
Abstract
Background
In the present study, the electropharmacological activity of traditional Chinese medicine, Ginkgo biloba extract (GBE), on human hyperpolarization-activated nucleotide-gated (HCN) channels and the underlying “funny” currents was investigated.
Methods
Standard two-electrode voltage-clamp recordings were employed to examine the properties of cloned HCN subunit currents expressed in Xenopus oocytes under controlled conditions and GBE administration.
Results
We found that GBE irreversibly inhibited the HCN2 and HCN4 channel currents in a concentration-dependent fashion and that the HCN4 current was more sensitive to GBE compared with HCN2. In addition, GBE inhibition of the current amplitudes of HCN2 and HCN4 currents was accompanied by a decrease in the activation and deactivation kinetics.
Conclusion
The results of this study contribute toward illustrating the antiarrhythmic mechanism of GBE, which might be useful for the treatment of arrhythmia.
Zusammenfassung
Hintergrund
In der vorliegenden Studie wurde die elektropharmakologische Aktivität einer Form traditioneller chinesischer Medizin, des Ginkgo-biloba-Extrakts (GBE), auf humane hyperpolarisationsaktivierte nukleotidgesteuerte (HCN-)Kanäle und die zugrunde liegenden Ströme, „funny currents“, untersucht.
Methoden
Standardisierte Aufnahmen mit 2‑Elektroden-Spannungsklemmen wurden verwendet, um die Eigenschaften der Ströme geklonter HCN-Untereinheiten zu untersuchen, die in Xenopus-Oozyten unter kontrollierten Bedingungen und Anwendung von GBE exprimiert wurden.
Ergebnisse
Die Autoren stellten fest, dass GBE die HCN2- und HCN4-Kanalströme konzentrationsabhängig irreversibel hemmte und dass der HCN4-Strom empfindlicher auf GBE reagierte als HCN2. Darüber hinaus trat als Begleiterscheinung der GBE-Inhibition bei den Stromamplituden der HCN2- und HCN4-Ströme eine Verminderung der Aktivierungs- und Deaktivierungskinetik auf.
Schlussfolgerung
Die Ergebnisse der vorliegenden Studie tragen zur Veranschaulichung der antiarrhythmischen Mechanismen des GBE bei, welcher sich möglicherweise für die Behandlung von Arrhythmien eignet.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
As is well known, the sino-atrial (SA) node is the pacemaker region of the mammalian heart. One of the most prominent features of SA node cells is the “funny” current (If; [1,2,3,4]). Activation of If at the termination of an action potential is the main process responsible for the generation of the diastolic depolarization and spontaneous activity of cardiac pacemaker cells. The inward If current that is carried by Na+ and K+ is activated upon membrane hyperpolarization within a voltage range [5, 6]. If is modulated by the direct binding of cyclic adenosine monophosphate (cAMP), which flows through the hyperpolarization-activated cyclic nucleotide-gated (HCN) channels. The molecular identity of this current has been identified through heterologous expression. The four isoforms of the mammalian HCN channels (HCN1–4), which belong to the superfamily of voltage-dependent K+ and cyclic nucleotide-gated channels, combine to form tetrameric channels in the heart and nervous system [7,8,9,10]. Previous studies have shown that all four channel isoforms have been detected in the heart. HCN4 is the most prominent HCN transcript in the SA node, whereas HCN2 is the dominant transcript in the ventricles of all the species investigated. If has been considered to produce automatic activity from other cardiac regions, including the Purkinje fibers, atrioventricular node, atrium, and ventricle [11,12,13,14].
Ginkgo biloba extract (GBE) has been used worldwide as a herbal medicine that possesses a number of constituents, and is mainly composed of flavonoids, including quercetin and rutin, and terpenoids such as bilobalide and ginkgolides A, B, and C [15, 16]. The effects of GBE and bilobalide (its main constituent) on the pacemaker activity and the underlying ionic currents in rat SA nodal cells have been investigated, indicating that GBE and bilobalide decreased the slope of the diastole (phase 4 depolarization; [17]). Satoh [18] demonstrated that the current of If was inhibited by this drug in rat SA nodal cells. Although these results indicated that GBE may affect the function of cardiac HCN channels and the spontaneous activity, the mechanism of its action has not been explained.
