
Abstract
Termites, and in particular species that remove live plant material, play an important role in rangeland deterioration in the arid areas of the world. In the arid parts of southern Africa, where the sparse vegetation is important for subsistence and large-scale game farming, it is vital to understand termite ecology and feeding behaviour. Here, we focus on the termite species, Baucaliotermes hainesi, the most common termite species within its range in the arid interior of southern Africa. We determine B. hainesi foraging and nest maintenance activities, predator response and the preferred food plants. The study was conducted during 2014 in the Nama-Karoo biome of South Africa. We found that Baucaliotermes hainesi is strictly a nocturnal species and that temperature is the most important abiotic determinant of foraging activity. Below 14 °C, no foraging takes place, whilst maximum foraging activity is observed only above 20 °C. Only three plant species are foraged upon, with the grass Stipagrostis uniplumis utilised the most. We suggest that B. hainesi can significantly reduce grass cover, particularly in overgrazed areas and in dry years. Further studies to elucidate the effects of drought, overgrazing and the role of B. hainesi in veld degradation are critical, in particular under future climate scenarios that the predict aridification of B. hainesi habitat.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Termites are globally recognised as one of the most important soil engineers and are crucial to ecosystem functioning (Harris 1971; Jouquet et al. 2006). Since termites construct large extensive epigeal nests or large underground systems, as ecosystem engineers, they improve soil hydrology and composition, and are important as decomposers and in nutrient cycling, particularly in semi-arid regions (see, for example, Whitford et al. 1992; Jones et al. 1994; Dangerfield et al. 1998). In contrast, many species of termite are considered pests, and are responsible for huge economic losses (Harris 1971; Verma et al. 2009). Other than damage to wooden structures and agricultural crops, termites are also responsible for the deterioration of rangelands (Nel 1968; Van Ark 1969; Bodin et al. 2006).
From an ecological viewpoint, termites are an important basal link in the food chain and are an important food source for many species, including ants (e.g., Mirenda et al. 1980), reptiles (e.g., Mouton et al. 2000), amphibians (Harris 1971), birds (such as the crowned plover, Vanellus coronatus, with a diet consisting of 30 % termites Urban et al. 1986), and mammals (e.g., Taylor et al. 2002). One mammal, the aardwolf (Proteles capensis) in Africa has become so specialized that it almost solely feeds on termites (Richardson 1987; Koehler and Richardson 1990).
Africa is a particularly species rich continent for termites, with an estimated thousand species, out of the 2600 described species (Kambhampati and Eggleton 2000; Uys 2002). In general, termite diversity, abundance and biomass decrease with an increase in latitude (Eggleton 2000). However, southern Africa with 54 genera and 165 termite species (Uys 2002) is a region unusually rich at such a southern latitude (Eggleton 2000). Surprisingly, for the drier areas of southern Africa, only two species, Hodotermes mossambicus and Trinervitermes trinervoides, have received substantial attention, H. mossambicus as an important pest species (Coaton 1958; Nel 1968; Nel and Hewitt 1969; Van Ark 1969; Basson 1972; Duncan and Hewitt 1989) and T. trinervoides initially because of its role in the deterioration of degraded rangelands (Coaton 1948a, b, 1953; Hartwig 1955), but subsequently as important food source for the specialized myrmecophagous aardvark and aardwolf (Richardson 1987; Taylor et al. 2002; Anderson 2004). Despite the importance of termites globally and in particular in the grasslands of southern Africa, other termite species in southern Africa have received far less attention. One such species, whose ecology is largely unknown, is Baucaliotermes hainesi.
Baucaliotermes hainesi is potentially a keystone species, since it is the most abundant termite species in most of its distribution range (Coaton and Sheasby 1973), it influences plant community composition, builds massive nests (Tschinkel 2010), acts as soil engineer and provides an important food source for the myrmecophagous aardvark (pers. obs). Furthermore, similar to the closely related T. trinervoides which is the most common termite species in South Africa (Sands 1965), B. hainesi might play a role in the further deterioration of already degraded rangeland in the arid areas of southern Africa (Dougill et al. 2016). Under future climate change scenarios, these arid areas are predicted to become drier which potentially will exacerbate termite impacts (Midgley et al. 2001).
If the distribution of B. hainesi is superimposed on the distribution of the closely related Trinervitermes species, it is clear that B. hainesi occupies the ecological niche in the most arid parts of southern Africa occupied by Trinervitermes species in the wetter areas. Therefore, it is surprising that B. hainesi has not been well studied. Other than a species description, distribution data (Coaton and Sheasby 1973) and a more recent and detailed study on nest structure (Tschinkel 2010), there are, to our knowledge, no studies on B. hainesi. In this study, we explore its ecology and influence on land degradation by determining (1) foraging behaviour and preferred food plants, (2) influence of climate on foraging activity, (3) nest maintenance activities, and (4) predator response.
