
Abstract
Paddy rice is a typical wetland plant species, and mercury (Hg) accumulation in this rice has received much attention over the last two decades. The role of root iron plaque on rice Hg accumulation is not well understood. The effects of iron plaque on Hg0 uptake, translocation, and volatilization in rice seedlings were investigated under hydroponic conditions using different rice genotypes. After induction of iron plaque on rice roots with pretreatment solutions containing 0, 15 and 30 mg Fe2+L−1, rice seedlings were transplanted into specially designed airtight culture chambers, where roots were separated from the aerial parts and exposed to saturated Hg0 vapor. The results showed the following: (1) There were significant differences in the amount of iron plaque formed on the rice roots among the three genotypes. (2) A significant correlation was observed between the concentrations of Hg and Fe in the iron plaque of the root surface for the three genotypes (R2 = 0.933, p < 0.01). (3) Iron plaque may act as a barrier for Hg0 behavior, i.e., inhibiting the process of Hg0 uptake and translocation from the rhizosphere.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Mercury (Hg) is listed as a priority pollutant by many international agencies because of its persistence, bioaccumulation, and toxicity in the environment (Jiang et al. 2006). It exists in nature in three forms, i.e., elemental (or metallic) mercury, inorganic mercury compounds, and organic mercury compounds. Among all forms of Hg, its organic form, methylmercury (MeHg), is a neurotoxin with high bioaccumulation potential in the food chain and has thus attracted concern from the public (Elsayed et al. 2020). In previous studies, fish consumption was recognized as the main pathway of human MeHg exposure (Hong et al. 2012). However, at the beginning of this century, rice grains were found to bioaccumulate MeHg in Hg-contaminated areas, causing local exposure of MeHg to the population (Horvat et al. 2003; Zhang et al. 2010). Enriched Hg isotopes were artificially added to paddy soil or ambient air to trace the contributions of Hg from the soil or atmosphere (Strickman and Mitchell 2017). Studies have shown that paddy soil is the main source of MeHg in rice tissue, whereas rice plants receive inorganic mercury (IHg) both from the soil and the atmosphere (Meng et al. 2010, 2014). Due to contamination from Hg mining and smelting activities, as well as the discharge of industrial effluent and wastewater irrigation in agriculture, Hg pollution of soils and crops is becoming a global problem (Zagar et al. 2014). In recent years, Hg pollution incidents in rice have been frequently reported. For example, Zhang et al. (2010) found that the contents of IHg and MeHg in rice grain were 94 (7.4 ~ 460) μg/kg and 11 (1.2 ~ 44) μg/kg, respectively, in a Hg mining area, which is higher than the limit of the Chinese national standards for cereal (20 μg/kg, GB2762-2005). To reduce Hg accumulation in rice, many studies have been conducted on the processes and mechanisms of the uptake and translocation of IHg and MeHg in the soil‒rice system (Zhang et al. 2010; Strickman and Mitchell 2017). As a typical wetland plant species, the well-developed aerenchyma formed in rice roots promotes O2 transportation from the atmosphere to the rhizosphere. The released O2 and other oxidizing substances form an oxidizing environment in the root zone and finally induce the precipitation of the oxides of Fe and Mn on the root surface, thus forming a reddish-brown iron plaque on the root surface (Fu et al. 2010; Wang et al. 2022). It has been shown that iron plaque induced on the root surface can act as a barrier that significantly decreases the uptake and translocation of both IHg and MeHg in rice, which is an important regulatory mechanism to reduce Hg accumulation in rice plants (Li et al. 2014, 2015).
