
Abstract
In the last two decades, a large number of literature had focused on the biosynthesis of silver nanoparticles (AgNPs) from silver ions by bacteria and other microorganisms. This study infers that bacteria and other microorganisms do not per se synthesize AgNPs. All tested auto- and heterotrophic microorganisms in this study were killed by silver ions and could not as viable cells produce AgNPs. Microbial cell viability represented in colony-forming units and metabolic viability represented in aerobic respiration in all investigated microorganisms as well as photosynthesis in photoautotrophic microorganisms ceased by silver ions too early before AgNPs formation. The time required for AgNPs synthesis inversely related to the incubation temperature of the investigated microorganisms with silver ions where it requires only few minutes for nanoparticles formation at high temperature or autoclaving. The minimum inhibitory and minimum bactericidal and fungicidal concentrations of silver ions were significantly lower than AgNPs, indicating that silver ions are more efficient antimicrobial. The results presented in this study indicate that formation of AgNPs by eubacteria, cyanobacteria and fungi is not a vitally regulated cellular metabolic process and the mechanism occurs via bioreduction of silver ions to nanoparticles by organics released from the dead cells.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Over the past few decades, nanoparticles, with at least one dimension less than 100 nm, of metals attracted high attention for its new applications in various fields of industry. Among these nanomaterials, silver nanoparticles (AgNPs) seem to have attracted the most interest in terms of its potential applications. Chemical, physical and biological syntheses of AgNPs are hot topics of research due to the wide range of AgNPs applications and their antibacterial, antifungal and antiviral activities (Kim et al. 2007; Pal et al. 2007; Rai et al. 2009; et al. De Gusseme et al. 2010; Govindaraju et al. 2011; Lara et al. 2011; Brandt et al. 2012; Kim et al. 2012; Mohanty et al. 2012; Tamboli and Lee 2013; Eckhardt et al. 2013; Lok et al. 2014). Because of their antimicrobial properties, AgNPs are incorporated in apparel, footwear, paints, wound dressings, appliances, cosmetics and plastics (Sharma et al. 2009; Li et al. 2010).
In spite of the antimicrobial properties of AgNPs, microbial synthesis of these nanoparticles (Klaus et al. 1999, 2001; Mandal et al. 2006; Gurunathan et al. 2009; Sintubin et al. 2009) has been proposed as one of the alternative ecofriendly green methods to chemical and physical procedures that are fraught with many problems including the use of toxic solvents, generation of hazardous by-products and high energy consumption (Thakkar et al. 2010). Many microorganisms have been used for intra- or extra-cellular biosynthesis of AgNPs. Silver-containing nanocrystals of different compositions were synthesized by Pseudomonas stutzeri AG259 bacterium (Klaus et al. 1999, 2001). The silver reduction by the bacterium Proteus mirabilis PTCC 1710 to produce intra- and extra-cellular nanoparticles was suggested to occur through energy-dependent processes (Samadi et al. 2009). Possible involvement of proteins in synthesizing AgNPs was observed in the filamentous cyanobacterium, Plectonema boryanum UTEX 485 (Lengke et al. 2006). The white rot fungus, Phaenerochaete chrysosporium, reduced silver ions to form AgNPs where a protein was suggested to cause the reduction (Vigneshwaran et al. 2006a). Jain et al. (2011) indicated that silver nanoparticle synthesis by Aspergillus flavus occurs initially by a 33-kDa protein followed by a 35-kDa protein electrostatic attraction which stabilizes the nanoparticles by forming a capping agent. Geobacter sulfurreducens reduced silver ions to nanoparticles via a mechanism involving c-type cytochromes (Law et al. 2008). Exposure of Verticillium fungal biomass to aqueous silver ions resulted in the intracellular formation of AgNPs below the cell wall surface, possibly due to the reduction of the metal ions by enzymes present in the cell membrane (Mukherjee et al. 2001). Intracellular formation of AgNPs was also reported in the filamentous fungus Neurospora crassa (Castro-Longoria et al. 2011). In Fusarium oxysporum fungus, the reduction of silver ions to nanoparticles was attributed to an enzymatic process involving NADH-dependent reductase (Durán et al. 2005). Synthesis of AgNPs using α-NADPH-dependent nitrate reductase and phytochelatin in vitro has been demonstrated (Anil-Kumar et al. 2007), and the nitrate reductase present in the bacteria was suggested to aid in the synthesis of AgNPs (Kalimuthu et al. 2008). Nitrate reductase was also suggested to initiate AgNPs formation from silver ions in a Penicillium species (Naveen et al. 2010). However, silver ions used for the biosynthesis of AgNPs can also be used as antimicrobial agent (Slawson et al. 1992; Feng et al. 2000; Yamanaka et al. 2005). This study was devoted to explore the controversy between biosynthesis of AgNPs by microorganisms and using AgNPs as antimicrobial agents.
