
Abstract
Soft-bodied preservation is common in the Cambrian but comparatively rare in the Ordovician. Here, a new deposit preserving soft-bodied fossils is reported from the Middle Ordovician (Dapingian-Darriwilian) upper Valongo Formation of northern Portugal. The deposit contains the first known occurrences of soft-bodied fossils from the Middle Ordovician (Dapingian-Darriwilian) of Portugal and is the first Ordovician example of soft-tissue preservation involving carbonaceous films from the Iberian Peninsula. It also represents the lone deposit of soft-bodied fossils from the Middle Ordovician of northern Gondwana. Thus temporally, it lies between the exceptional deposits of the Lower Ordovician of Fezouata (Morocco) and the Upper Ordovician of the Soom Shale (South Africa); it also serves as a biogeographic link between these and the various Ordovician soft-bodied deposits in Laurentia. The soft-bodied fossils come from the deep-water slates of the upper part of the Valongo Formation and include a discoidal fossil questionably referable to Patanacta, wiwaxiid sclerites, and a possible pseudoarctolepid arthropod.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Deposits preserving soft-bodied fossils are critical for understanding the diversity and origins of early animals, as they preserve organisms that are prone to decay and would usually not be preserved in the fossil record (e.g., Butterfield 1990; Schiffbauer et al. 2014; Muscente et al. 2017; Daley et al. 2018). These deposits are most common in Cambrian Series 2 and 3 (Conway Morris 1989; Gaines 2014; Robison et al. 2015; Kimmig et al. 2019), and their record declines subsequently (e.g., Lerosey-Aubril et al. 2018), with only a few deposits known from the Ordovician (Allison and Briggs 1993; Butterfield 1995; Gaines 2014; Van Roy et al. 2015; Briggs et al. 2018). The most diverse Ordovician Burgess Shale type (BST) deposit known to date is the Lower Ordovician Fezouta biota of Morocco (Van Roy et al. 2010, 2015; Martin et al. 2016). Other deposits preserving soft-tissue remains include the Lower Ordovician Afon Gam Biota of Wales (Botting et al. 2015); the Middle Ordovician Winneshiek Lagerstätte of Iowa (Liu et al. 2006, 2009; Briggs et al. 2018) and Llanfallteg Formation in Wales (Hearing et al. 2016); and the Upper Ordovician Beecher’s Trilobite Bed of New York (Briggs et al. 1991), Big Hill Formation of Michigan (Lamsdell et al. 2017), Martinsburg Formation of Pennsylvania (Meyer et al. 2018), “Upper Ordovician Lagerstätten” from Manitoba, Canada (Young et al. 2007, 2012), Bardahessiagh Formation of Ireland (MacGabhann and Murray 2010), and Soom Shale of South Africa (Aldridge et al. 1994; Gabbott 1998). The information preserved in these Ordovician exceptional deposits has greatly improved knowledge on the expansion of biodiversity during the Great Ordovician Biodiversification Event (GOBE) (Webby et al. 2004; Servais and Harper 2018). These deposits have also revealed that several clades of soft-bodied taxa once thought to have gone entirely extinct during the Cambrian in fact survived into the Ordovician (e.g., Van Roy et al. 2010; Botting et al. 2015; Hearing et al. 2016).
Here, we report on the first Middle Ordovician (Dapingian-Darriwilian) soft-bodied fossils from northern Gondwana. They are preserved in the upper Valongo Formation of northern Portugal (Fig. 1), which is famous for its trilobite diversity (Delgado 1908; Romano and Diggens 1974; Romano 1991), as well as diverse nautiloids, gastropods, and echinoderms (Delgado 1908; Couto et al. 1997; Ausich et al. 2007; Couto 2013). The soft-bodied fossils comprise a discoidal fossil, possible wiwaxiid sclerites, and an arthropod carapace.

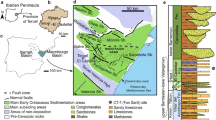
a Geological map of the Valongo Anticline area, northern Portugal, showing the locations of study specimens, indicated by the stars (S. Pedro da Cova, 41° 9′ N, 8° 29′ W; Belói 41° 8′ N, 8° 29′ W) (modified from Couto et al. 2013). A, Cambrian: SJF, Santa Justa Formation; VF, Valongo Formation; SF, Sobrido Formation; B, Silurian to Carboniferous. b Generalized lithostratigraphic column of the Paleozoic succession in the study area: 1, quartzites; 2, slates; 3, diamictites; 4, slates and siltstones; 5, conglomerates; 6, acidic volcanics; 7, basic volcanics (modified from Couto and Roger 2017)
Soft-tissue preservation, and especially BST preservation, is well documented in deposits of Cambrian age (Conway Morris 1989; Allison and Briggs 1993; Butterfield 1995; Orr et al. 1998; Briggs and Fortey 2005; Gaines 2014), but there are far fewer deposits known from the Ordovician (Van Roy et al. 2015; Muscente et al. 2017). This is partially due to alteration of ocean pH and Eh; oxygenation of upper sediment layers by increasing and deeper bioturbation; and major erosional and facies differences related to global tectonics, which altered chemical availability for cement formation and changed ocean shelf areas (Gaines et al. 2012a, b; Daley et al. 2018). Another reason for the sparse preservation of these organisms is their delicate nature and their tendency to decompose before preservation. In most early Paleozoic cases, these organisms are preserved as two-dimensional carbon or aluminosilicate films in carbonaceous shales (Gaines 2014; Muscente et al. 2017). The best-known Ordovician deposits with soft-tissue preservation in a plethora of taxa are the Lower Ordovician Fezouta Lagerstätte in Morocco (Van Roy et al. 2015; Martin et al. 2016) and the Upper Ordovician Soom Shale of South Africa (Gabbott 1998; Gabbott et al. 2017). These two Lagerstätten allow snapshots into the biodiversity of the Ordovician and show the changes that happen during this period. The Fezouata Lagerstätte has yielded several clades of typical Cambrian taxa (i.e., radiodonts, and marellomorphs) together with typical members of later Paleozoic faunas (Van Roy et al. 2015). The Soom Shale in contrast is dominated by typical Paleozoic animals (i.e., conodonts, orthoconic nautiloids, brachiopods) and has few survivors of the Cambrian radiation (Gabbott et al. 2017).
Some of the most common soft-bodied fossils in the Ordovician are discoidal fossils. They have been described from the deposits of Ordovician age (Fig. 2 and Suppl. 2) that otherwise contain minimal to no other instances of soft-tissue preservation, including New York State (Discophyllum peltatum Hall 1847), Sweden (Patanacta pedina Cherns 1994), Ireland (Septus pomeroii MacGabhann and Murray 2010), and Morocco (D. peltatum; “Eldonia”). The Valongo Formation of northern Portugal preserves a diverse well-skeletonized fauna (Delgado 1908; Couto et al. 1997; Ausich et al. 2007; Couto 2013), but the herein described associated soft-bodied fauna is sparse and usually poorly preserved. The only previously described soft-bodied fossil from the Lower Ordovician of Portugal is a discoidal fossil referred to as Discophyllum plicatum Hall 1847 by Delgado (1892) from the slightly older Armoricain Quartzite Formation of Buçaco (Central Portugal), which is coeval with the Santa Justa Formation from the Valongo Anticline (Tremadocian-Floian, the fossils are from the Floian interval), but the specimen is currently reported missing.