The aim of the present study was to examine whether GBE affects human HCN2 (hHCN2) and hHCN4-mediated currents heterologously expressed in Xenopus oocytes. The ability of GBE to modulate If in the oocytes injected with the HCN gene may contribute to anti-arrhythmic therapy.
Materials and methods
Study approval
The animals used in the study were treated in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and all the experimental methods were approved by the Animal Research Committee of the First Clinic College of Wuhan University (Wuhan, China).
In vitro transcription and functional expression in Xenopus oocytes
Wild-type hHCN2 and hHCN4 complementary DNA (cDNA) inserted into the pcDNA3 vector were kindly provided by Professor Ludwig and Professor Stieber (Friedrich-Alexander-Universität Erlangen-Nürnberg, Erlangen, Germany). cRNAs that were used for injection into oocytes were prepared with the T7 kit (Ambion, Austin, TX, USA) following linearization of the expression constructs with XbaI (Takara, Kyoto, Japan). RNA quality was checked by gel electrophoresis and its concentration was quantified by ultraviolet spectroscopy (UV-2201; Shimadzu Corp., Kyoto, Japan).
Voltage clamp of Xenopus oocytes
Xenopus frogs were anesthetized by cooling on crushed ice for 30–40 min. Ovarian lobes were digested with 1 mg/ml type IA collagenase in Ca2+-free ND96 solution for 30 min to remove follicle cells. Stage IV and V Xenopus oocytes were injected with 30 nl (1 µg/µl) of hHCN2 and hHCN4 cRNAs per oocyte using a Nanoject micro-dispenser and cultured in ND96 solution supplemented with 100 U/ml penicillin, 100 U/ml streptomycin, and 2.5 mM pyruvate at 17 ˚C for 2–3 days before being used in voltage clamp experiments. Recordings were performed 2–12 days after injection. A standard two-microelectrode voltage-clamp technique was used to record currents at 21–23 ˚C.
Drugs and reagents
Collagenase type I, zatebradine, CsCl, 4‑aminopyridine and HEPES were purchased from Sigma-Aldrich (St. Louis, MO, USA). Bovine serum albumin and pronase E were obtained from Roche (Basel, Switzerland). The GBE was provided by Wuhan Jianmin Pharmaceutics Co. (Wuhan, China), and was dissolved in dimethyl sulfoxide (DMSO) stock solution. The DMSO final concentration in the bathing solution was 0.5%. In order to maintain a constant concentration of ions and drugs, the perfusion rate was strictly controlled using the perfusion device BPS‑4 (ALA Scientific Instruments, Inc., Westbury, NY, USA) and a constant-flow pump.
Data acquisition and statistical analysis
All the data were stored on the computer hard disk and analyzed off-line using Clampfit 10.0 (Axon Instruments, San Jose, CA, USA) and Origin 8.0 software (Origin Laboratory, Northampton, MA, USA). The amplitude of HCN-mediated currents was defined as the time-dependent component (Istep) at the end of hyperpolarizing pulses or the peak tail current (Itail) at the beginning of depolarizing pulses. To construct I–V associations, the currents were normalized to their own maximum current measured prior to drug treatment and subsequently plotted as a function of the test potential (Vt). Voltage dependent on HCN current activation was determined by the analysis of Itail measured at depolarizing potentials. All the tail current amplitudes from an individual oocyte were normalized to their own Imax, plotted as a function of Vt, and fitted again with a Boltzmann function: I / Imax = 1 / [1 + exp(Vt − V1/2) / k] to determine the values of the half-point (V1/2) and the slope (k). The time constants for the HCN current activation or deactivation (activation or deactivation) at different Vt were determined using the standard exponential curve fitting. Activating or deactivating currents were fitted to a single exponential function: I(t) = Ae −t / τ + C. The concentration–effect curves were fitted using the Hill equation in the form, f = 1 / [1 + (IC50 / D)n], where f was the increase in HCN currents, expressed as percentage change compared with the control values, IC50 was the concentration of GBE for half-maximum inhibitory, D was the concentration of GBE, and n was the Hill coefficient. Data are presented as the mean ± standard deviation. Student’s t test was used for statistical analysis of the paired observations, and an analysis of variance was performed to test the difference among the groups; p<0.05 was considered to indicate a statistically significant difference.
