
Abstract
This study investigated density and some anatomical features, and also assessed the correlations among them in stem and branch woods of Entandrophragma cylindricum, Entandrophragma angolense and Khaya ivorensis. Branch and stem wood samples were sampled from two natural forest reserves. Wood density was determined in accordance with ISO 3131 whereas anatomical investigations followed IAWA committee’s recommendations-1989. Results indicated that at 5% level of significance, wood density was significantly higher in branchwood than stemwood for all three species. Generally for all the three species, significant differences were found in fibre length and vessel diameter between branchwood and stemwood of same species but no significant differences were found in both ray and axial parenchyma quantities. Stemwood density correlated significantly and positively with fibre (r = 0.408, p < 0.01) and ray parenchyma (r = 0.280, p < 0.05) proportions but negatively with fibre length (r = −0.528, p < 0.01), vessel lumen diameter (r = −0.304, p < 0.05) and axial parenchyma proportion (r = −0.679, p < 0.01). However, branchwood density correlated significantly and positively with fibre (r = 0.505; p < 0.01) and axial parenchyma (r = 0.385; p < 0.01) proportions but negatively with fibre length (r = −0.316, p < 0.05), vessel proportion (r = −0.486, p < 0.01) and ray parenchyma proportion (r = −0.357, p < 0.01). The study suggests that wood type (stem or branch) is an important factor to consider in terms of their effect on some properties of products produced from them, since differences in fibre length, vessel lumen diameter and vessel proportion could affect strength, porosity, leachability, natural durability and drying properties of wood.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
1 Introduction
The dwindling trend of timber resources globally appears to be threatening the continuous existence and expansion of wood industries and therefore, it has become almost critical that drastic changes are made in the pattern of raw material utilization without further delay. In Ghana, the forest is depleting at a faster rate of about 2% per annum (Ministry of Lands and Natural Resources—MLNR 2012). The worst hit trees are the most valued commercial timber species in the furniture export market such as Entandrophragma cylindricum (sapele), Entandrophragma angolense (edinam) and Khaya ivorensis (mahogany) (Dadzie 2011) which are mostly used for glulam, mouldings, architraves, and other tertiary wood products for building and interior architectural works, for both domestic and export markets (Pleydell 1994).
Regarding the apparent threat of wood raw material shortages on the continuous existence and possible expansion of wood and construction related industries, researchers are finding means of dealing with the problem of timber shortages. One step is the exploration of lesser utilized (LU) and lesser known (LK) tropical hardwood species. Researchers, such as Kumi-Woode (1996), Okai (1998), Oteng-Amoako et al. (1998) and Quartey (2009) investigated the potential of some LK and LU wood species to substitute some of the most preferred wood species. The more recent study by Quartey (2009) found that lesser utilized species such as Albizia ferruginea, Sterculia rhinopetala and Blighia sapida could be used as substitutes for some of the commercial hardwood species, in terms of natural durability and bending strength properties. However, findings from such research works appear not yet accepted by manufacturers and users of wood products to any appreciable level, since many stakeholders in the industry appear to be still concentrating on the premium and commercial species. Another alternative but potentially acceptable approach to increasing timber availability is the use of other tree parts like branches, off-cuts and roots (i.e. logging residues) of the already known and accepted commercial timber tree species. It is reported that when stem logs were abundant, the general notion about such residues was that they are inferior to stemwood in technical properties, but currently the utilization of logging residues has now turned out to be a viable alternative in many industrialized countries (Bhat et al. 1985). Thus exploring the utilization of branchwood of the commercial and premium species appears to be one immediate alternative and readily available solution to finding alternatives and supplements to stemwood to augment timber availability (Bhat et al. 1985). This is because, at least, the stemwood of the tree species is already known and materials from same species whose properties compare favourably with those of the already used stemwood may be accepted better than the lesser utilized and lesser known wood species. To this end, Okai (2002, 2003) has investigated some mechanical properties of Terminalia ivorensis (emire) and Aningeria robusta (asanfena) whereas Amoah et al. (2012) also worked on Terminalia ivorensis (emire) and Milicia excelsa (iroko) and found that MOE of the wood types of both species were comparable, whereas MOR of emire was significantly higher in branchwood than in stemwood. Further, Dadzie et al. (2016) have found kiln-dried branchwood of E. cylindricum to be naturally more durable than its stemwood counterpart.
Branchwood and stemwood of same trees can differ in density and anatomical structures as well as the relationship among them, since some kinds of cells are more abundant or less in wood from branches than wood from the main bole (Haygreen and Bowyer 1996; Wiedenhoeft and Miller 2005). These differences induce variability in wood which in turn is the single most important factor that detracts the performance of wood and therefore influences utilization of wood as a material (Desch and Dinwoodie 1996). For instance, high wood density is associated with thick fibre walls and a higher proportion of fibres which are also some of the very important qualities which contribute to strength and hence, in the absence of any other data about the properties of a particular species, wood density is used as a guide to its hygroscopicity, shrinkage and swelling, etc. and subsequent utilization (Shrivastava 2000).
However, wood density can be a complex trait and an expression of differential anatomical characteristics, especially in hardwoods where fibres and vessels are surrounded by other cells such as rays and parenchyma (Zhang and Zhong 1992) and therefore, understanding variation in anatomical properties of wood is imperative for good utilization (Zobel and van Buijtenen 1989).