Stratigraphic chart showing the occurrences of soft-bodied fossils in the Ordovician. Deposits with soft-bodied discoidal fossils are shown in bold
Geological setting
The deposit described herein is located near the village of S. Pedro da Cova, in northern Portugal. Placed in the Central-Iberian Zone, it is part of the Valongo Anticline, a major ante-Stephanian asymmetrical antiform anticline trending NW-SE, whose axis plunges 5° to 15 ° to the NW, with an axial plane 60° to the NE (Ribeiro et al. 1987). The anticline is surrounded by Variscan granites (Fig. 1a). Lower Paleozoic metasediments in this region range in age from Cambrian to Devonian (Fig. 1b), and at this locality, the Cambrian is represented by the Montalto Formation which consists of interbedded slates, quartzites, polygenic conglomerates, and volcanics (Couto 1993). Three Ordovician formations were defined by Romano and Diggens (1974), later revised by Couto (2013) and Couto et al. (2013). These are, in ascending order, (1) the Santa Justa Formation (Tremadocian-Floian), which is formed by volcanic rocks (Tremadocian) and platform deposits, which lie below interbedded fine-grained and coarse-grained clastic sediments and volcano-sedimentary layers, indicating subsiding and tectonically unstable sedimentation conditions; (2) the Valongo Formation (Dapingian-Darriwilian), which is formed by fine-grained clastic sediments deposited in shallow to deep-water environments; and (3) the Sobrido Formation (Hirnantian), which contains glacially influenced marine rocks deposited on the north Gondwana platform.
The Valongo Formation varies locally from 300 to 400 m thick and consists of a succession of slightly metamorphosed (greenschist facies) slates and siltstones (Fig. 1b). The base is formed by fossiliferous pink siltstones, followed by dark gray siltstones probably of Dapingian age. Gray siltstones and dark gray fossiliferous shales of Dapingian-Darriwilian age overlay these strata. These shales are gradually replaced by slates, which host the soft-bodied fauna. These in turn are overlain by fossiliferous light gray siltstones (Romano and Diggens 1974; Couto 1993). Petrographic study of these metasediments showed that they are mainly comprised of chlorite and muscovite (Couto 1993). The exact beds that preserve the soft-bodied fossils are uncertain, as all soft-bodied fossils have been collected from the scree, but the lithology corresponds to the slates in the upper part of the Valongo Formation.
Fossils
The Valongo Formation is one of the most fossiliferous units in Portugal and comprises a diverse biota of benthic and pelagic animals. In addition to the putative Patanacta, wiwaxiid sclerites, and bivalved arthropod, the fauna includes over 150 different species of trilobites, graptolites, brachiopods, gastropods, bivalves, cephalopods, echinoderms, and several groups of uncertain affinity (Delgado 1892, 1897, 1908; Thadeu 1949; Curtis 1961; Romano 1975, 1976, 1980, 1982a, b, 1990, 1991; Romano and Diggens 1974; Romano and Henry 1982; Rábano 1989; Tauber and Reis 1994; Couto et al. 1997; Couto and Gutiérrez-Marco 1999, 2000; Ausich et al. 2007; Couto 2013). The assemblage is dominated by taxa characteristic of Ordovician communities, particularly trilobites and molluscs. The most common trilobites in the Valongo Formation are Actinopeltis (Valongia) wattisoni, Colpocoryphe rouaulti, Dionide mareki, Eccoptochile almadenensis, Prionocheilus mendax, Ectillaenus giganteus, Eodalmanitina macrophtalma, Eodalmanitina destombesi, Eoharpes cristatus, Isabelinia glabrata, Neseuretus tristani, Nobiliasaphus nobilis, Nobiliasaphus hammanni, Parabarrandia crassa, Phacopidina sp., Placoparia tournemini, Placoparia (Coplacoparia) borni, Protolloydolithus sp., Salterocoryphe salteri, Selenopeltis gallica, Uralichas hispanicus, Zeliszkella toledana, and Zeliszkella torrubiae (Delgado 1908; Romano 1976; Couto et al. 1997). Among mollusk taxa in the upper Valongo Formation, the most common are Cameroceras sp., Cardiolaria beirensis, Clathrospira sp., Hemiprionodonta lusitanica, Praenucula costae, Redonia deshayesi, Sinuites sp., Trocholites fugax, and various orthoceratids as well as rostroconchs (Babin et al. 1996; Couto and Gutiérrez-Marco 2000). Other taxa in the deposit are hyolithids, machaeridians (Plumulites sp.), ostracods, echinoderms such as Mitrocystella incipiens miloni, and rare graptolites referable to Orthograptus calcaratus (Delgado 1908; Babin et al. 1996; Couto et al. 1997; Couto and Gutiérrez-Marco 2000; Gutiérrez-Marco et al. 2000). In addition to the body-fossil record, a diverse ichnofossil record is present in the Valongo Formation, including burrows and microbially mediated traces (Neto de Carvalho et al. 2016).
Depositional environment
The sedimentary rocks of the Lower Ordovician of the study area (Fig. 1) were deposited in a shallow sea that formed due to rifting related to the opening of the Rheic Ocean (Couto et al. 2014). The facies hosting the soft-bodied (and biomineralized) fossils in the upper Valongo Formation is comparable to those known from Cambrian deep-water deposits containing BST fossils (Powell et al. 2003; Gabbott et al. 2008; Kimmig and Pratt 2016), as they are mostly comprised of aluminosilicates, chlorite and occasionally muscovite, and quartz (Couto 1993). In contrast to some other BST deposits, which are interpreted as having been deposited under oxic conditions (Powell et al. 2003; McKirdy et al. 2011; Kimmig and Pratt 2016; Sperling et al. 2018), the bottom waters of the Valongo Formation have been interpreted as at least temporarily dysoxic to anoxic based on ichnological information (Neto de Carvalho et al. 2016).
The depositional environment of the Valongo Formation appears to have been a relatively low energy setting, as there are no indications of ripples. The biota also supports this, as there are no putatively photosynthetic organisms preserved. The soft-bodied fossils were likely not transported very far, as the discoidal fossil and the wiwaxiids do not have any apparent damage.
Material and methods
The fossil specimens are housed in the collection of the Faculty of Sciences of the University of Porto, Department of Geosciences, Environment and Spatial Planning, Portugal (FCUP/DGAOT).
Elemental mapping utilizing energy-dispersive X-ray spectroscopy (EDS) was conducted at the University of Kansas Microscopy and Analytical Imaging Laboratory using an Oxford Instruments 80 mm2 x-Max silicon drift detector (SDD), mounted on an FEI Versa 3D Dual Beam. Analyses used a horizontal field width of 2.39 mm, a kilovolt of 10, a spot size of 4.5, and a 1000-μm opening (no aperture). EDS maps were collected at a pixel resolution of 512 × 512 with a total of 18 passes.
The fossils were photographed using a Canon EOS 5D Mark II digital SLR camera with a 50-mm Canon macro lens. The soft-bodied fossils (FCUP/DGAOT 3SPC and FCUP/DGAOT 5SPC) were photographed submerged in alcohol. The contrast, color, and brightness of the images were adjusted in Adobe Photoshop.
Results
Systematic palaeontology
Cnidaria Verrill, 1865
Medusozoa Petersen, 1979
Patanacta Cherns, 1994
Type species. P. pedina
Diagnosis. Ovoid to circular disc, weakly concavo-convex and with entire and distinct margin; convex and concave surfaces correspond closely. Circular central area, beyond which coarse, shallow, radial ridges originate and traverse to reach the margin, intersected by perpendicular, evenly spaced ridges (amended from Cherns 1994).
Patanacta? sp. indet. (Fig. 3a–e)

Patanacta? sp. indet. (FCUP/DGAOT 3SPC) from the upper part of the Valongo Formation of northern Portugal. a Dorsal view of the specimen. b Line drawing illustrating the preserved structures in a. c–e Close-ups of different parts of the specimen. c The center of the specimen, showing the circular central area and ridges connected to it. d The center bottom of the specimen, showing the edge of the fossil and ridges. e The top left of the specimen, showing the edge of the fossil and possible ridges. Scale bars are 5 mm
Material. One specimen in dorsoventral view (FCUP/DGAOT 3SPC)
Occurrence. Middle Ordovician slates of the upper part of the Valongo Formation, near S. Pedro da Cova, northern Portugal
Description. Ellipsoidal in outline. 59.8 mm long and 49.4 mm wide. Central circular area 5.7 mm in diameter, with ridges radiating out toward the edge of the specimen. At least 15 ridges, consistently about 3.1 mm wide and between 19.9 and 31.3 mm long. The ridges are straight and have no signs of branching. Margin mostly smooth, though slightly scalloped where the ridges are shorter (Fig. 3d).
