
Abstract
The native Yellowstone cutthroat trout (Oncorhynchus clarkii bouvieri; YCT) of Yellowstone Lake, Yellowstone National Park, Wyoming, USA, presents a metapopulation that inhabits an ecologically simple, uniquely pristine, and highly protected environment. Recent investigation of a key YCT spawning stock’s 1977–1992 dynamics, before predation from an illegally introduced and growing lake trout (Salvelinus namaycush) population measurably affecting YCT population dynamics, showed the new climate regime that began in the 1970s largely explained declining YCT spawning runs, although the concurrent and growing predation effect soon became dominant. The present study's time series analyses of annual (1969–2010) gillnet catches from 11 lake locations showed the metapopulation’s age-5 YCT abundance rapidly declined to low levels 7 years after the spawning stock’s (median YCT age, 5 years) similar decline. Since the 1980s, the Yellowstone region has experienced increasing drought that has reduced wetlands and associated tributaries used by the lake’s spawning YCT. While small tributaries were first to become unsuitable for YCT spawning and early rearing, the spawning stock’s tributary was of moderate size and its population persisted later. Two conditions are necessary for YCT metapopulation recovery to historically high levels: (1) climatic conditions must return to those prevalent around 1980 and (2) the lake trout population must be reduced to its mid-1990s levels. The most recent time of YCT metapopulation abundance and broadest spatial extent occurred under those simultaneous conditions. Similarities between the metapopulation’s structure and dynamics and those reported for metapopulations of anadromous salmonids are described, including adverse effects of ongoing climate change.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
For many salmonid fish species, the metapopulation, whose foundational concept is attributed to Levins (1969), can be simply described as a group of spatially separated populations that converge during some part of the species’ life history. Population separation occurs during spawning and early rearing of young in specialized habitats and population convergence during subsequent growth and maturation elsewhere, notably in regions of high species’ productivity (e.g., Schtickzelle and Quinn 2007). The metapopulation’s spatial extent is indicated by movements of its individuals among locations of population separation and convergence. Furthermore, in the sense of Pulliam (1988), the most stable (and likely largest) populations are the source of recruits for the least stable (and likely small) sink populations. These recruits are called strays, and straying is important to sink population maintenance and establishment or restoration of populations. Availability of recruits depends upon the source population’s compensatory reserve (Rose et al. 2001); specifically, its excess reproduction capacity is not needed then to offset stochastic effects on its own mortality. Consequently, metapopulation persistence ultimately depends upon maintaining excess compensatory reserve in its source populations.
The native Yellowstone cutthroat trout (Oncorhynchus clarkii bouvieri; YCT) of Yellowstone Lake, Wyoming, USA, ostensibly spawned (around June) in 68 of the lake’s approximately 124 tributaries during the 1970s and 1980s, as well as in the lake’s outlet stream, the Yellowstone River (Gresswell and Varley 1988). Based on these and other data described below, Gresswell et al. (1994) concluded the YCT (an iteroparous fish and obligate stream spawner) of Yellowstone Lake constituted a metapopulation. After all, soon after emerging from tributary gravels in July or August, most age-0 YCT emigrate to the lake where together they grow to maturity over ≥ 3 years (Kaeding and Koel 2011).
Located wholly within Yellowstone National Park, Yellowstone Lake and its catchment (partly located in a federally protected wilderness area) present an ecologically simple and remarkably uncommon aquatic environment today, one where typical human impacts of fishery over-harvest, water pollution and water removal, habitat degradation, dams and their adverse effects, and the stocking of hatchery fish do not exist and thus are not potential confounding factors in the examination of YCT metapopulation dynamics. Furthermore, those YCT have shown a profound and nearly continuous decline since the 1970s, which Koel et al. (2005) speculated resulted from one or more of three factors: (1) predation from an illegally introduced and growing population of lake trout (Salvelinus namaycush), a nonnative species discovered in the lake in 1994; (2) debilitating effects on infected fish of “whirling” disease, found in the lake’s YCT in 1998; (3) a regional drought that those authors believed had begun in 1998 and harmed YCT reproduction. Results of subsequent studies assessed by Kaeding (2020) indicated the relative importance of each factor to the decline.