Materials and methods
Organisms and growth conditions
The cyanobacterium Spirulina platensis strain IAM M-135 was grown photoautotrophically in Spirulina–Ogawa–Terui (SOT) medium (Ogawa and Terui, 1970) under continuous illumination at 32 ± 1 °C with aeration (Morsy et al. 2008a). Growth was assessed by chlorophyll a (chl a) content. Chl a was dark-extracted under cooling in 90 % methanol overnight, clarified by centrifugation under cooling (4 °C) at 7000×g for 15 min and quantified spectrophotometrically according to the equations of Holden (1976). The mutant Escherichia coli HD701 constructed previously (Sauter et al. 1992) from wild-type E. coli MC4100 was maintained aerobically on nutrient agar and grown on nutrient broth with shaking (100 rpm) (Penfold et al. 2003; Morsy 2011a) at 35 °C. Growth of E. coli was followed spectrophotometrically at 600 nm. Aspergillus terreus was maintained on Sabouraud dextrose agar (Moss and McQuown 1969) slants and grown on Sabouraud sucrose broth (Koutb and Morsy 2011) at 30 °C. The composition of Sabouraud sucrose broth (SSB) is (g/L): sucrose 40, peptone 10, Actidione 0.5 and chloramphenicol 0.25. Growth of A. terreus was assessed by dry mass per 100 ml culture.
Bioreduction of silver ions and formation of AgNPs
Three culture replicates were incubated with shaking in dark in case of E. coli and A. terreus and in light in case of S. platensis. Silver nitrates were added to the 100-ml culture of each organism up to 1 mM AgNO3 at the exponential phase of the growth. The formation of the AgNPs was monitored by UV–visible spectroscopy at 430 nm. The effect of temperature on AgNPs synthesis was investigated.
Microbial cell viability during biosynthesis of AgNPs
Microbial cell viability was followed by monitoring the colony-forming units (CFU) in time intervals after addition of silver nitrates to each culture. Synthesis of AgNPs and the revival of E. coli and A. terreus cells as well as metabolic cell viability represented in its oxygenic respiration activity at aerobic conditions were followed in nutrient broth and Sabouraud dextrose broth, respectively. Formation of AgNPs by the cyanobacterium S. platensis as well as the cyanobacterial cell viability represented in its photosynthesis and respiration activities was followed up for 7 days in SOT medium.
Photosynthesis and respiration
Photosynthetic oxygen evolution in case of S. platensis and aerobic respiration for all tested microorganisms were followed in time intervals after addition of silver ions to each culture. Photosynthetic oxygen evolution of S. platensis was measured using a Clark-type oxygen electrode computerized to an oxygen monitoring system (OMS; Hansatech Instruments, Inc.). Oxygen evolution was measured in actinic light (44.5 µE m−1 s−1) passed through a red optical filter (Morsy 2011b). All measurements were taken at 32 °C ± 1 stabilized by a circulating water bath. Respiratory O2 uptake by each investigated microorganism was measured using the same above-mentioned Clark electrode and OMS at respective optimum temperature of each microorganism but in the dark.
Reduction of silver ions to nanoparticles by cyanobacterial polysaccharides of S. platensis
Exopolysaccharides (EPS) of S. platensis were prepared as described previously (Filali-Mouhim et al. 1993), lyophilized and stored at 4 °C for further experiments. The reduction of silver ions to nanoparticles by cyanobacterial polysaccharides of S. platensis was conducted at 15 psi and 121 °C for 5 min. The effect of silver ions and EPS concentrations on AgNPs formation was investigated.
Characterization of AgNPs
After formation of AgNPs, the cultures were centrifuged at 4000×g for 5 min at 20 °C. The pellet was discarded, and the supernatant containing AgNPs was centrifuged at 15,000×g for 20 min at 20 °C. After washing the precipitate (AgNPs) three times with distilled water, it was suspended in distilled water and stored in the dark at room temperature for further characterizations.
UV–visible spectroscopy
The wavelength absorption spectrum of AgNPs was monitored by UV–visible spectroscopy by recording the spectra between 300 and 900 nm and monitoring the appearance of the characteristic peak at 400–440 nm using UV–vis spectrophotometer (Thomson Scientific, model: Evolution 3000).
Transmission electron microscopy (TEM)
The morphology of AgNPs was characterized by TEM. An aliquot (10 μL) of aqueous suspension of AgNPs was transferred onto a carbon-film-coated copper grid and allowed to dry for 30 min. The grid was scanned, and the images of AgNPs were obtained by a JEOL-JEM-2010XII microscope operated at 200 kV with a magnification of up to 5 × 105 times. The TEM data of AgNPs shape and size were analyzed.
Minimum inhibitory concentration (MIC) and minimum bactericidal (MBC) and fungicidal (MFC) concentrations of silver ions versus nanoparticles
The concentration of silver nanoparticle solutions was calculated using the Beer–Lambert law, which correlates the optical density (OD) with concentration. Due to the linear relationship between OD and concentration, silver nanoparticle concentration in diluted solutions was calculated from a standard curve of the optical density at 430 nm (maximum absorbance wavelength) versus mass concentration (ppm). The minimum inhibitory concentration (MIC) of silver ions and nanoparticles was determined as the lowest concentration that inhibits the growth of a microorganism. A series of proper medium broth of each subject microorganism were mixed with the serially diluted silver ions or nanoparticles solutions, and a standard inoculum of the microorganism was added. The minimum inhibitory concentration (MIC) was determined as that in the first broth in which growth of the microorganism has been inhibited by the antimicrobial effect of silver ions or nanoparticles. The minimum bactericidal (MBC) and fungicidal (MFC) concentrations were determined from broth dilution MIC tests by subculturing to appropriate medium agar plates of each subject microorganism without silver ions or nanoparticles. MBC and MFC were determined as the lowest concentration of silver ions or nanoparticles required to kill the microorganism where no colony appeared upon subculturing.