Although a more recent study by Dadzie et al. (2016) found significant differences in some anatomical characteristics in stem and branch woods of Terminalia superba and Pterygota macrocarpa and another study by Dadzie and Amoah (2015) has also found some significant differences in density and some anatomical characteristics of stem and branch woods of sapele and mahogany, studies on anatomical characteristics of edinam and other anatomical characteristics such as fibre length, axial and ray parenchyma quantities of sapele, and mahogany and their influence on density of branchwood, both alone or in comparison with their stemwood counterparts have not been sighted and therefore appear limited or unavailable. The interest in edinam in addition to sapele and mahogany stems from three reasons. First, these three hardwood species are among the commercial ‘redwood’ timber species in the natural forests of Ghana, which are mostly preferred to be used together as ‘mixed redwoods’ for the production of lumber, plywood, furniture and finger-jointed products for both local and export markets (Pleydell 1994) and which are fast depleting (MLNR 2012). Second, branch woods of sapele, edinam and mahogany constitute a significant proportion of their merchantable volume (Dadzie 2013; Dadzie and Amoah 2014). Finally, to the authors` knowledge, there are scarce studies devoted to investigating the anatomical characteristics of branchwood of the wood species, in spite of the high monetary values of the stemwood of the species (Dadzie et al. 2014). However, in addition to the above mentioned recent studies some appreciable number of studies on the anatomical data of the stemwood rather than branchwood of the species has been carried out (Ayensu and Bentum 1974; Richter and Dallwitz 2000; Kémeuzé 2008; Tchinda 2008; Lemmens 2008; Oteng-Amoako et al. 2008; Forest Products Laboratory 2010).
It is in the light of the above reasons that this study of expanded comparative anatomical characteristics investigations of stem and branch woods of the three species and how they influence density is necessary.
2 Materials and methods
2.1 Samples collection and preparation
Stem and branch wood of each species were extracted from six trees from two natural forest reserves within two ecological zones of Ghana (three trees of each species from each forest reserve). Because samples were from natural forests, the age of the trees was unknown. However, the average diameter at breast height (dbh) of the species was 1200 mm for sapele, 1080 mm for edinam and 1120 mm for mahogany. The forest reserves were Asukawkaw reserve at Awronsua near Nkawkaw in the Eastern Region which is within the boundaries of longitude 0° 1° W and latitude 6° 7° N (a Moist Semi-Deciduous-South East type forest) and Suie reserve at Nsawora near Sefwi Wiawso in the Western Region which is within the boundaries of longitude 2° 3° W and latitude 6° 7° N (a Moist Evergreen forest). The reserves are concessions of Logs and Lumber Limited (LLL)—a timber processing firm in Kumasi, Ghana, and used for this study. In all, 36 branch logs (i.e. 2 branches × 6 trees × 3 species) with diameters ranging from 300 to 520 mm and lengths from 1.5 to 2 m were extracted for the study. Stemwood samples were also obtained from a total of 36 sections of stem logs (i.e. 2 sections × 6 trees × 3 species). All stem and branch wood samples were conveyed from the forests to LLL for processing. Both through-and-through and quarter sawn methods were used for the conversion of the logs to lumber using the same vertical bandmills as used for the conversion of logs in the factory. The logs were initially processed into rough lumber boards of 25 mm thickness with varied widths and to lengths corresponding to the lengths of the various log samples. After conversion, both branch and stem boards were re-sawn and crosscut into dimensions of 25 mm × 60 mm × 420 mm and both stem and branch wood grouped into 36 each according to the three species and the two reserves from which they were obtained, hence obtaining 72 groups (i.e. 2 wood types × 3 species × 6 trees × 2 reserves). Clear heartwoods were then sampled from each group (i.e. samples free of knots, fuzzy and woolly surfaces which are the evidences of reaction wood and any other visible defects). All samples were conditioned in an air-drying shed of LLL to an average MC of 12 ± 3% at same temperature and relative humidity as used by the company to air-dry wood.
2.2 Data collection
2.2.1 Density of stemwood and branchwood
For determining density, all samples were weighed using an electronic balance with accuracy of 0.01 g to note their masses, whereas their dimensions were also determined with an electronic vernier caliper with accuracy of 0.1 mm as specified in ISO 3131 at 12 ± 3% air-dry moisture content. Volumes and density (measured as \(\frac{{{mass}}}{{{volume}}}\)) were determined in accordance with ISO 3131 (1975). In all, a total of 432 samples of stem and branch woods were used for density estimations {i.e. (6 samples × 2 stem logs × 3 species × 3 trees × 2 sites = 216) + (6 samples × 2 branch logs × 3 species × 3 trees × 2 sites = 216)}. Moisture contents were measured with resistance type moisture meter (MO210 designed to measure MC of wood up to 44% as specified by manufacturers) and which has been found to have accuracy of ±2% upon validation with oven-dry method (Dadzie and Amoah 2015; Dadzie et al. 2016). The use of moisture meter, including resistance type, is one of the acceptable means to measure MCs in wood properties studies as has been done by some researchers, including Beaulieu et al. (1987), Ayarkwa et al. (2000), and Amoah et al. (2012).
