
Abstract
Nitric oxide (NO) has biphasic effects on regulating osteoblast survival and death. This study was aimed to evaluate the effects of NO pretreatment on hydrogen peroxide (HP)-induced insults of rat osteoblasts and the possible mechanisms. Exposure of osteoblasts prepared from rat calvarias to HP significantly increased intracellular reactive oxygen species levels, decreased alkaline phosphatase activity and cell survival, and ultimately induced cell apoptosis. However, NO pretreatment lowered HP-induced oxidative stress and apoptotic insults. In parallel, HP increased Bax levels and its translocation from the cytoplasm to mitochondria. NO pretreatment caused significant attenuations in HP-induced modulations in Bax synthesis and translocation. In contrast, pretreatment with NO enhanced levels and translocation of antiapoptotic Bcl-XL protein in rat osteoblasts. RNA analyses further revealed that HP inhibited Bcl-XL mRNA expression without affecting Bax mRNA levels. In comparison, NO induced Bcl-XL mRNA production and alleviated HP-caused inhibition of this mRNA expression. As to the mechanism, HP suppressed RNA and protein levels of transcription factor GATA-5 in rat osteoblasts. Pretreatment with NO induced GATA-5 mRNA and protein expressions and simultaneously attenuated HP-induced inhibition of this gene’s expression. Consequently, GATA-5 knockdown using RNA interference inhibited Bcl-XL mRNA expression and concurrently lowered NO’s protection against HP-induced apoptotic insults. Therefore, this study showed that NO can protect rat osteoblasts from HP-induced apoptotic insults. The protective mechanisms are mediated by GATA-5-mediated transcriptional induction of Bcl-X L gene, and translocational modulation of Bcl-XL and Bax proteins.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Nitric oxide (NO), synthesized from l-arginine metabolism by NO synthases, contributes to the regulation of tissue/cell activities, including vasodilatation, neurotransmission, immunoresponses, and tumorigenesis (Moncada et al. 1991; Chen et al. 2005a, b; Cochain et al. 2013). In bone-derived cells, both constitutive and inducible NO synthases were detected, and NO was implicated as an important signaling molecule that can regulate the physiology and pathophysiology of bone tissues (Evans and Ralston 1996; Jamal et al. 2013). A previous study showed that NO could decrease bone mineral density in inflammation-induced osteoporosis (Armour et al. 1999). In cyclosporine A-treated rats, administration of l-arginine increased NO levels and simultaneously prevented bone loss and bone collagen breakdown (Fiore et al. 2000). Recent cohort studies also reported that treatment of postmenopausal women with organic nitrates, which can act as NO donors, was associated with increased bone mineral density and with lower risks of hip fracture (Jamal et al. 2013). Thus, NO exerts biphasic effects on bone activities (Ralston 1997; Tai et al. 2007). High levels of NO have cytotoxic effects which can induce bone diseases, whereas NO at low concentrations can potentiate bone metabolism.
Osteoblasts, which differentiate from stromal stem cells, play a central role in mediating bone formation (Takeda and Karsenty 2001; Sims and Vrahnas 2014). A large array of systemic and local factors participate in regulating osteoblast activities (Takeda and Karsenty 2001; Giustina et al. 2008). One such factor, NO, appears to form an important facet of the many complicated communication pathways controlling osteoblast physiology and bone remodeling (Collin-Osdoby et al. 1995). Our previous studies showed that NO from exogenous decomposition of sodium nitroprusside (SNP) or endogenous responses to stimulation by interferon-γ and lipopolysaccharide induced osteoblast death via an apoptotic mechanism (Chen et al. 2002, 2005a, b). Mitochondria were shown to play an important role in NO-induced osteoblast apoptosis (Chen et al. 2005a, b; Ho et al. 2005). In addition to its deadly effects, previous studies revealed that low levels of NO can protect a variety of cells from stress-induced apoptotic insults (Das et al. 2005; Simon et al. 2014). In ischemia and reperfusion injury, NO preconditioning was shown to produce protective effects on cardiomyocytes, hepatocytes, and endothelial cells by modulating mitochondrial function (Cottart et al. 1999; Murillo et al. 2011). Our previous studies further showed that low concentrations of NO can protect osteoblasts from high concentrations of NO-induced apoptotic insults (Tai et al. 2007; Chang et al. 2006; Ho et al. 2009). However, a more detailed molecular mechanism of NO’s protection of osteoblasts needs to be explored.
Bax, one of the proapoptotic multidomain Bcl-2 members with homology in the BH1 ~ BH3 domains, functions as an essential gateway to mediate mitochondrion-dependent cell apoptosis (Pagliari et al. 2005; Monaco et al. 2013). When exposed to various apoptotic stimuli, Bax proteins are specifically translocated to the outer membranes of mitochondria from the cytoplasm. After forming oligomers, the Bax protein can insert itself into the outer mitochondrial membrane, permeabilizing the membrane and triggering the release of cytochrome c (Cyt c) (Hsu et al. 1997; Goyal 2001). In contrast, Bcl-XL, another Bcl-2 family member, functions to prevent Bax-induced apoptotic events and is thought to have antiapoptotic effects (Pagliari et al. 2005). In ischemia-/reperfusion-induced heart injury, overexpression of the Bcl-X L gene can suppress Bax translocation, leading to suppression of Cyt c release and cardiac cell apoptosis (Huang et al. 2005). Under apoptotic stimulation, these apoptosis-related gene expressions can be regulated. GATA-DNA-binding proteins (GATAs), a family of transcriptional regulators, contain two zinc fingers with the motif Cys-X2-Cys-X17-Cys-X2-Cys that directly binds to the nucleotide sequence element (A/T)GATA(A/G) (Evans and Felsenfeld 1989; Suzuki et al. 1996). There are 6 members in the GATA family of transcription factors. In general, GATA-1, GATA-2, and GATA-3 are known to regulate critical events in hematopoietic lineages, while GATA-4, GATA-5, and GATA-6 are reported to mainly be expressed in nonhematopoietic tissues, including the heart and gut (Burch 2005). Our previous study demonstrated that GATA-3 can mediate survival signals of osteoblasts (Chen et al. 2010a, b). GATA-5 contributes to the regulation of cell proliferation and differentiation (Capo-Chichi et al. 2005; Peters et al. 2014). GATA-DNA-binding elements are found in the 5′-end promoter region of the Bcl-X L gene (Yu et al. 2005). However, the roles of GATA-5 in osteoblasts are still unknown. In this study, we specifically evaluated the effects of NO pretreatment on oxidative stress-induced osteoblast injuries, especially the roles of GATA-5, Bcl-XL, and Bax in such events.