Kaeding (2020) dismissed whirling disease as an important factor but tentatively concluded that both lake trout predation and climate variation could have equally reduced YCT survival, at least across the preceding decade of data records or perhaps longer. Because population effects of predation and climate were therefore statistically inseparable, contemporary model selection techniques (e.g., Hilborn and Mangel 1997) were not useful. Consequently, Kaeding (2020) used a population model that had a local climate index (described below) as its only driver (regulating age-0 YCT survival) and Bayesian methods to examine a key YCT spawning stock from 1977 to 1992, that is, before lake trout discovery and, as argued, before their predation measurably affected YCT population dynamics. The YCT spawning stock was that of Clear Creek, a Yellowstone Lake tributary of moderate size.
Kaeding (2013a) had shown the climate index (specifically, total annual mean-daily air temperatures > 0 °C at the lake’s north shore) was negatively correlated with Clear Creek discharge and positively correlated with creek temperature during the YCT spawning and early rearing season. Furthermore, the index’s trajectory illustrated increasingly dry and warm climatic conditions evident across the Yellowstone Lake region during recent decades. Using his model for population forecasting through 2007 (the final empirical data year), Kaeding (2020) showed the new climate regime that began in the 1970s (described below) had an overarching effect and largely explained declining YCT spawning runs, although a growing predation effect became evident soon after lake trout discovery and quickly became the major YCT population driver.
Objectives of the present study were to: (1) examine data for the lake’s annual (1969–2010) YCT gillnet catches, which presumptively represented the metapopulation; (2) comprehensively assess those results, along with conclusions from preceding studies, in the context of climate-change and nonnative-piscivore impacts on YCT metapopulation dynamics; (3) identify requirements for YCT metapopulation recovery to historically high levels; (4) describe similarities between YCT metapopulation structure and dynamics and those reported for anadromous salmonids, including adverse effects of ongoing climate change and how interpretations of metapopulation dynamics based on time series data for a presumed “index” population can affect management actions.
Methods
Study area and data collection
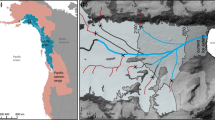
The Yellowstone Lake study area, the lake’s fishes, gillnetting procedures and data taken from netted YCT were detailed by Kaeding and Koel (2011). In brief, beginning in 1969, one net was set in mid-September at each of 21 sampling sites around the lake. In 1977, four nets were set at each site. Finally, beginning in 1978 and continuing through 2010 (the sampling program’s last year), 5 nets were set at each of 11 of the original 21 sites (Fig. 1).

Map of Yellowstone Lake, Yellowstone National Park, Wyoming, showing the 11 sampling sites (●) and their codes, as well as Clear Creek
Those 11 sites were chosen based on among-site similarity in YCT catches and a preponderance of the nonnative longnose sucker (Catostomus catostomus) catches (Jones et al. 1977). Set in the afternoon and retrieved the next morning, gill nets were 38 m long and 1.5 m deep and consisted of five ordered, 7.6-m panels of 19-mm, 25-mm, 32-mm, 38-mm or 51-mm (bar measure) monofilament netting. Beginning with smallest mesh in water ~ 1.5 m deep, nets were set perpendicular to shoreline and ~ 100 m apart. All netted YCT were measured to total length (TL, mm).
Data reduction
The data were those collected at the 11 sites between 1969 and 2010, when 1854 gillnet nights (Supplementary Table S1) caught 22,454 YCT (12.1 YCT/net night). The aggregate length-frequency distribution for those fish (Fig. 2a) had conspicuous size classes centered around 180 mm TL and 250 mm TL, which Kaeding and Koel (2011) showed predominantly consisted of age-2 and age-3 YCT, respectively. (Supplementary Figure S1 provides annual TL distributions.)

a Length-frequency distribution for 22,454 Yellowstone cutthroat trout in gillnet catches, Yellowstone Lake, 1969–2010; b age-TL relation (n = 2248) used to assign ages to each fish; c aggregate frequency distribution for the 500 ages assigned to each fish
Because YCT catch rate would be based on abundance of a single age class, an age-TL key (Hilborn and Walters 1992) was used to assign ages (years) to each fish. The key’s scale-age data were those determined for these YCT by Kaeding and Koel’s (2011) analyst 8, which provided the most accurate age estimates, and were pooled in consecutive 10-mm TL bins (Fig. 2b). For each captured YCT, an arbitrary 500 random draws (with replacement) were made from among ages in its respective TL bin of the key. That accounted for measurement uncertainty associated with age assignments and provided 500 sets of age-based catches for subsequent analyses. The TL measurements were assumed to have no uncertainty. The 259 YCT (1.2%) that had TL ≥ 499 mm (a size class that had scant scale-aged fish) were assigned age 10.