Statistical analysis
The results of the MIC and MBC of silver ions versus nanoparticles were subjected to analysis of variance (one-way ANOVA) using PCSTAT program (version 1A). The treatment means were compared using the least significant difference (LSD) values at a significance level of P < 0.05.
Results
This study was devoted to explore the controversy between the biosynthesis of AgNPs by microorganisms and the antimicrobial activities of these nanoparticles. For this purpose, experiments were conducted to follow up simultaneously the microbial cell viability and the formation of AgNPs in time intervals after addition of silver ions to each microbial culture. Autotrophic microorganisms represented by the cyanobacterium S. platensis and heterotrophic microorganisms represented by the eubacterium E. coli HD701 and the fungus A. terreus were used in this investigation.
Upon addition of silver ions to the cultures, microbial cell viability was followed by monitoring the colony-forming units (CFU) in time intervals (Fig. 1). All tested auto- and heterotrophic microorganisms in this study were killed within 2 to 4 h of exposure to silver ions (Fig. 1). Metabolic viability represented in aerobic respiration in all investigated microorganisms as well as photosynthesis in autotrophic microorganisms was followed in time intervals after addition of silver ions to the culture (Fig. 2) of each microorganism at the exponential growth phase. For S. platensis, silver ions were added at the third day of culture where photosynthesis and respiration ceased within one hour of silver ions addition (Fig. 2a). The growth represented by chlorophyll a concentration in the culture ceased after addition of silver ions, and a decline phase appeared (Fig. 2a). For E. coli, silver ions were added at the fourth hour of culture. Aerobic respiration of E. coli (Fig. 2b) ceased upon addition of silver ions to the culture. Growth curve represented by bacterial optical density declined after silver ion addition (Fig. 2b). Similar results were obtained for the wild-type strain E. coli EGY previously isolated and identified by 16 sRNA (Abd-Alla et al. 2011). For the fungus A. terreus, silver ions were added at the second day of culture where aerobic respiration ceased within 2 h of silver ions addition and the growth curve declined (Fig. 2c).

Microbial cell viability represented as colony-forming units (CFU) of E. coli (a), S. platensis (b) and A. terreus (c) in response to 1 mM silver ions. Viable cells were enumerated on an agar plates of the appropriate medium and growth conditions for each microorganism as described in Materials and methods. The bars show the standard errors of three replicates

Cease of growth, photosynthesis and aerobic respiration by silver ions. a Photosynthesis, respiration and growth of S. platensis in response to 1 mM silver ions. Growth (open squares), photosynthesis (open circles) and respiration (closed triangles) were monitored at normal growth condition as described in Materials and methods. At the third day of culture, silver nitrate was added to the culture up to 1 mM and the growth (closed squares), photosynthesis (closed circles) and respiration (open cubes) were followed. b, c Cease of growth and aerobic respiration of E. coli and A. terreus, respectively, by silver ions. Growth (open squares) and aerobic respiration (closed squares) were monitored as described in Materials and methods. Upon addition of silver nitrate to the culture up to 1 mM, the growth (open circles) and aerobic respiration (closed circles) were also followed. The bars show the standard errors of three replicates
In contrast to the fast loss of microbial cell viability upon addition of silver ions to the cultures (Figs. 1, 2), the formation of AgNPs (Fig. 3) required several days at normal growth temperature (Fig. 4) of each microorganism where cells of all investigated microorganisms were killed by silver ions too early before formation of AgNPs. The bioreduction of silver ions to nanoparticles was strongly enhanced at high temperature (Fig. 4) which enhance the cell lyses as well as the bioreduction rate of silver ions to nanoparticles by the released cellular organics. The prolonged exposure to silver ions of all investigated microorganisms leads to the formation of silver particles in the nanorange (Fig. 3), indicating that the formation of AgNPs by the released cellular organics is not a specific metabolic process to a certain microorganism. The formed nanoparticles by E. coli, S. platensis and A. terreus (Fig. 3) showed a brown to reddish color in aqueous solution and showed a typical wavelength spectrum of AgNPs with a maximum wavelength of around 430 nm. The synthesized AgNPs by E. coli, S. platensis and A. terreus (Fig. 3) showed a size range of 5–30, 10–40 and 4–21 nm, respectively, as analyzed using transmission electron microscope (Fig. 3).

Transmission electron microscope (TEM) images of silver nanoparticles prepared by E. coli (a), S. platensis (b) and A. terreus (c). Histograms d, e and f are for the particle size frequency derived by counting over multiple TEM images of AgNPs prepared by E. coli, S. platensis and A. terreus, respectively

Temperature versus time during the synthesis of silver nanoparticles. The temperature and time required for the synthesis of silver nanoparticles (OD430 = 1) were recorded for E. coli (closed squares), S. platensis (open squares) and A. terreus (open cubes). The bars show the standard errors of three replicates
The cyanobacterium Anabaena variabilis ATCC 29413 grown photoautotrophically in liquid cultures in an eightfold dilution of the medium of Allen and Arnon (1955) also non-viably formed AgNPs (Fig. 5a) after several days of exposure to silver ions and showed similar results to S. platensis. Mixed phytoplankton collected from the Nile River as previously described (Morsy 2011c) and maintained in Bold’s basal medium (Bold 1949, Bischoff and Bold 1963) formed AgNPs (Fig. 5b) also after full death of cells and could not as viable cells produce these nanoparticles similar to S. platensis and A. variabilis. Average size of various AgNPs synthesized in this study as analyzed using TEM is shown in Table 1.