Methods
Study area
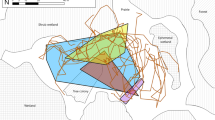
The study area is located in the Northern Cape Province of South Africa on a game farm, Tierkop (Fig. 1). This is a 7400 hectare game farm which stocks low densities of Hartmann’s mountain zebra, red hartebeest, eland, springbok, blue wildebeest and gemsbok. Vegetation consists of low karroid shrubland (Nama-Karoo biome) on flat gravel and sandy plains. This vegetation is seen as a transition vegetation between Kalahari proper (Savanna Biome) and the northern Nama-Karoo (Mucina and Rutherford 2006). Approximately 60 % of the Nama-Karoo has moderately to severely degraded soils and vegetation (Hoffman and Ashwell 2001). At our study sites, Stipagrostis ciliata, S. obtusa and S. uniplumis var. uniplumis were the dominant grasses (see Table S3 for a complete list of plant species occurring at the study sites).

Study plots and the main study area indicated in relation to rivers and roads. Inset map indicates locality in South Africa, with the hatched polygon indicating the known Baucaliotermes hainesi distribution (redrawn from Coaton and Sheasby 1973)
The mean annual precipitation in the area is between 100 and 200 mm, with most rainfall in late summer and early autumn. In southern Africa, only the Desert biome has a higher variability in annual rainfall. As a consequence, droughts are unpredictable and can be prolonged, and rivers are non-perennial. Solar radiation is high. Mean maximum temperatures are as high as 39.5 °C in January with mean minimum temperature in midwinter (July) as low as −4.2 °C.
Study species
The harvester termite, Baucaliotermes hainesi (Fuller, 1922), is an endemic in southern Namibia and the northwestern parts of South Africa (Coaton and Sheasby 1973). Baucaliotermes hainesi belongs to the Nasutitermitinae and a subfamily of the Termitidae (Krishna et al. 2013). The composition and the role in defence of the frontal gland secretions of the Nasutitermitinae are well studied (reviewed in Šobotník et al. 2010). Baucaliotermes hainesi workers and soldiers reach their foraging areas by way of a network of subterranean galleries (see Tschinkel 2010 for details on nest structure). This complex tunnel system radiates at least 10–15 m from the nest, with connections to the surface at 50–75 cm intervals (Tschinkel 2010).
Surface access point surveys, foraging and maintenance activities
Sampling was done in autumn (March 2014), due to high B. hainesi activity this time of year (pers. obs.; Adam et al. 2008). Five plots of 20 m long by 10 m wide were used (for a total area of 1000 m2). Plots were permanently marked with small poles with different colour coded reflectors. Plots were monitored by trained observers for two subsequent nights. Monitoring started at 18h00 (sunset at 19h01) and repeated every hour, on the hour, throughout the night until the last surface access point closed.
In contrast to other studies, we use the term “surface access points” instead of “foraging holes”, since holes are not only used for foraging but also for maintenance activities. Termites counter erosion and nest damage with a continuous transport of soil to the surface (Turner et al. 2006). These quantities of soil moved by termites can be substantial (Pomeroy 1976). Fresh surface deposition consists of a roughly textured soil of a darker colour. We define these activities as nest maintenance activity.
Recording and observation of active surface access points is a common method of assessing the foraging activity of harvester termites (Nel and Hewitt 1969; Basson 1972; Ohiagu and Wood 1976; Ohiagu 1979a; Adam et al. 2008). Every hour all new surface access points were marked with a colourful flag. The time at which a surface access point become active or inactive was recorded. The number and length of foraging columns going out from a service access point (n = 325 surface access points and 502 foraging columns counted over the study period) were determined. The number of workers returning with a piece of grass was counted for five 1-min intervals (with 5 min waiting time in-between) for 20 randomly selected surface access points.
After peak foraging time was determined on the first two nights of detailed sampling, plots were only monitored during peak activity times for three subsequent nights to determine usage constancy of surface access points. In addition, during three consecutive nights, 100 × 10 m (for a total of 1000 m2) plots were monitored at 13 haphazardly selected sites during peak activity periods, i.e., early evening (Table S1). With multiple observers, more than one plot was sometimes monitored simultaneously. A 100 m rope was laid out and observers walked with a 1 m stick on both sides of the line, noting active surface access points. In total 285 h of observations were done over the five nights.
Influence of climatic conditions on foraging and maintenance
Temperature is known to be one of the main variables influencing foraging of termites (Kofoid 1934). To determine the importance of temperature, humidity and wind on B. hainesi foraging and maintenance activities, during all surveys, these climate variables were measured at ground level with an Acurite Pro Weather Station. The significance of these variables was assessed in two separate general regression models (GRM), with either foraging or maintenance as the response variable and temperature, humidity and wind as predictor variables. The assumptions of the GRM were investigated using probability plots, whose residuals were normally distributed. All analyses were conducted in Statistica 12 (StatSoft, Inc. 2013, Tulsa, USA).
