide (Cytoxan) therapy. J Urol 136(2):497–500
Guan NN, Svennersten K, de Verdier PJ, Wiklund NP, Gustafsson LE (2015) Receptors involved in the modulation of guinea pig urinary bladder motility by prostaglandin D2. Br J Pharmacol 172(16):4024–4037
Hegde SS (2006) Muscarinic receptors in the bladder: from basic research to therapeutics. Br J Pharmacol 147(Suppl 2):S80–S87
Hu VY, Malley S, Dattilio A, Folsom JB, Zvara P, Vizzard MA (2003) COX-2 and prostanoid expression in micturition pathways after cyclophosphamide-induced cystitis in the rat. Am J Physiol Regul Integr Comp Physiol 284(2):R574–R585
Irwin DE, Kopp ZS, Agatep B, Milsom I, Abrams P (2011) Worldwide prevalence estimates of lower urinary tract symptoms, overactive bladder, urinary incontinence and bladder outlet obstruction. BJU Int 108(7):1132–1138
Jeremy JY, Mikhailidis DP, Dandona P (1984) The rat urinary bladder produces prostacyclin as well as other prostaglandins. Prostaglandins Leukot Med 16(2):235–248
Jeremy JY, Tsang V, Mikhailidis DP, Rogers H, Morgan RJ, Dandona P (1987) Eicosanoid synthesis by human urinary bladder mucosa: pathological implications. Br J Urol 59(1):36–39
Klinger MB, Dattilio A, Vizzard MA (2007) Expression of cyclooxygenase-2 in urinary bladder in rats with cyclophosphamide-induced cystitis. Am J Physiol Regul Integr Comp Physiol 293(2):R677–R685
Kothari HV, Lee WH, Ku EC (1987) An alternate mechanism for regulation of leukotriene production in leukocytes: studies with an anti-inflammatory drug, sodium diclofenac. Biochim Biophys Acta 921(3):502–511
Lecci A, Birder LA, Meini S, Catalioto RM, Tramontana M, Giuliani S, Criscuoli M, Maggi CA (2000) Pharmacological evaluation of the role of cyclooxygenase isoenzymes on the micturition reflex following experimental cystitis in rats. Br J Pharmacol 130(2):331–338
Lee T, Hedlund P, Newgreen D, Andersson KE (2007) Urodynamic effects of a novel EP(1) receptor antagonist in normal rats and rats with bladder outlet obstruction. J Urol 177(4):1562–1567
Leslie CA, Pavlakis AJ, Wheeler JS Jr, Siroky MB, Krane RJ (1984) Release of arachidonate cascade products by the rabbit bladder; neurophysiological significance? J Urol 132(2):376–379
Linares-Fernandez BE, Alfieri AB (2007) Cyclophosphamide induced cystitis: role of nitric oxide synthase, cyclooxygenase-1 and 2, and NK(1) receptors. J Urol 177(4):1531–1536
Luheshi GN, Zar MA (1991) The effect of streptozotocin-induced diabetes on cholinergic motor transmission in the rat urinary bladder. Br J Pharmacol 103(3):1657–1662
Macedo FY, Baltazar F, Almeida PR, Tavora F, Ferreira FV, Schmitt FC, Brito GA, Ribeiro RA (2008) Cyclooxygenase-2 expression on ifosfamide-induced hemorrhagic cystitis in rats. J Cancer Res Clin Oncol 134(1):19–27
Maggi CA (1992) Prostanoids as local modulators of reflex micturition. Pharmacol Res 25(1):13–20
Maggi CA, Patacchini R, Santicioli P, Giuliani S (1991) Tachykinin antagonists and capsaicin-induced contraction of the rat isolated urinary bladder: evidence for tachykinin-mediated cotransmission. Br J Pharmacol 103(2):1535–1541
Narumiya S, Sugimoto Y, Ushikubi F (1999) Prostanoid receptors: structures, properties, and functions. Physiol Rev 79(4):1193–1226
Nile CJ, Gillespie JI (2012) Interactions between cholinergic and prostaglandin signaling elements in the urothelium: role for muscarinic type 2 receptors. Urology 79(1):240.e17–240.e23
Nile CJ, de Vente J, Gillespie JI (2010) Stretch independent regulation of prostaglandin E(2) production within the isolated guinea-pig lamina propria. BJU Int 105(4):540–548
Ohmura M, Kondo A, Saito M (1997) Effects of ethanol on responses of isolated rabbit urinary bladder and urethra. Int J Urol 4(3):295–299
Orie NN, Clapp LH (2011) Role of prostanoid IP and EP receptors in mediating vasorelaxant responses to PGI2 analogues in rat tail artery: evidence for Gi/o modulation via EP3 receptors. Eur J Pharmacol 654(3):258–265
Ortiz MI, Granados-Soto V, Castaneda-Hernandez G (2003) The NO-cGMP-K+ channel pathway participates in the antinociceptive effect of diclofenac, but not of indomethacin. Pharmacol Biochem Behav 76(1):187–195
Oter S, Korkmaz A, Oztas E, Yildirim I, Topal T, Bilgic H (2004) Inducible nitric oxide synthase inhibition in cyclophosphamide induced hemorrhagic cystitis in rats. Urol Res 32(3):185–189