Transmission electron microscope (TEM) images of silver nanoparticles obtained by the bioreduction of silver ions to nanoparticles by Anabaena variables ATCC 29413 (a) or mixed phytoplankton collected from the Nile River (b). Histograms c and d are for the particle size frequency derived by counting over multiple TEM images of AgNPs prepared by Anabaena variables and mixed phytoplankton, respectively
To confirm that the biosynthesis of AgNPs by microorganisms is just a bioreduction process of silver ions by the organics released from the dead cultures, the synthesis of AgNPs (Fig. 6a) by cyanobacterial exopolysaccharides (EPS) of S. platensis cells was investigated. Various initial concentrations of silver nitrate (0.0 to 10 mM) were used to study the effect of silver ion concentration on AgNPs formation in the presence of 10 mg/ml polysaccharides (Fig. 6b), and the reaction was autoclaved at 15 psi and 121 °C for 5 min. The formation of AgNPs improved by increasing the initial concentration of silver salt (Fig. 6b). The effect of EPS concentration (0.0–14 mg/ml) on AgNPs formation was followed in the presence of 8 mM silver ions. Increasing the initial concentration of EPS increased the formation of AgNPs (Fig. 6c). The synthesized nanoparticles had a brown to reddish color in aqueous solution and showed a typical wavelength spectrum of AgNPs (Fig. 7) with a maximum wavelength of 430 nm. The synthesized AgNPs showed a size range of 8–39 nm as analyzed using transmission electron microscope (Fig. 6D; Table 1).

Bioreduction of silver ions to nanoparticles by exopolysaccharides (EPS) of S. platensis. TEM image (a). b The effect of silver ions concentration in the presence of 10 mg/ml EPS on AgNPs formation. The effect of EPS concentration on AgNPs formation was followed in the presence of 8 mM silver ions (c). The bars show the standard errors of three replicates. The histogram (d) is for the particle size frequency derived by counting over multiple TEM images of AgNPs prepared by EPS of S. platensis

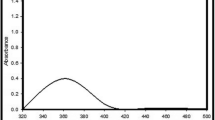
UV–visible spectrum of AgNPs prepared by the bioreduction of silver ions to nanoparticles by exopolysaccharides (EPS) of S. platensis
Minimum inhibitory concentration (MIC) and minimum bactericidal (MBC) and fungicidal (MFC) concentrations of silver ions versus nanoparticles were determined. The formed AgNPs by E. coli culture was used for investigating its bactericidal effect against the same microorganism where these nanoparticles showed antibacterial properties against that microorganism supporting that E. coli did not viably synthesize AgNPs (Fig. 8a). In addition, silver ions showed a more efficient antibacterial activity against E. coli (Fig. 8b) where the MIC and MBC of silver ions were significantly lower than AgNPs (Fig. 9). Similarly, AgNPs synthesized by other microorganisms were investigated as antimicrobial agent against all investigated microorganisms in this study. All AgNPs prepared in this study showed antimicrobial properties against all tested microorganisms even the same microorganism used for its synthesis (Table 2). Silver ions showed also antimicrobial properties against all tested microorganisms (Table 2). Both silver ions and nanoparticles showed a slight difference between the MIC and MBC or MFC against each microorganism (Table 2), suggesting that both act as a potent microbicidal agent against all subject microorganisms (E. coli, S. platensis and A. terreus). Although both silver ions and nanoparticles showed antimicrobial activities (Table 2), the minimum inhibitory concentration and minimum bactericidal and fungicidal concentrations of silver ions were significantly lower than AgNPs (Fig. 9; Table 2) indicating that silver ions are more efficient antimicrobial.

Determination of the minimum inhibitory concentration. The MIC of AgNPs (a) and silver ions (b) was determined by following the inhibitory effect on growth of E. coli as described in “Materials and methods”

Minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of silver ions versus nanoparticles against E. coli. The bars on columns show the standard errors of three replicates. Columns followed by the same letter are not significantly different at LSD (P < 0.05)
Discussion
Due to its wide range of applications and in spite of its antimicrobial properties, the biosynthesis of silver nanoparticles (AgNPs) by bacteria and other microorganisms became a hot topic of research in the recent two decades seeking a cheap mass scale industrial production of these nanoparticles. The results shown in this study reveal the controversy among literature on the biosynthesis of AgNPs by microorganisms and those describing the use of AgNPs as antimicrobial agent against similar organisms in other literature. Upon addition of silver ions to the cultures, all tested auto- and heterotrophic microorganisms in this study were killed and could not as viable cells produce AgNPs. Photosynthesis and respiration of the cyanobacterium S. platensis ceased by silver ions. Similar results were obtained for Anabaena variabilis ATCC 29413. Aerobic respiration of E. coli and A. terreus also ceased upon the addition of silver ions to the culture medium. Similar results were obtained for Epicoccum purpurascens and Aspergillus fumigatus. The formation of AgNPs by all investigated microorganisms at normal growth conditions was very slow process. Such formation of AgNPs occurs after full culture death and requires several days of incubation at normal growth temperature of all tested microorganisms. Mixed phytoplankton could not also as viable cells produce AgNPs and showed similar results to S. platensis and Anabaena variabilis. Such slow bioreduction was concomitant with the slow microbial cell lyses and release of organics to the culture medium. The bioreduction was strongly enhanced and occurs within few minutes at high temperature or autoclaving which enhance the cell lyses and the bioreduction rate of silver ions to nanoparticles by the released cellular organics.