2.2.2 Anatomical features
Four 20 mm cubes subsamples were prepared from each of the six trees from both sites/forest reserves for each wood type and species totalling 144 cubes (i.e. 4 replicates × 2 wood types × 3 trees × 2 sites × 3 species = 144). Samples were softened before sectioning with a sliding microtome, by placing in water for 21 days followed by soaking in a mixture of ethanol and glycerol, at a ratio of 1:1 for a period of 21–30 days depending on species.
Thin sections of 25 μm thickness were cut from transverse surfaces of the samples using a sliding microtome. The sections were first washed in distilled water and then stained in 1% safranin in 50% ethanol solution for about 10–15 min. Afterwards, the sections were rewashed in distilled water and dehydrated in increasing concentration of ethanol from 30, 50, 70, 80, 90 and 100% for 5–10 min. They were then immersed in xylene to remove little traces of water. The sections were then finally mounted permanently in Canada balsam after which the slides were dried in an oven at 60 °C overnight.
For maceration process, two match-stick sized specimens were plucked from each of the anatomical subsamples. These match-stick sizes were placed in separate labelled containers and immersed in a mixture of glacial acetic acid and hydrogen peroxide (6%) prepared at a ratio of 1:1. The specimens in the solution were incubated in an oven at 60 °C till complete maceration was attained. Macerated cells were temporarily mounted in glycerol for measurements of fibre lengths.
Photomicrographs were taken from the sections and macerated slides separately at 40∇ magnification using light microscope (Micromaster Premier) with a digital camera. All measurements of anatomical tissues were done manually on the photomicrographs using imageJ software (National Institute of Health, Bethesda, MD, USA). In all, for both stem and branch wood anatomy a total of 288 photomicrographs with image sizes of 682 × 512 pixels were used (i.e. 2 replicates × 4 cubes × 2 wood types × 3 species × 3 trees × 2 sites; for sections) and same quantities were used for the macerates for fibre length measurements. Fibre lengths and vessel diameters were manually determined using the straight line method in imageJ (Fig. 1a, b). In determining the proportions of the 3 main hardwood tissues [vessel, fibres, parenchyma (ray and axial)], the micrographs were inclined at 45° in imageJ with reference to the ray parenchyma orientation. Thereafter, line scale grids with 30 intersecting points were randomly placed on the micrographs of transverse sections. The number of intersects that fell on each particular tissue type were counted using the cell counter in imageJ. The number of counts obtained by each tissue was expressed as percentages of the total of 30 intersecting points of the grid area (Fig. 1c). For both wood types and all 3 species, vessel diameter, fibre length and the proportions of the tissues were estimated by taking a total of 2880 measurements (i.e. 10 measurements × 2 replicates × 4 cubes × 3 species × 3 trees × 2 sites; for macerates and same quantities for sections). This implies that each anatomical element of each wood type for each species was estimated from averages of 480 measurements {i.e. 2880/(2 wood types × 3 species)}. Wood anatomical property characterization and terminology were done in accordance with IAWA committee’s recommendations for hardwoods (IAWA Committee 1989).

Measurements of tissues dimensions and proportions using straight line and line scale grid in ImageJ; a fibre length measurement, b vessel diameter measurement, and c tissue proportion determination
2.3 Data analyses
Both descriptive and inferential statistics, and also regression analyses were performed using SPSS 17.0 version to analyze the data obtained. The descriptive statistical analysis was conducted to obtain means, standard deviations, and percentages of the measured properties. Inferential statistical analyses (ANOVA, Tuckey test and T test) were also carried out. Analysis of Variance ANOVA and Tuckey-multiple comparison test were used to ascertain the differences in the measured properties in either stem or branch woods among the species (within species variations). The independent sample T test was however conducted to evaluate the differences in the properties between stem and branch woods of same species (within tree variations). Differences among all mean values were tested at 95% confidence level. Regression analyses were also performed to ascertain the relationships among density of stem and branch wood and their respective anatomical properties.
3 Results and discussions
3.1 Density of stemwood and branchwood
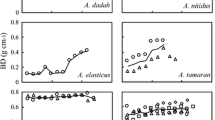
For all three species, branchwood exhibited relatively higher density than their stemwood (Fig. 2). Expectedly, the species with the highest stemwood density (sapele) also had the highest branchwood density, whereas the one with the lowest stemwood density (mahogany) also had the lowest branchwood density. Generally, branchwood density at 12 ± 3%MC was 15.88%, 30.38% and 15.74% higher than their stemwood counterparts for sapele, edinam and mahogany, respectively and these differences are significant (p < 0.05) (Fig. 1). Hence, within the same species, stem and branch wood density were significantly different for sapele (T = −5.397; p < 0.01), edinam (T = −12.240; p < 0.01) and mahogany (T = −7.482; p < 0.01). There also were significant differences in stemwood density (F = 55.886; p = 0.000) and branchwood density (F = 76.773; p = 0.000) among the three species (Fig. 2).

