
Abstract
Winter stream temperatures, though infrequently studied, exert important influences on aquatic communities. To quantify effects of watershed physical characteristics on stream winter thermal regime, 54 streams (watershed area = 0.2–7.9 km2; altitude < 1300 m) on the Olympic Peninsula, Washington, USA were monitored hourly for 4 years. During the study, an exceptionally warm winter (2015) was used to evaluate influences of watershed characteristics under climatic conditions similar to those projected for mid-twenty-first century. Four watershed characteristics were hypothesized to influence winter stream temperature: stream size, elevation, solar exposure, and presence of glacial materials overlying bedrock. Larger streams were associated with colder winter water temperatures and higher thermal sensitivity to atmospheric conditions. Elevation—the strongest driver of winter stream temperatures—was negatively correlated to stream temperature, except on the coldest 15 days of winter when it had no influence. Watershed solar exposure had only a marginal influence on how cold streams were in winter but was positively correlated to diel stream temperature variation and thermal sensitivity. Streams in watersheds with glacial material overlying the sedimentary bedrock were colder and had less diel variation and lower thermal sensitivity than streams in watersheds where glacial material was not present. During the warm 2015 winter, the influences of watershed characteristics on temperature tended to be weaker compared with the other years. These insights improve our understanding of how watershed physical characteristics influence stream winter thermal regimes and how these winter thermal regimes vary across landscapes, facilitating development of predictive models, a first step in designing management plans that account for winter thermal habitat needs.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Water temperature influences the rate of biological and chemical processes in streams, including the productivity, behavior, and life histories of aquatic organisms (Allan and Castillo 2007). These biological influences occur year-round across trophic levels, affecting production of periphyton, invertebrate growth and phenology, vertebrates including fish and amphibians, and community composition (Ward and Stanford 1982; Taniguchi et al. 1998; Welsh and Hodgson 2008). Though rates of biological activity peak in summer when stream temperatures are warmest, winter stream temperatures also exert important influences on stream biota at all trophic levels (Vannote and Sweeney 1980; Morin et al. 1999; Kishi et al. 2005; Durance and Ormerod 2007; Shuter et al. 2012). For fall-spawning salmonids in particular, winter water temperature regimes influence spawning, embryonic development rates, timing of emergence, juvenile rearing, and migration (Bjornn and Reiser 1979; Murray and McPhail 1988; Holtby et al. 1989; Elliott and Elliott 2010). These various influences are nuanced in that anadromous salmonids may have increased juvenile survival and growth under warmer winter stream temperatures, but accompanying earlier migration may result in reduced marine survival (Holtby 1988; Schindler and Rogers 2009; Mantua et al. 2011; Hawkins et al. 2020).
Landscape topography and the position of a stream within a drainage network affect the relative influences of the thermal processes that determine stream temperature. Stream temperature patterns generally vary with stream size, as small, headwater streams have a proportionally greater thermal influence from groundwater than do larger streams lower in a network (Macan 1958; Caissie 2006). Higher elevations generally have lower stream temperatures (Ward 1985), though the elevational gradient in stream temperature does not always exist during winter months (Isaak et al. 2018). Aspect, slope, and shading from topography and vegetation modify the amount of incoming solar radiation, which is often the dominant source of heat in streams (Brown 1969; Webb and Zhang 1997; Johnson 2004; Danehy et al. 2005). Geology within a watershed can also influence the stream thermal regime via hydrologic influences, as underlying lithology affects the movement of groundwater (Kelson and Wells 1989; Carlier et al. 2018; Wirth et al. 2020), which in turn influences stream temperature (Tague et al. 2007; Krause et al. 2012; Rosenberry et al. 2016; Briggs et al. 2018). Where glacially derived materials overlie bedrock, the groundwater stored can significantly influence phreatic groundwater inputs and streamflow dynamics (Ward et al. 1999; Caballero et al. 2002; Käser and Hunkeler 2016) and may thus influence the stream thermal regime.