Materials and methods
Preparation of rat osteoblasts
Rat osteoblasts were prepared from 3-day-old Wistar rat calvaria according to a previously described collagenase digestion method (Chen et al. 2002). All procedures were performed according to National Institutes of Health Guidelines for Use of Laboratory Animals and were pre-approved by the Institutional Animal Care and Use Committee of Taipei Medical University. Osteoblasts were seeded in Dulbecco’s modified Eagle’s medium (DMEM; Gibco-BRL, Grand Island, NY, USA) supplemented with 10 % heat-inactivated fetal bovine serum, l-glutamine, penicillin (100 IU/ml), and streptomycin (100 μg/ml) in 75-cm2 flasks at 37 °C in a humidified atmosphere of 5 % CO2. Osteoblasts were grown to confluence prior to drug treatment. Only the first passage of rat osteoblasts was used in this study.
Sources of cellular oxidative stress and NO
SNP was purchased from Sigma (St. Louis, MO, USA), was freshly dissolved in phosphate-based saline (PBS) buffer (0.14 M NaCl, 2.6 mM KCl, 8 mM Na2HPO4, and 1.5 mM KH2PO4), and was protected from light. Our previous study showed that administration of 0.3 mM SNP to osteoblasts significantly increased cellular NO levels without affecting cell viability (Chang et al. 2006). However, 0.2 mM hydrogen peroxide (HP), purchased from Sigma, significantly induced cellular oxidative stress and cell damage. Thus, 0.3 mM SNP and 0.2 mM HP were used in the present study as respective sources of NO and oxidative stress.
Determination of cellular oxidative stress
Levels of intracellular reactive oxygen species (ROS) were quantified to evaluate cellular oxidative stress to osteoblasts after exposure to HP according to a previously described method (Chen et al. 2010a, b). Briefly, osteoblasts (5 × 105 cells per well) were cultured in 12-well tissue culture plates overnight and then were cotreated with HP and 2′,7′-dichlorofluorescin diacetate (Molecular Probes, Eugene, OR, USA), an ROS-sensitive dye. After drug treatment, osteoblasts were harvested and suspended in 1 × PBS buffer. Relative fluorescent intensities in osteoblasts were quantified using a flow cytometer (EPICS XL, Beckman Coulter, Fullerton, CA, USA).
Quantification of cellular NO levels
Cellular NO levels were determined by detecting levels of total nitrite, an oxidative product of NO, in culture medium of osteoblasts following the technical bulletin of the Bioxytech NO assay kit (OXIS International, Portland, OR, USA) as described previously (Chen et al. 2013). In this kit, nitrate reductase is provided to reduce nitrate to nitrite, and then, the total nitrite in the culture medium is detected. After drug treatment, the culture medium of osteoblasts was collected and centrifuged. The supernatant fractions were collected and reacted with nitrate reductase. Following reaction of the supernatant with sulfanilamide and N-1-naphthylethylenediamine, a colorimetric azo compound was formed and quantified using an Anthos 2010 microplate photometer (Anthos Labtec Instruments, Lagerhausstrasse, Wals/Salzburg, Austria).
Assay of alkaline phosphatase (ALP) activity
After drug treatment, the ALP activity of osteoblasts was determined by detecting the formation of p-nitrophenol, a product of p-nitrophenyl phosphate catalyzed by ALP, according to a colorimetric procedure provided by a Sigma Diagnostics Alkaline, Acid and Prostatic Acid Phosphatase kit (Sigma) as described previously (Ho et al. 2009).
Assay of cell viability
Effects of NO preconditioning and HP on cell viability were assayed using a trypan blue exclusion method as described previously (Chang et al. 2011). Briefly, osteoblasts (2 × 104 cells) were seeded in 24-well tissue culture plates. After drug treatment, cells were trypsinized with 0.1 % trypsin–EDTA (Gibco-BRL). Following centrifugation and washing, neuro-2a cells were suspended in PBS and stained with trypan blue dye (Sigma). Fractions of dead cells with a blue signal were visualized and counted using a reverse-phase microscope (Nikon, Tokyo, Japan).
Quantification of DNA fragmentation
DNA fragmentation was quantified with a cellular DNA fragmentation enzyme-linked immunosorbent assay (ELISA) kit (Boehringer Mannheim, Indianapolis, IN, USA) as previously described (Lin et al. 2012). Briefly, osteoblasts (2 × 105 cells) were subcultured in 24-well tissue culture plates and labeled with 5-bromo-2′-deoxyuridine (BrdU) overnight. Cells were harvested and suspended in the culture medium. One hundred microliters of cell suspension was added to each well of 96-well tissue culture plates. After drug treatment, amounts of BrdU-labeled DNA in the cytoplasm were quantified using an Anthos 2010 microplate photometer (Anthos Labtec Instruments).
Analysis of apoptotic cells
Apoptotic osteoblasts were determined using propidium iodide (PI) to detect DNA fragments in nuclei according to a previously described method (Chio et al. 2013). After drug treatment, osteoblasts were harvested and fixed in cold 80 % ethanol. Following centrifugation and washing, fixed cells were stained with PI and analyzed by flow cytometry (Beckman Coulter).