Gaseous elemental mercury (Hg0) is usually present in the atmosphere. Using stable isotope tracer techniques, numerous studies have shown that the atmosphere is the principal source of Hg to the aboveground parts of rice plants through direct stomatal uptake of Hg0 (Yin et al. 2013; Strickman and Mitchell 2017; Qin et al. 2020). Moreover, Hg concentrations in rice grains increase with Hg0 levels in air (Tang et al. 2021), which will directly pose human exposure problems. However, previous studies on Hg0 were mostly limited to the plant‒atmosphere system. For wetland habitats, redox reactions can occur by aquatic microorganisms, which favors the production of Hg0. Bouffard and Amyot (2009) showed that under anaerobic conditions, the reduction of Hg(II) in sediments driven by microorganisms and other abiotic factors led to Hg0 concentrations in sediments accounting for 7.4%–28.4% of the total Hg concentration, indicating that Hg0 is also a very important form of Hg in wetland ecosystems. The seasonal irrigation of rice during its growth period makes the paddy field a unique wetland ecosystem. Our previous research showed that the aerenchyma of rice roots could play an important role in affecting Hg0 uptake by rice (Shang et al. 2017). However, the factors affecting Hg0 uptake, translocation and volatilization by rice have not been well studied. Does the iron plaque act as a barrier that inhibits the process of Hg0 uptake, translocation, and volatilization from the rhizosphere to the air? Does the iron plaque amount affect this process? This work aimed to investigate the effect of different amounts of iron plaque on the Hg0 process mentioned above. In addition, the differences among genotypes affecting the formation of iron plaque and Hg0 uptake, translocation and volatilization were also investigated. This study will provide a scientific basis for ensuring the food safety of rice in mercury-contaminated areas and contribute to a better understanding of the Hg biogeochemical cycle in wetland ecosystems.
Materials and Methods
Seeds of three rice (Oryza sativa L.) cultivars, including IIYou 1259 (IIY), Tiyou 267 (TU267) and Wuyunjing 21 (WYJ21), were used in this study. Cultivar IIY belongs to indica hybrid rice, while cultivars TU267 and WYJ21 belong to japonica rice. Seeds were sterilized in 10% H2O2 (v/v) diluted with 30% H2O2 solution for 20 min and washed thoroughly with distilled water. They were then germinated on wet filter papers in Petri dishes and placed in a constant-temperature incubator at 28°C for 3 days. After transferring seedlings to sand medium for 5 days, uniform seedlings were selected and transported to PVC pots (5 L, one plant per pot) containing Kimura B nutrient solution. The nutrient solution pH value was adjusted to 5.5 using 0.1 M KOH or HCl and renewed every 3 days. All pots were placed in a controlled environment growth chamber with a 14 h light period (260 µmol·(m2·s)−1) and a 10 h dark period. The temperature was kept at 28°C during the day and 22°C during the night. The relative humidity was 70%. Iron plaque was induced on the roots after 30 days of cultivation with Kimura B nutrient solution. Briefly, uniform seedlings were selected, and the whole root systems were put into a 0.5 mmol·L−1 CaCl2 solution for 2 h and then transferred into a solution with 0, 15 or 30 mg·L−1 of ferrous ions for 24 h (Fe2+ as FeSO4·7H2O). These treatments were referred to as Fe0, Fe15 and Fe30, respectively. The solution pH was adjusted to 5.5 using 0.1 M KOH or HCl. Seedlings were subsequently grown again in Kimura B nutrient solution (pH 5.5) for 2 d before exposure to Hg0.
The effects of root iron plaque on Hg0 volatilization were examined using a specially designed airtight chamber after iron plaque was induced on rice root surfaces. The device was customized with high-temperature resistant silicon boride material and was divided into two parts, the upper leaf chamber and the lower root chamber (Fig. 1). After the rice roots were washed with distilled water, the junction of the roots and shoots was placed in a silica gel plug and then inserted into a hole with a diameter of 4 cm between the upper and lower air chambers. After 3 g of elemental Hg was put into the lower root chamber (the Hg vapor concentration was approximately 0.0215 g·m−3 calculated according to the saturated vapor pressure of Hg at 27°C), the inflation pump was used to provide a constant airflow for the upper leaf chamber with a flow rate of 200 mL·min−1 through the float flowmeter. Hg in the airflow was absorbed by one Hg sorbent trap. The Hg0 volatilized from the leaves reached the Hg sorbent trap (Hg adsorbent) 1 and 2 with the airflow. Roots were exposed to saturated Hg0 for 2.5 h. Each exposure treatment was performed in triplicate. All treatments were performed under the same conditions, i.e., room temperature (20–25°C) and indoor light.