Baucaliotermes hainesi response to predation
Ant predation behaviour was observed throughout the 285 h of observations. Ants were identified with http://antsofafrica.org and http://www.antwiki.org and confirmed with a verified voucher from the Iimbovane collection at Stellenbosch University.
Since B. hainesi are potentially preyed upon by various predators, the reaction to disturbance of B. hainesi was explored. Fifteen foraging parties were removed entirely with a soft brush and dustpan and surface access points observed subsequently for 10 min. To test the termite response of damage to surface access points, a small brush was used to sweep over surface access points and termite response observed for 10 min.
Baucaliotermes hainesi densities and plant species utilisation
To determine B. hainesi aboveground ratio of soldiers to workers, termites in a maintenance and foraging patch were collected with a dustpan and broom (n = 10 maintenance and eight foraging patches) and counted.
To determine the favoured food source of B. hainesi, foraging workers were observed for 40 h over four nights (5 h per night for two observers). All plant species foraged upon were identified with relevant field guides (Le Roux 2005; Derichs and du Plessis 2012).
Results
Surface access points
Prior to emerging, workers remove the plug of soil from the round (5 mm in diameter, minimum 3 mm; maximum 7 mm; n = 10) surface access points. Soldiers appear first and spread out, facing outward (Fig. 2a). Surface access points are rarely used on consecutive nights, with 93 % only used once during five consecutive nights. After emergence, activity can either be foraging or maintenance. Only workers partake in foraging and maintenance, with soldiers providing protection. Workers (3.56 mg ± 0.80; mean ± SD; n = 11) are significantly heavier than soldiers (1.07 mg ± 0.33; mean ± SD; n = 10) within the same colony (Mann–Whitney U test U = 0.0, Z = 3.8, P < 0.01).

a Baucaliotermes hainesi soldiers guarding a surface access point. b B. hainesi forages only at night, and when a light is shown on the surface access point, the urge to get away from the light source is stronger than to return underground. c Foraging column splitting up towards two different tufts of grass. d Area of past high Baucaliotermes hainesi activity. This is indicated by the nests that slightly protrude above the soil surface following the removal of topsoil by sheet erosion. e Foraging B. hainesi worker transporting a piece of grass is guarded by a number of soldiers. Note the size difference. f Multiple queens from a single nest
Surface access point activity can switch between maintenance and foraging. Twenty-five access points started as maintenance and switched to foraging, while for 59, it was vice versa. The remainder had the same activity throughout (n = 319 over the two nights of detailed sampling). The shortest maintenance bout of 32 min was followed by 40 min of foraging, whilst the longest foraging bout of 433 min was followed by 322 min of maintenance (18h47–07h22). It is only during peak foraging time that the number of foraging surface access points surpasses that of maintenance (Fig. S1).
Baucaliotermes hainesi is very sensitive to light, and no activity was observed before sunset or after sunrise. A torch 1.5 m above an established foraging column is tolerated, but when brought closer, the foraging column dammed up in front of the light. Foraging is then terminated and termites return underground (Fig. 2b). A light above a newly opened surface access point terminates all activity and the surface access point remains unused for the remainder of the night.
Maintenance activities
Nest maintenance involves a column of workers, each emerging with a grain of sand in their mandibles, which they deposit just outside the surface access point. After a worker disposed the grain of sand, it will return to the same surface access point. The amount of sand increases up to a height of 33 mm (range 10–33 mm). Maintenance activities lasted on average 2 h (range 27–576 min) with an average B. hainesi maintenance patch diameter of 6 cm (range 1–14 cm).
Foraging activities
Initially, foraging involves a single column one to two workers wide, which develops into a column of two to four workers wide, or rarely up to seven workers wide. Foraging activity lasted on average 142 min (range: 20–543 min). Foraging starts just before sunset (earliest started at 18h37, with the latest only commencing at 1h12), and even though foraging continues throughout the night, peak foraging time is between 20h00 and 23h00.
Foraging columns can increase in length to a maximum of 1.06 m (median 0.25 m). Along the length of these foraging columns, soldiers move up and down and take position at higher elevations on stones and grass tussocks (Video 1).
At the food source, workers fan out to collect plant material. This is done by climbing into a grass tussock or shrub and cutting grass blades or leaves with their mandibles. Up to four foraging columns can emerge from a single surface access point, each foraging at a separate grass tuft (Fig. 2c). Cut grass is also collected underneath the grass tussock. Large pieces are cut into smaller parts before being carried back to the nest (median length ± 95 % CI 5.8 ± 0.2 mm; Fig. S2, Fig. 2e). Eighty-three percent of workers return without a piece of grass (n = 3910 workers observed).