Though winter stream temperatures have been much less studied than summer temperatures due in part to regulatory emphasis on summer maximum temperature (e.g., U.S. Environmental Protection Agency 1976, 2003), widespread observations of long-term warming of streams (Webb et al. 2008; Kaushal et al. 2010; Van Vliet et al. 2013) convey an urgency for a broader approach. Trends toward greater winter precipitation (Warner et al. 2015), warmer winter air temperatures, warmer rain, and reduced snow cover could have considerable impacts on winter stream temperatures and on aquatic biota (Regonda et al. 2005; Crozier et al. 2008; Leach and Moore 2014; Liu et al. 2015). Thus, to manage stream ecosystems year-round under a changing climate, it is necessary to deepen our understanding of both how and why winter stream thermal regimes vary across the landscape. In this study, we investigated small, fish-bearing streams draining 54 physically diverse watersheds in the coastal Pacific Northwest region, USA. The primary objective was to quantify the influence of watershed characteristics (stream size, elevation, solar exposure, and lithology) on the winter thermal regime of these small streams. Hypothesized watershed influences were:
H1
Stream size is inversely correlated to winter water temperature, with larger streams exhibiting colder water temperatures owing to greater atmospheric exposure.
H2
Higher elevation is associated with colder winter water temperature in streams.
H3
Solar exposure of a watershed—a function of aspect, slope, and topographic shading—is positively correlated to its winter stream temperature.
H4
In watersheds characterized by glacial materials overlying bedrock, winter stream temperatures will be moderated at both seasonal and diel timescales, and thus not as cold overall.
A second objective of this study was to assess how these factors influenced the winter thermal regime during an exceptionally warm winter (2015), representative of winter conditions projected for the mid-twenty-first century (Marlier et al. 2017; Steel et al. 2019).
Methods
Study area
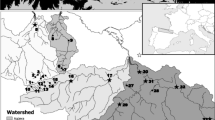
The study was conducted in 54 small, forested watersheds in the Olympic Experimental State Forest planning area (OESF), located on the western side of the Olympic Peninsula, Washington, USA (Fig. 1). This part of the peninsula is bordered by the Pacific Ocean to the west and the Strait of Juan de Fuca to the north. From a population of the 648 watersheds within the OESF that contained land managed by Washington Department of Natural Resources (WDNR), the study watersheds were selected through stratified random sampling to represent the range of physiographic conditions of WDNR-managed forestland in the OESF. Stratification was based on median watershed slope, a surrogate for the correlated attributes of distance from coast, elevation, and annual precipitation. Within strata, further criteria for selection of study watersheds were: (1) stream size at the outlet is Washington state’s Type 3 stream, equivalent to the smallest fish-bearing streams and typically 2nd and 3rd order (Strahler 1957), and (2) at least 50% of the watershed is WDNR-managed land (with the exception of four watersheds in Olympic National Park). The randomly selected study watersheds average 2.1 km2 in size (SD = 1.7 km2; min. = 0.2 km2; max = 7.9 km2). Near the outlet of each study watershed, a sample stream reach with length equal to 20 times the stream’s mean bankfull width (but always ≥ 100 m) was established. Sample reach gradient ranged from 0.8 to 21.1%. The study watersheds were hydrologically unconnected and spanned an elevational range from 27 to 1,288 m above mean sea level, from the lowest to the highest point in the watersheds.

Locations of 54 monitored watersheds and lithology on the western Olympic Peninsula, Washington, USA
The study area’s maritime climate is cool and mild. Estimated 30-year mean daily minimum air temperature for individual study watersheds in January ranges from − 1.5 to 2.8 °C, and mean daily maximum air temperature in August ranges from 20.1 to 24.6 °C (PRISM Climate Group 2018). Estimated 30-year mean annual precipitation among the study watersheds ranges from 2.15 to 4.14 m, generally increasing with elevation (PRISM Climate Group 2018). An average of 77% of annual precipitation falls during the 6-month period 1 October through 31 March, when precipitation averages ≥ 0.30 m/month in the study area. Winter precipitation is predominately rain, though approximately half of the watersheds are partially in the rain-on-snow zone (Mote et al. 2005; Nolin and Daly 2006), locally estimated to have a lower elevational boundary between 300 and 450 m (C. Snyder, WDNR, Olympia, Washington, personal communication, 2018). None of the streams in this study are glacially fed.