The results presented in this study showed two similar antimicrobial characteristics of silver ions and nanoparticles. First, both silver ions and nanoparticles showed a broad-spectrum antimicrobial activities against all subjected microorganisms where all synthesized AgNPs in this study showed bactericidal and fungicidal properties even against microorganisms used for its synthesis supporting that the biosynthesis is not a vital metabolic process. Second, both silver ions and nanoparticles act as microbicides. As antimicrobials are usually regarded as bactericidal if the MBC is no more than four times the MIC (French 2006), the slight difference between the MIC and MBC or MFC of either silver ions or nanoparticles suggests that both of them act as potent microbicidal agents against all tested microorganisms. However, silver ions are more efficient as antimicrobial than its nanoparticles. Thus, the microbial cell death occurs by silver ions too early before formation of AgNPs.
Cell extracts of all investigated microorganisms could also reduce silver ions to form AgNPs at high temperature in a fast rate and at low rate at normal growth temperature of each microorganism. Even the cyanobacterial polysaccharides fraction extracted from S. platensis cells could efficiently reduce silver ions to nanoparticles in a fast rate at high temperature or autoclaving and at low rate at normal growth temperature of that microorganism. These results suggest that the biosynthesis of AgNPs by microorganisms is just a non-specific bioreduction process of silver ions into nanoparticles by the organics released in the microbial cell lysate from the dead cultures. Such bioreduction of silver ions into nanoparticles is similar to that bioreduction occurs by using many reported organic materials including pure and crude organics such as starch and plant extracts which have been successfully used for AgNPs formation (Raveendran et al. 2003; Vigneshwaran et al. 2006b; Mittal et al. 2013).
Although photoreduction of silver ions to nanoparticles was previously reported (Tan et al. 2007; Sato-Berrú et al. 2009), in this study, photoreduction of silver ions to nanoparticles did not occur in control experiments in light using SOT medium without S. platensis cells or pure EPS. This might be because SOT medium is inorganic, while photoreduction of silver ions to nanoparticles requires the presence of organic materials such as sodium citrate (Sato-Berrú et al. 2009) or a mixture of branched polyethyleneimine (BPEI) and 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) where the mechanism for the reduction of silver ions in the BPEI/HEPES mixtures entails oxidative cleavage of BPEI chains that results in the formation of positively charged BPEI fragments enriched with amide groups as well as in the production of formaldehyde, which serves as a reducing agent for silver ions (Tan et al. 2007). Control experiments were also conducted in dark to explore whether there is a role of photoreduction in AgNPs formation or not in the presence of S. platensis cells or EPS. In these control experiments conducted in dark using S. platensis at normal growth temperature of 32 °C, formation of AgNPs was also observed similar to light conditions indicating that neither light nor vitality of S. platensis cells was required for AgNPs formation. In addition, purified EPS also reduced silver ions to nanoparticles in dark at 32 °C confirming that organics are the bioreductant of silver ions to nanoparticles with no role of photoreduction or cell vitality. These results confirm that organics released from dead S. platensis cells reduce silver ions to nanoparticles.
It is well known that a microorganism that synthesize an antibiotic or a toxin has a mechanism for protecting itself from the microbicidal action of such substances. In contrast to biologically synthesized antibiotics, AgNPs act as antimicrobial and a powerful microbicide for even the microbes that have been hypothesized to biosynthesize these nanoparticles. Unlike iron magnetic nanoparticles in magnetotactic bacteria, it is not reported that intracellular biosynthesis of AgNPs is a natural phenomenon in any microorganism. Besides, a natural intracellular biosynthesis of AgNPs would require a mechanism for active influx and accumulation of silver ions inside the cell. The magnetotactic bacteria that in vivo synthesize magnetite nanoparticles in magnetosomes are believed to synthesize siderophores (Bazylinski and Frankel 2004) which have a high affinity for Fe3+ to be transferred to inside the cell and then to be reduced to Fe2+ and accumulated in magnetosomes by active iron transport proteins (Nakamura et al. 1995; Schübbe et al. 2003) where magnetite (or greigite) nanocrystals are formed. In contrast, no gene in currently sequenced microbial genomes is functionally confirmed by knockout as encoding for silver ion influx transporter in microorganisms. A survey in NCBI (National Center for Biotechnology Information) Web site and all sequenced genomes revealed nothing for a gene expressing a biochemically or physiologically characterized silver ion influx transporter in any of the current genome-sequenced microorganisms. Unlike iron, silver is not considered as a mineral macro-nutrient or even micro-nutrient for microorganisms and is not actively accumulated for a natural phenomenon. One better alternative to microorganisms for the biosynthesis of AgNPs is the plant extracts which are easily available, safe and have a broad variety of metabolites that can aid in the reduction of silver ions faster than microorganisms in the synthesis. The use of microbial cell extracts (Morsy 2014) or fractions such as cyanobacterial polysaccharides which can be massively prepared (Morsy et al. 2008b) for the bioreduction of silver ions to nanoparticles (Morsy et al. 2014) at high temperature would also be economically feasible for industrial-scale preparations of AgNPs.
