
Abstract
Acid DNase activity in marine organisms displays a sensitive response to pollutants. In the present study, ex situ effect of toxic seawater organic extracts (Rijeka, Vranjic, Ploče) and in situ impact of environmental factors at referent (St. Andrew) and a polluted site (ACI Rovinj) on acid DNase in the digestive gland and gills of mussels was investigated. In the digestive gland, enzyme activity was related to potential toxicity of seawater organic extracts. Seawater organic extracts showed an increase of the enzyme activity in digestive gland following exposure. In in situ study, polluted environment affected mussel’s acid DNase activity in digestive gland, when compared to gills. Acid DNase activity in the digestive gland expressed significant difference between the referent and polluted site, particularly from early spring through summer. Both ex situ and in situ studies displayed more sensitive acid DNase response to the presence of toxic mixtures in the digestive gland.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
The use of biological parameters measured at the molecular or cellular level have been proposed as sensitive ‘early warning’ tool which should provide a general view of mussel’s health in environmental quality assessment. The enzyme systems measurements in marine organisms express many advantages (high sensitivity, selectivity, economically efficient equipment and experimental techniques) that allow integrated information on environmental pollution (Menzorova et al. 2014). Many enzyme systems have become of great interest as candidates for investigation of water pollution.
Acid DNase, also named DNase II is a lysosomal enzyme implicated in many cellular processes. Early studies have shown that acid DNase function is required during degradation of DNA within phagolysosomes, during infection and during degradation of apoptotic cells by phagocytic cells (Evans and Aguilera 2003.). Recent work of deoxyribonucleases (DNases) from bivalves (Menzorova and Rasskazov 2007; Fafanđel et al. 2008), snail (Popov et al. 2003, 2008) and sea urchin (Menzorova and Rasskazov 1999, 2007, 2009) correlated the DNase response to the presence of model marine pollutants: heavy metals, oil and detergents. Kovačić and Medić (2016) showed the responce of acid DNase from mussel digestive gland to pesticide. At the same time, acid DNase have shown sensitiveness to evaluate the marine ecosystem in in situ studies. DNase from mussel Crenomytilus grayanus was used during the analysis of seawater samples from Troitsa Bay (Menzorova and Rasskazov 2007). Fafanđel et al. (2008) showed the response of acid DNase activity in Mytilus galloprovincialis to model pollutants and in Rijeka Bay using specific acid DNase activity in digestive gland and haemocytes ratio (Hep/Hem). At the same time, simultaneous testing of acid DNase (from one single tissue) on 96-multiwell plates allowed considerably shortening time for analysis (Kovačić and Medić 2016).
A good enzyme system to be used in the field studies should be responsive to environmental pollutant mixtures (Olmstead and LeBlanc 2005), with no or low sensitivity to natural environmental conditions. In this respect, the understanding of natural changes of enzyme activity is necessary for interpretation of in situ results, and to discriminate the onset of biological disturbance from the natural variability (Bocchetti and Regoli 2006).
The aim of this study was to detect the suitable tissue for acid DNase measurement in mussel, sensitive enough to detect contaminant presence and anthropogenic impact, under environmental conditions of fluctuating ecological factors, including seasonal variation of mussel physiology.
Materials and Methods
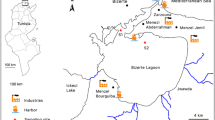
Seawater samples (30 L) were collected in March, June, August and October from 1999 to 2005 at three sampling sites along the Adriatic coast, Croatia (Fig. 1) representing a small rural area (Ploče), shipyard (Rijeka), urban sewage and industrial outflow (Vranjic). Extraction of the organic fraction occurred at each sampling time according to Bihari et al. (2004a). Potential toxicity and genotoxicity of seawater organic extracts was determined by bioluminescent assay Microtox® test and the umu-test, respectively (Bihari et al. 2004a, b). The toxic potential of seawater organic extracts was characterized as low (Ploče), moderate (Rijeka) and high (Vranjic) toxicity (Final Report 2014). Pools from each sampling site of the seawater organic extracts were made, and stored at +4 °C in dimethyl sulfoxide (DMSO).