Remarks. The specimen is questionably attributed to Patanacta, based on the circular central area, the radiating ridges, and the absent concentric rings. The smooth, uniform appearing edge of the fossil is likely due to compression or other taphonomic factors, it is not the same width throughout the specimen. The specimen seems to be preserved at somewhat of an angle to a dorsoventral plane, as the ridges vary in length between 19.9 and 31.3 mm. Patanacta was first described by Cherns (1994) from the Upper Ordovician-Silurian of Sweden. The Swedish specimen is preserved as a shiny, inorganic, and possibly silicate film (Cherns 1994), resembling in general the style of preservation of the Portuguese specimen. The Swedish specimen preserves 22–24 ridges, but the specimen from the Valongo Formation only preserves 15 complete ridges (Fig. 3a, b). This difference could be due to taphonomic factors, including compression, ontogenetic variation, or it could represent intra- or intergeneric variation. Given that this is the single known specimen, it is not assigned to species level.
Another Ordovician discoidal genus that shares features with the Portuguese specimen is Rutgersella Johnson and Fox, 1968 from the Australian Grindstone Range; it is preserved as three-dimensional external molds (Rettalack 2009). The specimens are elliptical in outline and preserve curving outward ribs and a central ellipsoidal depression (Rettalack 2009). Patanacta? illustrated herein, however, does not have curving outward ribs, and the center appears to be circular not elliptical. Additionally, there is controversy regarding Rutgersella as it has been considered a pseudofossil by Cloud (1973), although Rettalack (2009, 2015) argued that new fossils recovered from the Grindstone Range Sandstone in southern Australia support a biogenic origin. By contrast, in a review of the Dawson Hill Member of the Grindstone Range Sandstone Jago et al. (2010) argued that most, if not all, of the fossils described by Retallack (2009) are of inorganic origin.
Patanacta? from the Valongo Formation also differs from Ordovician and Cambrian representatives of Discophyllum Hall 1847 (see MacGabhann 2012; Lieberman et al. 2017) or the Ordovician Septus (MacGabhann and Murray 2010), as it lacks concentric rings, although this difference could be taphonomic (Briggs 2003; Kimmig and Pratt 2016).
The specimen does not appear to be a coprolite as these, when recovered from BST deposits, usually preserve carbon flakes or skeletal material (Vannier and Chen 2005; Kimmig and Strotz 2017; Kimmig and Pratt 2018), yet these are absent in Patanacta? Further, no radiating structures have been observed in coprolites.
Finally, the specimen does not appear to be a radiodontan oral cone of the type discussed in Pates et al. (2018) as the size of the central circular area is relatively far too small to represent the central opening of an oral cone, there is no evidence of possible marginal teeth/spines protruding into that central circular area, as would be expected if it was the central opening of an oral cone, and the radiating ridges do not show the linearity that would be expected if they represented the margins of plates of a radiodontan oral cone.
Lophotrochozoa Halanych et al., 1995
Wiwaxiidae? Walcott, 1911 (Fig. 4a)

a Wiwaxiid? sclerites from the upper part of the Valongo Formation in lateral view (FCUP/DGAOT 5SPC). b Ventro-lateral sclerite of Wiwaxia herka from the Cambrian Spence Shale of Utah in lateral view, collected by the Gunther Family (KUMIP 286302; see Conway Morris et al. 2015). Scale bars are 5 mm
Material. Two specimens (FCUP/DGAOT 5SPC) preserved in lateral view on one slab
Diagnosis. (See Conway Morris 1985)
Occurrence. Middle Ordovician slates of the upper part of the Valongo Formation, near S. Pedro da Cova, northern Portugal
Description. Two isolated recurved sclerites. The first one is 28 mm long and 2.4 mm wide in the center. The second is 23 mm long and missing both ends; it is 3.1 mm wide in the center. Both sclerites are thickest in the center and pinch out toward the ends. The more complete specimen is slimmer at one end than the other.
Remarks. The sclerites are tentatively attributed to Wiwaxiidae based on similarities to Wiwaxia corrugata from the Burgess Shale (see Fig. 145 in Conway Morris 1985) and Wiwaxia herka (see Conway Morris et al. 2015; Kimmig et al. 2019) from the Spence Shale (Fig. 4b). The assignment is, however, tentative because no microstructures are preserved. If indeed wiwaxiids, they likely represent ventro-lateral sclerites, based on their recurved shape. The sclerites would extend the range of the group into the Middle Ordovician; wiwaxiids have also been reported from the early Ordovician Fezouata biota (Van Roy et al. 2015).
Arthropoda von Siebold 1848
Pseudoarctolepidae? Brooks and Caster 1956 (Fig. 5a, b)

a Pseudoarctolepid? arthropod valves from the upper part of the Valongo Formation in lateral view (FCUP/DGAOT 20BE), posterior end of right valve showing possible dorsoventral compression. b Line drawing illustrating the preserved structures in a. c Butterflied valves of Pseudoarctolepis sharpi from the Cambrian Wheeler Formation of the Wheeler Amphitheater in the House Range of Western Utah, collected by Robert Harris (KUMIP 153913). Scale bars are 5 mm
Material. One specimen in lateral view (FCUP/DGAOT 20BE)
Diagnosis. (See Brooks and Caster 1956)
Occurrence. Middle Ordovician slates of the upper part of the Valongo Formation, near Belói, northern Portugal.
Description. Two valves partly overlying each other. Hinge line approximately straight. Total valve length 49.4 mm. Prominent ventral process 18.8 mm long, projecting from ventral margin well anterior of midline, weakly curved, with concave margin posteriorly, thinning slightly ventrally to end in weakly rounded point. Prominent posterior process 27.3 mm, only weakly thinning posteriorly; no anterior process preserved. Valve 14.1 mm at widest point (excluding ventral process).
Remarks. The specimen is strongly weathered, especially the valve that is displaced anteriorly and also weakly declined ventrally and slightly rotated counterclockwise. The valves preserve no or possibly weak, millimeter-sized trapezoid, ornamentation, and only the margins are well preserved. Valves appear to have been separated along the putative hinge line before burial, as the two valves are offset (Fig. 5a, b), with the posterior process only visible in the overlying valve. The specimen is tentatively assigned to the Pseudoarctolepidae Brooks and Caster 1956, as it shares with members of that family the shape of the valves and the prominent ventral process, which seems to originate at the same position of the valve. Further, the ventral processes bear roughly the same shape as those in Pseudoarctolepis sharpi Brooks and Caster 1956 (Fig. 5c, d). The shape of the posterior process is also reminiscent of what is known for the family, although it seems somewhat larger and more prominent than what is known in P. sharpi. All previously identified representatives of the family are from the Cambrian (e.g., Brooks and Caster 1956; Robison and Richards 1981; Yuan et al. 2011), so this would constitute a range extension for the group. The specimen does differ from typical Cambrian specimens of the family in not having the two valves joined along the hingeline. However, this may be analogous to the situation in phyllocarid crustaceans (e.g., Rode and Lieberman 2002; Briggs et al. 2004), or the Cambrian carapaced arthropod? Perspicaris dilatus (Kimmig and Pratt 2015), where some species are typically preserved with the two valves joined, yet others are represented by individual valves. Notably, the specimen differs from phyllocarid crustaceans in the presence of the prominent ventral process.
The prominent posterior process could also indicate a possible affinity to the bivalved arthropod Isoxys, but this genus usually does not preserve a ventral process (García-Bellido et al. 2009; Kimmig and Pratt 2015).
Another possible affinity might be the marrellid Furca (e.g., Van Roy et al. 2010; Legg 2016), which is known from deposits relatively close in space and time to the Valongo Formation. In this case, the fossil would either represent two very poorly preserved specimens, or one fractured specimen. In this case, what has been interpreted as the ventral process would instead be the projection coming off of the righthand side and center of a Furca carapace, and the interpreted posterior process would be the process coming off of the righthand side and posterior of the carapace.