Results
Electrophysiological properties of hHCN channels heterologously expressed in Xenopus oocytes.
For voltage-clamp recording, the hHCN2 current was elicited by hyperpolarization pulses of 4000 ms from a holding potential of −40 to −140 mV in 10-mV decrements at 0.1 Hz and subsequently clamped back to 0 mV for 1000 ms, whereas the hHCN4 current by hyperpolarization pulses of 4000 ms was from a holding potential of −60 mV to −150 mV in 10-mV decrements at 0.1 Hz and subsequently clamped back to 10 mV for 2000 ms (Fig. 1). Thereafter, selective and non-selective f‑channel blockers, zatebradine and CsCl, were utilized to confirm the HCN channel biophysical properties. The hHCN2 (n = 3) and hHCN4 (n = 4) currents were readily and completely blocked by 5 mM CsCl. By contrast, 10 µM zatebradine markedly inhibited the hHCN2 and hHCN4 currents by 72.3 ± 14.4 (n = 3) and 86.3 ± 9.7% (n = 4), respectively (Fig. 1).

Activation curves of the hHCN2 and hHCN4 currents and the inhibitory effects of zatebradine or CsCl on currents heterologously expressed in the Xenopus oocytes. a The representative current traces of hHCN2 and subsequent to administration of 10 µM zatebradine or 5 mM CsCl are indicated. b The representative current traces of hHCN4 and subsequent to administration of 10 µM zatebradine or 5 mM CsCl are indicated
Concentration-dependent blockage of hHCN currents by GBE
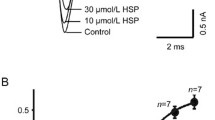
The results indicated the percentage inhibition of the hHCN2 and hHCN4 currents induced by GBE (0.01–1 mg/ml) at the test potentials of −110 mV investigated (Fig. 2a). The association between the decreased fraction of If and the concentrations of GBE at −120 mV is clearly shown in Fig. 2b, c, with IC50 values of 0.25 ± 0.01 and 0.12 ± 0.05 mg/ml, and Hill coefficients of 0.09 ± 0.03 (n = 8) and 0.06 ± 0.02 (n = 8), respectively. In addition, the time course of the effects of GBE on HCN currents was assessed. Blockages of currents occurred rapidly in the presence of 0.3 mg/ml GBE, but apparently were not reversed following washout for 5 min. After 15–20-min washout, the current failed to recover completely to the control level, but was recovered to ~55–75% of the control values (Fig. 2d).

Concentration-dependent blockage of Gingko biloba extract (GBE) on hHCN2 and hHCN4 currents in Xenopus oocytes. a The percentage inhibition of the hHCN2 and hHCN4 currents, induced by GBE (0.01–1 mg/ml), at the test potentials of −110 mV investigated. b Original current tracing of the If channel was superimposed prior to (control) and subsequent to superfusion of GBE (0.01–1 mg/ml). c The concentration–response curves were plotted based on data from b and fitted by the Hill equation, and IC50 values were calculated. d Time course of the effects of GBE on If. Pacemaker cells were perfused with Tyrode solution for 5 min before application of 0.3 mg/ml and again with Tyrode solution (washout). The normalized currents were plotted during the recording course
Effects of GBE on activation and deactivation kinetics of hHCN2 and hHCN4 channels
The effects of GBE on activation curves of hHCN2 and hHCN4 currents recorded in oocytes are shown in Fig. 3. The activation curves were plotted by amplitudes of tail currents at a depolarizing potential to 0 mV (Fig. 3a, b). The representative traces of hHCN2 and hHCN4 currents and the expanded traces of the outward tail current prior to (control) and subsequent to 0.03 or 0.3 mg/ml GBE treatment are illustrated in Fig. 3c, d. Superfusion of GBE (0.03 or 0.3 mg/ml) reduced normalized Itail at voltages in the range of −90–−140 mV, with more pronounced effects at the more hyperpolarized voltages (n = 8, p < 0.05). Furthermore, GBE (0.03 or 0.3 mg/ml) caused a shift of V1/2 from −93.6 ± 6.3 to −90.5 ± 4.6 mV (n = 8, p < 0.05) or −86.7 ± 5.6 mV (n = 8, p < 0.05), respectively. Similarly, GBE had inhibitory actions on hHCN4. The average value for V1/2 was −118.3 ± 7.2 mV under the control conditions, and −107.4 ± 4.9 mV and −102.1 ± 3.5 mV following the addition of 0.03 and 0.3 mg/ml GBE, respectively (n = 8, p > 0.05).