Schematics of the Hg0 volatilization experiment using rice seedlings
Iron plaque on fresh root surfaces was extracted using the dithionite-citrate-bicarbonate (DCB) method (Taylor and Crowder 1983). This solution is composed of 0.03 mol·L−1 sodium citrate (Na3C6H507·2H20) and 0.125 mol·L−1 sodium bicarbonate (NaHCO3), with the addition of 0.6 g sodium dithionite (Na2S2O4). The entire root system of each rice seedling was incubated in 60 mL of DCB solution for 60 min and rinsed three times with deionized water. The rinse was then added to the extract from the same sample, and the combined sample was made up to 100 mL with deionized water. The Fe concentrations in the extractant were determined directly by ICP‒OES. In addition, the rice seedlings (roots and shoots) used for Hg0 volatilization experiments were freeze-dried after DCB extraction and weighed for biomass.
Hg concentrations in DCB extracts, roots, shoots, and Hg adsorbents were determined by a DMA-80 direct mercury analyzer (Milestone, Italy). After thermal decomposition, catalytic conversion, and Hg homogenization, the total Hg concentration in each sample was measured by atomic absorption spectrophotometry at 253.7 nm. The certified reference materials of citrus leaves (GBW10020, China) were measured for quality control, and the recoveries ranged from 88.2% to 110.6%, with RSD < 10%.
The data were analyzed using repeated measures analysis of variance (ANOVA) at a significance level of 0.05. If there was a statistically significant difference between groups, pairwise comparisons were further performed by the Bonferroni method with the significance level set to 0.05/n (n = 3). All statistical analyses were conducted using the statistical software R version 3.4.3.
Results
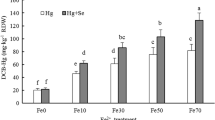
Compared with the control (Fe0), the iron plaque was visible as a reddish-brown coating on the root surface after pretreatment with Fe2+. The concentrations of Fe (DCB-Fe) and Hg (DCB-Hg) in DCB extracts of three rice genotypes are shown in Fig. 2. The contents of Fe in the iron plaque formed on rice roots were significantly different among genotypes (p < 0.05, Fig. 2a). The amount of iron plaque in Indica rice was significantly higher than that in Japonica rice. The amount of iron plaque in genotype IIY was 2.35 and 2.01 times that of genotype TU267 and 3.55 and 3.68 times that of genotype WYJ21 under the Fe15 and Fe30 treatments, respectively. The Fe levels in the iron plaque increased with increasing Fe2+ concentrations in the pretreatment solutions for the three genotypes (Fig. 2a). For genotype TU267, the amount of iron plaque differed significantly between Fe15 and Fe30 (p < 0.05). For IIY and WYJ21, although the Fe2+ concentration in the pretreatment solution of Fe30 was higher, the amount of iron plaque did not significantly increase compared to that of Fe15. The content of total Hg involved in the interaction between Hg0 and iron plaque was investigated by measuring the Hg concentrations in the DCB extracts. The results are shown in Fig. 2b. The pretreatment solution of Fe15 and Fe30 significantly increased Hg concentrations in DCB extracts of three rice genotypes compared with the Fe0 treatment (Fe0, p < 0.05, Fig. 2b). Hg concentrations in DCB extracts correlated significantly with Fe levels in iron plaque for the three genotypes (R2 = 0.933, p < 0.01, Fig. 3).

Concentrations of Fe(a) and Hg(b) in DCB extracts of different pretreatments. Plants were pretreated in solutions with different concentrations of Fe2+ (Fe0, Fe15, and Fe30: 0, 15, and 30 mg Fe2+ L-1 solutions) for 24 h. Different lowercase letters refer to significant differences at the p < 0.05 level between genotypes, and different capital letters refer to significant differences at the p < 0.05 level between different pretreatments

Correlation between concentrations of Fe and Hg in DCB extracts
The concentrations of Fe2+ in the pretreatment solutions had a significant effect on the Hg concentrations in the roots and shoots for all cultivars. Compared with Fe0, the Hg concentrations in the roots and shoots of the three genotypes decreased significantly under Fe15 and Fe30 (p < 0.05, Fig. 4). Under the pretreatment of Fe15 and Fe30, compared with Fe0, Hg concentrations in roots of genotype IIY decreased by 25.8% and 38.7%, genotype TU267 decreased by 35.6% and 61.5%, and genotype WYJ21 decreased by 67.6% and 74.5%, respectively. Similarly, compared with Fe0, Hg concentrations in shoots of genotype IIY decreased by 42.9% and 55.3%, genotype TU267 decreased by 31.7% and 47.8%, and genotype WYJ21 decreased by 44.2% and 57.1%, respectively.