A final possible affinity might be an arthropod appendage with a spine on each podomere.
Preservation
EDX analysis of Patanacta? and the wiwaxiid sclerites (Suppl. 3 and 4) identified carbon throughout but no consistent film, likely due to the metamorphic alteration of the rock and possibly to weathering on the scree slope (note a similar preservational style regarding carbon was found in Cambrian Discophyllum by Lieberman et al. 2017). While the sclerites are visually different, the elemental composition does not significantly differ from Patanacta? No sulfur was detected in the rock, suggesting that pyrite is not playing a role in mediating preservation. In addition to carbon, the bulk mineralogy of the specimens was determined to comprise aluminosilicates (SiAlO or SiFeAlO), likely muscovite (Kal2AlSi3O10(OH)2) and/or chlorite ((Mg, Fe, Al)6 (Al, Si)4O10(OH)8); examination via petrographic microscope revealed these were dispersed across the surrounding rock and the fossil. Spectral maps of Patanacta? (Suppl. 3) indicated the following variations in percentage by weight for different detectable elements: O, 40.2–47.1%; Si, 15.8–22.1%; Al, 14.8–16.3%; C 4.1–13.8%; Fe, 4.4–10.3%; K, 2.8–4.2%; Na, 0.7–0.9%; Mg, 0.4–0.6%; Ti, 0–0.8%; and P 0–0.3%. Although the Patanacta? specimen was found in scree, it appears that the elemental signature is primary. The fossil has an enhanced signature of carbon (from 6.7 at the edge to 13.8% near the center of the fossil) relative to the matrix (4.1–4.8%). Al, Si, O, Na, Mg, P, and Ti were found to be close to identical in the fossil and the surrounding matrix. It appears unlikely that the fossils are of inorganic nature, as no S was present in detectable levels during the EDX analysis, confirming that the fossils are no pyrite residue. Iron is slightly more prominent in the matrix (9.4–10.3%) than in the fossil (4.4–6.2%) of FCUP/DGAOT 3SPC, again arguing against a prominent role for pyrite in soft-bodied preservation.
In total, the EDX data indicate the specimens are originally preserved via carbonaceous films and aluminosilicate replacement, broken up by weathering and low temperature metamorphism. Scanning electron microscopy of the soft-bodied fossils revealed no pyrite framboids in the fossil or the surrounding matrix, but pyrite framboids can occur in some slate layers. There is no indication of phosphatisation in any of the soft-bodied or biomineralized fossils. The low temperature metamorphism (epizone) that overprinted the rocks (Couto 1993) might have altered part of the mineralogical composition of the host-rock and the fossils, but still the soft-bodied preservation in the upper Valongo Formation appears to have followed the standard path of BST preservation (Butterfield 1995; Orr et al. 1998; Cai et al. 2012; Gaines 2014).
Discussion
The upper Valongo Formation preserves relatively few taxa of soft-bodied fossils compared to many of the other Ordovician soft-body deposits (Fig. 2 and Suppl. 1), but it still adds to our knowledge of soft-bodied fossils as it represents the only deposit preserving such fossils of Dapingian–Darriwilian age from Gondwana, filling the gap between the Tremadocian–Floian Fezouata biota and the Sandbian fossils of the Tafilalt Biota (MacGabhann 2012; Gutiérrez-Marco and García-Bellido 2015; Van Roy et al. 2015). In addition, the upper part of the Valongo Formation represents one of the few deeper-water BST deposits in the Ordovician; finally, it is the first deposit preserving soft-bodied fossils from the Ordovician of the Iberian Peninsula.
In regard to the fossils, the best-preserved specimen is the discoidal fossils referred to Patanacta?. Discoidal fossils comprise an enigmatic, probably polyphyletic assemblage of organisms known from the late Neoproterozoic and throughout much of the Phanerozoic (Cartwright et al. 2007). They have been referred to many phyla including Cnidaria, Mollusca, and Echinodermata. One of the challenges of studying discoidal fossils is determining whether or not they are truly biogenic, a topic discussed in detail in Hofmann et al. (1991), Gehling et al. (2000), Ruiz et al. (2004), MacGabhann (2007), Kirkland et al. (2016), and Lieberman et al. (2017). Focusing on examples that appear to represent bona fide organic remains, soft-bodied discoidal fossils are particularly well known from the Ediacaran (Glaessner 1971; Fedonkin 1981; Sun 1986; Gehling et al. 2000; MacGabhann 2007; Tarhan et al. 2015; Lieberman et al. 2017). They are also known from the Phanerozoic as well, especially from the Cambrian (Masiak and Zylinska 1994; Waggoner and Collins 1995; Landing and Narbonne 1992; Hagadorn et al. 2002; Zhu et al. 2002; Van Roy 2006a, b; Cartwright et al. 2007; Young and Hagadorn 2010; Sappenfield et al. 2017), with fewer localities preserving these fossils known from the Ordovician (e.g., Ruedemann 1916; Yochelson 1984; Cherns 1994; MacGabhann and Murray 2010; Botting et al. 2015; Fig. 2a and Suppl. 1) and later time periods (e.g., Ossian 1973; Stanley and Kanie 1985; Kirkland et al. 2016).
Cherns (1994) considered P. pedina to be a medusoid of uncertain affinities, based on its concentric center, which she treated as the stomach, and its radial ridges, which could be part of the gastrovascular system. An interpretation as a medusozoan is not unreasonable, though currently there is much debate about the interpretation of discoidal fossils and their taxonomic affinities (Cartwright et al. 2007; MacGabhann 2007; Young and Hagadorn 2010; Botting et al. 2015; Tarhan et al. 2015; Kirkland et al. 2016; Lieberman et al. 2017; Landing et al. 2018). The absence of concentric rings in Patanacta was treated by Cherns (1994) as either reflecting taphonomic factors or the absence of actual biological structures. Notably, these are again absent in the specimen described herein, perhaps suggesting that this might not be a taphonomic feature. Other Ordovician discoidal fossils have been attributed to Medusozoa (Young et al. 2007, 2012; Lamsdell et al. 2017).
An alternative affinity for Patanacta would be as an eldoniid, although a diagnostic coiled sac is not visible. Eldoniids are discoidal soft-bodied fossils thought to represent stem group deuterostomes (Caron et al. 2010; MacGabhann 2012; Kimmig et al. 2018). Once thought to be limited to the Cambrian, now specimens from other periods have been reported (Alessandrello and Bracchi 2003; MacGabhann 2007, 2012). The eldoniids are characterized by branching radial ridges and a coiled sac (Alessandrello and Bracchi 2003; MacGabhann 2007, 2012); many specimens also preserve concentric rings, especially if Discophyllum is considered to be an eldoniid (see discussion in Lieberman et al. 2017). In the last two decades, several eldoniids have been described from the Ordovician, and they now appear to comprise the most common soft-bodied discoidal fossils from this time period (Table 1; Alessandrello and Bracchi 2003; MacGabhann 2012; Gutiérrez-Marco and García-Bellido 2015). MacGabhann (2012) in particular argued that specimens of Discophyllum (and other discoidal forms that resemble this genus such as Paropsonema) from the Ordovician and other time periods were likely to represent eldoniids. However, there still is active debate about whether these taxa might comprise eldoniids or instead porpitids (Lieberman et al. 2017; Landing et al. 2018). Discophyllum has also been reported from the Cambrian of California (Lieberman et al. 2017), the Ordovician of Morocco (MacGabhann 2012), the Ordovician of Portugal (Delgado 1892), the Ordovician of France (Phillips and Slater 1848; Barrois 1891), and the Silurian of England (Fryer and Stanley 2004). If all these specimens actually belong to Discophyllum, the genus is long lived and likely had a worldwide distribution. At this time, based on the available evidence in the literature and the new specimen presented herein, we build on Cherns’ (1994) interpretation and suggest Pantanacta is likely a medusozoan of some kind, though the precise class, i.e., hydrozoan or scyphozoan (there is no evidence for cubozoan affinity), is indeterminate. More, better preserved specimens are needed to verify and constrain its taxonomic affinities. Furthermore, the lack of concentric rings, the lack of branches on the radial ridges, and the lack of a prominent coiled sac in the specimens from the Valongo Formation and from Sweden suggest that at this time there is a paucity of character evidence supporting an eldoniid interpretation.