Lithology of the study area is predominantly marine sedimentary rocks (sandstone and shale), accreted and uplifted during subduction of the oceanic plate beneath the continental plate (Fig. 1). In some areas, sedimentary bedrock is overlain by glacial drift produced by at least four Pleistocene glaciations that extended from the Olympic Range westward through the major river valleys (Crandell 1964). The area is characterized by steep, erodible terrain, becoming mountainous at higher elevations. Soils are moderately well drained to well drained and are predominantly classified as Andisols, owing to volcanic ash influence across the region (Soil Survey Staff 2018).
The climax vegetation zones in the study area are Sitka spruce (Picea sitchensis (Bong.) Carrière) from 0 to 150 m elevation, western hemlock (Tsuga heterophylla (Raf.) Sarg.) from 150 to 550 m elevation, and Pacific silver fir (Abies amabilis (Douglas ex Loudon) Douglas ex Forbes) from 550 to 1300 m elevation (Franklin and Dyrness 1973). Within the study watersheds, the most prevalent tree species are naturally established western hemlock and planted Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco var. menziesii); Sitka spruce also is common at lower elevations, as is Pacific silver fir at higher elevations. Red alder (Alnus rubra Bong.) is common in riparian and disturbed areas.
Within the OESF, 50 of the 54 study watersheds are on majority WDNR-managed land, and four are in Olympic National Park. Whereas the latter four are in old-growth forest that was never harvested, all WDNR-managed study watersheds have a history of forest management, with initial harvests beginning in the early 1900s and peaking in the 1960s–1980s. At present, these watersheds consist of a mosaic of second-growth forest, younger third-growth forest, and unharvested old growth. Among the 50 watersheds, 35 contain less than 20% young forest (i.e., ≤ 25 years of age), 12 contain 20–40% young forest, and 3 contain > 40% young forest. Eight of these 50 watersheds contain more than 40% old-growth, 16 contain 20–40% old growth, and 26 contain < 20% old growth. Extensive clearcutting during the 1960s–1980s influenced many streams through resulting sedimentation, debris flows, and the removal of wood from streams following harvests (Martens et al. 2019). Debris flows certainly occurred in some of the study streams during that era, but stream temperature effects are unlikely to persist, as rapid reestablishment of vegetation in the region attenuates temperature effects as early as 6 years after a debris flow (Foster et al. 2020). Unharvested riparian stream buffers were first implemented in the OESF in the 1980s, and since the late 1990s, management under a habitat conservation plan effectively created an unharvested buffer at least 30 m wide on both sides of all small fish-bearing and perennial non-fish-bearing streams including those in this study (WDNR 1997). Owing to these buffers, the riparian forests of all monitored watersheds are characterized by dense canopies, typically second-growth or, in some locations, old growth. Stream shade, measured at six locations per sample reach in summer using hemispherical photography (180° field of view), was consistently high for the sample reaches, averaging 92.1% canopy closure (SD = 1.7%; range 83.4–93.9%).
The Olympic Peninsula contains a dense network of streams supporting seven species of salmonids; the most prevalent fish species in the study streams are juvenile coho salmon (Oncorhynchus kisutch), steelhead/rainbow trout (O. mykiss), coastal cutthroat trout (O. clarkii clarkii), and sculpins (Cottus spp.) (Martens et al. 2019).
Data collection
Stream water temperature (Tw) was measured in one location per sample reach at a 60-min interval for water years 2014–2017 (1 October–30 September) using a TidbiT® v2 temperature logger (UTBI-001; Onset Computer Corp., Bourne, MA, USA). The temperature loggers have a manufacturer’s stated accuracy of ± 0.21 °C; calibration of each logger was verified using an ice bath prior to deployment. Depending on stream channel morphology and substrate size, loggers were mounted instream using either a weighted tether assembly or a mount secured to a boulder by epoxy. To quantify terrestrial microclimate and to help identify potential dewatering of each stream temperature logger (resulting from channel migration or periods of low baseflow), a second TidbiT® logger was installed to monitor air temperature (Ta) at a shaded streambank location at least 1.0 m above ground but within 2.5 vertical meters and 5.0 horizontal meters from the stream logger.