Influence of climatic conditions on foraging and maintenance
Baucaliotermes hainesi activity peaks in early evening with a significant decline until activity ceases. This is strongly influenced by temperature, with higher temperature favouring foraging activity (Table 1; Fig. 3). Temperature is negatively correlated to humidity (Table S4). In contrast, maintenance activities are not influenced by the climatic variables measured (Table S2).

Total number of foraging columns recorded at various temperatures over 5 nights of observation at 13 plots randomly distributed over 7400 hectares. Under higher temperatures, there is more Baucaliotermes hainesi foraging activity (see Table 1)
Response to predation
When ants, of the species Tetramorium grandinode (subfamily Myrmicinae), were close to surface access points, only a few soldiers would emerge, and no foraging or maintenance activities would take place. Tetramorium grandinode actively catch workers when they have the opportunity. Strangely, surface access points were not closed and could remain open for hours, even after all ants had left. Most surface access points were then unused for the entire night, whilst in a few cases, soldiers would appear and maintenance activities would begin.
Ants generally waited at a distance until a foraging column had formed before attacking. Attacks consisted either of an ant rushing in from a distance, grabbing an isolated termite with its mandibles (Video 2) or a more subtle approach, where an ant would use cover to get close to the foraging column before striking.
When foraging columns were removed by hand, there was no reaction within the 10 min observation period. Similarly, when service access points were damaged, there was no immediate reaction, and service access points could remain open for hours.
Baucaliotermes hainesi densities and plant species utilisation
Maintenance patches were dominated by soldiers with an average 145 soldiers and only 24 workers (n = 10). Foraging columns also contained more soldiers with 154 soldiers and 91 workers (n = 8 single foraging columns of 15–48 cm in length).
During a single night, there are 2180 maintenance and 1940 foraging B. hainesi surface access points per hectare (average over the two nights of detailed sampling on 0.1 hectare). On average, there are 843720 B. hainesi individuals on the soil surface per hectare per night. Due to extensive tunnelling, the number of nests per hectare could not be determined, but interestingly more than one queen can be found in a nest (Fig. 2f).
The grass species Stipagrostis uniplumis is the preferred food source for B. hainesi. Another abundant species, S. obtusa, is also utilised, whilst S. ciliata was never utilised, as were six other less abundant grass species (see Table S3 for full plant species list of the study area). In areas with limited Stipagrostis, the herbaceous Zygophyllum decumbens was utilised, but only leaves and young twigs.
Discussion
The foraging behaviour of Baucaliotermes hainesi is very similar to that reported for other Nasutitermitinae species in Africa, such as Trinervitermes trinervoides and T. geminatus (Sands 1961; Ohiagu and Wood 1976; Ohiagu 1979a; Adam et al. 2008). Foraging tunnels of B. hainesi are very similar to those of other species, such as Macrotermes bellicosus (Pomeroy 1976), M. michaelseni (Darlington 1982) and T. trinervoides, in that foraging tunnels can extend between 30 and 50 m from the mound (Tschinkel 2010). These underground structures ensure that workers rarely have to forage far from safety. In fact, B. hainesi usually forages only half a metre and never more than a metre from a surface access point. This is much closer than T. geminatus, which can forage up to 3 m (Ohiagu and Wood 1976), and also closer than T. trinervoides that can forage up to 1.4 m (Adam et al. 2008) from a surface access point. An interesting observation is that B. hainesi forage columns do not merge, in one instance, foraging from two surface access points only 2 cm apart, moved in different directions. In contrast, in T. trinervoides, up to six surface access points as far as 40 cm apart frequently merge into a single foraging column (Adam et al. 2008).
While Uys (2002) and Grohmann (2010) found that B. hainesi foraged on leaf litter, decaying wood and herbivore dung, we never encountered B. hainesi utilising these food items. Similar to T. geminatus, B. hainesi cuts down and forages on life plant material (Ohiagu and Wood 1976). In contrast, T. trinervoides shows a preference for grass litter (Adam et al. 2005). An unexpected large number of workers (83 %) returned to the nest without any food. This was also noted in T. geminatus (Ohiagu and Wood 1976) and T. trinervoides (Adam et al. 2008) although at much lower incidence (1–30 %). The previous studies were unable to determine the function of this, but we speculate that this is to regurgitate food to soldiers, nymphs and larvae (McMahan 1963), although this was not observed in this study. We speculate that the higher percentage of workers returning without grass [compared to the closely related T. trinervoides (Adam et al. 2008)] might be related to the higher soldier to worker ratio in B. hainesi. With a higher soldier to worker ratio, more soldiers must be fed by regurgitating workers, which will result in fewer workers returning with a piece of plant material.