Density of stem and branch wood of the three test species measured at 12 ± 3%mc. Means with same letters are significantly different (p < 0.05): small letters wood type (stem or branch) effect, capital letters wood species effects. NB: N = 216; error bars are SE
Density is reported to be one of the most important properties that determines most properties of wood (e.g. bending strength, compression strength, natural durability, pulp yield, joint efficiencies in finger-jointed lumber), but it is highly influenced by the anatomical characteristics of the wood (Panshin and de Zeeuw 1980; Tsoumis 1991; Haygreen and Bowyer 1996; Desch and Dinwoodie 1996), hence, the findings in this study are informative. However, it is important to indicate that the mean density values for stemwood of Entandrophragma cylindricum (sapele), Entandrophragma angolense (edinam) and Khaya ivorensis (mahogany) were respectively 0.8, 0.6 and 0.4% higher than their values reported previously (Pleydell 1994). It is however worth to note that the stemwood densities found in this study appear to be within the range of other findings previously reported by researchers such as Ayensu and Bentum (1974), Tchinda (2008), Lemmens (2008), Kémeuzé (2008), Chudnoff (1984), and Richter and Dallwitz (2000) on density of the studied species. The findings in this study also appear to confirm that whereas softwood branchwood generally has densities ranging from 5 to 20% lower than their stemwood counterparts, hardwood branchwood has densities that range from higher in some species to lower or the same in others (Haygreen and Bowyer 1996; Bhat et al. 1985; Okai 2002, 2003; Gurau et al. 2008; Samariha et al. 2011; Amoah et al. 2012; Dadzie and Amoah 2015; Dadzie et al. 2016). Branchwood densities being higher than their counterpart stemwood in most hardwoods of same species are reported to be attributable to differences in growth rates of branch and stem woods. Branchwood grows more slowly than stemwood and this usually results in shorter cells with thicker cell walls (Fegel 1941; Patel 1970; Jane 1970) that subsequently lead to high density. Additionally, branchwood is known to contain reaction wood which also results in higher density than in stemwood (Tsoumis 1991).
3.2 Anatomical features and their correlations with density in stem and branch woods
Upon visual inspection of micrographs of transverse sections and fibres of the species (Figs. 3, 4, 5), some similarities and marginal differences in both arrangements and sizes of some wood cells in stem relative to branch woods of the same species were observed. These qualitative assessments relative to each species are presented as follows:

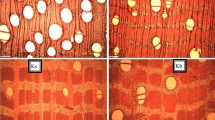
Sections and fibres of stem and branch woods of Entandrophragma cylindricum (sapele); E.C ai = stemwood cross section, E.C aii = stemwood tangential section, E.C aiii = stemwood fibres, E.C bi = branchwood cross section E.C bii = branchwood tangential section and E.C biii = branchwood fibres. Scale bar 200 μm

Sections and fibres of stem and branch woods of Entandrophragma angolense (edinam); E.A ai = stemwood cross section, E.A aii = stemwood tangential section, E.A aiii = stemwood fibres, E.A bi = branchwood cross section, E.A bii = branchwood tangential section and E.A biii = branchwood fibres. Scale bar 200 μm

Sections and fibres of stem and branch woods of Khaya ivorensis (mahogany); K.I ai = stemwood cross section, K.I aii = stemwood tangential section, K.I aiii = Stemwood fibres, K.I bi = branchwood cross section, K.I bii = branchwood tangential section and K.I. biii = branchwood fibres. Scale bar 200 μm
Entandrophragma cylindricum (sapele) Vessels in stem and branch wood were observed to be partly solitary and partly in radial multiples of 2–4 with rounded outlines and some occluded with some deposits. Axial parenchyma appears to be predominantly confluent paratracheal and seemingly marginal bands in both stem and branch wood (Fig. 3E.C ai and bi).
Entandrophragma angolense (edinam) Vessels in both stem and branch wood of edinam were found to be partly solitary and partly in radial multiples of 2–4 with rounded outlines and some are occluded with some deposits (Fig. 4E.A ai and bi). Also, axial parenchyma in both stemwood and branchwood appeared to be predominantly vasicentric. Some confluent parenchyma were also visible in both stem and branch woods.
Khaya ivorensis (mahogany) Vessels in stemwood were found to be predominantly solitary compared to vessels in branchwood but all appear to be partly in radial multiples of 2–4 with rounded outlines and having some deposits.
The observations made on the micrographs in this study about stemwood appeared to compare favourably with reports in literature about the species (Chudnoff 1984; Kémeuzé 2008; Lemmens 2008; Tchinda 2008). However, though descriptions of the anatomical features of branchwood of the studied species appear limited or unavailable and therefore not sighted, by inferring from and extending the consistencies of the findings on stemwood by findings in aforementioned literature, it could be said that the descriptions obtained in this study for the branchwood of the species also represent the true features of branchwood anatomy of the species.
Generally, quantitatively, it appeared that branchwood of the species exhibited relatively lower mean values of fibre length (μm), vessel lumen diameter (μm), vessel proportion (%), and axial parenchyma proportion (%) than their stemwood counterparts (Table 1). However, branchwood fibre proportion (%) and ray parenchyma proportion (%) appeared to be higher than those in their stemwood counterparts. The exception in these trends is mahogany in which the branchwood exhibited higher fibre length (μm), higher vessels proportion (%), higher ray and axial parenchyma proportions (%), but rather lower fibre proportion (%) than its stemwood counterpart. These general trends and dimensions of the anatomical features however, appear to agree with earlier research findings including those of Stokke and Manwiller (1994); Haygreen and Bowyer (1996), and Samariha et al. (2011) that branchwood has relatively shorter fibres and smaller vessel lumen diameter than their stemwood counterparts in most hardwoods.