Confocal microscopic analysis of Bcl-XL and Bax protein translocation
Bcl-XL and Bax proteins in osteoblasts were recognized by specific antibodies and visualized using confocal microscopy as described previously (Chen et al. 2010a, b). Briefly, after drug treatment, osteoblasts were fixed with a fixing reagent (acetone/ methanol = 1: 1) at −20 °C for 10 min. Following rehydration, cells were incubated with 0.2 % Triton X-100 at room temperature for 15 min. Bcl-XL and Bax proteins in osteoblasts were immunodetected by reacting with rabbit polyclonal antibodies, respectively, raised against mouse Bcl-XL and Bax proteins (Transduction Laboratories, Lexington, KY, USA) at 4 °C overnight. After washing, osteoblasts were sequentially reacted with the 2nd antibodies with biotin-SP-conjugated AffiniPure goat antirabbit immunoglobulin G (IgG; Jackson ImmunoResearch, West Grove, PA, USA) at room temperature for 1 h. After washing, the third antibody with Cy3-conjugated streptavidin (Jackson ImmunoResearch) was added to the cells and reacted at room temperature for 30 min. Mitochondria of osteoblasts were stained with 3,3′-dihexyloxacarbocyanine (DiOC6) (Molecular Probes), a positively charged dye, at 37 °C for 30 min. A confocal laser scanning microscope (Model FV500, Olympus, Tokyo, Japan) was utilized for sample observation. An Olympus PlanApo 60x/1.4 NA oil-immersion objective lens was used to visualize the microtubules. The excitation wavelength was set to 568 nm, while a 585-nm band-pass filter was utilized to collect the emitted light. Images were acquired using FluoView software (version 4.0, Olympus Optical, Tokyo, Japan).
Reverse-transcription (RT) and real-time polymerase chain reaction (PCR) assays
Messenger (m)RNA from control and drug-treated osteoblasts was prepared for RT-PCR analyses of Bcl-XL, Bax, GATA-5, and β-actin mRNAs according to instructions of the ExpressDirect™ mRNA Capture and RT System for an RT-PCR kit (Pierce, Rockford, IL, USA). Oligonucleotides for the PCR analyses of rat Bcl-XL, Bax, GATA-5, and β-actin were designed and synthesized by Clontech Laboratories (Palo Alto, CA, USA). The oligonucleotide sequences of the respective upstream and downstream primers for these mRNA analyses were 5′-TTGGACAATGGACTGGTTG-3′ and 5′-GTAGAGTGGATGGTCAGTG-3′ for Bcl-XL (Yamanaka et al. 2002), 5′-AGGATCGAGCAGAGAGGATG-3′ and 5′-GGTCCCGAAGTAGGAGAGGA-3′ for Bax (Yamanaka et al. 2002), 5′-GAACCAGTGTGCAACGCCTG-3′ and 5′-CTAGGCCAGGGCCAGGGCACACCAG-3′ for GATA-5 (Nemer et al. 1999), and 5′-TATGGAGAAGATTTGGCACC-3′ and 5′-TATGGAGAAGATTTGGCACC-3′ for β-actin (Chen et al. 2010a, b). After the PCR assay, the products were loaded and separated in a 1.8 % agarose gel containing 0.1 μg/ml ethidium bromide. The intensities of the DNA bands in the agarose gel were quantified with the aid of an UVIDocMw version 99.03 digital imaging system (UVtec Limited, Cambridge, UK). A real-time PCR analysis was carried out using iQSYBR Green Supermix (Bio-Rad, Hercules, CA, USA) and a MyiQ single-color real-time PCR detection system (Bio-Rad) as described previously (Wu et al. 2013).
Immunodetection of GATA-5 protein
Amounts of GATA-5 were immunodetected as described previously (Ho et al. 2009). After drug treatment, osteoblasts were washed with 1 × PBS buffer. Cell lysates were prepared in ice-cold radioimmunoprecipitation assay (RIPA) buffer (25 mM Tris–HCl (pH 7.2), 0.1 % sodium dodecylsulfate (SDS), 1 % Triton X-100, 1 % sodium deoxycholate, 0.15 M NaCl, and 1 mM EDTA). To avoid protein degradation, a mixture of proteinase inhibitors, including 1 mM phenyl methyl sulfonyl fluoride, 1 mM sodium orthovanadate, and 5 µg/ml leupeptin, was added to the RIPA buffer. Protein concentrations were quantified by a bicinchonic acid protein assay kit (Pierce). Cytosolic proteins (100 μg/well) were subjected to SDS–polyacrylamide gel electrophoresis (PAGE) and transferred to nitrocellulose membranes. Membranes were blocked with 5 % nonfat milk at 37 °C for 1 h. Immunodetection of GATA-5 was carried out using a rabbit polyclonal antibody against mouse GATA-5 (Santa Cruz Biotechnology, Santa Cruz, CA, USA). The cellular β-actin protein was immunodetected using a mouse monoclonal antibody against mouse β-actin (Sigma) as an internal standard. Intensities of the immunoreactive bands were determined using an UVIDocMw version 99.03 digital imaging system (UVtec Limited).
GATA-5 knockdown
Expression of GATA-5 in osteoblasts was knocked down using an RNA interference (RNAi) method following a small interfering (si)RNA transfection protocol provided by Santa Cruz Biotechnology as described previously (Wu et al. 2013). GATA-5 siRNA was a pool of 3 target-specific 20 ~ 25-nt siRNAs designed to knockdown GATA-3’s expression. After culturing osteoblasts in antibiotic-free DMEM at 37 °C in a humidified atmosphere of 5 % CO2 for 24 h, the siRNA duplex solution, which was diluted in the siRNA transfection medium (Santa Cruz Biotechnology), was added to the osteoblasts. Scrambled siRNA, purchased from Santa Cruz Biotechnology, was applied to control cells as a negative standard. After transfection for 24 h, the medium was replaced with normal DMEM, and osteoblasts were treated with NO or HP.
Statistical analysis
Statistical differences between the control and drug-treated groups were considered significant when the p value of Duncan’s multiple range test was <0.05. Statistical analysis between drug-treated groups was carried out using a two-way analysis of variance (ANOVA).