Mussel (square) and seawater (dot) sampling sites along the Adriatic coast, Croatia
For the laboratory experiment, mussels M. galloprovincialis of similar shell length (55–65 mm) were obtained from the mariculture area of Lim Bay (45°07′50′′N 13°44′10′′E) (Fig. 1) and maintained in seawater basins for acclimation. After 48 h, 120 mussel specimens were randomly allocated into four aerated experimental aquaria (30 specimens per basin) containing seawater. In first basin, control mussel group consisted of 30 mussel injected with 50 µl DMSO. In the other three basins, each of 30 mussel specimen was injected with 50 µl of one of seawater organic extract (30 specimens/extract). Ten specimens per basin (extract treatment) were randomly sampled at 3, 24 and 48 h for tissue dissection.
For in situ study, ten mussel specimens per site were collected monthly from August 2012 to August 2013. Collection of mussels was carried out at two sampling sites in Northern Adriatic, Croatia (Fig. 1) representing a reference (St. Andrew) and polluted marina with a small shipyard close to urban sewage area (ACI Rovinj). Reference site is located in Rovinj coastal area at 14 km distance from urban and sewage outflow area. ACI Rovinj was characterized by elevated pollutant concentration (metals, PAH) in sediment and biota compared to nearby undisturbed environment (Bihari et al. 2007; Fafanđel and Bihari 2007; Final report 2014). After collection, specimens were transported to the laboratory where they were immediately dissected.
Mussel tissues were homogenised and fluorometric enzyme activity measurement was done using dsDNA-binding fluorochrome PicoGreen® (Molecular Probes Inc., USA) in a Fluoroscan Ascent microplate reader (Labsystems, Finland) (Fafanđel et al. 2008; Kovačić et al. 2015; Kovačić and Medić 2016). Enzyme activity was defined as change in the amount of the dsDNA-PicoGreen® complex (Delta fluorescence) after hydrolytic cleavage of DNA by the acid DNase present in the sample. It was calculated by subtracting the measured fluorescence of the reaction mixture containing tissue homogenate from the blank value obtained from the reaction mixture containing heat denaturated (95°C, 15 min) homogenate. Results were presented as specific enzyme activity, ∆F mg − 1 min − 1.
Western blot analysis was done according to Kovačić et al. (2015).
All data were expressed as mean ± standard deviation (SD). For statistical analyses, STATISTICA (version 6.0.) was used. Significant differences (p < 0.05) among groups were studied using Kruskal–Wallis test, followed by Mann Whitney tests to differentiate the groups of data.
Results and Discussion
The present study investigated acid DNase response to environmental pollutant mixtures in ex situ and in situ study. In all investigated samples, the acid DNase activity was higher in exposed mussel then in control (Fig. 2). Observed results are in accordance with reports on model pollutants eliciting effects at acid DNase activity level (Popov et al. 2003; Fafanđel et al. 2008). Ex situ study was done to investigate whether the acid DNase activity is responsive to seawater organic extracts distributed in mussel tissues. The response of acid DNase in mussel digestive gland and gills exposed to toxic organic seawater extracts (ex situ study) was tissue-specific (Fig. 2). Acid DNase activity in digestive gland was related to the level of toxicity of seawater organic extracts. Unlike digestive gland, the acid DNase activity in gills had no correspondence to the seawater organic extract toxic potential. In the digestive gland, time response of increased acid DNase activity was detected when compared to gills. All organic seawater extracts induced a significant increase in acid DNase activity after 3 h. After 24 h, the acid DNase activity decreased but was still elevated when compared to 3 h in mussels treated with seawater organic extracts with moderate and high potential toxicity. After 48 h, the acid DNase activity fell to a basal level in mussels exposed to seawater organic extracts with low and moderate potential toxicity, while the enzyme activity remained elevated in mussels exposed to seawater organic extract with high potential toxicity. In gills, acid DNase activity was not affected after exposure to seawater organic extracts with low potential toxicity. Significantly elevated (p < 0.05) acid DNase activity was observed in gills after 3 and 48 h of exposure to seawater organic extracts with moderate potential toxicity and after 24 h of exposure to organic seawater extracts with high potential toxicity. Different time-response pattern of each organic mixture extract in both tissues is considered as the result of tissue-specific susceptibility to a particular contaminant in the mixture and its metabolic pathway. The observed tissue-specific response may be due to distinctive physiochemical activities specific for these two organs, especially with respect to either the activation or detoxification of pollutants or the repair of different types of DNA breaks (de Almeida et al. 2003). Since the damaged DNA is metabolised by deoxyribonucleases (Evans and Aguilera 2003), it could be expected that the digestive gland as tissue with higher metabolic activity had more DNA damage (Al-Subiai et al. 2012). Therefore, the activity of haepatopancreatic DNase could be affected more than the gills’. Acid DNase response in mussel tissues could not be related with the application of a toxicant. Injection of a toxicant under the mantle enable the distribution of seawater organic extracts through the body with open circular system. Moreover, our previous results showed toxicant distributed in mussel organs after 90 min from injection (Bihari et al. 1990; Mičić et al. 2001).