The aggregate age frequency showed YCT fully recruited into the gillnet catch at age 5 (Fig. 2c; Supplementary Text S1). Therefore, age-5 abundance was calculated as total annual age-5 catch at each site for each of the 500 sets of age-based catches, divided by the site-specific number of net nights. Pearson’s product moment correlations between median age-5 abundance across study years (catch-rate analyses restricted to data for 1977–2010, when more than one net had been set at each site) indicated highly significant (p ≤ 0.01) associations among all sites (Supplementary Table S2). Accordingly, age-5 abundances for each year were averaged among all 11 sites, thereby giving the metapopulation’s age-5 abundance. To broadly show the study area’s climate trajectory over the past century, the climate index and its 5-point moving average were calculated for 1920–2020 using air temperature data that included statistical estimates for the few previously missing daily measurements (Kaeding 2013a).
Age-5 abundance and run-size trajectories and their change points
The YCT spawning run was that of a moderate-size tributary, whereas age-5 abundance ostensibly represented aggregate spawning stocks of all the lake’s tributaries, which varied in size from rivulets up to the upper Yellowstone River itself (Gresswell et al. 1997). Furthermore, median age of YCT in spawning runs was 5 years (Kaeding and Koel 2011). I therefore hypothesized that changing climate would first degrade YCT spawning and early-rearing habitats of the smallest tributaries, and their spawning stocks would be eliminated. Even so, age-5 abundance may not concurrently decline because source populations still had excess compensatory reserve. Nevertheless, a marked decline in age-5 abundance would be expected should those adverse climatic effects increase and excess reserves become exhausted. Thus, understanding the temporal relationship between spawning run and age-5 abundance declines was critical to interpreting metapopulation dynamics.
The statistical package BEAST (Bayesian Estimator of Abrupt change, Seasonality, and Trend)Footnote 1 and MATLAB platform (MathWorks 2021) were used to identify abrupt change points in time series for age-5 abundance and run size. BEAST is a model-averaging algorithm that decomposes time series data into individual components and estimates the location and probability of abrupt change points (Zhao et al. 2019). Because age-5 abundance ostensibly represented aggregate spawning stocks of all the lake’s tributaries, in contrast with Clear Creek’s single YCT spawning run, fitting of Kaeding’s (2020) climate-driven model could not provide equally unambiguous results.
Results
Age-5 abundance and run-size trajectories and change points
Age-5 abundance was mostly stable between 1977 and 1998, apart from its marked increase in 1981 (Fig. 3a); those particular age-5 YCT were produced in 1975 (this study’s aging convention had first-year fish as age-0), when mean June discharge was especially low (Supplementary Text S2). In any case, age-5 abundance declined rapidly between 1998 and 2002 and remained low thereafter. The spawning run’s trajectory showed no pronounced peak in 1981 and dropped to small sizes between 1987 and 1994, near which it remained through 2007 (the final empirical data year; Fig. 3b). The brief bump-up in run size in the late 1990s resulted from preceding climatic conditions that were favorable to YCT reproduction (Kaeding 2020).

a Median annual age-5 Yellowstone cutthroat trout abundance (solid line) and the full range in age-5 abundance estimates (dashed lines), 1977–2010, and b number of Yellowstone cutthroat trout in the annual Clear Creek spawning run, 1977–2007 (taken from Kaeding 2020); dashed vertical lines indicate change points, the probabilities for which are also indicated
The estimated locations (calendar years) and associated probabilities of change points were 2001 and 0.74 for age-5 abundance (Fig. 3a) and 1987 (0.37) and 1994 (0.32) for the spawning run, respectively (Fig. 3b; see also Supplementary Text S3). However, only the run’s 1994 change point indicated an end of time series decline, as did that for age-5 abundance in 2001.