Results
To determine the effects of NO on cellular oxidative stress and osteoblast injuries, levels of intracellular ROS, ALP activity, cell survival, and apoptotic cells were analyzed (Table 1). Exposure of osteoblasts to a low concentration (0.3 mM) of SNP for 24 h caused a significant 32 % increase in cellular NO amounts but did not lead to cell damage (data not shown). Thus, such a treatment was used in the present study as the source of NO pretreatment. In comparison, administration of 0.2 mM HP to osteoblasts for 24 h caused a 5.6-fold increase in levels of intracellular ROS (Table 1). Pretreatment with NO for 24 h did not induce significant oxidative stress to osteoblasts but significantly lowered HP-induced intracellular ROS production by 49 %. HP decreased ALP activity in osteoblasts by 57 % (Table 1). NO pretreatment did not affect the ALP activity but significantly lessened the HP-induced reduction in enzyme activity by 41 %. Exposure to HP caused 51 % of osteoblasts to die (Table 1). However, pretreatment with NO did not cause osteoblast death but led to a 49 % rebound in HP-induced cell death. Administration of HP to osteoblasts triggered 54 % of cells to undergo apoptosis (Table 1). After NO precondition, HP-induced cell apoptosis was attenuated by 55 %.
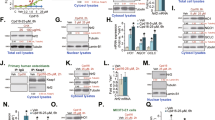
The cellular distributions of Bax and Bcl-XL proteins in osteoblasts were visualized to determine whether NO’s protection was mediated by alterations in the levels and translocation of these 2 apoptosis-related proteins (Fig. 1; Table 2). Exposure of osteoblasts to HP obviously increased cellular Bax levels (Fig. 1a). Translocation of the Bax protein from the cytoplasm to mitochondria was augmented after administration of HP. NO pretreatment did not affect cellular Bax protein levels and its translocation to mitochondria. After pretreatment with NO, the HP-caused enhancement in cellular levels and translocation of Bax protein were suppressed (Fig. 1a). In comparison, pretreatment with NO increased basal levels of the Bcl-XL protein and its translocation from the cytoplasm to mitochondria (Fig. 1b). These fluorescent signals were quantified and statistically analyzed (Table 2). Exposure of osteoblasts to HP increased Bax levels and its translocation to mitochondria by 3.5- and 3.8-fold, respectively. Meanwhile, NO pretreatment decreased HP-induced augmentations in the level and translocation of Bax by 43 and 47 %, respectively (Table 2). In contrast, NO pretreatment caused 3.7- and 2.6-fold increases in the Bcl-XL protein level and its translocation to mitochondria (Table 2).

Effects of nitric oxide (NO) pretreatment on hydrogen peroxide (HP)-induced translocation of Bax and Bcl-XL proteins from the cytoplasm to mitochondria. Osteoblasts prepared from neonatal rat calvaria were pretreated with 0.3 mM sodium nitroprusside for 24 h as the source of NO and then exposed to HP for a further 6 h. Distributions of Bax (a) and Bcl-XL (b) were immunodetected using antibodies with Cy3-conjugated streptavidin as described in “Materials and methods.” Mitochondria of osteoblasts were stained with 3,3′-dihexyloxacarbocyanine, a positively charged dye. Fluorescent images were visualized using a confocal laser scanning microscope at ×200
To validate the roles of apoptosis-related Bcl-X L and Bax gene expressions in NO’s protection against HP-induced osteoblast insults, RT- and real-time PCR analyses were carried out on osteoblasts (Fig. 2). In untreated osteoblasts, Bcl-XL mRNA was detected (Fig. 2a, top panel, lane 1). Levels of Bcl-XL mRNA were inhibited following administration of HP (lane 2). NO pretreatment induced Bcl-XL mRNA synthesis in osteoblasts (lane 3). After pretreatment with NO, the HP-caused inhibition of Bcl-XL mRNA dropped (lane 4). Bax mRNA was detected in untreated osteoblasts (Fig. 2a, middle panel, lane 1). Neither HP, NO pretreatment, nor a combination of NO pretreatment and HP changed Bax mRNA levels in osteoblasts (lanes 2 ~ 4). Amounts of β-actin mRNA were determined as the internal standard (Fig. 2a, bottom panel). These DNA bands were quantified and statistically analyzed (Fig. 2b). HP inhibited Bcl-XL mRNA production by 56 %. Pretreatment with NO induced 55 % of Bcl-XL mRNA expression and completely ameliorated HP-induced suppression of this mRNA synthesis. Bax mRNA production was not affected by HP, NO pretreatment, or a combination of NO pretreatment and HP (Fig. 2b). These effects were further confirmed using real-time PCR analyses (Fig. 2c). NO pretreatment led to 86 % induction of Bcl-XL mRNA in osteoblasts and alleviated HP-triggered inhibition of antiapoptotic mRNA levels by 82 %. Amounts of Bax mRNA in osteoblasts did not change following treatment with HP or NO pretreatment (Fig. 2c).

Effects of nitric oxide (NO) pretreatment on hydrogen peroxide (HP)-induced modulations of Bax and Bcl-XL mRNA expressions. Osteoblasts prepared from neonatal rat calvaria were pretreated with 0.3 mM sodium nitroprusside for 24 h as the source of NO and then exposed to HP for a further 6 h. Levels of Bax and Bcl-XL mRNA in osteoblasts were determined by RT-PCR analyses (a). Amounts of β-actin mRNA were quantified as the internal control. These DNA bands were quantified and statistically analyzed (b). A real-time PCR was further conducted to confirm such effects (c). Each value represents the mean ± SEM, n = 6. The symbols asterisk and hash symbol indicate that a value significantly (p < 0.05) differed from the respective control and HP-treated groups, respectively
RNA and protein levels of GATA-5 were quantified to evaluate the roles of this transcription factor in NO’s protection (Fig. 3). The GATA-5 protein was immunodetected in untreated osteoblasts (Fig. 3a, top panel, lane 1). HP decreased GATA-5 protein synthesis (lane 2). NO preconditioning enhanced cellular GATA-5 levels and eased the HP-induced reduction in GATA-5 levels (lanes 3 and 4). β-Actin was immunodetected as the internal control (bottom panel). These protein bands were quantified and statistically analyzed (Fig. 3b). HP significantly decreased GATA-5 protein synthesis by 64 %. NO pretreatment augmented cellular GATA-5 amounts by 36 % and completely lowered HP-induced reduction in the synthesis of this transcription factor (Fig. 3b). GATA-5 mRNA was detected in untreated osteoblasts (Fig. 3c, top panel, lane 1). After administration of HP, GATA-5 mRNA production was inhibited (lane 2). NO pretreatment slightly induced GATA-5 mRNA synthesis (lane 3). Pretreatment with NO alleviated HP-caused inhibition of GATA-5 mRNA synthesis (lane 4). β-Actin mRNA in osteoblasts was detected as the internal standard (bottom panel). Real-time PCR analyses further showed that HP significantly inhibited GATA-5 mRNA expression by 66 % (Fig. 3d). Pretreatment with NO caused a 35 % induction of GATA-5 mRNA expression and a complete rebound from HP-induced suppression of this mRNA synthesis.