References
Abd-Alla MH, Morsy FM, El-Enany AE (2011) Hydrogen production from rotten dates by sequential three stages fermentation. Int J Hydrogen Energy 36:13518–13527
Allen MB, Arnon DI (1955) Studies on nitrogen-fixing blue-green algae. I. Growth and nitrogen fixation by Anabaena cylindrica Lemm. Plant Physiol 30:366–372
Anil-Kumar S, Abyaneh MK, Gosavi Sulabha SW, Ahmad A, Khan MI (2007) Nitrate reductase-mediated synthesis of silver nanoparticles from AgNO3. Biotechnol Lett 29:439–445
Bazylinski DA, Frankel RB (2004) Magnetosome formation in prokaryotes. Nat Rev Microbiol 2:217–230
Bischoff H, Bold HC (1963) Phycological studies. IV. Some soil algae from enchanted rock and related algal species. University of Texas Publication No. 6318, 95 pp
Bold HC (1949) The morphology of Chlamydomonas chlamydogama, sp. nov. Bull Torrey Bot Club 76:101–108
Brandt O, Mildner M, Egger AE, Groessl M, Rix U, Posch M, Keppler BK, Strupp C, Mueller B, Stingl G (2012) Nanoscalic silver possesses broad-spectrum antimicrobial activities and exhibits fewer toxicological side effects than silver sulfadiazine. Nanomed Nanotech Biol Med 8(4):478–488
Castro-Longoria E, Vilchis-Nestor A, Avales-Borja M (2011) Biosynthesis of silver, gold, and bimetallic nanoparticles using the filamentous fungus Neurospora crassa. Colloids Surf B Biointerfaces 83:42–48
De Gusseme B, Sintubin L, Baert L, Thibo E, Hennebel T, Vermeulen G, Uyttendaele M, Verstraete W, Boon N (2010) Biogenic silver for disinfection of water contaminated with viruses. Appl Environ Microbiol 76:1082–1087
Durán N, Marcato PD, Alves OL, De Souza GIH, Esposito E (2005) Mechanistic aspects of biosynthesis of silver nanoparticles by several Fusarium oxysporum strains. J Nanobiotechnol 3:8
Eckhardt S, Brunetto PS, Gagnon J, Priebe M, Giese B, Fromm KM (2013) Nanobio silver: its interactions with peptides and bacteria, and its uses in medicine. Chem Rev 113(7):4708–4754
Feng QL, Wu J, Chen GQ, Cui FZ, Kim TN, Kim JO (2000) A mechanistic study of the antibacterial effect of silver ions on Escherichia coli and Staphylococcus aureus. J Biomed Mater Res 52:662–668
Filali-Mouhim R, Cornet JF, Fontaine T, Fournet B, Dubertret G (1993) Production, isolation and preliminary characterization of the exopolysaccharide of the cyanobacterium Spirulina platensis. Biotechnol Lett 15:567–572
French GL (2006) Bactericidal agents in the treatment of MRSA infections—the potential role of daptomycin. J Antimicrob Chemother 58:1107–1117
Govindaraju K, Tamilselvan S, Kiruthiga V, Singaravelu G (2011) Silver nanotherapy on the viral borne disease of silkworm Bombyx mori L. J Nanopart Res 13:6377–6388
Gurunathan S, Kalishwaralal K, Vaidyanathan R, Venkataraman D, Pandian SR, Muniyandi J, Hariharan N, Eom SH (2009) Biosynthesis, purification and characterization of silver nanoparticles using Escherichia coli. Colloids Surf B Biointerfaces 74:328–335
Holden M (1976) In: Goodwin TW (ed) Chlorophylls in chemistry and biochemistry of plant pigments, 2nd edn, vol 2. Academic Press, London, pp 1–37
Jain N, Bhargava A, Majumdar S, Tarafdar J, Panwar J (2011) Extracellular biosynthesis and characterization of silver nanoparticles using Aspergillus flavus NJP08: a mechanism perspective. Nanoscale 3(2):635–641
Kalimuthu K, Babu RS, Venkataraman D, Mohd B, Gurunathan S (2008) Biosynthesis of silver nanocrystals by Bacillus licheniformis. Colloids Surf B Biointerfaces 65:150–153
Kim JS, Kuk E, Yu KN, Kim JH, Park SJ, Lee HJ, Kim SH, Park YK, Park YH, Hwang C-Y, Kim YK, Lee Y-S, Jeong DH, Cho M-H (2007) Antimicrobial effects of silver nanoparticles. Nanomed Nanotech Biol Med 3(1):95–101
Kim SW, Jung JH, Lamsal K, Min JS, Lee YS (2012) Antifungal effects of silver nanoparticles (AgNPs) against various plant pathogenic fungi. Mycobiol 40(1):53–58
Klaus T, Joerger R, Olsson E, Granqvist CG (1999) Silver-based crystalline nanoparticles, microbially fabricated. Proc Natl Acad Sci USA 96:13611–13614
Klaus T, Joerger R, Olsson E, Granqvist CG (2001) Bacteria as workers in the living factory: metal-accumulating bacteria and their potential for materials science. Trend Biotechnol 19(1):15–20
Koutb M, Morsy FM (2011) A potent lipid producing isolate of Epicoccum purpurascens AUMC5615 and its promising use for biodiesel production. Biomass Bioenergy 35:3182–3187