Again at 5% significance level, there were significant differences in stemwood vessel lumen diameter (F = 11.016; p = 0.000), vessels proportion (F = 9.066; p = 0.000), fibre proportion (F = 4.577; p = 0.012), ray parenchyma proportion (F = 21.829; p = 0.000) and axial parenchyma proportion (F = 27.867; p = 0.000) among the three species (Table 1). There were also significant differences in branchwood fibre length (F = 10.089; p = 0.000), vessel lumen diameter (F = 2.837; p = 0.062), vessels proportion (F = 7.798; p = 0.001), ray parenchyma proportion (F = 4.630; p = 0.011) and axial parenchyma proportion (F = 11.961; p = 0.000) among the three species. However, there were no significant differences in stemwood fibre lengths (F = 2.118; p = 0.124), branchwood fibre proportion (F = 2.136; p = 0.122), and both stem and branch wood total parenchyma proportion among the three species. Moreover generally, within the same species, stem and branch wood ray, axial and total parenchyma proportions were not significantly different (Table 1). However, this study recorded significant differences in stem and branch wood fibre length (T = 4.308; p < 0.01-for sapele: T = 5.557; p < 0.01-for edinam: and T = −5.946-for mahogany), vessels lumen diameter (T = 1.928; p < 0.1-for sapele: T = 3.936; p < 0.01-for edinam: and T = 1.852; p < 0.1 for mahogany), and vessel proportion (T = 2.973; p < 0.05-for sapele: and T = −4.357; p < 0.01- for mahogany). The generally significant decrease in vessel lumen diameter and significant decrease in fibre lengths in branchwood of the species especially for sapele and edinam (Table 1) could have contributed significantly to the significant increase in density of branchwood of the species as could be observed in Fig. 2.
Shorter fibres in wood imply slow growth of the wood and subsequent thicker fiber cell walls which in addition to narrow vessel diameter produce wood with relatively higher density (Fegel 1941; Patel 1970; Jane 1970). In contrast, the significantly shorter fibre length, decrease in percent vessel area in addition to significant increase in percent fibre area in stemwood of mahogany did not result in high stemwood density compared to the density of its branchwood counterpart. It is worth to note that on average generally, the vessels diameter was significantly narrower in branchwood than in stemwood of mahogany (though some vessels in branchwood appeared visually similar in size to those in stemwood). This finding apparently suggests that the effects that fibre length and percent fibre proortion could have had on density of the stemwood might have been neutralized by the significant increase in vessels diameters. It is also important to recognize that the numerous deposits or occlusions in mahogany branchwood vessels compared to its stemwood (Fig. 4) could have also contributed to the significantly higher density of the branchwood though they could pose treatability difficulties (Desch and Dinwoodie 1996). These differences in vessel lumen diameter, vessel proportions, and fibre proportions (Table 1) could result in porosity, shrinkage, natural durability, permeability and treatability differences between the stem and the branch woods of the species (Desch and Dinwoodie 1996; Haygreen and Bowyer 1996; Ncube 2010; Skadsen 2007; Tsoumis 1991). Therefore, some level of caution may be exercised if the wood is to be dried or treated with preservative.
Moreover, in this study, total (axial + ray) parenchyma was not significantly different among both stemwood (F = 2.020; p = 0.138) and branchwood (F = 0.926; p = 0.399) of all three species (Table 1). Because parenchyma cells are carbohydrate storage sites in wood and as such they are the first points of attack by biological agents of wood destruction (Eaton and Hale 1993; Haygreen and Bowyer 1996), presupposes that the combined effects of axial and radial parenchyma among the three species are not different and as such, at same conditions and with the same biological agents, there would be no significant difference in their level of destruction. This appears to support literature that stemwood of all these three studied species is moderately durable (Pleydell 1994). It is however worth noting that parenchyma cells have some advantages in furniture and other tertiary wood products that mostly need application of finishes. The presence of ray and axial parenchyma in wood contributes enormously to give wood some beautiful figure that results in relatively nice appearance upon application of finishes to furniture and other tertiary wood products made from them (Walton 1974; Desh and Dinwoodie 1996). This also appears to be a contributing factor that has made the three species most preferred for furniture, interior panelling and other tertiary products manufacturing in Ghana for both local consumption and exports (Pleydell 1994; Dadzie et al. 2014).
Tables 2 and 3 report the correlation matrixes of the relationships among density and the various anatomical features of stemwood, and those among density and the anatomical features in branchwood.
Except vessel proportion (%) in stemwod, and vessel lumen diameter and total parenchyma proportion (%) in branchwood, the rest of the selected anatomical properties significantly (i.e. either p < 0.01 or p < 0.05) influenced density. However, these influences were positive for some and inverse for others. It however, appeared generally that the correlation between density and the selected anatomical features was either moderate or weak. Further, some same anatomical features had relatively higher correlation coefficients in stemwood than in branchwood while others had higher coefficients in branchwood than stemwood, but with varied levels of significance (Tables 2, 3). For instance fibre length correlated moderately but negatively with stemwood density (r = −0.528, P < 0.01) compared to branchwood (r = −0.316, P < 0.05). Moreover it was found that only vessel proportion correlated positively but insignificantly (r = 0.105, p > 0.05) with stemwood density but inversely and significantly (r = −0.486, p < 0.01) with branchwood density, the rest of the anatomical properties correlated in similar directions to density of both stem and branch woods. These findings appear to corroborate a report by Zobel and Jett (1995) that in hardwoods, vessel proportion, ray proportion, fibre length and parenchyma cells affect wood density. Samariha et al. (2011) have also reported that short fibres tend to give a dense and uniform sheet structure in wood. Therefore, the relatively higher density of branchwood found in this present study could be attributable to their relatively shorter fibres. This finding also appears to support the negative correlation of fibre length to the density of both stem and branch wood found in this study. On the contrary, a weak but positive correlation has been found between fibre length and oven-dry density of some Ghanaian tropical hardwoods (Okoh 2014).