Effects of nitric oxide (NO) pretreatment on hydrogen peroxide (HP)-induced suppression of GATA-5 mRNA and protein expressions. Osteoblasts prepared from neonatal rat calvaria were pretreated with 0.3 mM sodium nitroprusside for 24 h as the source of NO and then exposed to HP for a further 6 or 24 h. Amounts of GATA-5 were immunodetected (a, top panel). Levels of β-actin mRNA were quantified as the internal control (bottom panel). These protein bands were quantified and statistically analyzed (b). Levels of GATA-5 and β-actin mRNA in osteoblasts were determined by RT-PCR analyses (c). A real-time PCR was further conducted to evaluate the effects of NO pretreatment and HP on GATA-5 mRNA expression (d). Each value represents the mean ± SEM, n = 6. The symbols asterisk and hash symbol indicate that a value significantly (p < 0.05) differed from the respective control and HP-treated groups, respectively
Translation of GATA-5 mRNA was knocked down to further evaluate the roles of this transcription factor in Bcl-XL mRNA expression and NO’s protection (Fig. 4). Transforming GATA-5 siRNA into osteoblasts significantly decreased protein levels of this transcription factor (Fig. 4a, top panel, lane 2). β-Actin was immunodetected as the internal standard (bottom panel). These protein bands were quantified and analyzed (Fig. 4b). Application of GATA-5 siRNA to osteoblasts led to a 72 % reduction in cellular GATA-5 amounts. In parallel with this decrease, the expression of Bcl-XL mRNA was inhibited following GATA-5 siRNA administration (Fig. 4c, top panel, lane 2). β-Actin was immunodetected as the internal standard (bottom panel). A real-time PCR analysis showed that application of GATA-5 siRNA inhibited Bcl-XL mRNA expression by 75 % (Fig. 4d). Scrambled siRNA did not affect production of Bcl-XL mRNA by osteoblasts (Fig. 4e). However, application of GATA-5 siRNA completely inhibited NO pretreatment-induced Bcl-XL mRNA expression. The NO pretreatment-involved alleviation of HP-triggered inhibition of Bcl-XL mRNA expression was also totally decreased by GATA-5 siRNA (Fig. 4e).

Effect of GATA-5 knockdown on Bcl-X L gene expression. Osteoblasts were treated with GATA-5 siRNA for 24 h. Scrambled siRNA was administered to control cells as the negative standard. Levels of GATA-5 protein were immunodetected (a). An RT-PCR analysis was carried out to determine Bcl-XL mRNA production by osteoblasts (c). These protein and DNA bands were quantified and statistically analyzed (b and d). After knocking down GATA-5 mRNA expression, osteoblasts were pretreated with 0.3 mM sodium nitroprusside for 24 h as the source of nitric oxide (NO) and then exposed to hydrogen peroxide (HP) for a further 6 h. Real-time PCR analyses were conducted to determine Bcl-XL mRNA expression (E). Each value represents the mean ± SEM, n = 6. The symbols asterisk, hash symbol and ampersand indicate that a value significantly (p < 0.05) differed from the control, HP-, and NO + HP-treated groups, respectively
ALP activity, cell survival, DNA fragmentation, and cell apoptosis were assayed to further determine the roles of GATA-5 in NO pretreatment-involved cell protection (Fig. 5). Exposure of osteoblasts to HP caused 55 and 54 % reductions in ALP activity and cell survival, respectively (Fig. 5a, b). NO preconditioning caused respective HP-induced decreases in ALP activity and cell viability by 48 and 52 %. However, application of GATA-5 siRNA to osteoblasts, respectively, led to significant 100 and 85 % decreases in NO’s protection against HP-induced alterations in ALP activity and cell survival (Fig. 5a, b). As to the mechanism, HP-induced DNA fragmentation and cell apoptosis were, respectively, suppressed following NO pretreatment by 81 and 75 % (Fig. 5c, d). In comparison, after administration of GATA-2 siRNA, NO’s protection against DNA fragmentation and cell apoptosis was significantly alleviated (Fig. 5c, d).

Survival-mediating roles of GATA-5 in osteoblasts. Osteoblasts were treated with GATA-5 siRNA for 24 h. Scrambled siRNA was administered to control cells as the negative standard. After knocking down GATA-5 expression, osteoblasts were pretreated with 0.3 mM sodium nitroprusside for 24 h as the source of nitric oxide (NO) and then exposed to hydrogen peroxide (HP). Alkaline phosphatase (ALP) activity was analyzed according to colorimetric methods (a). Cell survival was assayed using a trypan blue exclusion method (b). DNA fragmentation (c) and apoptotic cells (d) were quantified using flow cytometry. Each value represents the mean ± SEM, n = 12. The symbols asterisk, hash symbol and ampersand indicate that a value significantly (p < 0.05) differed from the control, HP-, and NO + HP-treated groups, respectively
Discussion
This study showed that NO can protect osteoblasts against HP-induced insults. Pretreatment of osteoblast with SNP slightly elevated cellular NO levels but did not cause cell insults. In comparison, exposure of osteoblasts to HP raised amounts of intracellular ROS and simultaneously induced cell death. Interestingly, NO pretreatment defended osteoblasts against HP-induced cell damage. Our previous studies showed that pretreatment with low concentrations of NO could lower cell insults induced by high concentrations of NO (Tai et al. 2007; Chen et al. 2010a, b). In ischemia-/reperfusion-induced cell injury, NO has been shown to produce protective effects on cardiomyocytes, hepatocytes, and endothelial cells (Cottart et al. 1999; Murillo et al. 2011). NO has biphasic effects on bone activities (Ralston 1997). Overproduction of NO led to osteoblast apoptosis (Chen et al. 2002; 2005a, b). This study further showed the protection of NO pretreatment against HP-induced injury to osteoblasts. Osteoblasts play a key role in bone formation (Sims and Vrahnas 2014). Das et al. reported that NO pretreatment had the therapeutic potential for preventing myocyte cell death following ischemia/reperfusion injury (Das et al. 2005). Therefore, NO may provide a de novo therapeutic strategy for bone diseases such as bone fracture.