Concentrations of Hg in shoots (a) and roots (b) of different pretreatments
With the increase in Fe2+ concentrations in the pretreatment solutions, Hg0 volatilization by leaves of the three genotypes decreased significantly (p < 0.05). Under the pretreatment of Fe15 and Fe30, compared with Fe0, the Hg0 volatilization from leaves of genotype IIY decreased by 34.2% and 46.9%, that of genotype TU267 decreased by 27.1% and 44.2%, and that of genotype WYJ21 decreased by 23.8% and 36.9%, respectively. In addition, there was no significant difference in Hg0 volatilization by leaves among the three genotypes (Fig. 5).

Hg0 volatilization of rice shoots after different pretreatments
Discussion
The results showed that the amounts of iron plaque induced on the root surfaces were significantly different among genotypes, which was consistent with the results obtained by Liu et al. (2014), and in this study, the results showed that the amounts of iron plaque on root surfaces of 14 rice genotypes varied by 2.6 times (Liu et al. 2014). The iron plaque amounts may correlate with the root surface area. Deng et al. (2010) showed that treated roots with lower radial oxygen loss formed more iron plaque, mainly because the larger surface area provided a larger adsorption area for Fe2+, thereby increasing the amount of iron plaque. The root surface areas of the three rice genotypes were analyzed in our previously published article (Shang et al. 2017). The results showed that the root surface area of genotype IIY was the largest (31.70 cm2), while that of genotype WYJ21 was the smallest (14.78 cm2). In the present study, our results demonstrated that the amount of iron plaque induced on the root surface of genotype IIY was the highest, while genotype WYJ21 was the lowest (Fig. 2a), which was consistent with the change in root surface area, indicating that the difference in the amount of iron plaque induced on the root surface caused by genotype may be mainly affected by root surface area. Thus, the amount of iron plaque increased with increasing Fe2+ concentrations in the pretreatment solution, as reflected by the similar amount of iron plaque under the Fe15 and Fe30 treatments (Fig. 2a). However, for genotype TU267, the amount of iron plaque varied significantly by pretreatment with the three Fe2+ solutions, i.e., Fe0, Fe15, and Fe30 (Fig. 2a), indicating that iron plaque could be affected by other factors. From the mechanism of iron plaque formation, the difference in the amounts of iron plaque is associated with the oxidizing power of rice roots of different genotypes, which could be mainly determined by the structure of rice root aerenchyma and two components, i.e., oxygen release and enzymatic oxidation (Ando et al. 1983). Meng et al. (2008) showed that there was a positive correlation between the aerenchyma (characterized by porosity, %) of rice roots and the amounts of iron plaque induced on the root surface. Thus, other factors, such as rice root aerenchyma and the related effects of oxygen release and enzymatic oxidation, need to be further explored.
Studies have shown that iron plaque exhibits high affinities for anions and cations due to the high absorption capacity generated by the large specific surface area of iron plaque (generally more than 200 m2/g) (Trivedi and Axe 2001) and special electrical properties of oxides (such as iron oxide, goethite, and squamous carbon) contained in iron plaque. Thus, iron plaque may change the distribution ratio of ions in the solid‒liquid phase through adsorption and coprecipitation (de Araujo et al. 2020), thereby affecting the uptake and translocation of these ions by plants. We found that the iron plaque has a certain adsorption capacity for Hg0 (Fig. 2a). The physisorption (by van der Waals force bonding) may be formed between Hg0 and the iron plaque (Wu et al. 2007). Compared to the chemisorption of ionic compounds with iron plaque, this effect could be much weaker. Additionally, the formation of iron plaque transferred Hg0 to Hg cations, as revealed by the detected Hg in DCB extraction. Furthermore, the DCB-extracted Hg correlated significantly with the amounts of iron plaque for all three genotypes (Fig. 3), which indicated that the significant difference in the amount of Hg0 adsorbed and transformation by iron plaque may be largely due to the varied ability of iron plaque formation among genotypes.