It is therefore important to note that opinions of researchers are reported to be divided regarding the correlations of density with anatomical properties in different species of wood (Sreevani and Rao 2014). For instance Taylor (1973) reported that density increases with increases in fibre proportion and decreases with increases in parenchyma proportion in Eucalyptus, but on the contrary, Sreevani and Rao (2014) found that density of Eucalyptus is weakly influenced by fibre proportion and rather increases with increases in its parenchyma proportion. However, findings in this present study appear to support Taylor (1973) rather than Sreevani and Rao (2014). Grzeskowiak, et al. (2000) also found that density is positively influenced by vessel percentage which is also in conflict with the findings by Sreevani and Rao (2014) that density is strongly but rather negatively influenced by vessel percentage.
The foregoing findings in literature appear to paint some picture of complexity of the nature of the interrelationships existing between wood density and its anatomical features. It is therefore not strange to find in this present study that percentage vessel and ray parenchyma proportions correlated weakly and positively with stemwood density whereas they correlated moderately but negatively with branchwood density. Moreover, axial parenchyma proportion correlated moderately with both stems and branch wood density, but whereas the correlation was positive in branchwood it was negative in stemwood.
4 Conclusion
This study sought to characterize and compare the density and some anatomical features of stem and branch wood of Entandrophragma cylindricum (sapele), Entandrophragma angolense (edinam) and Khaya ivorensis (mahogany), and also determine the influence of the anatomical features of stem and branch wood on their respective densities. The three wood species are among the most preferred wood species used for furniture and other tertiary wood products for both local and international markets. Therefore, the expectation was that findings from this study will provide information that could inspire confidence and generate interest in the use of branchwood of the species as supplementary materials for wood products manufacturing in the face of the continuous depletion of timber resources in Ghana. From the results obtained in this study, the following conclusions were drawn;
-
1.
Branchwood of sapele, edinam and mahogany exhibited higher density than their stemwood counterparts. Among the three studied species, the one with high stemwood density (sapele) expectedly had high branchwood density, whereas the one with lower stemwood density also had the lowest branchwood density.
-
2.
The results showed that branchwood of all three species exhibited shorter fibre lengths and smaller vessel lumen diameters than their respective stemwood counterparts. Banchwood of the species has also been found to possess relatively higher fibre and total parenchyma percentages than their respective stemwood counterparts. Though total parenchyma percentage may have some implication on natural durability, the finding that the difference between stem and branch wood of same species was not significantly different could mean that, all things being equal, their natural durability could also not be much different. Further, fibre length, vessel lumen diameter and total parenchyma proportions strongly but negatively influence density of both stem and branch wood and this was consistent with findings reported in literature.
-
3.
The study has also established that strong positive and significant (i.e. at p < 0.01 or p < 0.05) relationships exist between fibre percentage and density of both stem and branch wood. Moreover, it appeared that except vessel proportion in the case of stemwood, and vessel lumen diameter in the case of branchwood all the anatomical features determined in this study could be reliable predictors of stem and branch wood density of sapele, edinam and mahogany.