NO defended against HP-induced insults to osteoblasts via an antiapoptotic mechanism. Administration of HP time-dependently decreased ALP activity and caused osteoblast death. ALP is one of the critical markers of osteoblast metabolism (Collin-Osdoby et al. 1995). Thus, HP can augment cellular oxidative stress, leading to osteoblast dysfunction or even death. Exposure of osteoblasts to HP induced DNA fragmentation and cell cycle arrest at the sub-G phase. DNA fragmentation and the appearance of a hypodiploid sub-G1 peak indicate that osteoblasts are undergoing apoptosis (Lin et al. 2012; Kawatani and Imoto 2003). Thus, HP induces insults to osteoblast via an apoptotic mechanism. In comparison, the present results revealed that pretreatment of osteoblasts with NO lowered HP-induced osteoblast dysfunction and cell death. Additionally, NO led to significant alleviation of DNA fragmentation and cell cycle arrest at the sub-G1 phase induced by HP. In ischemia/reperfusion injury, NO pretreatment can modulate mitochondrial function and subsequently produces protective effects on various types of cells (Cottart et al. 1999; Murillo et al. 2011). Our previous studies also demonstrated that NO pretreatment attenuated oxidative stress-induced alterations in the mitochondrial membrane potential and caspase activation (Chang et al. 2006). As a result, NO can protect osteoblasts from HP-induced cell damage through a mitochondrion-dependent antiapoptotic mechanism.
NO had differential effects on the regulation of Bcl-XL and Bax protein translocation from the cytoplasm to mitochondria in osteoblasts. Exposure to HP increased cellular Bax levels. In parallel, translocation of the Bax protein from the cytoplasm to mitochondria was also enhanced by HP. However, pretreatment with NO decreased such levels and translocation. Mitogen-activated protein kinases (MAPKs) participate in phosphorylation of the Bax protein and stimulate its translocation from the cytoplasm to mitochondria (van Laethem et al. 2004). Our previous study also showed that oxidative stress decreases the activities of p38 MAPK, c-Jun N-terminal kinase (JNK), and Erk1/2 in osteoblasts (Chen et al. 2010a, b). Meanwhile, NO pretreatment lowered the suppressive effects. Thus, pretreatment with NO may lower the HP-induced increase in translocation of the Bax protein possibly via suppression of MAPK activities. Bax translocation from the cytoplasm to mitochondria is essential for inducing a cascade of apoptotic signals, including depolarization of the mitochondrial membrane and release of apoptotic factors (Hsu et al. 1997; Goyal 2001). Thus, a decrease in the proportion of Bax protein translocated to mitochondria is another important determinant inducing NO’s protection against apoptotic insults. In parallel with induction of Bcl-XL mRNA, NO augmented cellular Bcl-XL protein levels and its translocation from the cytoplasm to mitochondria. Bcl-XL has antiapoptotic effects by stopping Bax-induced disruption of the mitochondrial membrane (Pagliari et al. 2005). Therefore, NO decreased Bax translocation but enhanced Bcl-XL levels and its translocation. Taken together, the differential effects on translocation of Bax and Bcl-XL proteins are critical reasons to further explain how NO protects osteoblasts from apoptotic insults.
Differential regulation of Bcl-X L and Bax gene expressions is involved in NO’s protection against HP-induced osteoblast apoptosis. Administration of HP inhibited Bcl-XL mRNA production without affecting Bax mRNA synthesis. Bcl-XL is thought to be an antiapoptotic protein because it can suppress Bax-mediated apoptosis (Pagliari et al. 2005; Monaco et al. 2013). Huang et al. (2005) reported that overexpression of the Bcl-X L gene inhibits Bax translocation and Cyt c release and prevents cardiac cells from apoptosis (Huang et al. 2005). A dominant negative form of the Bcl-XL protein produced by deleting the BH1 domain can trigger cells to undergo apoptosis (Kawatani and Imoto 2003). Thus, the rebound in the inhibition of Bcl-X L gene expression induced by HP may be one of the important reasons explaining NO’s protection against osteoblast apoptosis. HP had no effect on Bax mRNA production. Our previous results, as with the present data from a confocal microscopic analysis, revealed that HP increases cellular levels of Bax protein in osteoblasts (Chen et al. 2002). NO pretreatment can lower the HP-induced increase in the Bax protein. Thus, NO protects osteoblasts from HP-induced apoptotic insults mainly through pretranscriptional induction of Bcl-XL mRNA synthesis and posttranscriptional downregulation of cellular Bax protein levels.
GATA-5 regulates Bcl-X L gene expression and thus plays a critical role in NO’s protection. Exposure of osteoblasts to HP suppressed RNA and protein levels of GATA-5. Pretreatment with NO significantly increased GATA-5 mRNA and protein expressions and concurrently alleviated the suppressive effects induced by HP. GATA-DNA-binding elements, (A/T)GATA(A/G), are found in the 5′-end promoter regions of the Bcl-X L gene (Chen et al. 2010a, b; Yu et al. 2005). To further validate the role of this antiapoptotic protein in NO’s protection, GATA-5 translation was knocked down using RNAi. In parallel with downregulation of the GATA-5 protein, the rebound effects of NO pretreatment on HP-induced suppression of Bcl-XL mRNA synthesis were lowered. Thus, NO-mediated Bcl-XL mRNA induction is involved by the upregulation of GATA-5 protein and RNA levels in osteoblasts. Consequently, after transformation of GATA-5 RNAi, a decrease in Bcl-X L gene expression lowered the protective effects of NO on ALP activity, cell survival, DNA fragmentation, and cell apoptosis. GATA-5, like other members GATA-1 ~ 4 and GATA-6, participates in regulating cell proliferation and differentiation (Capo-Chichi et al. 2005; Peters et al. 2014). Our previous study showed the roles of GATA-3 in mediating survival signal in osteoblasts (Chen et al. 2010a, b). This is the first study to demonstrate that GATA-5 has antiapoptotic effects on NO’s protection against HP-induced osteoblast apoptosis via upregulation of Bcl-X L gene expression.