Our results showed that the formation of iron plaque significantly reduced the Hg content in roots (Fig. 4b), which is consistent with some previous findings on inorganic Hg (Li et al., 2014) and other ionic compounds (Liu et al. 2004, Hu et al. 2013; Peng et al. 2018). The results suggested that the formation of iron plaque may act as a barrier for Hg0 behavior, i.e., inhibiting the process of Hg0 uptake from the rhizosphere. Within a similar amount of iron plaque, we found that the adsorption capacity of iron plaque for Hg0 was low (1.45 ~ 15.69 mg/kg Fig. 2b) in comparison with previous studies on the adsorption capacity of iron plaque for other metal ions (Deng et al. 2010; Hu et al. 2013; Peng et al. 2018) or other forms of Hg (all > 100 mg/kg) (Li et al. 2014, 2015). Although the adsorption capacity of iron plaque for Hg0 was low, it led to a significant decrease in the uptake of Hg0 by roots (p < 0.05, Fig. 4b). Compared with Fe0 without iron plaque, the Hg content in rice roots with iron plaque decreased by a maximum of 74.5% in Fe30 (Fig. 4b), suggesting that iron plaque could play a key role in inhibiting Hg0 uptake by roots. We speculate that the adsorption of Hg0 and its transferred Hg cations by iron plaque may be one of the reasons for the reduction in the Hg content in roots. In addition, the barrier effect of iron plaque on Hg0 uptake by rice roots may also be due to the encasement of rice root systems by iron plaque colloids, which reduces the gas permeability of the rice root surface as well as the contact between rice root cells and the external environment, thereby inhibiting Hg0 uptake by rice roots.
In the present study, the formation of iron plaque significantly reduced the Hg content in shoots (Fig. 4a). For the same genotype of rice, the differential changes in Hg concentrations in shoots and roots under pretreatment with three concentrations of Fe2+ solution showed similar decreasing trends (Fig. 4), which indicates that the Hg concentration in roots is an important factor affecting the Hg concentration in shoots. The translocation factor (TF) was used as an indicator to evaluate the transfer ability of Hg from the roots to aboveground parts in rice (TF = Tshoot-Hg/Troot-Hg, Tshoot-Hg and Troot-Hg represent the Hg content in the shoots and roots, respectively) (Table 1). The results indicated that the formation of iron plaque effectively inhibited the translocation of Hg0 from the roots to the shoots (Table 1). Therefore, we speculate that the barrier effect of iron plaque on Hg0 uptake by the roots leads to a decrease in Hg content in the roots and, in turn, to a decrease in the translocation of Hg to the aboveground part.
Our results showed that increasing amounts of iron plaque significantly decreased Hg0 volatilization by rice (Fig. 5), which was consistent with the variation pattern of Hg concentration in shoots. Therefore, we calculated the aboveground Hg0 volatilization rate (%) under different pretreatments (Table 2), that is, the proportion of Hg0 volatilization in the total Hg uptake in the aboveground part. The results showed that the aboveground Hg0 volatilization rate was much lower than 0.1%, indicating that only a very small part of Hg can be released from the rice leaf. In addition, there was no significant difference in the Hg0 volatilization rate under different pretreatments relative to the control (p > 0.05), which indicated that the formation of iron plaque had little effect on the volatilization process of Hg0 from shoots to air.
Our results showed that the formation of iron plaque on rice roots was influenced by the rice genotype. The formed iron plaque on the rice root surface had a certain adsorption capacity for Hg0, thereby reducing the Hg content in roots and shoots. The large difference in the amount of Hg0 adsorbed by iron plaque may be largely due to the varied ability of iron plaque formation among genotypes. Overall, the introduction of iron plaque could act as a barrier that inhibits the process of Hg0 uptake and translocation by rice.