Preceding century’s climate indexes
Although the climate index varied widely across the preceding century, it showed a decidedly increasing trajectory between 1980 and 2020 (Fig. 4).

Climate indexes for years 1920–2020 and their 5-point moving average (solid line)
Discussion
Do the YCT constitute a metapopulation?
Defining characteristics of an Oncorhynchus metapopulation were provided by Schtickzelle and Quinn (2007) for sockeye salmon (O. nerka) that spawned and early reared in a large complex of lakes that drained into Alaska’s Bristol Bay. Those authors concluded the sockeye constituted a metapopulation because: (1) spawning habitats were discrete and spawning populations spatially separated; (2) measurable asynchrony in dynamics was evident among populations (ostensibly, highly improbable perfect statistical synchrony would instead indicate a single population); (3) movements of fish linked populations. Although Schtickzelle and Quinn (2007) provided no evidence of local population extirpations or recolonizations—likewise considered characteristic of metapopulations (Levins 1969)—those events undoubtedly happened during prehistoric development of the sockeye metapopulation.
Schtickzelle and Quinn’s (2007) criteria are consistent with characteristics of Yellowstone Lake’s YCT metapopulation, specifically: (1) YCT ostensibly spawned in 68 lake tributaries during the 1970s and 1980s (Gresswell and Varley 1988), thus indicating numerous former, spatially separated, reproducing populations; (2) spawning times for these populations varied because of differences in aspect and size among tributary drainages (Gresswell et al. 1997), thus demonstrating asynchrony of the populations’ dynamics; (3) mark-recapture studies indicated roughly 5% of mature YCT strayed among lake tributaries during spawning (Gresswell et al. 1994), thus showing YCT movements among populations. Furthermore, Teisberg et al. (2014) found that, among 22 Yellowstone Lake tributaries used by spawning YCT and visited by foraging bears (Ursus spp.) during 1997–2000, only 16 of those tributaries hosted spawning YCT during 2007–2009 and only 2 did so in all 3 years. Those observations evidenced local YCT population extirpations and possible attempted recolonization events and also substantiated metapopulation occurrence (Levins 1969).
Climate regime shift, drought and tributary flows
Mantua et al. (1997) reported a 1977 shift in the Pacific Decadal Oscillation (PDO), a robust index of North Pacific Basin climate. Based on subsequent analyses of 31 climatic and 69 biological time series, Hare and Mantua (2000) confirmed the 1977 shift and provided a comprehensive review of numerous supporting studies. That 1977 shift heralded a new climate regime in North Pacific and North American regions, accompanied by drought. The climate index’s broadly increasing trajectory over 1980–2020 corroborated that shift.
For 17 Missouri River Basin locations, including the Yellowstone River drainage, Martin et al. (2020) reported persistent, severe drought events increased since the 1980s eventually exceeding conditions during the Dust Bowl period (the 1930s) and that warming air temperatures have accompanied increased regional precipitation deficits and decreased tributary flows. Martin et al. (2020) concluded increasing drought will continue under ongoing climate change. At the same time, the drought’s drying of wetlands at Yellowstone National Park locations, including the Yellowstone Lake drainage, caused severe declines in four once-common amphibian species (McMenamin et al. 2008). Those former wetlands often had small tributaries used by spawning YCT from the lake. Teisberg et al. (2014) had documented elimination of several of those spawning populations.
The decline in Yellowstone Lake’s tributary flows was driven by that climate shift and subsequent climatic regime. Kaeding (2013a) showed the climate index was negatively correlated with Clear Creek discharge during YCT spawning. Furthermore, the overarching climate trend (Mantua et al. 1997) is part of a broader, ongoing pattern of climate warming especially evident in western North America (Vose et al. 2017). Since the investigation of Hare and Mantua (2000), the PDO has undergone substantial variations between cool and warm phases, yet much of western North America (including the Yellowstone region) has shown a general warming trend, accompanied by substantial variation in annual precipitation (Easterling et al. 2017; Vose et al. 2017). Those climatic trends were recently substantiated for North America (Jones and Driscoll 2022). A return to climates more favorable to YCT reproduction is highly unlikely (Vose et al. 2017).