Effects of Gingko biloba extract (GBE) on voltage dependence of the activation of hHCN2 and hHCN4 currents. a Normalized activation curves of hHCN2 current expressed in the oocytes in ND96 solution and the presence of 0.03 or 0.3 mg/ml GBE. b Normalized activation curves of hHCN4 current expressed in the oocytes in ND96 solution and the presence of 0.03 or 0.3 mg/ml GBE. c Current traces of hHCN2 channel expressed in the Xenopus oocytes were superimposed prior to (control) and subsequent to superfusion of GBE (0.03 or 0.3 mg/ml). d Current traces hHCN4 channel expressed in the Xenopus oocytes were superimposed prior to (control) and subsequent to superfusion of GBE (0.03 or 0.3 mg/ml). Tail currents were normalized to the peak tail currents in the control condition for each oocyte and the data were fitted with the Boltzmann function
Effects of GBE on activation and deactivation kinetics of hHCN channels
Adding GBE from −140 mV to −90 mV in the testing potential channel increased τactivation significantly, and when the test potential became more negative, this change was more evident. Compared with the control, a lower (0.03 mg/ml) and a higher concentration (0.3 mg/ml) of GBE did not significantly affect the deactivation curves of the hHCN currents (n = 8, P > 0.05; Fig. 4a, b), which may indicate that the drug did not have a notable effect. However, the values of τactivation were increased markedly by GBE at a Vt of −110 mV from 504.6 ± 39.8 (n = 8) to 588.4 ± 21.7 ms (0.03 mg/ml, n = 8; p < 0.05), and 1176.4 ± 57.3 (0.3 mg/ml, n = 8; p < 0.05) and 1330.5 ± 59.8 ms (n = 8) to 1973.1 ± 83.6 (0.03 mg/ml, n = 8; p < 0.05) and 2814.5 ± 107.6 ms (0.3 mg/ml, n = 8; p < 0.05), respectively (Fig. 4c, d).

Effects of Gingko biloba extract (GBE) on the activation and deactivation kinetics of hHCN2 and hHCN4 channels in Xenopus oocytes. a The time constants for deactivation of hHCN2 currents and subsequent to GBE (0.03 and 0.3 mg/ml) were plotted by the holding potentials. b The time constants for deactivation of hHCN4 currents and subsequent to GBE (0.03 and 0.3 mg/ml) were plotted by the holding potentials. c The time constants for deactivation of hHCN2 currents and subsequent to GBE (0.03 and 0.3 mg/ml) were plotted by the holding potentials. d The time constants for deactivation of hHCN4 currents and subsequent to GBE (0.03 and 0.3 mg/ml) were plotted by the holding potentials. τactivation of the HCN currents from −90 mV to −140 mV and τdeactivation of HCN currents from −70 mV to −20 mV were determined using the standard exponential curve fitting
Discussion
The HCN family of ion channels represent the molecular substrate for If in cardiomyocytes. HCN2 was abundantly expressed in the ventricles and HCN4 in the SA node. The Xenopus oocytes were used as a heterologous expression system and here we discuss the effects of GBE on the expression of hHCN2 and hHCN4 individually. The hHCN2 and hHCN4 channels expressed in Xenopus oocytes showed that the electrophysiological properties were consistent with those reported previously [19,20,21]. The results of the present study indicated that blockage of GBE on the hHCN2 and hHCN4 channels is concentration-dependent, and that GBE markedly shifts the activation curve of hHCN2 and hHCN4 currents toward more negative potentials and slows the kinetics of activation, but not of deactivation. In addition, previous studies [17, 22, 23] have reported that GBE prolonged the action potential duration, inhibited the calcium channel currents, and delayed the outward rectifier potassium channel currents and inward rectifier potassium currents in guinea pig ventricular myocytes. In addition, Chen et al. [24] found that 0.01–0.1% GBE significantly inhibited the sodium current, L‑type calcium current, and transient outward potassium current in a concentration-dependent manner in rat ventricular myocytes. In hippocampal neuronal cells, GBE and its constituents inhibited receptor-gated chloride channels [25]. Cermak et al. [26] showed that quercetin (one of the constituents of GBE) activated basolateral potassium channels in the colon epithelium. These noteworthy findings indicated that GBE may be a potential multi-ion channel blocker, which could be widely applied in antiarrhythmic therapy.