References
Ando T, Yoshida S, Nishiyama I (1983) Nature of oxidizing power of rice roots. Plant Soil 72:57–71. https://doi.org/10.1007/bf02185094
Bouffard A, Amyot M (2009) Importance of elemental mercury in lake sediments. Chemosphere 74:1098–1103. https://doi.org/10.1016/j.chemosphere.2008.10.045
de Araujo T, Isaure M, Alchoubassi G et al (2020) Paspalum urvillei and Setaria parviflora, two grasses naturally ada-pted to extreme iron-rich environments. Plant Physiol Biochem 151:144–156. https://doi.org/10.1016/j.plaphy.2020.03.014
Deng D, Wu S, Wu F et al (2010) Effects of root anatomy and Fe plaque on arsenic uptake by rice seedlings grown in solution culture. Environ Pollut 158:2589–2595. https://doi.org/10.1016/j.envpol.2010.05.015
Elsayed H, Yigiterhan O, Al-Ansari E et al (2020) Methylmercury bioaccumulation among different food chain levels i-n the EEZ of Qatar (Arabian Gulf). Reg Stud Mar Sci 37:101334. https://doi.org/10.1016/j.rsma.2020.101334
Fu Y, Yu Z, Cai K et al (2010) Mechanisms of iron plaque formation on root surface of rice plants and their ecologi-cal and environmental effects: a review. Plant Nutr Fertitizer Sci 16:1527–1534. https://doi.org/10.4028/www.scientific.net/AMM.37-38.1549
Hong Y, Kim Y, Lee K (2012) Methylmercury exposure and health effects. J Prev Med Public Health 45:353–363. https://doi.org/10.3961/jpmph.2012.45.6.353
Horvat M, Nolde N, Fajon V et al (2003) Total mercury, methylmercury and selenium in mercury polluted areas in the province Guizhou, China. Sci Total Environ 304:231–256. https://doi.org/10.1016/s0048-9697(02)00572-7
Hu Y, Huang Y, Liu Y (2013) Influence of iron plaque on chromium accumulation and translocation in three rice (Oryza sativa L.) cultivars grown in solution culture. Chem Ecol 30:29–38. https://doi.org/10.1080/02757540.2013.829050
Jiang G, Shi J, Feng X (2006) Mercury pollution in China. An overview of the past and current sources of the t-oxic metal. Environ Sci Technol 40:3673–3678. https://doi.org/10.1021/es062707c
Li Y, Zhao J, Gao Y et al (2014) Effects of iron plaque and selenium on the absorption and translocation of inorgani-c Mercury and Methylmercury in Rice (Oryza sativa L.). Asian J Ecotoxicol 9:972–977. https://doi.org/10.7524/aje.1673-5897-20140519002
Li Y, Zhao J, Zhang B et al (2015) The influence of iron plaque on the absorption, translocation and transformation o-f mercury in rice (Oryza sativa L.) seedlings exposed to different mercury species. Plant Soil 398:87–97. https://doi.org/10.1007/s11104-015-2627-x
Liu C, Chen C, Gong X et al (2014) Progress in research of iron plaque on root surface of wetland plants. Acta Ecol Sin 34:2470–2480. https://doi.org/10.5846/stxb201304110685
Liu W, Zhu Y, Smith F et al (2004) Do iron plaque and genotypes affect arsenate uptake and translocation by rice seedlings (Oryza sativa L.) grown in solution culture? J Exp Bot 55:1707–1713. https://doi.org/10.1093/jxb/erh205
Meng B, Feng X, Qiu G et al (2010) Distribution patterns of inorganic mercury and methylmercury in tissues of rice (Oryza sativa L.) plants and possible bioaccumulation pathways. J Agric Food Chem 58:4951–4958. https://doi.org/10.1021/jf904557x
Meng M, Li B, Shao J et al (2014) Accumulation of total mercury and methylmercury in rice plants collected from di-fferent mining areas in China. Environ Pollut 184:179–186. https://doi.org/10.1016/j.envpol.2013.08.030
Meng D, Zhu Y, Zhou J (2008) Study on the relationship between aerenchyma of rice root systems and iron Plaque outside roots. Modern Agric Sci 04:55–58 (in Chinese)