Metapopulation dynamics
In metapopulation theory (e.g., Hanski and Simberloff 1997), availability of excess compensatory reserve is determined by stochastic processes that are demographic within populations and environmental at spatial scales that range from the sink population’s patch to the species’ entire distribution. Even so, the least stable sink population cannot persist if: (scenario 1) the source population’s compensatory reserve remains diminished for too long or (scenario 2) spawning and early-rearing habitats of the sink population remain diminished for so long that the population can no longer be sustained despite available source recruits. For either scenario, however, if the stochastic variables causing diminishment have no overarching temporal trend—that is, they simply vary independently around eternal mean values—both scenarios are precluded and the sink population will persist. Alternatively, if the overarching temporal trend of those stochastic variables is toward effecting either scenario, sink populations would be eliminated according to their susceptibility until only the source population remained but was then likewise eliminated. It is impossible for a sink population to recolonize a former source population when the diminishing stochastic variable has an overarching trend.
Clear Creek’s spawning run decline to low levels preceded that of metapopulation age-5 abundance by 7 years and thus was informative of metapopulation dynamics. Specifically, owing to their vulnerability to drying, wetlands’ small tributaries were first to become unsuitable for YCT spawning and early rearing (but not necessarily entirely dry) and their YCT populations were eliminated. But Clear Creek’s drainage was of moderate size and its YCT population persisted. Even so, there came a time when the metapopulation’s excess compensatory reserve was inadequate to offset combined adverse effects of climate change and lake trout predation on Clear Creek’s YCT spawning run, and the run rapidly declined.
Yellowstone Lake’s YCT metapopulation is evidently functioning under conditions of scenario 2, a conclusion based on: (1) the empirically supported environment-recruitment relationship for Clear Creek’s spawning run that indicated an overarching climatic driver (Kaeding 2020); (2) climate projections cited by the preceding studies that indicated adverse effects of that driver will increase under ongoing climate change; (3) loss of YCT sink populations that had spawned and early reared in former wetland rivulets and other small lake tributaries; (4) near elimination of the spawning run of Clear Creek, a moderate-size tributary. Clearly, metapopulation persistence is threatened by the overarching climatic effect. Furthermore, given the predicted climate trend, the source population for Yellowstone Lake’s YCT metapopulation will eventually occur in the upper Yellowstone River and its tributaries above the lake, where suitable YCT spawning and rearing habitats may remain but lake trout are absent. Based on recent observations (Koel et al. 2020), one could conclude that condition is now evident.
Requirements for metapopulation recovery
From the fishery management perspective, whether climate or lake trout predation is now having the most adverse effect on YCT metapopulation dynamics is an academic question. Elimination of lake trout, were that possible, would not provide climatic conditions more favorable to YCT reproduction, and attaining such climatic conditions would not eliminate lake trout predation. For metapopulation recovery to historic-high levels, effects of both drivers must be mitigated.
The present study indicates two conditions are necessary for that recovery: (1) climatic conditions must return to those prevalent around 1980 and (2) the lake trout population must be reduced to its mid-1990s levels. Historically high YCT metapopulation abundances and broadest spatial extent occurred under those simultaneous conditions. Even so, in the mid-1990s there was already an apparent emerging effect of lake trout predation on Clear Creek’s spawning run because subsequent runs were consistently smaller than median run sizes predicted by Kaeding’s (2020) climate-driven population model.
In their assessment of effects of shifting baseline syndrome (SBS) in conservation, Papworth et al. (2008) made clear that, for effective conservation, “Accurate assessments of change are required when conservation aims to restore former conditions.” They defined SBS as “a cautionary tale referring to changing human perceptions of biological systems due to loss of experience about past conditions.” Age-5 abundances in the mid-1990s reveal objective baseline conditions and provide an unambiguous goal for YCT metapopulation recovery. In addition, Ruzycki et al. (2003) provided an estimate of lake trout population in 1996, 2 years after discovery: 8300 age 3–5 fish and 3000 age 6–23 fish. To quote Soga and Gaston (2018), “Arguably, the fundamental driver of SBS is the lack, or paucity, of relevant historical data on the natural environment.” Such data are not lacking for Yellowstone Lake’s YCT metapopulation, however. Thus, SBS is avoidable when setting recovery goals.