References
Amoah M, Appiah-Yeboahand J, Okai R (2012) Characterisation of physical and mechanical properties of Branch, Stem and Root wood of Iroko and Emire Tropical Trees. Res. J Appl Sci Eng Technol 4(12):1754–1760
Ayarkwa J, Hirashima Y, Sasaki Y (2000) Effects of finger geometry and end pressure on the flexural properties of finger-jointed tropical African hardwoods. Forest Prod J 50(11/12):53–63
Ayensu E S, Bentum A (1974) Commercial trees of West Africa, City of Washington, p 68
Beaulieu C, Verreault C, Gosme C, Samson M (1987) Experimental assessment of the effect of length on the tensile strength of structural finger-jointed lumber. For Prod J 47(10):94–100
Bhat KM, Bhat KV, Dhamodaran TK (1985) Wood and Bark properties of branches of selected tree species growing in Kerala, KFRI research report No. 29, Kerala Forest Research Institute, Peechi, Thrissur, p 34
Chudnoff M (1984) Tropical Timber of the world, Agricultural handbook 607. U.S. Department of Agriculture, Forest Service Washington D.C., p 466
Dadzie PK (2011) Value addition to wood resources, the case of garden furniture production and export in Ghana-study of a local firm, conference paper presented at the 65th international convention of the Forest Products Society (F.P.S), Portland, Oregon, USA. 19th–21st June. http://www.slideshare.net/julielang/slideshows/2. Accessed 20 June 2013
Dadzie PK (2013) The potential contribution of branchwood quantity, left after logging operations towards reducing depletion rate and preserving Ghana’s forest ecosystem. Am J Agric For 1(2):32–39
Dadzie PK, Amoah M (2014) Timber merchantable residue quantities and harvesting efficiency in tropical forests of Ghana; drivers of wood residue utilization for forest conservation. Glob J Sci Front Res H Environ Earth Sci 14(5/10):33–46
Dadzie PK, Amoah M (2015) Density, some anatomical properties and natural durability of stem and branch wood of two tropical hardwood species for ground applications. Eur J Wood Prod 73:759–773
Dadzie PK, Amoah M, Tekpetey SL (2014) Preliminary assessment of wealth creation in wood products’ business in Ghana: the perspective of lumber and furniture production and implications for entrepreneurship. Int J Bus Econ Res 3(6):243–249
Dadzie PK, Amoah M, Frimpong-Mensah K, Sheldon QS (2016) Comparison of density and selected microscopic characteristics of stem and branch wood of two commercial trees in Ghana. Wood Sci Technol 50(1):91–104
Desch HE, Dinwodie JM (1996) Timber structure, properties, conversion and use, 7th edn. Macmillan Press, London, p 306
Eaton RA, Hale MDC (1993) Wood decay, pest and protection. In: Quartey GA, Zurcher E, Frimpong-Mensah K (2008) Durability investigations of lesser-known Ghanaian wood species, a Conference Paper, International Research Conference on Lesser Known Timber Species for Bridge Construction, 8th–11th Sept, pp 41–45
Fegel AC (1941) Comparative anatomy and varying physical properties of Trunk, Branch and Root wood in certain Northeastern trees. New York State College of Forestry, Technology Bulletin No. 55
Forest Products Laboratory (2010) Wood as an engineering material centennial edition, general technical report FPL—GTR—190; U.S. Department of Agricultural Service, Madison, Wisconsin, p 509
Grzeskowiak V, Turner OP, Megown RA (2000) The use of densitometry and image analysis techniques to predict pulp strength properties in Eucalyptus plantations. Conference paper presented at TAPPSA conference ‘African paper week 2000 and beyond’ Durban, South Africa, pp 1–18
Gurau L, Cionca M, Mansfield-Williams H, Sawyer G, Zeleniuc O (2008) Comparison of the mechanical properties of branch and stem wood for three species. Wood Fibre Sci 40(4):647–656
Haygreen JG, Bowyer JL (1996) Forest products and wood science—an introduction, 3rd edn. IOWA State University Press, Ames, p 485
IAWA Committee (1989) IAWA list of microscopic features for hardwood identification with appendix on non-anatomical information. In: Wheeler EA, Bass P, Gasson P (eds)
ISO 3131 (1975) International standard; wood-determination of density for physical and mechanical tests, 1st edn. International organisation for standardisation, UK
Jane FW (1970) The structure of wood, London. A & C Black, London, p 420
Kémeuzé VA (2008) Entandrophragma cylindrium (Sprague) [internet]. Record from PROTA4U. In: Louppe D, Oteng-Amoako AA, Brink M (eds) PROTA (plant resources of Tropical Africa, Wageningen, Netherlands (http://www.prota4u.org/search-asp). Accessed 9 April 2014
Kumi-Woode BG (1996) Natural Decay resistance of some Ghanaian timbers and wood decay hazard potential for Ghana. In: Quartey G (2009) Relationships between some anatomical, physical and durability properties of the wood of some lesser utilized Ghanaian hardwoods, A Phd. Thesis Submitted to the Department of Wood Science and Technology, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana, p 150
Lemmens RHMJ (2008) Khaya ivoransis A. Chev. [internet] Record from PROTA4U. In: Louppe D, Oteng-Amoako AA, Brink M (eds) PROTA (Plant Resource of Tropical Africa), Wagenningen, Netherlands. http://www.prota4u.org/search.asp. Accessed 9 April 2014
Ministry of Lands and Natural Resources-MLNR (2012) Ghana-forest investment program (FIP) plan. Draft-March 2012: 5–66.
Ncube E (2010) Durability of Tropical Timbers against white Rot (Trametes versuolor) Fungi: Part 1. Durability classification of six Lesser-used Hardwoods; Copper belt University, School of Natural Resources, Kitwe-Zambia. 1–7. http://www.congress.sfb.bg.ac.rs/PDF/wood/rad23w.pdf. Accessed 30th Dec 2012
Okai R (1998) Recent advances in the machining of lesser-used species. In: Proceedings of ITTO/FORIG/TEDB-VAHPU Conference, City Hotel, Kumasi, Ghana, 17–19 February 1998, pp 94–105.