In conclusion, this study has shown that HP raised cellular oxidative stress and simultaneously caused reductions in ALP activity, cell survival, and induction of cell apoptosis. Pretreatment with NO significantly lowered the toxic effects via an antiapoptotic mechanism. After NO pretreatment, the HP-induced increase in the proportion of Bax protein being translocated from the cytoplasm to mitochondria was attenuated, but Bcl-XL’s translocation was enhanced. Pretreatment with NO selectively alleviated the HP-induced suppression of the GATA-5 transcription factor. RNAi analyses further showed that GATA-5 was involved in upregulating Bcl-X L gene expression and contributed to NO’s protection against HP-induced osteoblast insults. Therefore, NO can protect osteoblasts from HP-induced cell apoptosis via multiple mechanisms, including (1) the GATA-5-mediated transcriptional induction of Bcl-X L gene expression; (2) the posttranscriptional regulation of Bax gene expression; and (3) the differential regulation of Bax and Bcl-XL protein translocation.
References
Armour KE, van’t Hof RJ, Grabowski PS, Reid DM, Ralston SH (1999) Evidence for a pathogenic role of nitric oxide in inflammation-induced osteoporosis. J Bone Miner Res 14:2137–2142
Burch JB (2005) Regulation of GATA gene expression during vertebrate development. Semin Cell Dev Biol 16:71–81
Capo-Chichi CD, Rula ME, Smedberg JL, Vanderveer L, Parmacek MS, Morrisey EE, Godwin AK, Xu XX (2005) Perception of differentiation cues by GATA factors in primitive endoderm lineage determination of mouse embryonic stem cells. Dev Biol 286:574–586
Chang CC, Liao YS, Lin YL, Chen RM (2006) Nitric oxide protects osteoblasts from oxidative stress-induced apoptotic insults via a mitochondria-dependent mechanism. J Orthop Res 24:1917–1925
Chang HC, Chen TG, Tai YT, Chen TL, Chiu WT, Chen RM (2011) Resveratrol attenuates oxidized LDL-evoked Lox-1 signaling and consequently protects against apoptotic insults to cerebrovascular endothelial cells. J Cereb Blood Flow Metab 31:842–854
Chen RM, Lin YL, Jean WC, Chen JS, Wang JH, Liu HC (2002) Nitric oxide induces osteoblast apoptosis through the de novo synthesis of Bax protein. J Orthop Res 20:295–302
Chen RM, Chen TL, Chiu WT, Chang CC (2005a) Molecular mechanism of nitric oxide-induced osteoblast apoptosis. J Orthop Res 23:462–468
Chen RM, Chen TL, Lin YL, Chen TG, Tai YT (2005b) Ketamine reduces nitric oxide biosynthesis in human umbilical vein endothelial cells through downregulating endothelial nitric oxide synthase expression and intracellular calcium levels. Crit Care Med 33:1044–1049
Chen RM, Lin YL, Chou CW (2010a) GATA-3 transduces survival signals in osteoblasts through upregulation of bcl-xL gene expression. J Bone Miner Res 25:2193–2204
Chen TL, Wu GJ, Hsu CS, Fong TH, Chen RM (2010b) Nitrosative stress induces osteoblast apoptosis through downregulating MAPK-mediated NFκB/AP-1 activation and subsequent Bcl-XL expression. Chem Biol Interact 184:359–365
Chen RM, Tai YT, Chen TG, Lin TH, Chang HC, Chen TL, Chen RM (2013) Propofol protects against nitrosative stress-induced breakage of the blood-brain barrier through reducing apoptotic insults to cerebrovascular endothelial cells. Surgery 154:58–68
Chio CC, Lin JW, Cheng HA, Chiu WT, Wang YH, Wang JJ, Chen RM (2013) MicroRNA-210 targets antiapoptotic Bcl-2 expression and mediates hypoxia-induced apoptosis of neuroblastoma cells. Arch Toxicol 87:458–468
Cochain C, Channon KM, Silvestre JS (2013) Angiogenesis in the infarcted myocardium. Antioxid Redox Signal 18:1100–1113
Collin-Osdoby P, Nickols GA, Osdoby P (1995) Bone cell function, regulation, and communication, a role for nitric oxide. J Cell Biochem 57:399–408
Cottart CH, Do L, Blanc MC, Vaubourdolle M, Descamps G, Durand D, Galen FX, Clot JP (1999) Hepatoprotective effect of endogenous nitric oxide during ischemia-reperfusion in the rat. Hepatology 29:809–813
Das A, Xi L, Kukreja RC (2005) Phosphodiesterase-5 inhibitor sildenafil preconditions adult cardiac myocytes against necrosis and apoptosis. Essential role of nitric oxide signaling. J Biol Chem 280:12944–12955
Evans T, Felsenfeld G (1989) The erythroid-specific transcription factor Eryf1, a new finger protein. Cell 58:877–885
Evans DM, Ralston SH (1996) Nitric oxide and bone. J Bone Miner Res 11:300–305
Fiore CE, Pennisi P, Cutuli VM, Prato A, Messina R, Clementi G (2000) L-arginine prevents bone loss and bone collagen breakdown in cyclosporin A-treated rats. Eur J Pharmacol 408:323–326
Giustina A, Mazziotti G, Canalis E (2008) Growth hormone, insulin-like growth factors, and the skeleton. Endocr Rev 29:535–559
Goyal L (2001) Cell death inhibition, keeping caspases in check. Cell 104:805–808
Ho WP, Chen TL, Chiu WT, Tai YT, Chen RM (2005) Nitric oxide induced osteoblast apoptosis through a mitochondrial-dependent pathway. Ann NY Acad Sci 1042:460–470