Koel et al. (2020) detailed the history of management actions intended to control Yellowstone Lake’s lake trout population since its 1994 discovery. Commercial-scale gill netting removed 3.35 million lake trout from the lake through 2019, when the estimated population of lake trout ≥ age 2 that remained was 674,000. The central assumption of the park’s Native Fish Conservation Plan (Koel et al. 2010) is that gill netting alone will sufficiently reduce the lake trout population and thereby lead to natural recovery of Yellowstone Lake’s YCT metapopulation, although not specifically to historically high levels. In any case, that assumption ignores the overarching effect of climate and climate change on the metapopulation’s dynamics.
Similarities to anadromous salmonid metapopulations
Schtickzelle and Quinn (2007) pointed out that the metapopulation concept “is rarely used, and barely tested” for anadromous salmonids. That point remains noteworthy. After all, as life history strategies, semelparity (as in sockeye) and iteroparity (as in YCT) are now seen as endpoints along a continuum (Hughes 2017). Furthermore, Cole (1954) long ago pointed out that the negative effect of semelparity on population dynamics (specifically, spawner death) is offset by recruiting one additional, biologically equivalent adult to the subsequent spawning population. That recruiting is a function of the population’s compensatory reserve. Perhaps researchers of population dynamics of anadromous salmonids are not aware that they are studying metapopulations.
Jones et al. (2020) recently reported associations among eight climate metrics statistically reconstructed at stream-drainage scales and productivity of 15 Chinook salmon (O. tshawytscha) populations that spawned and early reared in streams entering Cook Inlet, Alaska. Those authors found negative associations for precipitation during spawning and positive associations for precipitation during juvenile rearing for all populations. The effect of stream temperature ranged from positive to negative among populations, although productivity steeply declined for two populations when maximum weekly water temperatures during spawning increased from 18 °C to 22 °C across study years. Jones et al. (2020) cited studies indicating climatic drivers of precipitation and temperature that negatively affected the Chinook populations will increase under ongoing climate change. While their study seemingly was of a Chinook metapopulation, the authors did not use that term. Straying probably occurred among populations because Chinook of unspecified hatchery origin were common enough to warrant their exclusion from analyses. In any case, like YCT, the putative Cook Inlet Chinook metapopulation (as I term it here) appears to be functioning under conditions of scenario 2, based on: (1) empirically supported environment-recruitment relationships that indicated overarching climatic drivers; (2) climate projections that indicated adverse effects of those drivers will increase under ongoing climate change; (3) probable heat deaths of spawners in two populations. Persistence of this metapopulation is plainly threatened.
Bouchard et al. (2022) recently estimated the extent of inter-population synchrony among 18 Atlantic salmon (Salmo salar) populations that composed a metapopulation in Brittany, France. They found that measurable synchrony was common and driven by an effect of summer stream flow on fry survival during fry emergence from redds, specifically, low flows reduced survival and thus synchronized population dynamics at a broad spatial scale. Bouchard et al. (2022) pointed out that such spatial synchrony among populations (that is, the Moran effect) increased risk of simultaneous population extinction, particularly owing to extreme climatic events that are likely to increase in the Brittany region with climate change. Thus, metapopulations of anadromous salmonids are beginning to be recognized and their dynamics critically examined.
Metapopulation index populations
Appreciable synchrony of dynamics would be expected among anadromous salmonid populations that had roughly similar age structures and vital rates, comparably suitable spawning and rearing habitats, and were aggregated such that they shared major environmental (in particular, climatic) drivers. Even so, Cooper and Mangel (1999) used a simple source-sink metapopulation model that considered those biological and ecological conditions plus the effect of dispersal among populations to show how variation in salmonid metapopulation characteristics (that is, its structure) may affect population dynamics. Among their results were examples of how interpretation of metapopulation dynamics based on time series data for a presumed “index” population can be misleading and thus result in incorrect management actions.
For the YCT metapopulation of Yellowstone Lake, that index population was Clear Creek’s spawning run, which was chosen a century ago because it was useful as a spawning stock for hatchery operations, not for population monitoring. When new management policies ended hatchery operations in the 1950s, run monitoring continued for research purposes. As it turned out, that spawning run was informative of metapopulation dynamics (Kaeding 2013b). Moreover, the present study showed another informative index was provided by gillnet data, specifically, age-5 YCT abundance. As metapopulation indexes, however, age-5 YCT abundance lagged 7 years behind the spawning run. Finally, although not originally intended as such, monitoring of YCT spawning in rivulets and other small tributaries visited by foraging bears provided a particularly useful index because those sink populations were the “miner’s canary” of metapopulation decline.