Conclusions
The discovery of soft-bodied fossils in the upper Valongo Formation extends the occurrence of deposits with soft-tissue preservation into the Dapingian-Darriwilian of northern Gondwana and closes a gap in the soft-bodied fossil record between the Tremadocian–Floian Fezouata biota and the Sandbian fossils of the First Bani Group in Morocco. Additionally, it expands the extent of BST deposits in deep-water settings into northern Gondwana. Moreover, it represents the first occurrence of soft-bodied fossils in Portugal and the first Ordovician soft-tissue preservation on the Iberian Peninsula. The discovery of the soft-bodied fossils in the upper Valongo Formation also shows that it is likely that many soft-bodied fossils in the Ordovician remain to be discovered, and a new look at deep-water shales and slates of this time period is warranted.
References
Aldridge RJ, Theron JN, Gabbott SE (1994) The Soom Shale: a unique Ordovician fossil horizon in South Africa. Geol Today 10:218–221
Alessandrello A, Bracchi G (2003) Eldonia berbera n. sp., a new species of the enigmatic genus Eldonia Walcott, 1911 from the Rawtheyan (Upper Ordovician) of Anti-Atlas (Erfoud, Tafilalt, Morocco). Atti della società italiana di scienze naturali e del museo civico di storia naturale di Milano 144:337–358
Allison PA, Briggs DEG (1993) Paleolatitudinal sampling bias, Phanerozoic species diversity, and the end-Permian extinction. Geology 21:65–68
Aris MJ, Palomo M (2014) Primer registro de una fauna Ordovícica ‘tipo Burgess Shale’ en Argentina y Sudamérica. XIX Congreso Geológico Argentino, Córdoba. Junio 2014, Asociación Geológica Argentina, Buenos Aires, Abstracts, S2–S4
Aris MJ, Corronca JA, Quinteros S, Pardo PL (2017) A new marrellomorph euarthropod from the Early Ordovician of Argentina. Acta Palaeontol Pol 62:1–8
Ausich WI, Sá A, Gutiérrez-Marco JC (2007) New and revised occurrences of Ordovician crinoids from southwestern Europe. J Paleontol 81:1374–1383
Babin C, Becq-Giraudon JF, Lardeux H, Gutiérrez-Marco JC (1996) Présence de Trocholites (Cephalopoda, Nautiloidea) dans L’Ordovicien du Massif Armoricain et du Portugal. Bulletin de la Société des Sciences de l’Ouest de la France, nouvelle série 18:105–112
Baliníski A, Sun Y (2013) Preservation of soft tissues in an Ordovician linguloid brachiopod from China. Acta Palaeontol Pol 58:115–120
Baliński A, Sun Y, Dzik J (2014) Probable advanced hydroid from the Early Ordovician of China. Palaeontol Z 88:1–10
Barrois C (1891) Mémoire sur la Faune du Grès armoricain. Annales de la Société Géologique du Nord 19:134–237
Botting JP, Muir LA, Jordan N, Upton C (2015) An Ordovician variation on Burgess Shale-type biotas. Sci Rep 5(9947):1–11. https://doi.org/10.1038/srep09947
Briggs DEG (2003) The role of decay and mineralization in the preservation of soft-bodied fossils. Annu Rev Earth Pl Sc 31:275–301
Briggs DEG, Fortey RAF (2005) Wonderful strife: systematics, stem groups, and the phylogenetic signal of the Cambrian radiation. Paleobiology 31:94–112
Briggs DEG, Bottrell SH, Raiswell R (1991) Pyritization of soft-bodied fossils: Beecher’s Trilobite Bed, Upper Ordovician, New York State. Geology 19:1221–1224
Briggs DEG, Sutton MD, Siveter DJ, Siveter DJ (2004) A new phyllocarid (Crustacea: Malacostraca) from the Silurian Fossil-Lagerstätte of Herefordshire, UK. Proc R Soc Lond B 271:131–138
Briggs DEG, Liu HP, McKay RM, Witzke BJ (2018) The Winneshiek biota: exceptionally well-preserved fossils in a Middle Ordovician impact crater. J Geol Soc London 175:865–874. https://doi.org/10.1144/jgs2018-101
Brooks HK, Caster KE (1956) Pseudoarctolepis sharpi n. gen. n. sp. (Phyllocarida) from the Wheeler Shale (Middle Cambrian) of Utah. J Paleontol 30:9–14
Budil P, Fatka O, Bruthansová J (2003) Trilobite fauna of the Šárka Formation at Praha–Červeny vrch Hill (Ordovician, Barrandian area, Czech Republic). Bull Geosci 78:113–117
Butterfield NJ (1990) Organic preservation of non-mineralizing organisms and the taphonomy of the Burgess Shale. Paleobiology 16:272–286
Butterfield NJ (1995) Secular distribution of Burgess-Shale-type preservation. Lethaia 28:1–13
Caron J-B, Conway Morris S, Shu D (2010) Tentaculate fossils from the Cambrian of Canada (British Columbia) and China (Yunnan) interpreted as primitive deuterostomes. PLoS One 5:1–13
Cai Y, Schiffbauer JD, Hua H, Xiao S (2012) Preservational modes in the Ediacaran Gaojiashan Lagerstätte: pyritization, aluminosilicification, and carbonaceous compression. Palaeogeograp Palaeoclimatol Palaeoecol 326-328:109117
Cartwright P, Halgedahl SL, Hendricks JR, Jarrard RD, Marques AC, Collins AG, Lieberman BS (2007) Exceptionally preserved jellyfishes from the Middle Cambrian. PLoS One 2:e1121. https://doi.org/10.1371/journal.pone.0001121
Cherns L (1994) A medusoid from the Late Ordovician or Early Silurian of Jamtland, Central Sweden. J Paleontol 68:716–721
Chlupáč I (1999) Some problematical arthropods from the Upper Ordovician Letná Formation of Bohemia. J Czech Geol Soc 44:79–92
Cloud P (1973) Pseudofossils: a plea for caution. Geology 1:123–127
Conway Morris S (1985) Cambrian Lagerstätten: Their distribution and significance. Philo Trans Royal Soc Ser B 311:49e65
Conway Morris S (1989) The persistence of Burgess Shale-type faunas: implications for the evolution of deeper-water faunas. Trans R Soc Edinb Earth Sci 80:271–283
Conway Morris S, Selden PA, Gunther G, Jamison PG (2015) New records of Burgess Shale-type taxa from the middle Cambrian of Utah. J Paleontol 89:411–423
Couto H (1993) As mineralizações de Sb-Au da região Dúrico-Beirã. 2 Vols. (Vol. Texto; Vol. Anexos: 32 Estampas e 7 Mapas). Unpublished PhD thesis, University of Porto:607 pp
Couto H (2013) The Ordovician of Valongo Anticline (Northern Portugal): state of art. Geology, Exploration and Mining, SGEM2013 Conference Proceedings 1:203–208
Couto H, Gutiérrez-Marco JC (1999) Nota sobre algunos Diplo porita (Echinodermata) de las pizarras de la Formación Valongo (Ordovicico Medio, Portugal). Temas Geológico-Mineros ITGE 26:541–545
Couto H, Gutiérrez-Marco JC (2000) Alguns fósseis raros ou pouco conhecidos na Formação de Valongo (Ordovícico Médio, norte de Portugal). In: Díez JB, Balbino AC (eds) I Congresso Ibérico de Paleontologia/XV Jornadas de la Sociedad Española de Paleontología, Évora, pp 191–192
Couto H, Roger G (2017) Palaeozoic Magmatism Associated with Gold-Antimony-Tin-Tungsten-Lead-Zinc and Silver Mineralization in the Neighbouring of Porto, Northern Portugal. IOP Conf Ser: Earth Environ Sci 95:022054