Individual HCN subunits have six transmembrane segments (S1–S6), with a highly positively charged S4 domain that is the putative voltage sensor and the P domain between S5 and S6 acting as the ion-conducting pore and selectivity filter [27,28,29,30]. Wahl-Schott and Biel [29] believed that HCN channels were dually activated by voltage and by cyclic nucleotides, so that each one of the four subunits of the tetrameric channel was independently gated by voltage. When a voltage sensor switched to the activated state, the probability for channel opening increased. The present results demonstrated that GBE principally affected the activation of the two channels, which would decrease the probability of channel opening. This may be one reason why GBE inhibited the HCN channel current.
Furthermore, it was revealed that the response of GBE on hHCN2 and hHCN4 was almost irreversible, and it was predicted that GBE may be a drug with long-lasting action, which may exhibit distinct characteristics by blood perfusion. Notably, the results showed distinct effects of GBE (0.03 and 0.3 mg/ml) in a single concentration and, therefore, we had reason to believe that the final concentration of GBE active in the myocardium in the clinical study by Satoh [23] may be lower than the concentration used in the present study. This would also explain why cardiac arrhythmias are not observed during GBE therapy.
A previous study regarding the cardiac-specific and inducible knockout model of HCN4 and mutations in HCN4 channels indicated that the HCN4 current provided a fundamental contribution to basal heart rate maintenance and modulation since its removal leads to basal bradycardia and a strongly reduced response to sympathetic stimulation [31]. Blockade of If has the potential to reduce the heart rate without reducing contractility. This is particularly important in patients with heart failure, ischemia, or both. If is up-regulated in atrial and ventricular myocytes during heart failure, hypertrophy, and atrial fibrillation in rats and humans [4, 32,33,34,35]. Blocking these channels may be useful in reducing the heart rate without causing undesirable systemic cardiovascular reactions in patients with congestive heart failure and in patients suffering from conditions such as inappropriate sinus tachycardia. Therefore, controlling the heart via If blockers or down-regulation of If expression may be used in numerous clinical conditions.
Limitations
However, the data in the present study were obtained from a heterologous expression system, which could not be directly associated with arrhythmia genesis. In addition, despite the fact that GBE affected the function of hHCN channels, further experiments are required to elucidate each component acting on the HCN channels and interacting with each other.
Conclusion
In the present study, we demonstrated that GBE irreversibly inhibited the HCN2 and HCN4 channel currents in a concentration-dependent fashion and the HCN4 current was more sensitive to GBE than HCN2. In addition, GBE inhibition of the current amplitudes of HCN2 and HCN4 currents were accompanied by decreasing in the activation and deactivation kinetics. Therefore, these results may contribute to illustrate the antiarrhythmic mechanism of GBE, and might be useful for the treatment of arrhythmia.