Lara HH, Garza-Treviño EN, Ixtepan-Turrent L, Singh DK (2011) Silver nanoparticles are broad-spectrum bactericidal and virucidal compounds. J Nanobiotech 9:30
Law N, Ansari S, Livens FR, Renshaw JC, Lloyd JR (2008) The formation of nano-scale elemental silver particles via enzymatic reduction by Geobacter sulfurreducens. Appl Environ Microbiol 74:7090–7093
Lengke M, Fleet M, Southam G (2006) Biosynthesis of silver nanoparticles by filamentous cyanobacteria from a silver(I) nitrate complex. Langmuir 10:1021–1030
Li WR, Xie XB, Shi QS, Zeng HY, Ou-Yang YS, Chen YB (2010) Antibacterial activity and mechanism of silver nanoparticles on Escherichia coli. Appl Microbiol Biotechnol 85(4):1115–1122
Lok C-N, Zou T, Zhang J-J, Lin IW-S, Che C-M (2014) Controlled-release systems for metal-based nanomedicine: encapsulated/self-assembled nanoparticles of anticancer gold(III)/platinum(II) complexes and antimicrobial silver nanoparticles. Adv Mater 26:5550–5557
Mandal D, Bolander ME, Mukhopadhyay D, Sarkar G, Mukherjee P (2006) The use of microorganisms for the formation of metal nanoparticles and their application. Appl Microbiol Biotechnol 69:485–492
Mittal AK, Chisti Y, Banerjee UC (2013) Synthesis of metallic nanoparticles using plant extracts. Biotechnol Adv 31:346–356
Mohanty S, Mishra S, Jena P, Jacob B, Sarkar B, Sonawane A (2012) An investigation on the antibacterial, cytotoxic, and antibiofilm efficacy of starch-stabilized silver nanoparticles. Nanomed Nanotech Biol Med 8(6):916–924
Morsy FM (2011a) Hydrogen production from acid hydrolyzed molasses by the hydrogen overproducing Escherichia coli strain HD701 and subsequent use of the waste bacterial biomass for biosorption of Cd(II) and Zn(II). Int J Hydrog Energy 36:14381–14390
Morsy FM (2011b) Acetate versus sulfur deprivation role in creating anaerobiosis in light for hydrogen production by Chlamydomonas reinhardtii and Spirulina platensis: two different organisms and two different mechanisms. Photochem Photobiol 87(1):137–142
Morsy FM (2011c) A simple approach to water and plankton sampling for water microbiological and physicochemical characterizations at various depths in aquatic ecosystems. Ann Limnol 47:65–71
Morsy FM (2014) Hydrogen production by Escherichia coli without nitrogen sparging and subsequent use of the waste culture for fast mass scale one-pot green synthesis of silver nanoparticles. Int J Hydrog Energy 39:11902–11912
Morsy FM, Nakajima M, Yoshida T, Fujiwara T, Sakamoto T, Wada K (2008a) Subcellular localization of ferredoxin-NADP+ oxidoreductase in phycobilisome retaining oxygenic photosynthetic organisms. Photosynth Res 95(1):73–85
Morsy FM, Kozuha S, Takani Y, Sakamoto T (2008b) Novel thermostable glycosidases in the extracellular matrix of the terrestrial cyanobacterium Nostoc commune. J Gen Appl Microbiol 54:243–252
Morsy FM, Nafady NA, Abd-Alla MH, Abd-Elhady D (2014) Green synthesis of silver nanoparticles by water soluble fraction of the extracellular polysaccharides/matrix of the cyanobacterium Nostoc commune and its application as a potent fungal surface sterilizing agent of seed crops. Univ J Microbiol Res 2(2):36–43
Moss ES, McQuown AL (1969) Atlas of medical mycology, 3rd edn. Williams & Wilkins Co, Baltimore
Mukherjee P, Ahmad A, Mandal D, Senapati S, Sainkar S, Khan M, Parishcha R, Ajaykumar P, Alam M, Kumar R, Sastry M (2001) Fungus-mediated synthesis of silver nanoparticles and their immobilization in the mycelial matrix; a novel biological approach to nanoparticle synthesis. Nano Lett 1(10):515–519
Nakamura C, Kikuchi T, Burgess JG, Matsunaga T (1995) An iron-regulated gene, magA, encoding an iron transport protein of Magnetospirillum sp. strain AMB-1. J Biol Chem 270:28392–28396
Naveen H, Kumar G, Karthik L, Roa B (2010) Extracellular biosynthesis of silver nanoparticles using the filamentous fungus Penicillium sp. Arch App Sci Res 2:161–167
Ogawa T, Terui G (1970) Studies on the growth of Spirulina platensis I on the pure culture of Spirulina platensis. J Ferment Technol 48:361–367
Pal S, Tak YK, Song JM (2007) Does the antibacterial activity of silver nanoparticles depend on the shape of the nanoparticle? A study of the Gram-negative bacterium Escherichia coli. Appl Environ Microbiol 73:1712–1720