Ho WP, Chan WP, Hsieh MS, Chen RM (2009) Runx2-mediated Bcl-2 gene expression contributes to nitric oxide protection against oxidative stress-induced osteoblast apoptosis. J Cell Biochem 108:1084–1093
Hsu YT, Wolter KG, Youle RJ (1997) Cytosol-to-membrane redistribution of Bax and Bcl-XL during apoptosis. Proc Natl Acad Sci USA 94:3668–3672
Huang J, Nakamura K, Ito Y, Uzuka T, Morikawa M, Hirai S, Tomihara K, Tanaka T, Masuta Y, Ishii K, Kato K, Hamada H (2005) Bcl-XL gene transfer inhibits Bax translocation and prolongs cardiac cold preservation time in rats. Circulation 112:76–83
Jamal SA, Reid LS, Hamilton CJ (2013) The effects of organic nitrates on osteoporosis, a systematic review. Osteoporos Int 24:763–770
Kawatani M, Imoto M (2003) Deletion of the BH1 domain of Bcl-2 accelerates apoptosis by acting in a dominant negative fashion. J Biol Chem 278:19732–19742
Lin JW, Chen JT, Hong CY, Lin YL, Wang KT, Yao CJ, Lai GM, Chen RM (2012) Honokiol traverses the blood-brain barrier and induces apoptosis of neuroblastoma cells via an intrinsic Bax-mitochondrion-cytochrome c-caspase protease pathway. Neuro-Oncol 14:302–314
Monaco G, Vervliet T, Akl H, Bultynck G (2013) The selective BH4-domain biology of Bcl-2-family members, IP3Rs and beyond. Cell Mol Life Sci 70:1171–1183
Moncada S, Palmer RMJ, Higgs EA (1991) Nitric oxide, physiology, pathophysiology and pharmacology. Pharmacol Rev 43:109–142
Murillo D, Kamga C, Mo L, Shiva S (2011) Nitrite as a mediator of ischemic preconditioning and cytoprotection. Nitric Oxide 25:70–80
Nemer G, Qureshi ST, Malo D, Nemer M (1999) Functional analysis and chromosomal mapping of Gata5, a gene encoding a zinc finger DNA-binding protein. Mamm Genome 10:993–999
Pagliari LJ, Kuwana T, Bonzon C, Newmeyer DD, Tu S, Beere HM, Green DR (2005) The multidomain proapoptotic molecules Bax and Bak are directly activated by heat. Proc Natl Acad Sci USA 102:17975–17980
Peters I, Dubrowinskaja N, Kogosov M, Abbas M, Hennenlotter J, von Klot C, Merseburger AS, Stenzl A, Scherer R, Kuczyk MA, Serth J (2014) Decreased GATA5 mRNA expression associates with CpG island methylation and shortened recurrence-free survival in clear cell renal cell carcinoma. BMC Cancer 14:101
Ralston SH (1997) Nitric oxide and bone, what a gas. Br J Rheumatol 36:831–838
Simon JN, Duglan D, Casadei B, Carnicer R (2014) Nitric oxide synthase regulation of cardiac excitation-contraction coupling in health and disease. J Mol Cell Cardiol 73C:80–91
Sims NA, Vrahnas C (2014) Regulation of cortical and trabecular bone mass by communication between osteoblasts, osteocytes and osteoclasts. Arch Biochem Biophys 561:22–28
Suzuki E, Evans T, Lowry J, Truong L, Bell DW, Testa JR, Walsh K (1996) The human GATA-6 gene, structure, chromosomal location, and regulation of expression by tissue-specific and mitogen-responsive signals. Genomics 38:283–290
Tai YT, Cherng YG, Chang CC, Hwang YP, Chen JT, Chen RM (2007) Pretreatment with low nitric oxide protects osteoblasts from high nitric oxide-induced apoptotic insults through regulation of c-Jun N-terminal kinase/c-Jun-mediated Bcl-2 gene expression and protein translocation. J Orthop Res 25:625–635
Takeda S, Karsenty G (2001) Central control of bone formation. J Bone Miner Metab 19:195–198
van Laethem A, van Kelst S, Lippens S, Declercq W, Vandenabeele P, Janssens S, Vandenheede JR, Garmyn M, Agostinis P (2004) Activation of p38 MAPK is required for Bax translocation to mitochondria, cytochrome c release and apoptosis induced by UVB irradiation in human keratinocytes. FASEB J 18:1946–1948
Wu TT, Tai YT, Cherng YG, Chen TG, Chen TL, Chang HC, Cen RM (2013) GATA-2 transduces LPS-induced il-1β gene expression in macrophages via a toll-like receptor 4/MD88/MAPK-dependent mechanism. PLoS ONE 8:e72404
Yamanaka M, Shirai M, Shiina H, Shirai M, Tanaka Y, Fujime M, Okuyama A, Dahiya R (2002) Loss of anti-apoptotic genes in aging rat crura. J Urol 168:2296–2300
Yu YL, Chiang YJ, Chen YC, Papetti M, Juo CG, Skoultchi AI, Yen JJ (2005) MAPK-mediated phosphorylation of GATA-1 promotes Bcl-XL expression and cell survival. J Biol Chem 280:29533–29542
Acknowledgments
This study was supported by grants from Shin Kong Wu Ho-Su Memorial Hospital (SKH-8302-101-DR-24), Wan-Fang Hospital (104swf04), and the National Science Council (NSC101-2314-B-038-003-MY3), Taipei, Taiwan.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Wu, GJ., Wang, W., Lin, YL. et al. Oxidative stress-induced apoptotic insults to rat osteoblasts are attenuated by nitric oxide pretreatment via GATA-5-involved regulation of Bcl-X L gene expression and protein translocation. Arch Toxicol 90, 905–916 (2016). https://doi.org/10.1007/s00204-015-1491-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00204-015-1491-z