References
DiFrancesco D (1993) Pacemaker mechanisms in cardiac tissue. Annu Rev Physiol 55:455–472
Ravagli E, Bucchi A, Bartolucci C, Paina M, Baruscotti M, DiFrancesco D (2016) Cell-specific dynamic clamp analysis of the role of funny If current in cardiac pacemaking. Prog Biophys Mol Biol 120:50–66
Robinson RB, Siegelbaum SA (2003) Hyperpolarization-activated cation currents: from molecules to physiological function. Annu Rev Physiol 65:453–480
Baruscotti M, Bucchi A, Difrancesco D (2005) Physiology and pharmacology of the cardiac pacemaker (‘funny’) current. Pharmacol Ther 107:59–79
Frace AM, Maruoka F, Noma A (1992) External K+ increases Na+ conductance of the hyperpolarization-activated current in rabbit cardiac pacemaker cells. Pflugers Arch 421:97–99
Bucchi A, Baruscotti M, DiFrancesco D (2002) Current-dependent block of rabbit sino-atrial node I(f) channels by ivabradine. J Gen Physiol 120:1–13
Ludwig A, Zong X, Jeglitsch M, Hofmann F, Biel M (1998) A family of hyperpolarization-activated mammalian cation channels. Nature 393:587–591
Ishii TM, Takano M, Xie LH, Noma A, Ohmori H (1999) Molecular characterization of the hyperpolarization-activated cation channel in rabbit heart sinoatrial node. J Biol Chem 274:12835–12839
Altomare C, Terragni B, Brioschi C, Milanesi R, Pagliuca C, Viscomi C, Moroni A, Baruscotti M, DiFrancesco D (2003) Heteromeric HCN1-HCN4 channels: a comparison with native pacemaker channels from the rabbit sinoatrial node. J Physiol 549:347–359
Er F, Larbig R, Ludwig A, Biel M, Hofmann F, Beuckelmann DJ, Hoppe UC (2003) Dominant-negative suppression of HCN channels markedly reduces the native pacemaker current I(f) and undermines spontaneous beating of neonatal cardiomyocytes. Circulation 107:485–489
Hoppe UC, Beuckelmann DJ (1998) Characterization of the hyperpolarization-activated inward current in isolated human atrial myocytes. Cardiovasc Res 38:788–801
Shi W, Wymore R, Yu H, Wu J, Wymore RT, Pan Z, Robinson RB, Dixon JE, McKinnon D, Cohen IS (1999) Distribution and prevalence of hyperpolarization-activated cation channel (HCN) mRNA expression in cardiac tissues. Circ Res 85:e1–e6
Moosmang S, Stieber J, Zong X, Biel M, Hofmann F, Ludwig A (2001) Cellular expression and functional characterization of four hyperpolarization-activated pacemaker channels in cardiac and neuronal tissues. Eur J Biochem 268:1646–1652
Fernández-Velasco M, Goren N, Benito G, Blanco-Rivero J, Boscá L, Delgado C (2003) Regional distribution of hyperpolarization-activated current (If) and hyperpolarization-activated cyclic nucleotide-gated channel mRNA expression in ventricular cells from control and hypertrophied rat hearts. J Physiol 553:395–405
Huh H, Staba EJ (1992) The botany and chemistry of ginkgo biloba L. J Herbs Spices Med Plants 1:92–124
Huang P, Zhang L, Chai C, Qian XC, Li W, Li JS, Di LQ, Cai BC (2014) Effects of food and gender on the pharmacokinetics of ginkgolides a, b, c and bilobalide in rats after oral dosing with ginkgo terpene lactones extract. J Pharm Biomed Anal 100:138–144
Satoh H, Nishida S (2004) Electropharmacological actions of ginkgo biloba extract on vascular smooth and heart muscles. Clin Chim Acta 342:13–22
Satoh H (2005) Suppression of pacemaker activity by ginkgo biloba extract and its main constituent, bilobalide in rat sino-atrial nodal cells. Life Sci 78:67–73
Azene EM, Xue T, Marbán E, Tomaselli GF, Li RA (2005) Non-equilibrium behavior of HCN channels: insights into the role of HCN channels in native and engineered pacemakers. Cardiovasc Res 67:263–273
Bucchi A, Barbuti A, Baruscotti M, DiFrancesco D (2007) Heart rate reduction via selective ‘funny’ channel blockers. Curr Opin Pharmacol 7:208–213