Penfold DW, Forster CF, Macaskie LE (2003) Increased hydrogen production by Escherichia coli strain HD701 in comparison with the wild-type parent strain MC4100. Enzyme Microb Technol 33:185–189
Rai M, Yadav A, Gade A (2009) Silver nanoparticles as a new generation of antimicrobials. Biotechnol Advan 27(1):76–83
Raveendran P, Fu J, Wallen SL (2003) Completely “green” synthesis and stabilization of metal nanoparticles. J Am Chem Soc 125:13940–13941
Samadi N, Golkaran D, Eslamifar A, Jamalifar H, Fazeli MR, Mohseni FA (2009) Intra/extracellular biosynthesis of silver nanoparticles by an autochthonous strain of Proteus mirabilis isolated from photographic waste. J Biomed Nanotechnol 5(3):247–253
Sato-Berrú R, Redón R, Vázquez-Olmos A, Saniger JM (2009) Silver nanoparticles synthesized by direct photoreduction of metal salts. Application in surface-enhanced Raman spectroscopy. J Raman Spectrosc 40:376–380
Sauter M, Bohm R, Bock A (1992) Mutational analysis of the operon (hyc) determining hydrogenase 3 formation in Escherichia coli. Mol Microbiol 6:1523–1532
Schübbe S, Kube M, Scheffel A, Wawer C, Heyen U, Meyerdierks A, Madkour MH, Mayer F, Reinhardt R, Schüler D (2003) Characterization of a spontaneous nonmagnetic mutant of Magnetospirillum gryphiswaldense reveals a large deletion comprising a putative magnetosome island. J Bacteriol 185:5779–5790
Sharma VK, Yngard RA, Lin Y (2009) Silver nanoparticles: green synthesis and their antimicrobial activities. Adv Colloid Interface Sci 145(1–2):83–96
Sintubin L, De Windt W, Dick J, Mast J, Ha DV, Verstraete W, Boon N (2009) Lactic acid bacteria as reducing and capping agent for the fast and efficient production of silver nanoparticles. Appl Microbiol Biotechnol 84(4):741–749
Slawson RM, Van Dyke MI, Lee H, Trevors JT (1992) Germanium and silver resistance, accumulation, and toxicity in microorganisms. Plasmid 27:72–79
Tamboli DP, Lee DS (2013) Mechanistic antimicrobial approach of extracellularly synthesized silver nanoparticles against gram positive and gram negative bacteria. J Hazard Mater 260:878–884
Tan S, Erol M, Attygalle A, Du H, Sukhishvili S (2007) Synthesis of positively charged silver nanoparticles via photoreduction of AgNO3 in branched polyethyleneimine/HEPES solutions. Langmuir 23:9836–9843
Thakkar KN, Mhatre SS, Parikh RY (2010) Biological synthesis of metallic nanoparticles. Nanomed Nanotech Biol Med 6:257–262
Vigneshwaran N, Kathe AA, Varadarajan PV, Nachane RP, Balasubramanya RH (2006a) Biomimetics of silver nanoparticles by white rot fungus, Phaenerochaete chrysosporium. Colloids Surf B Biointerfaces 53:55–59
Vigneshwaran N, Nachane RP, Balasubramanya RH, Varadarajan PV (2006b) A novel one-pot ‘green’ synthesis of stable silver nanoparticles using soluble starch. Carbohydr Res 341:2012–2018
Yamanaka M, Hara K, Kudo J (2005) Bactericidal actions of a silver ion solution on Escherichia coli, studied by energy-filtering transmission electron microscopy and proteomic analysis. Appl Environ Microbiol 71(11):7589–7593
Acknowledgments
Sincere thanks to Professor Dr. L.E. Macaskie and Dr. Mark Redwood (School of Biosciences, University of Birmingham, Edgbaston, Birmingham, UK) for kindly providing E. coli HD701. Sincere thanks to Emeritus Prof. Dr. Keishiro Wada and Associate Professor Dr. Toshio Sakamoto (Division of Life Science, Graduate School of Natural Science and Technology, Kanazawa University, Kakuma, Kanazawa, Ishikawa, Japan) for their encouragements for study about cyanobacteria and the distinctive features and applications of cyanobacterial polysaccharides. The author is grateful to the precious suggestions and careful corrections made by the anonymous reviewers for further improvements of this paper.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Erko Stackebrandt.
Rights and permissions
About this article

Cite this article
Morsy, F.M. Toward revealing the controversy of bacterial biosynthesis versus bactericidal properties of silver nanoparticles (AgNPs): bacteria and other microorganisms do not per se viably synthesize AgNPs. Arch Microbiol 197, 645–655 (2015). https://doi.org/10.1007/s00203-015-1098-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00203-015-1098-z