The worker-to-soldier ratio of 1:1.7 is not unusual in the Nasutitermitinae (Haverty 1977) but is much higher than in T. trinervoides, where soldiers comprise 40 % (in contrast to 63 % in B. hainesi) of the number of termites on the surface (Adam et al. 2008). However, the lower percentage of T. trinervoides soldiers might not represent the composition of the entire colony, since the ratio of soldiers to workers can be much lower in the mound itself. Ratios of workers to soldiers varied from 1:1 in the smallest colonies to 5:1 in larger nests (Adam et al. 2012). The higher percentage of soldiers might be related to B. hainesi workers being half the size of T. trinervoides and only a third of the co-occurring Hodotermes mossambicus (worker weights of B. hainesi 3.5 mg, T. trinervoides 6.4 mg and H. mossambicus 9.5 mg; this study; Nel 1968). As would be expected, soldiers are extremely protective of the queen, but interestingly, we found more than one queen in one of the excavated nests (Fig. 2f). Whilst most species of termites are monogynous, i.e., colonies contain only one queen, polygynic colonies are not unusual in the Termitidae (Thorne 1982).
When potential threats are detected, the soldiers point their nasus in the direction of the threat. Sticky, noxious turpentine threads are squirted from the nasus at potential predators (Video 3). The chemical defence of Baucaliotermes hainesi is so successful in deterring predators that known termite predators, such as Schinz’s beaked blind snakes, Rhinotyphlops schinzi (Typhlopidae), stay clear of foraging columns (pers. obs.).
In this study, the ant, Tetramorium grandinode, preyed upon B. hainesi. If ants are present when B. hainesi opens a surface access point, there is no further termite activity, even though the surface access point remains open for a few hours. Ants were immobilised by the soldiers when they were detected, but surface access points remained open in the presence of ants. In contrast, when the co-occurring H. mossambicus is attacked by ants (Anoplolepis custodiens), it retreats underground and plugs the surface access points (Coaton 1958). In T. geminatus, predators, such as lizards, evoke this response, but ants do not lead to forage access point closure (Ohiagu and Wood 1976). A response to a threat, such as a predator or bright light, also elicits vibration behaviour by B. hainesi soldiers. This response has been well studied in Macrotermes natalensis and serves as a warning signal to other soldiers and workers with signals transmitted along the subterranean galleries (Hager and Kirchner 2013, 2014).
Foraging activities of African termite species have been well studied (e.g., Sands 1961; Ohiagu 1979b; Heidecker and Leuthold 1984; Duncan and Hewitt 1989; Olugbemi and Malaka 2007; Adam et al. 2008; Olugbemi 2010), but this study is the first to quantify nest maintenance activity in an African termite. Maintenance can happen from any surface access point and can switch to foraging and vice versa at any time. Maintenance is less influenced by climatic conditions than foraging. Part of the reason might be that during foraging more workers are above ground for longer periods and these workers are further away from the surface access point.
Temperature is the most important climatic factor in determining B. hainesi activity. Baucaliotermes hainesi avoids colder temperatures and only starts foraging at 14 °C, with peak foraging when temperatures reach the mid-1920 (Fig. 3). This is in contrast to T. trinervoides that becomes active at less than 10 °C and for which optimum foraging temperature is much lower at 10–14 °C (Adam et al. 2008). In contrast, the central African T. geminatus prefers much higher foraging temperatures of between 20 and 35 °C (Ohiagu 1979b). The co-occurring H. mossambicus also forages during the day, so when night time temperatures are too low, particular in winter, they can utilise the warmer days (Coaton 1958). Although T. trinervoides is mainly nocturnal, it sometimes forages after sunrise and in the afternoon (Adam et al. 2008), but this was never observed in B. hainesi. This difference is probably because T. trinervoides can forage in the shade of the litter which provides predator and light protection, whilst B. hainesi forages in grass tufts and is then very exposed. With very low temperature in winter (night temperature below 10 °C) at the study site, anecdotal observations for 2 weeks in winter confirm that the low winter temperatures prevent foraging. Autumn foraging is thus important, as has been shown for other Nasutitermitinae, with nest grass reserves depleted after the winter (see, for example, Adam et al. 2008). As in other studies, we noticed that precipitation ends all aboveground activity in B. hainesi (Coaton 1958). Due to the arid conditions, only a few raindrops were observed during the study period, but this was enough for all workers to hurriedly withdraw underground and seal up surface access points.