Acid DNase activity in the digestive gland (DG) and gills (G) of control mussels Mytilus galloprovincialis and exposed to seawater organic extracts with low (Ploče), moderate (Rijeka) or high (Vranjic) potential toxicity for 24, 48 and 72 h. Bar chart results are presented as mean ± SD ( n = 10). Asterisks (*) denote significant differences (p < 0.05) from the DMSO control
Tissue-specific response to toxic seawater organic extracts was also observed at the protein expression level (Fig. 3). Changes in the expression pattern of acid DNase were pronounced in the digestive gland. It is of particular importance to note that this induction was observed in all the samples of whatever the toxicity potential of the extract. Obtained results evidenced up regulation of 37 and 48 kDa immunoreactive bands. Those bands correspond to precursors of the acid DNase during enzyme maturation in mussel lysosomes (Kovačić et al. 2015) indicating induced de novo synthesis of the acid DNase following exposure to toxic organic pollutants.

Expression pattern of the acid DNase in the digestive gland (DG) and gills (G) of control mussels Mytilus galloprovincialis (DMSO, 24 h) and exposed to seawater organic extracts with moderate (Rijeka, 24 h) and high (Vranjic, 48 h) potential toxicity
Moreover, the potential of mussel acid DNase activity to discriminate sites with different pollution load in native mussel under environmental conditions (in situ observations) over 1-year period was investigated. Environmental mixtures of pollutants in the shipyard and urban sewage area (ACI Rovinj) affected the acid DNase that could be observed as a decrease in enzyme activity in both organs (Fig. 4). This decrease in enzyme activity in mussels sampled at the polluted site was in accordance with the observations reported for Crenomytilus grayanus (Menzorova and Rasskazov 2007). Similarly, a significant decrease in the activity of the acid DNase with a decrease in water quality was observed in the sea urchin DNase test system (Menzorova and Rasskazov 2009). Contrary, the increase in acid DNase activity in mussel tissues caused by pollution was observed in a previous in situ study in the Gulf of Rijeka, Croatia (Fafanđel et al. 2008). The response of enzyme activity to metal concentration in seawater samples could explain this. In fact, the presence of divalent metal ions (Ca2+, Cu2+, Co2+, Mg2+, Mn2+) in human COS-7 and Hep G2 cells as found by Yasuda et al. (1992) and Cu2+ in musses haemocytes (Fafanđel et al. 2008) have led to a suppression of acid DNase activity. In ACI Rovinj marina, heavy metals (lead, zinc, and cooper) were found to be bioconcentrated in mussel soft tissues from two to six times, compared to nearby undisturbed environment (Final Report 2014). Contaminant levels in tissues are likely to change during the season, reflecting variability in the discharge and metabolism of contaminants and/or changes in tissue weight and enzyme activities (Verlecar et al. 2007; Trisciani et al. 2012). An ideal parameter would show no seasonal variability in response to factors such as food supply or reproductive status and would vary only in response to pollutant exposure, but in practice, this seldom is the case (Sheehan and Power 1999). However, the environmental conditions in summer did not mask the pollution effect on acid DNase activity in the digestive gland when compared to gills (Fig. 4). Moreover, the results of this seasonal study revealed period when natural environmental and endogenous factors were less effective suggesting that monitoring of pollution effects in this part of the Adriatic Sea should be carried out in the period of spring—early summer. This was observed in previously published data of acid DNase from mussels sampled at the reference site (St. Andrew) in northern Adriatic (Kovačić et al. 2015), and other enzymes from mussels sampled in Greece (Koukouzika et al. 2009) and the Venice lagoon (Nesto et al. 2007).