Tamura A, Ogura T, Uemura H, Reien Y, Kishimoto T, Nagai T, Komuro I, Miyazaki M, Nakaya H (2009) Effects of antiarrhythmic drugs on the hyperpolarization-activated cyclic nucleotide-gated channel current. J Pharmacol Sci 110:150–159
Nishida S, Satoh H (2003) Mechanisms for the vasodilations induced by ginkgo biloba extract and its main constituent, bilobalide, in rat aorta. Life Sci 72:2659–2667
Satoh H (2003) Effects of ginkgo biloba extract and bilobalide, a main constituent, on the ionic currents in guinea pig ventricular cardiomyocytes. Arzneimittelforschung 53:407–413
Chen B, Cai J, Song LS, Wang X, Chen Z (2005) Effects of ginkgo biloba extract on cation currents in rat ventricular myocytes. Life Sci 76:1111–1121
Chatterjee SS, Kondratskaya EL, Krishtal OA (2003) Structure-activity studies with ginkgo biloba extract constituents as receptor-gated chloride channel blockers and modulators. Pharmacopsychiatry 36(1):S68–S77
Cermak R, Kuhn G, Wolffram S (2002) The flavonol quercetin activates basolateral K(+) channels in rat distal colon epithelium. Br J Pharmacol 135:1183–1190
Santoro B, Tibbs GR (1999) The HCN gene family: molecular basis of the hyperpolarization-activated pacemaker channels. Ann N Y Acad Sci 868:741–764
Biel M, Schneider A, Wahl C (2002) Cardiac HCN channels: structure, function, and modulation. Trends Cardiovasc Med 12:206–212
Wahl-Schott C, Biel M (2009) HCN channels: structure, cellular regulation and physiological function. Cell Mol Life Sci 66:470–494
Baruscotti M, Barbuti A, Bucchi A (2010) The cardiac pacemaker current. J Mol Cell Cardiol 48:55–64
Baruscotti M, Bucchi A, Viscomi C, Mandelli G, Consalez G, Gnecchi-Rusconi T, Montano N, Casali KR, Micheloni S, Barbuti A, DiFrancesco D (2011) Deep bradycardia and heart block caused by inducible cardiac-specific knockout of the pacemaker channel gene Hcn4. Proc Natl Acad Sci USA 108:1705–1710
Zicha S, Fernández-Velasco M, Lonardo G, L’Heureux N, Nattel S (2005) Sinus node dysfunction and hyperpolarization-activated (HCN) channel subunit remodeling in a canine heart failure model. Cardiovasc Res 66:472–481
Mangoni ME, Nargeot J (2008) Genesis and regulation of the heart automaticity. Physiol Rev 88:919–982
Stillitano F, Lonardo G, Zicha S, Varro A, Cerbai E, Mugelli A, Nattel S (2008) Molecular basis of funny current (If) in normal and failing human heart. J Mol Cell Cardiol 45:289–299
Wei-qing H, Qing-nuan K, Lin X, Cheng-hao G, Qi-yi Z (2011) Expression of hyperpolarization-activated cyclic nucleotide-gated cation channel (HCN4) is increased in hypertrophic cardiomyopathy. Cardiovasc Pathol 20:110–113
Funding
This work was supported by the National Science Foundation for Distinguished Young Scholars of China under Grant No. 81500255, and Hubei Provincial Natural Science Foundation of China under Grant No. 2018CFB420.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
H. Chen, Y. Chen, J. Yang, P. Wu, X. Wang and C. Huang declare that they have no competing interests.
For this article no studies with human participants were performed by any of the authors. All studies performed were in accordance with the ethical standards indicated in each case. The animals used in the study were treated in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and all the experimental methods were approved by the Animal Research Committee of the First Clinic College of Wuhan University (Wuhan, China).
Rights and permissions
About this article

Cite this article
Chen, H., Chen, Y., Yang, J. et al. Effect of Ginkgo biloba extract on pacemaker channels encoded by HCN gene. Herz 46, 255–261 (2021). https://doi.org/10.1007/s00059-020-04933-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00059-020-04933-z