The lengths of grass (5 mm long on average) transported back to the nest by B. hainesi are very similar to those collected by three Trinervitermes species from West Africa (Sands 1961), but much shorter than the lengths of 20–60 mm reported by Leuthold et al. (1976) and Duncan and Hewitt (1989) for larger termite species. Baucaliotermes hainesi only forages on three plant species, despite many species being available. In contrast, Sands (1961) found that West African Trinervitermes species forage on more than 20 grass species and Nel and Hewitt (1969) report that H. mossambicus foraged on 17 plant species. Similar to H. mossambicus, Stipagrostis uniplumis is the most preferred grass species for B. hainesi (Becker and Getzin 2000). However, in this study, B. hainesi was only observed in one area (although being 7400 hectares in size), and foraging behaviour could well differ across the range, which will increase the number of plant species foraged upon.
Considering the density of foraging access points as well as the size of B. hainesi foraging parties and the amount of grass pieces found in the nest and tunnels (Tschinkel 2010), the importance of B. hainesi in grass removal, tree-seedling growth and herbaceous vegetation composition (Okullo and Moe 2012; Muller and Ward 2013; Støen et al. 2013) requires further study. In this study, we observed that areas with a high number of B. hainesi surface access points are characterised by being largely devoid of grasses (Fig. 2d), pointing to a potential role in veld degradation. Under future climate change scenarios, B. hainesi habitat will receive less rainfall, which will result in a lower herbivore carrying capacity (Midgley et al. 2001; IPCC 2001). Termites tend to have the biggest impacts on already degraded land. Therefore, and in particular for most communal lands where stocking rates are already unsustainable, termites will further exacerbate land degradation under increased aridification. In contrast, B. hainesi could have a positive effect at low densities, since they act as conveyors of soil and water to the surface (Turner et al. 2006), but this requires further study. More speculatively, increases in temperatures could lead to extended foraging periods, either as extended seasons or per night, which could further enhance the negative impacts. A comprehensive assessment of the impact of B. hainesi on the vegetation is therefore warranted. An experimental exclusion plot design will be able to determine B. hainesi impacts whilst simultaneously controlling for moisture levels, overgrazing and effects of the co-occurring termite, H. mossambicus.
References
Adam RA, Mitchell JD, van der Westhuizen MC (2005) Food preferences in laboratory colonies of the harvester termite, Trinervitermes trinervoides (Sjöstedt) (Termitidae: Nasutitermitinae). Afr Entomol 13:193–200
Adam RA, Mitchell JD, van der Westhuizen MC (2008) Aspects of foraging in the harvester termite, Trinervitermes trinervoides (Sjöstedt) (Termitidae: Nasutitermitinae). Afr Entomol 16:153–161
Adam RA, Mitchell JD, van der Westhuizen MC (2012) The role of the harvester termite, Trinervitermes trinervoides (Termitidae: Nasutitermitinae), in a semi-arid grassland ecosystem in South Africa: Nest populations and caste composition. Afr Entomol 20:239–251
Anderson MD (2004) Aardwolf adaptations: a review. Trans R Soc South Afr 59:99–104
Basson NCJ (1972) Beskadiging van natuurlike weiveld in die Noord-Vrystaat deur die grasdraertermiet, Hodotermes mossambicus (Hagen). Phytophylactica 4:67–72
Becker T, Getzin S (2000) The fairy circles of Kaokoland (North-West Namibia)—origin, distribution, and characteristics. Basic Appl Ecol 1:149–159
Bodin Ö, Tengö M, Norman A, Lundberg J, Elmqvist T (2006) The value of small size: loss of forest patches and ecological thresholds in southern Madagascar. Ecol Appl 16:440–451
Coaton WGH (1948a) Trinervitermes species—the snouted harvester termite. Union South Afr Dep Agric Bull 261:1–19
Coaton WGH (1948b) The snouted harvester termite. Farming South Afr 23:97–106
Coaton WGH (1953) Termites and their control in cultivated areas in South Africa. Union of South Africa, Department of Agriculture, Division of Entomology, Bulletin No. 305, Pretoria
Coaton WGH (1958) The hodotermitid harvester termites of South Africa. Dep Agric Sci Bull South Afr Entomol Ser 375:431–512
Coaton WGH, Sheasby JL (1973) National survey of the Isoptera of southern Africa. 1. The genus Baucaliotermes Sands (Termitidae: Nasutitermitinae). Cimbebasia Ser A 3:1–7