All stream temperature data underwent a rigorous quality control process that began with automated detection of extreme Tw values (< 0 or > 20 °C) and large day-to-day changes in mean Tw (> 2.5 °C) (Sowder and Steel 2012). These and all other anomalous values were then examined graphically (plotted over time) and assessed in conjunction with temporally and spatially proximate stream temperature records to estimate whether data logger dewatering or malfunction had occurred. The Tw record for each stream was examined graphically in conjunction with the stream’s Ta record and field notes to identify instances of logger dewatering, burial of loggers under streambed sediments, and other irregularities. Data determined to have been recorded by dewatered, buried, or malfunctioning loggers were not used in analysis.
Metrics describing thermal regime
Many facets of the thermal regime are known to influence the health and life histories of aquatic biota (Steel et al. 2012; Vasseur et al. 2014; Maheu et al. 2016); therefore, a set of six metrics was formulated to describe components of the winter thermal regime (Table 1), including thermal sensitivity (i.e., the slope of the regression relating stream and air temperature; Kelleher et al. 2012). Based on preliminary analyses, these metrics were selected for local relevance and independence but ultimately to reflect facets of the thermal regime (Steel et al. 2017), in turn based on the natural flow regime (Poff et al. 1997). Metrics were calculated for every stream × water year combination (2014–2017) for which less than 5% of data during the winter analysis period were missing. For all metrics but DEGDAY, this winter analysis period was defined as the window of time that encompassed the 90 coldest Tw days (i.e., 24-h mean Tw) for all stream × water year combinations: 16 October through 15 May. Because degree-day accumulations for the DEGDAY metric were calculated from the beginning of the water year, the analysis period for DEGDAY began October 1. From all potential stream × water year combinations (n = 216), sufficient data were present to calculate metrics for 189. Because the reasons for data loss were varied, losses were not concentrated in any particular type of stream reach.
Predictors of thermal regime
The four watershed characteristics hypothesized to influence winter stream temperature, and associated data sources, are listed in Table 2. The spatial distribution of predictor values is shown in Fig. 2. Multicollinearity was assessed for predictors by variance inflation factor (values ranged from 1.17 to 1.57) and through scatterplots and Pearson correlation coefficients (values were between − 0.5 and 0.5). Other potential predictors, including modeled mean annual precipitation by watershed and distance from coast, were considered but not used because each was highly correlated to elevation. Similarly, watershed area was not used as a predictor because it was highly correlated to stream bankfull width which was measured in situ (r = 0.85); the two variables can be interpreted as largely equivalent in this study.

Values of four watershed characteristics used to predict winter stream temperature for 54 streams on the western Olympic Peninsula, Washington, USA: a bankfull width, b elevation of monitored reach, (c) watershed solar exposure, and (d) the fraction of the watershed on which bedrock is overlain by glacial material; see Table 2
An index of winter solar exposure was calculated for each watershed using modeled incoming solar radiation estimated by the ArcMap 10.5 Area Solar Radiation tool (ESRI 2016). Preliminary analyses showed that modeled insolation at the watershed level was a much stronger predictor of winter (and summer) stream temperature than modeled insolation at the stream itself; thus, whole-watershed insolation was used as a predictor. The Area Solar Radiation tool calculated insolation (kWh/m2) for each 10-m cell of a digital elevation model (DEM) raster surface, based on aspect, slope, latitude, elevation, sun angle, atmospheric transmissivity, proportion of radiation that is diffuse, and shadows from surrounding topography (Fu and Rich 2003). Values for diffuse radiation fraction and transmissivity were estimated to be 0.7 and 0.4, respectively, for winter in the study area using satellite data (NASA 2018). Insolation was modeled for three dates (winter solstice and 30 and 60 days post-solstice), averaged across all cells within each watershed, and then averaged across the three dates to produce an index value. Vegetative shading was not accounted for in the insolation model. Directly measured canopy closure was not used as a predictor for two reasons: (1) the dense and extremely consistent canopy closure among the 54 sample reaches prevented this from being a useful predictor, as indicated by preliminary analysis, and (2) canopy closure data were collected in summer and it is uncertain how well these measurements reflect winter values, owing to the presence of deciduous riparian trees (primarily red alder) and shrubs.