Okai R (2002) A study on the milling and strength properties of branchwood, Fellowship Report. ITTO Trop For Update 12(1):24–25. http://www.itto.int/direct/topics/topics_pdf_download/topics_id=1630000&no=1. Accessed 15 March 2012
Okai R (2003) Can logging residue be used to help meet timber demands for the downstream processing sector? In: Proceedings of the XII FAO World Forestry Congress, Quebec City, Canada. http://www.fao.org/docrep/ARTICLE/WFC/XII/0236-B4.HTM. Accessed 1st Feb 2013
Okoh ET (2014) Fibre, physical and mechanical properties of Ghanaian hardwoods. J Energy Nat Resour 3(3):25–30
Oteng-Amoako A, Ebanyenle E, Awuku F A (1998) The Identification of 14 under exploited promotable lesser used timber species of Ghana. In: Proceedings of ITTO/FORIG/TEDB-VAHPU Conference, City Hotel, Kumasi, Ghana. 17–19 February 1998, pp 46–51
Oteng-Amoako A A, Zurcher E, Agyakumhene I, Ebanyele E, Rogenmoser C (2008) 100 tropical African Timber trees from Ghana. Tree description and wood identification with notes on distribution, ecology, silviculture, ethnobotany and wood uses. Graphic Packaging, p 304
Panshin A J, Dezeeuw C (1980) Textbook of wood technology: In: Ncube E (2010) Durability of Tropical Timbers against white Rot (Trametes versuolor). Fungi: Part 1. Durability classification of six Lesser-used Hardwoods; Copper belt University, School of Natural Resources, Kitwe-Zambia, pp 1–7. http://www.congress.sfb.bg.ac.rs/PDF/wood/rad23w.pdf. Accessed 30 Dec 2012
Patel RN (1970) Anatomy of stem and root wood of Pinus radiata D. Don, New Zealand. J For Sci 1(1):37–40
Pleydell G (1994) Tropical timbers of Ghana. Timber Exports Development Board, Takoradi, Ghana, p 86
Quartey GA (2009) Relationships between some anatomical, physical and durability properties of the wood of some lesser utilised Ghanaian hardwoods, Thesis submitted to the Department of Wood Science and Technology at the Kwame Nkrumah University of Science and Technology, in partial fulfilment of the requirement for the degree of Doctor of Philosophy. p 150
Richter HG, Dallwitz (2000) Commercial timber; descriptions, illustrations, identification, and information retrieval. In England version 4th May, 2000. http://biodiversity.un.edu/delta. Accessed 15 May 2014
Samariha A, Kiaei M, Talaeipour M, Nemati M (2011) Anatomical structural differences between branch and Trunk in Ailanthus altissima wood. Indian J Sci Technol 4(12):1676–1678
Shrivastava MB (2000) Wood technology. Vikas publishing house PVT Ltd., Jangpura, p 180
Skadsen EN (2007) Basic wood anatomy and behaviour. In: Organic natural body jewellery (503) 240-0260, organic LLC. http://www.Organicjewellery.com/woodanatomy.html. Accessed 20 Dec 2012
Sreevani P, Rao R V (2014) Variation in basic density and anatomical properties of Eucalyptus Terticornis sm. Clones Res J Recent Sci 3(ISC-2013):271–274
Stokke DO, Manwiller FG (1994) Proportions of wood elements in stem, branch, and root wood of black oak (Quercus velutuia). IAWA J 15(3):301–310
Taylor FW (1973) Variations in the anatomical properties of South African grown Eucalyptus grandis, Appita. 27:171–178
Tchinda AT (2008) Entandrophragma angolense (Welw.) C.DC. In: Louppe D, Oteng-Amoako AA, Brink M (eds). PROTA (Plant Resource of Tropical Africa), Wagenningen, Netherlands. http://www.prota4u.org/search.asp. Accessed 15 Jan 2014
Tsoumis G (1991) Science and technology of wood; structure, properties, and utilization. Thessalomiki. Verlag Kessel, Greece, p 480
Walton JA (1974) Woodwork in theory and practice, 5th edn. The Australian Publishing Co. Pty Ltd., Sydney. Australia. pp 412–472
Wiedenhoeft A C, Miller R B (2005) Structure and functions of wood. In: Handbook of wood chemistry and wood composites. CRC press, New York: 2000 Boca, pp 10–33
Zhang SY, Zhong Y (1992) Structure-property relationship of wood in East Liaoning oak. Wood Sci Technol 26:139–149
Zobel BJ, Jett JB (1995) Genetics of wood production. Springer-Verlag, New York, p 337
Zobel BJ, Van Buijtenen JP (1989) Wood variation: its causes and control. Springer-Verlag, Berlin, p 363
Acknowledgements
We are grateful to the management and staff of Logs and Lumber Ltd. (LLL) especially Messrs; Cephas K. Kumah (HRM), Afful, Mher Snr. and Jnr., Sako, Manager Kofi (all of the forest section) and all the supervisors at the sawmill section. Also, Mr. Anthony Amamoo Eshun—TIDD’s Area Manager in Kumasi for his help. We are also indebted to Messrs, Michael Afful (now at Accra Poly.), Charles Cobbinnah, and Bright Agbeveade Y. Seenam (all of Kumasi Polytechnic) for their support. We also thank Prof. Sheldon Q. Shi and all staff of the Mechanical and Energy Engineering laboratory of the University of North Texas, Denton, USA, and staff of the Anatomy Laboratory of the Forest Research Institute of Ghana (FORIG)—Kumasi especially Anti Esi and Mr. Govina.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Dadzie, P.K., Amoah, M., Ebanyenle, E. et al. Characterization of density and selected anatomical features of stemwood and branchwood of E. cylindricum, E. angolense and K. ivorensis from natural forests in Ghana. Eur. J. Wood Prod. 76, 655–667 (2018). https://doi.org/10.1007/s00107-017-1195-6
Received:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00107-017-1195-6