Couto H, Piçarra JM, Gutiérrez-Marco JC (1997) El Paleozoico del Anticlinal de Valongo (Portugal) In: Grandal d'Anglade A, Gutiérrez-Marco JC, Santos Fidalgo L (eds) Comunicaciones XIII Jornadas de Paleontología y V Reunón Internacional PIGC 351. A Coruña, Libro de Resúmenes y Excursiones. Sociedad Española de Paleontologia, Madrid, pp 270–290
Couto H, Knight J, Lourenço A (2013) Late Ordovician ice-marginal processes and sea-level change from the North Gondwana platform: evidence from the Valongo Anticline (northern Portugal). Palaeogeograph Palaeoclimat Palaeocl 375:1–15
Couto H, Knight J, Lourenço A (2014) Rifting at the Cambrian–Ordovician transition in northwestern Portugal. Comunicações Geológicas 101 Especial I:251–254
Curtis MLK (1961) Ordovician trilobites from the Valongo area, Portugal. Cheiruridae, Pliomeridae and Dionididae. Boletim da Sociedade Geológica de Portugal 14:1–16
Cuvier G (1797) Tableau élémentaire de l’histoire naturelle des animaux. Baudouin, Paris, 770 p
Daley AC, Antcliffe JB, Drage HB, Pates S (2018) Early fossil record of Euarthropoda and the Cambrian explosion. PNAS (USA) 115:5323–5331
Delgado JFN (1892) Contributions à l’étude des terrains anciens du Portugal. Communicações da Commissão dos Trabalhos Geologicos de Portugal 2:216–228
Delgado JFN (1897) Fauna Silúrica de Portugal. Novas observações acerca de Lichas (Uralichas) Ribeiroi. Commissão dos Trabalhos Geologicos de Portugal: 1-34
Delgado JFN (1908) Système Silurique du Portugal. Étude de stratigraphie paléontologique Mémoire de la Commission du Service Géologique du Portugal:1–245
Farrell ÚC, Martin MJ, Hagadorn JW, Whiteley T, Briggs DEG (2009) Beyond Beecher’s Trilobite Bed: widespread pyritization of soft tissues in the Late Ordovician Taconic foreland basin. Geology 37:907–910
Fedonkin MA (1981) Belomorskaya biota venda (The Vendian White Sea biota). Trudy Geological Institute, Academy Nauk SSSR 342:1–100
Fryer G, Stanley GD Jr (2004) A Silurian porpitoid hydrozoan from Cumbria, England, and a note on porpitoid relationships. Palaeontology 47:1109–1119. https://doi.org/10.1111/j.0031-0239.2004.00402.x
Gabbott SE (1998) Taphonomy of the Ordovician Soom Shale Lagerstätte: an example of soft tissue preservation in clay minerals. Palaeontology 41:631–667
Gabbott SE, Zalasiewicz J, Collins D (2008) Sedimentation of the phyllopod bed within the Cambrian Burgess Shale Formation of British Columbia. J Geol Soc Lond 165:307–318
Gabbott SE, Browning C, Theron JN, Whittle RJ (2017) The late Ordovician Soom Shale Lagerstätte: an extraordinary post-glacial fossil and sedimentary record. J Geol Soc Lond 174:1–9
Gaines RR (2014) Burgess Shale-type preservation and its distribution in space and time. In M. Laflamme, J.D. Schiffbauer, and S.A.F. Darroch (eds.), Reading and writing of the fossil record: preservational pathways to exceptional fossilization. Paleontol Soc Paps 20:123–146
Gaines RR, Hammarlund EU, Hou X, Qi C, Gabbott SE, Zhao Y, Peng J, Canfield DE (2012a) Mechanism for Burgess Shale-type preservation. PNAS (USA) 109:5180–5184
Gaines RR, Droser ML, Orr PJ, Garson D, Hammarlund E, Qi C, Canfield DE (2012b) Burgess Shale-type biotas were not entirely burrowed away. Geology 40:283–286
García-Bellido DC, Paterson JR, Edgecombe GD, Jago JB, Gehling JG, Lee MSY (2009) The bivalved arthropods Isoxys and Tuzoia with soft-part preservation from the lower Cambrian Emu Bay Shale Lagerstätte (Kangaroo Island, Australia). Palaeontology 52:1221–1241
Gehling JG, Narbonne GM, Anderson MM (2000) The first named Ediacaran body fossil, Aspidella terranovica. Palaeontology 43:427–456. https://doi.org/10.1111/j.0031-0239.2000.00134.x
Glaessner MF (1971) The genus Conomedusites Glaessner and Wade and the diversification of the Cnidaria. Palaeontol Z 45:1–17. https://doi.org/10.1007/BF02989670
Gutiérrez-Marco JC, García-Bellido DC (2015) Micrometric detail in palaeoscolecid worms from Late Ordovician sandstones of the Tafilalt Konservat-Lagerstätte, Morocco. Gondwana Res 28:875–881
Gutiérrez-Marco JC, Rábano I, Couto H, Piçarra JM (2000) Plumulítidos (Machaeridia) de la Formación Valongo (Ordovícico Médio, Portugal). Livro de resumos do I Congresso Ibérico de Paleontologia/XVI Jornadas de la Sociedad Española de Paleontología:193–194
Hagadorn JW, Dott RH Jr, Damrow D (2002) Stranded on a late Cambrian shoreline: medusae from central Wisconsin. Geology 30:147–150. https://doi.org/10.1130/0091-7613(2002)030<0147:SOALCS>2.0.CO;2
Hall J (1847) Paleontology of New York, Vol. 1: natural history of New York, part 6. New York: Albany, 1–338
Hearing TW, Legg DA, Botting JP, Muir LA, McDermott P, Faulkner S, Taylor AC, Brasier MD (2016) Surival of burgess Shale-type animals in a Middle Ordovician deep-water setting. J Geol Soc Lond 173:628–633
Hofmann HJ, Mountjoy EW, Teitz MW (1991) Ediacaran fossils and dubiofossils, Miette Group of Mount Fitzwilliam area, British Columbia. Can J Earth Sci 28:1541–1552. https://doi.org/10.1139/e91-138
Jago JB, Gatehouse CG, Powell CMA, Casey T, Alexander EM (2010) The Dawson Hill member of the Grindstone Range Sandstone in the Flinders Ranges, South Australia. Trans Roy Soc S Austral 134:115–124
Jago JB, Gehling JG, Paterson JR, Brock GA, Zang W (2012) Cambrian stratigraphy and biostratigraphy of the Flinders Ranges and the north coast of Kangaroo Island, South Australia. Episodes 35:247–255
Johnson H, Fox SK (1968) Dipleurozoa from the Lower Silurian of North America. Science 162:119–120
Kimmig J, Pratt BR (2015) Soft-bodied biota from the middle Cambrian (Drumian) Rockslide Formation, Mackenzie Mountains, northwestern Canada. J Paleontol 89:51–71
Kimmig J, Pratt BR (2016) Taphonomy of the middle Cambrian (Drumian) ravens Throat River Lagerstätte, Rockslide Formation, northwestern Canada. Lethaia 49:150–169
Kimmig J, Pratt BR (2018) Coprolites in the ravens throat river lagerstätte of northwestern Canada: implications for the middle Cambrian food web. Palaios 33:125–140. https://doi.org/10.2110/palo.2017.038
Kimmig J, Strotz LC (2017) Coprolites in middle Cambrian (Series 2–3) Burgess Shale-type deposits of Nevada and Utah and their ecological implications. Bull Geosci 92:297–309
Kimmig J, Meyer RC, Lieberman BS (2018) Herpetogaster from the early Cambrian of Nevada (Series 2, Stage 4) and its implications for the evolution of deuterostomes. Geol Mag 156:172–178. https://doi.org/10.1017/S0016756818000389
Kimmig J, Strotz LC, Kimmig SR, Egenhoff SO, Lieberman BS (2019) The Spence Shale Lagerstätte: an important window into Cambrian biodiversity. J Geol Soc London. https://doi.org/10.1144/jgs2018-195