Data availability
The data that support the findings of this study are openly available in https://data.mendeley.com/datasets/sfpnd3xy2r/2
Notes
BEAST version 1.1.2.60 (4.1 MB) by Kaiguang is available at the MATLAB File Exchange and https://github.com/zhaokg/Rbeast
References
Bouchard C, Mathieu B, Clément L, Carlson SM (2022) Synchrony in population dynamics of juvenile Atlantic salmon: analyzing spatiotemporal variation and the influence of river flow and demography. Can J Fish Aquat Sci 79:782–794. https://doi.org/10.1139/cjfas-2021-0017
Cole L (1954) The population consequences of life history phenomena. Q Rev Biol 29:103–137
Cooper AB, Mangel M (1999) The dangers of ignoring metapopulation structure for the conservation of salmonids. Fish Bull 97:213–226
Easterling DR, Kunkel KE, Arnold JR et al (2017) Precipitation change in the United States climate science special report: fourth national climate assessment. In: Wuebbles DJ, Fahey DW, Hibbard KA et al (eds) US. Global change research program, Washington, DC, USA
Gresswell RE, Varley JD (1988) Effects of a century of human influence on the cutthroat trout of Yellowstone Lake. Am Fish Soc Symp 4:45–52
Gresswell RE, Liss WJ, Larson GL (1994) Life-history organization of Yellowstone cutthroat trout (Oncorhynchus clarki bouvieri) in Yellowstone Lake. Can J Fish Aquat Sci 51:298–309
Gresswell RE, Liss WJ, Larson GL, Bartlein PJ (1997) Influence of basin-scale physical variables on life history characteristics of cutthroat trout in yellowstone lake. North Am J Fish Manag 17:1046–1064
Hanski I, Simberloff D (1997) The metapopulation approach, its history, conceptual domain, and application to conservation. In: Hanski I (ed) Metapopulation biology Ecology, genetics, and evolution. Academic Press, San Diego, USA
Hare SR, Mantua NJ (2000) Empirical evidence for North Pacific regime shifts in 1977 and 1989. Prog Oceanogr 47:103–145
Hilborn R, Mangel M (1997) The ecological detective: confronting models with data. Princeton, Princeton University
Hilborn R, Walters CJ (1992) Quantitative fisheries stock assessment: choice, dynamics and uncertainty. Chapman and Hall, New York, p 570
Hughes PW (2017) Between semelparity and iteroparity: empirical evidence for a continuum of modes of parity. Ecol Evol 7:8232–8261
Jones JA, Driscoll CT (2022) Long-term ecological research on ecosystem responses to climate change. Bioscience 72:814–826
Jones RD, Varley JD, Jennings DE et al (1977) Fishery and aquatic management program in yellowstone national park: annual technical report for 1976. Fish and Wildlife Service, Yellowstone National Park Wyoming, US
Jones LA, Schoen ER, Shaftel R et al (2020) Watershed-scale climate influences productivity of Chinook salmon populations across southcentral Alaska. Glob Change Biol 26:4919–4936. https://doi.org/10.1111/gcb.15155
Kaeding LR (2013a) A climate-based index of historic spawning-stream hydrology. River Res Appl 29:380–388
Kaeding LR (2013b) Informative correlations among metrics of Yellowstone Lake cutthroat trout caught by two quantitative methods across three recent decades. North Am J Fish Manag 33:882–886
Kaeding LR (2020) New climate regime started and further shaped the historic Yellowstone Lake cutthroat trout population decline commonly attributed entirely to nonnative lake trout predation. Aquat Ecol 54:641–652
Kaeding LR, Koel TM (2011) Age, growth, maturity, and fecundity of Yellowstone Lake cutthroat trout. Northwest Sci 85:431–444
Koel TM, Bigelow PE, Doepke PD et al (2005) Nonnative lake trout result in Yellowstone cutthroat trout declines and impacts to bears and anglers. Fisheries 30(11):10–19
Koel TM, Arnold JL, Bigelow PE et al (2020) Yellowstone Lake ecosystem restoration: a case study for invasive fish management. Fishes 5:18. https://doi.org/10.3390/fishes5020018