Dangerfield JM, McCarthy TS, Ellery WN (1998) The mound-building termite Macrotermes michaelseni as an ecosystem engineer. J Trop Ecol 14:507–520
Darlington JPEC (1982) The underground passages and storage pits used in foraging by a nest of the termite Macrotermes michaelseni in Kajiado, Kenya. J Zool Lond 198:237–247
Derichs P, du Plessis N (2012) Flowering plants of the Augrabies Falls National Park
Dougill AJ, Akanyang L, Perkins JS, Eckardt FD, Stringer LC, Favretto N, Atlhopheng J, Mulale K (2016) Land use, rangeland degradation and ecological changes in the southern Kalahari, Botswana. Afr J Ecol 54:59–67
Duncan FD, Hewitt PH (1989) Observations on the foraging behaviour of the harvester termite, Hodotermes mossambicus (Hagen) (Isoptera: Hodotermitidae). Bull Entomol Res 79:631–642
Eggleton P (2000) Global patterns of termite diversity. In: Abe T, Bignell DE, Higashi M (eds) Termites: evolution, sociality, symbioses, ecology. Kluwer Academic Publishers, Dordrecht, pp 25–51
Fuller C (1922) The termites of South Africa. S Afr J Nat Hist 3:70–131
Grohmann C (2010) Termite mediated heterogeneity of soil and vegetation patterns in a semi‐arid savanna ecosystem in Namibia. Ph.D Dissertation, Julius‐Maximilians University of Würzburg, Würzburg
Hager FA, Kirchner WH (2013) Vibrational long-distance communication in the termites Macrotermes natalensis and Odontotermes sp. J Exp Biol 216:3249–3256
Hager FA, Kirchner WH (2014) Directional vibration sensing in the termite Macrotermes natalensis. J Exp Biol 217:2526–2530
Harris WV (1971) Termites: their recognition and control. Longman, Bristol
Hartwig EK (1955) Control of snouted harvester termites. Farming South Afr 30:361–366
Haverty MI (1977) The proportion of soldiers in a termite colonies: a list and a bibliography (Isoptera). Sociobiology 2:199–216
Heidecker JL, Leuthold RH (1984) The organisation of collective foraging in the harvester termite Hodotermes mossambicus (Isoptera). Behav Ecol Sociobiol 14:195–202
Hoffman MT, Ashwell A (2001) Nature divided: land degradation in South Africa. University of Cape Town Press, Cape Town
IPCC (2001) Climate change 2001: The scientific basis. Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change. Houghton JT, Ding Y, Griggs DJ, Noguer M, van der Linden PJ, Dai X, Maskell K., Johnson CA (eds.) Cambridge University Press, Cambridge and New York
Jones CG, Lawton JH, Shachak M (1994) Organisms as ecosystem engineers. Oikos 69:373–386
Jouquet P, Dauber J, Lagerlöf J, Lavelle P, Lepage M (2006) Soil invertebrates as ecosystem engineers: intended and accidental effects on soil and feedback loops. Appl Soil Ecol 32:153–164
Kambhampati S, Eggleton P (2000) Taxonomy and phylogeny of termites. In: Abe T, Bignell DE, Higashi M (eds) Termites: evolution, sociality, symbioses, ecology. Kluwer Academic Publishers, Dordrecht, pp 1–23
Koehler CE, Richardson PRK (1990) Proteles cristatus. Mammalian Species. Am Soc Mammal 363:1–6
Kofoid CA (1934) Climate factors affecting the local occurrence of termite and their geographical distribution. In: Kofoid CA (ed) Termites and termite control. University of California Press, Berkeley, pp 13–21
Krishna K, Grimaldi DA, Krishna V, Engel MS (2013) Treatise on the Isoptera of the world. Bull. Am. Mus. Nat. Hist. 377:1–2704
Le Roux A (2005) South African Wildflower Guide No. 1: Namaqualand. Botanical Society of South Africa
Leuthold RH, Bruinsma O, van Huis A (1976) Optical and pheromonal orientation and memory for homing distance in the harvester termite Hodotermes mossambicus (Hagen). Behav Ecol Sociobiol 1:127–139
McMahan EA (1963) A study of termite feeding relationship using radio-isotopes. Ann Entomol Soc Am 56:74–82
Midgley G, Rutherford M, Bond W (2001) The heat is on…; impacts of climate change on plant diversity in South Africa. National Botanical Institute, Cape Town, South Africa
Mirenda JT, Eakins DG, Gravelle K, Topoff H (1980) Predatory behavior and prey selection by army ants in a desert-grassland habitat. Behav Ecol Sociobiol 7:119–127
Mouton PFN, Geertsema H, Visagie L (2000) Foraging mode of a group-living lizard, Cordylus cataphractus (Cordylidae). Afr Zool 35:1–7
Mucina L, Rutherford MC (eds) (2006) The Vegetation of South Africa, Lesotho and Swaziland. Strelitzia 19. South African National Biodiversity Institute, Pretoria
Muller K, Ward D (2013) Direct and indirect effects of termites on savanna tree-seedling growth. Plant Ecol 214:811–819
Nel JJC (1968) Die grasdraertermiet, Hodotermes mossambicus (Hagen), as plaag van natuurlike weiveld. J Entomol Soc South Afr 31:309–321