Data analysis
Prior to analysis, the data distribution for each metric was examined to identify the theoretical probability distribution by which it could be most closely approximated. The metric DUR6C followed a Poisson distribution, specified in the R glm function as family = ‘poisson’; link = ‘log’, whereas all other metrics followed a normal distribution (R Core team 2016). The majority of data analyses were performed with a set of linear models designed a priori according to our hypotheses of the influences of watershed characteristics on each stream temperature metric. Because we were not attempting to create best-fit models, predictors were retained regardless of statistical significance. Model residuals were evaluated to verify that model form was reasonable. Linear regression modeling was performed in R using the ‘glm’ and ‘lmer’ functions of the ‘lme4′ package (R Core team 2016).
For the study’s primary objective—assessing hypothesized influences of watershed characteristics on winter thermal regime—a model was created to estimate the effect size of each of the four watershed characteristics in Table 2 for each of the six temperature metrics in Table 1, across all water years (hereafter, the “watershed model”):
where Mi is a temperature metric from Table 1 for the ith stream, β0 is the intercept, β1…5 are coefficients derived from the data, WYi is water year (a random effect), BFWi is bankfull width, Ei is elevation, Si is solar exposure, Gi is proportion of the watershed underlain by glacial material, and Ɛi is error. The hypothesized influences of each watershed predictor on temperature metrics are described in Table 3.
For the study’s secondary objective—assessing interannual variation with a focus on the 2015 winter—the watershed model was run for one study year at a time (i.e., without the WY term). Coefficients were compared graphically across years.
Maps of COLDEST90 were created to demonstrate how results could be used to provide spatial predictions of the winter thermal regime at unsampled locations. The watershed model was used to predict stream temperature metrics for the population of 648 watersheds from which the sampled watersheds were selected, using coefficient estimates from the sampled watersheds. Because bankfull width data were not available for the 594 non-sampled watersheds, bankfull width was predicted from watershed area using a relationship developed from the study sample:
where BFW is the bankfull width (m) of the stream at the watershed outlet and A is watershed area (ha). The other three predictors were available for all watersheds. For brevity, maps of the other metrics are not presented. Finally, residuals from the watershed model were mapped to identify potential spatial patterns in model fit.
Results
Winter stream temperatures were relatively mild, averaging 5.7 °C on the coldest 90 days of winter and 3.7 °C on the coldest 15 days of winter across all study years (Fig. 3). Sub-zero stream temperatures were recorded only in one watershed on two dates. Winter diel Tw variation averaged 0.8 °C across all watersheds. During winter, study-wide increases and decreases in mean daily Tw as large as 5 °C occurred over time periods of 10 or fewer days (Fig. 4). Thermal sensitivity averaged 0.38 study-wide (i.e., Tw increased 0.38 °C per 1.0 °C increase in Ta), with an average R2 of 0.75.

Mean values (with one standard deviation), by water year, of metrics describing the winter thermal regime of small, fish-bearing streams; dashed lines represent the 4-year mean. COLDEST15 = Tw on 15 coldest days; COLDEST90 = Tw on 90 coldest days; DUR6C = largest number of consecutive days with mean Tw < 6 °C; DIELVAR = mean diel Tw range; DEGDAY = count of days to accumulate 1,000 Tw degree days; THERMSEN = thermal sensitivity; see Table 1

Mean daily Tw for 54 streams (black line) with one standard deviation (grey shading) during four water years
Watershed predictors
The watershed models showed strong evidence that multiple facets of the winter thermal regime varied in association with stream bankfull width (Table 4). With greater bankfull width, both COLDEST15 and COLDEST90 were colder, and it took longer to accumulate degree days (DEGDAY). Greater bankfull width also was associated with longer cold periods (DUR6C). As bankfull width increased, THERMSEN increased, reflecting a greater Tw increase per degree of Ta increase in wider streams. Diel Tw variation (DIELVAR) was not related to bankfull width; of the six metrics, this was the only one that did not express the hypothesized effect of bankfull width (Table 3). Bankfull width was not correlated to elevation (r = -0.17) among the sampled streams.