Koel TM, Arnold JL, Bigelow PE et al (2010) Native Fish Conservation Plan: Environmental Assessment. US. Department of the Interior, National Park Service, Yellowstone National Park, WY, USA. Available online: https://parkplanning.nps.gov/projectHome.cfm?projectID=30504
Levins R (1969) Some demographic and genetic consequences of environmental heterogeneity for biological control. Bull Entomol Soc Am 15(3):237–240
Mantua NJ, Hare SR, Zhang Y et al (1997) A Pacific interdecadal climate oscillation with impacts on salmon production. Bull Am Meteor Soc 78:1069–1079
Martin JT, Pederson GT, Woodhouse CA et al (2020) Increased drought severity tracks warming in the United States’ largest river basin. Proce Nat Acad Sci US 117(21):11328–11336
MathWorks (2021) MATLAB, the language of technical computing, version 9.10.0.2015706 (R2021a). MathWorks, Natick, MA
McMenamin SK, Hadly EA, Wright CK (2008) Climatic change and wetland desiccation cause amphibian decline in Yellowstone National Park. Proce Nat Acad Sci US 105(44):16988–16993
Papworth SK, Rist J, Coad L, Milner-Gulland EJ (2008) Evidence for shifting baseline syndrome in conservation. Conserv Lett 2:93–100
Pulliam HR (1988) Sources, sinks, and population regulation. Am Nat 132:652–661
Rose KA, Cowan JH Jr, Winemiller KO et al (2001) Compensatory density dependence in fish populations: importance, controversy, understanding and prognosis. Fish Fish 2:293–327
Ruzycki JR, Beauchamp DA, Yule DL (2003) Effects of introduced lake trout on native cutthroat trout in Yellowstone Lake. Ecol Appl 13:23–37
Schtickzelle N, Quinn TP (2007) A metapopulation perspective for salmon and other anadromous fish. Fish Fish 8:297–314
Soga M, Gaston KJ (2018) Shifting baseline syndrome: causes, consequences, and implications. Front Ecol Environ 16(4):222–230. https://doi.org/10.1002/fee.1794
Teisberg JE, Haroldson MA, Schwartz CC et al (2014) Contrasting past and current numbers of bears visiting Yellowstone cutthroat trout streams. J Wildl Manag 78(2):369–378. https://doi.org/10.1002/jwmg.667
Vose RS, Easterling DR, Kunkel KE et al (2017) Temperature changes in the United States. In: Wuebbles DJ, Fahey DW, Hibbard KA et al (eds) Climate Science Special Report: Fourth National Climate Assessment. Global Change Research Program, Washington, DC, USA
Zhao K, Wulder MA, Hu T et al (2019) Detecting change-point, trend, and seasonality in satellite time series data to track abrupt changes and nonlinear dynamics: A Bayesian ensemble algorithm. Remote Sens Environ 232:111181. https://doi.org/10.1016/j.rse.2019.04.034
Kaeding L (2022) Yellowstone Lake cutthroat trout, annual gillnet data, 1969–2010. Mend Data. https://doi.org/10.17632/sfpnd3xy2r.2
Acknowledgements
I thank Todd Koel, Yellowstone National Park, and his staff for providing the most recent gillnet data; Phil Farnes, Snowcap Hydrology, for improved air temperature data used to calculate the climate index; Jim Mogen for Figure 1; and three anonymous reviewers for constructive comments on a preceding version of this report. I especially thank the innumerable employees of the US Fish and Wildlife Service and National Park Service, and volunteers, who collected these data between 1969 and 2010.
Author information
Authors and Affiliations
Contributions
The author compiled and analyzed the data and wrote the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The author did not receive financial support from any organization for the submitted work, nor has he competing interests to declare that are relevant to the content of this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Kaeding, L.R. Climate-change and nonnative-piscivore impacts on a renowned Oncorhynchus metapopulation, requirements for metapopulation recovery, and similarities to anadromous salmonid metapopulations. Aquat Sci 85, 88 (2023). https://doi.org/10.1007/s00027-023-00987-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00027-023-00987-3