Acid DNase activity in the digestive gland (DG) and gills (G) of mussels Mytilus galloprovincialis sampled at a reference (St Andrew) and a polluted (ACI Rovinj) site from August (VIII) 2012 to August (VIII) 2013. Bar chart results are presented as mean ± SD ( n = 10). Asterisks (*) denote significant differences (p < 0.05) between reference and polluted sites. Reference site data was presented in published study by Kovačić et al. (2015)
Since both, in situ and ex situ studies showed acid DNase activity response in the digestive gland as sensitive enough to discriminate sites with different potential toxicity, it could be proposed for the assessment of pollutant exposure instead of Hep/Hem ratio (Fafanđel et al. 2008). The advantage of the acid DNase activity measurement in one single tissue is shortening time in comparison to measuring in two mussel tissues.
References
Al-Subiai SN, Arlt VM, Frickers PE, Readman JW, Stolpe B, Lead JR, Moody AJ, Jha AN (2012) Merging nano-genotoxicology with eco-genotoxicology: an integrated approach to determine interactive genotoxic and sub-lethal toxic effects of C(60) fullerenes and fluoranthene in marine mussels, Mytilus sp. Mut Res 745:92–103
Bihari N, Batel R, Zahn RK (1990) DNA damage determination by the alkaline elution technique in the haemolymph of mussel Mytilus galloprovincialis treated with benzo[a]pyrene and 4-nitroquinoline-N-oxide. Aqua Toxicol 18:13–22
Bihari N, Mičić M, Fafandel M (2004a) Seawater quality along the Adriatic coast, Croatia, based on toxicity data. Environm Toxicol 19:109–114
Bihari N, Mičić M, Fafanđel M, Hamer B, Jakšić Ž, Batel R (2004b) Seawater quality of Adriatic coast, Croatia, based on toxicity, genotoxicity and DNA integrity assay. Acta Adriat 45:75–81
Bihari N, Fafanđel M, Piškur V (2007) Polycyclic Aromatic Hydrocarbons and Ecotoxicological Characterization of Seawater, Sediment, and Mussel Mytilus galloprovincialis from the Gulf of Rijeka, the Adriatic Sea, Croatia. Arch Environm Contam Toxicol 52:379–387
Bocchetti R, Regoli F (2006) Seasonal variability of oxidative biomarkers, lysosomal parameters, metallothioneins and peroxisomal enzymes in the Mediterranean mussel Mytilus galloprovincialis from Adriatic Sea. Chemosphere 65:913–921
de Almeida EA, Marques Sde A, Klitzke CF, Bainy AC, de Medeiros MH, Di Mascio P, Loureiro AP (2003) DNA damage in digestive gland and mantle tissue of the mussel Perna perna. Comp Biochem Physiol C Toxicol Pharmacol 135:295–303
Evans CJ, Aguilera RJ (2003) DNase II: genes, enzymes and function. Gene 322:1–15
Fafanđel M, Bihari N (2007) Temporal and spatial variations of seawater toxicity along Adriatic coast, Croatia: a long-term study. Fresh Environm Bull 16:1457–1463
Fafanđel M, Bihari N, Peric L, Cenov A (2008) Effect of marine pollutants on the acid DNase activity in the hemocytes and digestive gland of the mussel Mytilus galloprovincialis. Aqua Toxicol 86:508–513
Final report (2014) Coastal Cities Pollution project 2, The adriatic Sea Monitoring Program 2nd Phase. Center for Marine Research Split
Koukouzika N, Raftopoulou EK, Dimitriadis VK (2009) Seasonal differences of lysosomal, lipid and lipofuscin parameters in the digestive gland of the mussel Mytilus galloprovincialis. J Molluscan Stud 75:261–267
Kovačić I, Medić N (2016) The effect of chlorpyrifos on protein content and acid DNase activity in the mussel, Mytilus galloprovincialis. Mar Freshwat Behav Physiol 49:1–11