Elevation influenced five of the six metrics describing winter thermal regime, all in the hypothesized direction. Increasing elevation was associated with lower COLDEST90, slower accumulation of degree days (DEGDAY), longer cold periods (DUR6C), reduced thermal sensitivity (THERMSEN), and less diel variation in Tw (DIELVAR). On the coldest 15 days (COLDEST15), Tw did not vary with elevation.
Solar exposure influenced only one of the four metrics that broadly quantify seasonal coldness: greater solar exposure was associated with more rapid degree day accumulation (DEGDAY) but had no association with the coldest 15 or 90 days, or the duration of days below 6 °C (COLDEST15, COLDEST90, DUR6C, respectively), contrary to our hypotheses. However, solar exposure was associated with greater diel variation in Tw (DIELVAR) and with greater thermal sensitivity (THERMSEN), as hypothesized.
Glacial materials were hypothesized to moderate winter stream temperatures; no evidence was found supporting this hypothesis among the metrics describing seasonal coldness (COLDEST15, COLDEST90, DUR6C, and DEGDAY). To the contrary, the latter three of those metrics indicated that glacial materials were associated with colder winter stream temperatures. Glacial materials did, however, have a moderating effect on diel temperature range (DIELVAR) and also reduced thermal sensitivity (THERMSEN), as hypothesized.
Water year 2015
Mean air temperature for December 2014 through March 2015, measured in Forks, WA near the center of the study area, was 7.4 °C, 2.5 °C above the 100-year mean for that period (4.9 °C) and the second warmest during the past 100 years, behind 1992 (7.7 °C) (National Oceanic and Atmospheric Administration 2018). For the same December-March period, 2014, 2016, and 2017 had mean Ta in Forks of 4.8, 6.3, and 3.6 °C, respectively. Among the four water years in this study, stream temperature metrics showed that 2015 had the warmest winter temperatures (i.e., COLDEST15, COLDEST90, DUR6C, DEGDAY) and the greatest diel variation (DIELVAR) (Fig. 3). However, there was no clear indication that watershed predictors influenced temperature metrics differently in 2015 than in the other years (Fig. 5). For most of the predictor-metric combinations, the coefficients for 2015 were not the largest or smallest of the four-year study, as would be expected if watershed predictor influences truly differed in 2015. In nearly all cases where the predictor coefficients for 2015 were largest or smallest (e.g., the effect of glacial material on COLDEST90 or the effect of elevation on DUR6C), the 2015 coefficient was closer to zero than for the other years, indicating a smaller effect of the predictor in 2015. Of the 24 predictor-metric combinations in 2015, only three (13%) had coefficient confidence intervals that did not include zero. For the 72 such combinations in the other three study years, 36% had confidence intervals that did not include zero, indicating that watershed characteristics were weaker predictors of temperature metrics in 2015 than in other years.

Mean values (with 95% confidence intervals) for coefficients derived by using the watershed model to predict metrics describing the winter thermal regime of small, fish-bearing streams. Metrics are: COLDEST15 = Tw on 15 coldest days; COLDEST90 = Tw on 90 coldest days; DUR6C = largest number of consecutive days with mean Tw < 6 °C; DIELVAR = mean diel Tw range; DEGDAY = count of days to accumulate 1,000 Tw degree days; THERMSEN = thermal sensitivity; see Table 1
Predictive model
Derived from data from the 54 directly sampled watersheds, the following model was used to predict COLDEST90 for the 594 unsampled watersheds that constitute the remainder of the sampling frame:
where BFW is bankfull width (m), E is elevation (km), S is solar exposure (kWh/m2), and G is fraction of watershed underlain by glacial material. The resulting spatial distribution of predicted COLDEST90 temperatures showed a general trend toward warmer temperatures near the coast and colder temperatures inland at higher elevations but no clear patterns otherwise (Fig. 6a). The model’s residuals for the 54 directly monitored watersheds also showed no clear pattern that would indicate a spatial bias in the model (Fig. 6b).