Kirkland CL, MacGabhann BA, Kirkland BL, Daly JS (2016) Cryptic disc structures resembling Ediacaran discoidal fossils from the Lower Silurian Hellefjord Schist, Arctic Norway. PLoS One 11:e0164071. https://doi.org/10.1371/journal.pone.0164071
Lamsdell JC, LoDuca ST, Gunderson GO, Meyer RC, Briggs DEG (2017) A new Lagerstätte from the Late Ordovician Big Hill Formation, Upper Peninsula, Michigan. J Geol Soc Lond 174:18–22
Landing E, Narbonne GM (1992) Scenella and “A chondrophorine (medusoid hydrozoan) from the basal Cambrian (Placentian) of Newfoundland'”. J Paleontol 66:338–338. https://doi.org/10.1017/S0022336000033837
Landing E, Antcliffe JB, Geyer G, Kouchinsky A, Bowser SS, Andreas A (2018) Early evolution of colonial animals (Ediacaran Evolutionary Radiation-Cambrian Evolutionary Radiation-Great Ordovician Biodiversification interval). Earth-Sci Revs 178:105–135
Le Heron DP (2010) Trace fossils on a Late Ordovician glacially striated pavement in Algeria. Palaeogeograph Palaeoclimat Palaeocl 297:138–143
Legg DA (2016) A new marrellid arthropod from the Ordovician of Wales. Acta Palaeontol Pol 61:617–619
Lerosey-Aubril R, Gaines RR, Hegna TA, Ortega-Hernández J, Van Roy P, Kier C, Bonino E (2018) The Weeks Formation Konservat-Lager and the evolutionary transition of Cambrian marine life. J Geol Soc Lond 175:705–715. https://doi.org/10.1144/jgs2018-042
Lieberman BS, Kurkewicz R, Shinogle H, Kimmig J, MacGabhann BA (2017) Disc-shaped fossils resembling porpitids (Cnidaria: Hydrozoa) from the early Cambrian (Series 2: Stage 4) of western U.S.A. PeerJ 5(e3312):1–20. https://doi.org/10.7717/peerj.3312
Liu HP, McKay RM, Young JN, Witzke BJ, McVey KJ, Liu X (2006) A new Lagerstätte from the Middle Ordovician S. Peter Formation in northeast Iowa, USA. Geology 34:969–972
Liu HP, McKay RM, Witzke BJ, Briggs DEG (2009) The Winneshiek Lagerstätte and its depositional environments [in Chinese with English summary]. Geol J China Univs 15:285–295
MacGabhann BA (2007) Discoidal fossils of the Ediacaran biota: a review of current understanding. Geol Soc Spec Publ 286:297–313
MacGabhann BA (2012) A solution to Darwin's dilemma: differential taphonomy of Ediacaran and Palaeozoic non-mineralised discoidal fossils. PhD Thesis, National University of Ireland, Galway, 1–338. Available at https://aran.library.nuigalway.ie/bitstream/handle/10379/3406/2012MacGabhannPhDvol1.pdf?sequence=9
MacGabhann BA, Murray J (2010) Non-mineralised discoidal fossils from the Ordovician Bardahessiagh Formation, Co. Tyrone, Ireland. Irish J Earth Sci 28:1–12
Martin E, Pittet B, Gutiérrez-Marco JC, Vannier J, El Hariri K, Lerosey-Aubril R, Masrour M, Nowak H, Servais T, Vandenbroucke T, Van Roy P, Vaucher R, Lefebvre B (2016) The Lower Ordovician Fezouata Konservat-Lagerstätte from Morocco: age, environment and evolutionary perspectives. Gondwana Res 34:274–283
Masiak M, Zylinska A (1994) Burgess Shale-type fossils in Cambrian sandstones of the Holy Cross Mountains. Acta Palaeontol Pol 39:329–340
McKirdy DM, Hall PA, Nedin C, Halverson GP, Michaelsen BH, Jago JB, Gehling JG, Jenkins RJF (2011) Paleoredox status and thermal alteration of the lower Cambrian (Series 2) Emu Bay Shale Lagerstätte, South Australia. Aust J Earth Sci 58:259–272
Meyer MB, Ganis GR, Wittmer JM, Zalasiewicz JA, de Baets K (2018) A Late Ordovician planktic assemblage with exceptionally preserved soft-tissue problematica from the Martinsburg Formation, Pennsylvania. Palaios 33:36–46
Muscente AD, Schiffbauer JD, Broce J, Laflamme M, O’Donnell K, Boag TH, Meyer M, Hawkins AD, Huntley JW, McNamara M, MacKenzie LA, Stanley GD Jr, Hinman NW, Hofmann MH, Xiao S (2017) Exceptionally preserved fossil assemblages through geologic time and space. Gondwana Res 48:164–188
Neto de Carvalho C, Couto H, Figueiredo MV, Baucon A (2016) Microbial-related biogenic structures from the Middle Ordovician slates of Canelas (northern Portugal). Comunicações Geológicas 103:23–38
Orr PJ, Briggs DEG, Kearns SL (1998) Cambrian Burgess Shale animals replicated in clay minerals. Science 281:1173–1175
Ossian CR (1973) New Pennsylvanian scyphomedusan from western Iowa. J Paleontol 77:990–995
Pates S, Daley AC, Lieberman BS (2018) Hurdiid radiodontans from the middle Cambrian (Series 3) of Utah. J Paleontol 92:99–113
Petersen KW (1979) Development of coloniality in Hydrozoa. In: Larwood G, Rosen BR (eds) Biology and systematics of colonial animals. Academic Press, New York, pp 105–139
Phillips J, Slater JW (1848) Palaeontological appendix to Professor John Phillips' Memoir on the Malvern Hills compared with the Paleozoic districts of Abberley. Memoirs of the Geological Survey of Great Britain 2:331–386
Powell WG, Johnston PA, Collom CJ (2003) Geochemical evidence for oxygenated bottom waters during deposition of fossiliferous strata of the Burgess Shale Formation. Palaeogeograph Palaeocl 201:249–268
Rábano I (1989) El genero Uralichas Delgado, 1892 (Trilobita, Lichida) en al Ordovícico o de la Península Ibérica. Bol Geol Min 100:21–47
Rettalack GJ (2009) Cambrian-Ordovician non-marine fossils from South Australia. Alcheringa 33:355–391
Rettalack GJ (2015) Reassessment of the Silurian problematicum Rutgersella as another post-Ediacaran vendobiont. Alcheringa 39:573–588
Ribeiro A, Dias R, Pereira E, Merino H, Sodré Borges F, Noronha F, Marques M (1987) Guide-book for the Miranda do Douro-Porto excursion, paper presented at Conference on Plate Tectonics and Deformation, Univ. de Oviedo, Gijón-Oviedo, Spain
Robison RA, Richards BC (1981) Large bivalve arthropods from the middle Cambrian of Utah. U Kans Paleontol Contribs 106:1–28
Robison RA, Babcock LE, Gunther VG (2015) Exceptional Cambrian Fossils from Utah: A window into the age of trilobites. Utah Geological Survey, Miscellaneous Publications 15:97
Rode AL, Lieberman BS (2002) Phylogenetic and biogeographic analysis of Devonian phyllocarid crustaceans. J Paleontol 76:271–286
Romano M (1975) Harpid trilobites from the Ordovician of North Portugal. Comunicações Serviços geológicos de Portugal 59:27–36
Romano M (1976) The trilobite genus Placoparia from the Ordovician of the Valongo area, North Portugal. Geol Mag 113:11–28
Romano M (1980) The trilobite Eccoptochile from Ordovician of Northern Portugal. Palaeontology 23:605–616
Romano M (1982a) The Ordovician biostratigraphy of Portugal—a review with new data and re-appraisal. Geol J 17:89–110
Romano M (1982b) A revision of the Portuguese Odontopleuridae (Trilobita): Selenopeltis and Primaspis. Comunicações dos Serviços Geológicos de Portugal 68:213–223