Nel JJC, Hewitt PH (1969) A study of the food eaten by a field population of the harvester termite, Hodotermes mossambicus (Hagen) and its relation to population density. J Entomol Soc South Afr 32:123–131
Ohiagu CE (1979a) Nest and soil populations of Trinervitermes spp. with particular reference to T. geminatus (Wasmann) (Isoptera), in Southern Guinea Savanna near Mokwa. Nigeria. Oecologia 40:167–178
Ohiagu CE (1979b) A quantitative study of seasonal foraging by the grass harvesting termite, Trinervitermes geminatus (Wasmann) (Isoptera, Nasutitermitinae), in Southern Guinea Savanna, Mokwa, Nigeria. Oecologia 40:179–188
Ohiagu CE, Wood TG (1976) A method for measuring rate of grass-harvesting by Trinervitermes geminatus (Wasmann) (Isoptera, Nasutitermitinae) and observation on its foraging behaviour in Southern Guinea Savanna, Nigeria. J Appl Ecol 13:705–713
Okullo P, Moe SR (2012) Termite activity, not grazing, is the main determinant of spatial variation in savanna herbaceous vegetation. J Ecol 100:232–241
Olugbemi BO (2010) Influence of food on recruitment pattern in the termite, Microcerotermes fuscotibialis. J Insect Sci 10:154
Olugbemi BO, Malaka SLO (2007) The effect of food on pheromonal communication in the termite, Microcerotermes fuscotibialis Sjostedt. Afr J Ecol 45:216–219
Pomeroy DE (1976) Some effects of mound-building termites on soils in Uganda. J Soil Sci 27:377–394
Richardson PRK (1987) Aardwolf: the most specialized myrmecophagous mammal? South Afr J Sci 83:643–646
Sands WA (1961) Foraging behaviour and feeding in five species of Trinervitermes in West Africa. Entomol Exp Appl 4:277–288
Sands WA (1965) A revision of the termite subfamily Nasutitermitinae (Isoptera, Termitidae) from the Ethiopian region. Bull Br Mus Nat Hist Entomol Suppl 4:1–172
Šobotník J, Jirošová A, Hanus R (2010) Chemical warfare in termites. J Insect Physiol 56:1012–1021
Støen OG, Okullo P, Eid T, Moe SR (2013) Termites facilitate and ungulates limit savanna tree regeneration. Oecologia 172:1085–1093
Taylor WA, Lindsey PA, Skinner JD (2002) The feeding ecology of the aardvark Orycteropus afer. J Arid Environ 50:135–152
Thorne BL (1982) Polygyny in termites: multiple primary queens in colonies of Nasutitermes corniger (Motschuls) (Isoptera: Termitidae). Insect Soc 29:102–117
Tschinkel WR (2010) The foraging tunnel system of the Namibian desert termite, Baucaliotermes hainesi. J Insect Sci 10:1–17
Turner JS, Marais E, Vinte M, Mudengi A, Park WL (2006) Termites, water and soils. Agricola 16:40–45
Urban EK, Fry CH, Keith S (1986) The Birds of Africa, vol 2. Academic Press, London
Uys V (2002) A guide to the termite genera of southern Africa. Plant Protection Research Institute, Handbook No. 15, Agricultural Research Council, Pretoria
Van Ark H (1969) Control of Hodotermes mossambicus in the Karroo (Hodotermitidae, Isoptera). Phytophylactica 1:41–50
Verma M, Sharma S, Prasad R (2009) Biological alternatives for termite control: a review. Int Biodeterior Biodegrad 63:959–972
Whitford WG, Ludwig JA, Noble JC (1992) The importance of subterranean termites in semiarid ecosystems in South-Eastern Australia. J Arid Environ 22:87–91
Acknowledgments
We thank Henriëtte van der Linden, Dawie Snyman and Martin de Wit for assistance with fieldwork and comments on an earlier draft. We are grateful to Brigitte Braschler for assistance with ant identification. We thank the landowners, Isak and Hannes Nel, for permission to work on the farm Tierkop. We thank two anonymous reviewers and Yves Roisin whose suggestions helped improve and clarify this manuscript. SG acknowledges support from the National Research Foundation (Grant 87843).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
40_2016_488_MOESM3_ESM.mp4
Video 2. An ant, of the species Tetramorium grandinode (subfamily Myrmicinae), caught a Baucaliotermes hainesi soldier then encounters the foraging column again and makes a quick turnaround. (MP4 4660 kb)
40_2016_488_MOESM4_ESM.mp4
Video 3. Baucaliotermes hainesi soldiers squirting turpentine at an insect that landed close to a foraging column. (MP4 3021 kb)
Rights and permissions
About this article

Cite this article
Geerts, S., van der Linden, J. & van der Linden, E. The ecology and foraging behaviour of the harvester termite, Baucaliotermes hainesi in semi-arid grasslands in the northwestern interior of South Africa. Insect. Soc. 63, 457–465 (2016). https://doi.org/10.1007/s00040-016-0488-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00040-016-0488-9