Kovačić I, Fafanđel M, Bihari N (2015) Lysosomal deoxyribonuclease II from the mussel Mytilus galloprovincialis: Characterization and seasonal activity. Mar Biol Res 11:716–724
Menzorova NI, Rasskazov VA (1999) Application of the sea urchin Strongylocentrotus intermedius embryo DNAse test in the evaluation of marine water pollution by various toxicants. Russ J Mar Biol 25:60–64
Menzorova NI, Rasskazov VA (2007) Application of different test systems and biochemical indicators for environmental monitoring of the Troitsa Bay, Sea of Japan. Russ J Mar Biol 33:118–124
Menzorova NI, Rasskazov VA (2009) Evaluation of the ecological state of the Sea of Japan and the Sea of Okhotsk using the DNase test system. Oceanology 49:824–832
Menzorova NI, Seitkalieva AV, Rasskazov VA (2014) Enzymatic methods for the determination of pollution in seawater using salt resistant alkaline phosphatase from eggs of the sea urchin Strongylocentrotus intermedius. Mar Pollut Bull 79:188–195
Mičić M, Bihari N, Labura Z, Muller WE, Batel R (2001) Induction of apoptosis in the blue mussel Mytilus galloprovincialis by tri-n-butyltin chloride. Aqua Toxicol 55:61–73
Nesto N, Romano S, Moschino V, Mauri M, Da Ros L (2007) Bioaccumulation and biomarker responses of trace metals and micro-organic pollutants in mussels and fish from the Lagoon of Venice, Italy. Mar Pollut Bull 55:469–484
Olmstead AW, LeBlanc GA (2005) Toxicity assessment of environmentally relevant pollutant mixtures using a heuristic model. Integr Environm Assess Manag 1:114–122
Popov AP, Konichev AS, Tsvetkov IL (2003) Effect of Toxic Industrial Pollutants on the Activity and Isoforms of Acid DNase in the Freshwater Snail Viviparus viviparus L. Appl Biochem Microbiol 39:454–458
Popov AP, Tsvetkov IL, Konichev AS (2008) Separation and characterization of deoxyribonucleases from hepatopancreas of freshwater snail in normality and under in vivo model intoxication. Biochemistry (Mosc) 73:937–942
Sheehan D, Power A (1999) Effects of seasonality on xenobiotic and antioxidant defence mechanisms of bivalve molluscs. Comp Biochem Physiol C 123:193–199
Trisciani A, Perra G, Caruso T, Focardi S, Corsi I (2012) Phase I and II biotransformation enzymes and polycyclic aromatic hydrocarbons in the Mediterranean mussel (Mytilus galloprovincialis, Lamarck, 1819) collected in front of an oil refinery. Mar Environm Res 79:29–36
Verlecar XN, Jena KB, Chainy GBN (2007) Biochemical markers of oxidative stress in Perna viridis exposed to mercury and temperature. Chem Biol Interact 167:219–226
Yasuda T, Nadano D, Sawazaki K, Kishi K (1992) Genetic polymorphism of human deoxyribonuclease II (DNase II): low activity levels in urine and leukocytes are due to an autosomal recessive allele. Ann Hum Gen 56:1–10
Acknowledgements
The Croatian Ministry of Science, Education and Sport financially supported this study; Project No. 098-0982705-2725 (Dr. Nevenka Bihari).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Kovačić, I., Fafanđel, M., Perić, L. et al. Effect of Environmental Pollutant Mixtures on Acid DNase Activity in Mussel Mytilus galloprovincialis: Ex Situ and In Situ Study. Bull Environ Contam Toxicol 99, 433–437 (2017). https://doi.org/10.1007/s00128-017-2162-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00128-017-2162-y