Romano M (1990) The trilobite Protolloydohthus from the Middle Ordovician of North Portugal. Palaeontology 33:487–493
Romano M (1991) Trilobites from the Ordovician of Portugal. Palaeontology 34:329–355
Romano M, Diggens JN (1974) The stratigraphy and structure of Ordovician and associated rocks around Valongo, north Portugal. Comunicações dos Serviços Geológicos de Portugal 57:23–50
Romano M, Henry JL (1982) The trilobite genus Eoharpes from the Ordovician of Brittany and Portugal. Palaeontology 25:605–616
Ruedemann R (1916) Account of some new or little-known species of fossils. New York State Museum Bulletin 189:7–97
Ruiz JMG, Carnerup A, Christy AG, Wilhelm NJ, Hyde ST (2004) Morphology: an ambiguous indicator of biogenicity. Astrobiology 2:353–369
Sappenfield AR, Tarhan LG, Droser ML (2017) Earth's oldest jellyfish strandings: a unique taphonomic window or just another day at the beach? Geol Mag 154:859–874. https://doi.org/10.1017/S0016756816000443
Schiffbauer JD, Xiao S, Cai Y, Wallace AF, Hua H, Hunter J, Xu H, Peng Y, Kaufman AJ (2014) A unifying model for Neoproterozoic–Palaeozoic exceptional fossil preservation through pyritization and carbonaceous compression. Nat Commun 5:1–12. https://doi.org/10.1038/ncomms6754
Servais T, Harper DAT (2018) The Great Ordovician biodiversification event (GOBE): definition, concept and duration. Lethaia 51:151–164
Sperling EA, Balthasar U, Skovsted CB (2018) On the edge of exceptional preservation: Insights into the role of redox state in Burgess Shale–type taphonomic windows from the Mural Formation, Alberta, Canada, in Lyons, T.W. et al. Early Earth and the Rise of Complex Life: Emerging Topics in Life Sciences 2:311–323. https://doi.org/10.1042/ETLS20170163
Stanley GD, Kanie Y (1985) The first Mesozoic chondrophorine (medusoid hydrozoan) from the Lower Cretaceous of Japan. Palaeontology 28:101–109
Sun WG (1986) Precambrian medusoids: the Cyclomedusa-plexus and Cyclomedusa-like pseudofossils. Precambrian Res 31:325–360. https://doi.org/10.1016/0301-9268(86)90039-2
Tarhan LG, Droser ML, Gehling JG, Dzaugis MP (2015) Taphonomy and morphology of the Ediacara form genus Aspidella. Precambrian Res 257:124–136. https://doi.org/10.1016/j.precamres.2014.11.026
Tauber EH, Reis JM (1994) The trilobite Eoharpes cristatus Romano, 1975 from the Valongo Formation (Ordovician) of North Portugal. Comunicações do Instituto Geológico e Mineiro 80:35–49
Thadeu D (1949) Calimenídeos portugueses. Boletim da Socieadade Geológica de Portugal 8:129–134
Van Roy P (2006a) Non-trilobite arthropods from the Ordovician of Morocco. Unpublished PhD thesis, Ghent University, Ghent, 1–230
Van Roy P (2006b) An aglaspidid arthropod from the Late Ordovician of Morocco with remarks on the affinities and limitations of Aglaspidida. Trans R Soc Edinb Earth Sci 96:327–350
Van Roy P, Orr PJ, Botting JP, Muir LA, Vinther J, Lefebvre B, el Hariri K, Briggs DEG (2010) Ordovician faunas of Burgess Shale type. Nature 465:215–218
Van Roy P, Briggs DEG, Gaines RR (2015) The Fezouata fossils of Morocco; an extraordinary record of marine life in the Early Ordovician. J Geol Soc Lond 172:541–549. https://doi.org/10.1144/jgs2015-017
Vannier J, Chen J-Y (2005) Early Cambrian food chain: new evidence from fossil aggregates in the Maotianshan Shale biota, SW China. PALAIOS 20:3–26
Verrill AE (1865) Classification of polyps. Communications of the Essex Institute 4:145–152
von Siebold, C.T. (1848) Lehrbuch der vergleichenden Anatomie der Wirbellosen Thiere. Erster Theil, in von Siebold, C.T., and Stannius, H., eds., Lehrbuch der vergleichenden Anatomie: Berlin: Veit und Comp, 679 p
Waggoner BJ, Collins AG (1995) A new chondrophorine (Cnidaria, Hydrozoa) from the Cadiz Formation (Middle Cambrian) of California. Palaeontol Z 69:7–17. https://doi.org/10.1007/BF02985970
Webby BD, Cooper RA, Bergström SM, Paris F (2004) Stratigraphic framework and time slices. In: Webby BD, Paris F, Droser ML, Percival IG (eds) The great Ordovician biodiversification event. Columbia University Press, New York, pp 41–47
Yochelson EL (1984) North American Middle Ordovician Scenella and Macroscenella as possible chondrophorine coelenterates. Palaeontologr Am 54:148–153
Young GA, Hagadorn JW (2010) The fossil record of cnidarian medusae. Palaeoworld 19:212–221. https://doi.org/10.1016/j.palwor.2010.09.014
Young GA, Rudkin DM, Dobrzanski EP, Robson SP, Nowlan GS (2007) Exceptionally preserved Late Ordovician biotas from Manitoba, Canada. Geology 35:883–886
Young GA, Rudkin DM, Dobrzanski EP, Robson SP, Cuggy MB, Demski MW, Thompdon DP (2012) Great Canadian Lagerstätten 3. Late Ordovician Konservat-Lagerstätten in Manitoba. Geosci Can 39:201–213
Yuan J, Peng J, Zhao Y (2011) New bivalved arthropods from Mid-Cambrian Kaili Biota of southeastern Guizhou, southwest China. Acta Geol Sin 85:758–764
Zhu M-Y, Zhao Y-L, Chen J-Y (2002) Revision of the Cambrian discoidal animals Stellostomites eumorphus and Pararotadiscus guizhouensis from South China. Geobios 35:165–185
Acknowledgements
We would like to thank P. Thapa for his assistance using the SEM. P. Van Roy is thanked for his comments on a previous version of the manuscript. The editor, Allison Daley, William Ausich, and an anonymous referee are thanked for their comments. This is a contribution to ICT, Institute of Earth Sciences, Department of Geosciences, Environment and Spatial Planning, University of Porto, and to the IGCP Project 653.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by: Matthias Waltert
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
Table of Ordovician deposits with documented soft-tissue preservation, organized by age. (XLS 25 kb)
ESM 2
Table of Ordovician disc-shaped soft-bodied fossils. (XLSX 10 kb)
ESM 3
EDX spectrum and Elemental maps of Patanacta? sp. indet. (FCUP/DGAOT 3SPC). a Upper right of the specimen. b Center of the specimen, obscured by relief change. c The matrix. d Right of the center of the specimen. Locations based on Fig. 3. Scale bars are 1 mm. (PNG 2841 kb)
ESM 4
EDX spectrum and Elemental maps of the probable wiwaxiid sclerites (FCUP/DGAOT 5SPC). a Thin end of the right sclerite. b Wide end of the right sclerites. c The matrix. Locations based on Fig. 4. Scale bars are 1 mm. (PNG 3144 kb)
Rights and permissions
About this article

Cite this article
Kimmig, J., Couto, H., Leibach, W.W. et al. Soft-bodied fossils from the upper Valongo Formation (Middle Ordovician: Dapingian-Darriwilian) of northern Portugal. Sci Nat 106, 27 (2019). https://doi.org/10.1007/s00114-019-1623-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00114-019-1623-z