Palea S, Toson G, Pietra C, Trist DG, Artibani W, Romano O, Corsi M (1998) Pharmacological characterization of thromboxane and prostanoid receptors in human isolated urinary bladder. Br J Pharmacol 124(5):865–872
Papworth J, Colville-Nash P, Alam C, Seed M, Willoughby D (1997) The depletion of substance P by diclofenac in the mouse. Eur J Pharmacol 325(2–3):R1–r2
Rahnama'i MS, van Kerrebroeck PE, de Wachter SG, van Koeveringe GA (2012) The role of prostanoids in urinary bladder physiology. Nat Rev Urol 9(5):283–290
Rahnama'i MS, van Koeveringe GA, van Kerrebroeck PE, de Wachter SG (2013) The effect of indomethacin on the muscarinic induced contractions in the isolated normal guinea pig urinary bladder. BMC Urol 13:8
Rao P, Knaus EE (2008) Evolution of nonsteroidal anti-inflammatory drugs (NSAIDs): cyclooxygenase (COX) inhibition and beyond. J Pharm Pharm Sci 11(2):81s–110s
Selg E, Buccellati C, Andersson M, Rovati GE, Ezinga M, Sala A, Larsson AK, Ambrosio M, Lastbom L, Capra V, Dahlen B, Ryrfeldt A, Folco GC, Dahlen SE (2007) Antagonism of thromboxane receptors by diclofenac and lumiracoxib. Br J Pharmacol 152(8):1185–1195
Simmons DL, Botting RM, Hla T (2004) Cyclooxygenase isozymes: the biology of prostaglandin synthesis and inhibition. Pharmacol Rev 56(3):387–437
Takagi-Matsumoto H, Ng B, Tsukimi Y, Tajimi M (2004) Effects of NSAIDs on bladder function in normal and cystitis rats: a comparison study of aspirin, indomethacin, and ketoprofen. J Pharmacol Sci 95(4):458–465
Tanaka I, Nagase K, Tanase K, Aoki Y, Akino H, Yokoyama O (2011) Modulation of stretch evoked adenosine triphosphate release from bladder epithelium by prostaglandin E(2). J Urol 185(1):341–346
Uchiyama T, Chess-Williams R (2004) Muscarinic receptor subtypes of the bladder and gastrointestinal tract. J Smooth Muscle Res 40(6):237–247
Vesela R, Aronsson P, Tobin G (2011) Functional and morphological examinations of P1A1 purinoceptors in the normal and inflamed urinary bladder of the rat. Auton Neurosci 159(1–2):26–31
Wada N, Matsumoto S, Kita M, Watanabe M, Hashizume K, Kakizaki H (2013) Effect of intrathecal administration of E-series prostaglandin 1 receptor antagonist in a cyclophosphamide-induced cystitis rat model. Int J Urol 20(2):235–240
Whittle BJ, Silverstein AM, Mottola DM, Clapp LH (2012) Binding and activity of the prostacyclin receptor (IP) agonists, treprostinil and iloprost, at human prostanoid receptors: treprostinil is a potent DP1 and EP2 agonist. Biochem Pharmacol 84(1):68–75
Yan G, Wang Q, Shi H, Han Y, Ma G, Tang C, Gu Y (2013) Regulation of rat intrapulmonary arterial tone by arachidonic acid and prostaglandin E2 during hypoxia. PLoS One 8(8):e73839
Acknowledgements
We thank Prof. Mona Yehia, Ph.D., Professor and Head of the Department of Histochemistry and Cell Biology, Medical Health Institute, Alexandria University, Egypt, for her help in conducting histological studies. We also thank Dr. Takayuki Maruyama for providing the ONO compounds as a gift to Dr. Xavier Norel.
Funding
Part of this work was supported by Institue Francais en Egypte, French Agency for the Development of Higher Education and Research, and the Academy of Scientific Research and Technology in Egypt under the frame of “Imhotep Project N°: 31681XL” and “STDF/IFE Project N° 30630”.
Author information
Authors and Affiliations
Contributions
AS and TD conceived the idea. AS, XN, and TD designed research. WB conducted experiments. AS and WB analyzed and interpreted data. AS, WB, and TD revised data and wrote the manuscript. AS and XN were Egyptian and French PI of the project, respectively. All authors read and approved the manuscript.
Corresponding author
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(DOCX 3.62 MB)
Rights and permissions
About this article

Cite this article
Bassiouni, W., Daabees, T., Norel, X. et al. Hypoactivity of rat detrusor muscle in a model of cystitis: exacerbation by non-selective COX inhibitors and amelioration by a selective DP1 receptor antagonist. Naunyn-Schmiedeberg's Arch Pharmacol 392, 437–450 (2019). https://doi.org/10.1007/s00210-018-01599-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00210-018-01599-7