Peng C, Chen S, Shen C et al (2018) Iron plaque: a barrier layer to the uptake and translocation of copper oxide Na-noparticles by rice plants. Environ Sci Technol 52:12244–12254. https://doi.org/10.1021/acs.est.8b02687
Qin C, Du B, Yin R et al (2020) Isotopic fractionation and source appointment of methylmercury and inorganic me-rcury in a paddy ecosystem. Environ Sci Technol 54:14334–14342. https://doi.org/10.1021/acs.est.0c03341
Shang S, Tian P, Jiang Y et al (2017) Uptake and volatilization of gaseous elemental mercury by paddy rice. Huanjing Kexue 38:5308–5314. https://doi.org/10.13227/j.hjkx.201704034
Strickman R, Mitchell C (2017) Accumulation and translocation of methylmercury and inorganic mercury in Oryza sativa: an enriched isotope tracer study. Sci Total Environ 574:1415–1423. https://doi.org/10.1016/j.scitotenv.2016.08.068
Tang B, Chen J, Wang Z et al (2021) Mercury accumulation response of rice plant (Oryza sativa L.) to elevated atmo-spheric mercury and carbon dioxide. Ecotoxicol Environ Safe. https://doi.org/10.1016/j.ecoenv.2021.112628
Taylor G, Crowder A (1983) Use of the DCB technique for extraction of hydrous iron oxides from roots of wetla-nd plants. Am J Bot 70:1254–1257. https://doi.org/10.1002/j.1537-2197.1983.tb12474.x
Trivedi P, Axe L (2001) Modeling Cd and Zn soprtion to hydrous metal oxides 34:2000–2215. Environ Sci Technol 35:1892–1892. https://doi.org/10.1021/es010703w
Wang X, Jiang J, Dou F et al (2022) Zinc fertilizers modified the formation and properties of iron plaque and arsenic accumulation in rice (Oryza sativa L.) in a life cycle study. Environ Sci Technol 56:8209–8220. https://doi.org/10.1021/acs.est.2c01767
Wu S, Oya N, Ozaki M et al (2007) Development of iron oxide sorbents for Hg-0 removal from coal derived fuel ga-s: sulfidation characteristics of iron oxide sorbents and activity for COS formation during Hg-0 removal. Fuel 86:2857–2863. https://doi.org/10.1016/j.fuel.2007.03.004
Yin R, Feng X, Meng B (2013) Stable mercury isotope variation in rice plants (Oryza sativa L.) from the wanshan mercury mining district SW China. Environ Sci Technol 47:2238–2245. https://doi.org/10.1021/es304302a
Zagar D, Sirnik N, Cetina M et al (2014) Mercury in the Mediterranean. Part 2: processes and mass balance. EnvironSci Pollut Res 21:4081–4094. https://doi.org/10.1007/s11356-013-2055-5
Zhang H, Feng X, Larssen T et al (2010) Bioaccumulation of methylmercury versus inorganic mercury in rice (Oryza sativa L.) grain. Environ Sci Technol 44:4499–4504. https://doi.org/10.1021/es903565t
Acknowledgements
This research was supported by Shanghai Key Lab for Urban Ecological Processes and Eco-Restoration (SHUES2021C02) and the Fundamental Research Funds for the Central Universities and was funded by Technology Innovation Center for Land Spatial Eco-restoration in Metropolitan Area, Ministry of Natural Resources. We would also like to thank the National Natural Science Foundation of China (31400445) for supporting this study.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study. Experimental design and chemical analysis were performed by SS, and data processing was performed by XL, SC, and RL. The first draft of the manuscript was written by YW. WJ was responsible for data processing, content editing and language polishing of the revised manuscript. All authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Wang, Y., Shang, S., Li, X. et al. Effect of Iron Plaque on Gaseous Elemental Mercury Uptake, Translocation, and Volatilization by Paddy Rice. Bull Environ Contam Toxicol 109, 1155–1161 (2022). https://doi.org/10.1007/s00128-022-03617-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00128-022-03617-0