a Mean stream temperatures on the coldest 90 days of winter (COLDEST90) as predicted for 594 unsampled watersheds and as directly measured in the 54 sampled watersheds, and b COLDEST90 watershed model residuals for the 54 directly measured watersheds on the western Olympic Peninsula, Washington, USA
Discussion
Under a relatively mild maritime climate, watershed characteristics known to influence stream temperatures in summer also influenced the winter thermal regime, though not always in the same direction or to the same degree. Although stream temperatures were warmer during the anomalous winter of 2015, the influences of watershed physical drivers on thermal regime during that winter tended to be weaker than in other years.
Watershed predictors
There was strong evidence that, after adjusting for variation in other watershed predictors including elevation, larger streams were colder in winter, as expressed through multiple facets of the thermal regime. This was likely a result of variation in the relative magnitudes of two thermal influences: groundwater entering the stream and heat exchange at the air/water interface. The thermal influence of groundwater on stream temperature is generally stronger closer to a stream’s source, where streams are smallest (Macan 1958; Poole and Berman 2001; Caissie 2006). In these small streams, winter water temperature is often moderated by the advective heat influx of groundwater, with its relatively stable year-round temperature (Erickson and Stefan 2000; O’Driscoll and DeWalle 2006; Adelfio et al. 2019). Lower in the stream network, there is greater cumulative surface exposure of streams, with increased opportunity for longwave radiative heat loss as well as convective heat loss at the air/water interface under cold winter air temperatures. This increased influence of atmospheric conditions with greater stream size was evident in the association between thermal sensitivity and bankfull width (again controlling for effects of other watershed characteristics including elevation). This finding is in agreement with previous research that found smaller streams had greater groundwater influence and lower thermal sensitivity (Kelleher et al. 2012; Johnson et al. 2020). On a larger scale, a study of rivers across England and Wales showed that water temperature in smaller drainage basins was less sensitive to air temperature than that of larger basins (Garner et al. 2014). This correlation between thermal sensitivity and stream size in streams and smaller rivers is not expected to occur in very large rivers that have substantial thermal inertia and less diel variation (Caissie 2006).
The hypothesized decrease in stream temperature with increasing elevation was clearly supported by the results, even though the elevational range among the monitoring locations was relatively small (364 m). Despite the strong observed elevation effect, we cannot rule out that other variables highly correlated to elevation, such as stream gradient or modeled mean precipitation, may also have had some influence on stream temperature. The overall influence of elevation, however, conforms to the anticipated trend of cooler streams at higher elevations where air is cooler (Segura et al. 2015; Laizé et al. 2017). This elevational influence on winter stream temperature is not detectable where streams frequently approach 0 °C (Crisp and Howson 1982; Isaak et al. 2018), but such occurrences were quite rare in this study. We also observed lower diel variation and lower thermal sensitivity of streams at higher elevations (controlling for variation in stream size and other predictors). Decreases in thermal sensitivity with elevation were also reported across a diverse group of rivers on multiple continents within an elevational range comparable to this study (48–463 m) (Morrill et al. 2005) and for streams and rivers in the USA spanning elevations from 17 to 3680 m (Johnson et al. 2020). Streams from a wide elevational range across Washington and Oregon showed a pattern in which colder, higher-elevation streams had significantly lower thermal sensitivity during summer, a pattern attributed to the contributions of snowmelt (Luce et al. 2014). The presence of snow may have influenced the stream thermal regime of the higher-elevation watersheds in the present study, but the extent of such an effect is unknown owing to a lack of data on snow accumulation. In the absence of a snow monitoring station in the area, an attempt was made to quantify snow accumulation by using daily satellite imagery, but this was precluded by frequent cloud cover. One of the only studies directly examining rain-on-snow influence on temperature of small streams was conducted in a climatically comparable study area in British Columbia, Canada (Leach and Moore 2014). That study showed that the presence of snow on the ground decreased stream temperatures by 1–2 °C during rain-on-snow events, relative to rain-on-ground events.
Among the four watershed predictors, solar exposure had the weakest association with winter low stream temperatures. We attribute this pattern to a generally low level of solar radiation reaching the watersheds in winter. In addition to the frequent cloud cover resulting from maritime climate influences, the streams are well-shaded owing to a dense riparian forest canopy in combination with steep, deeply incised topography and a solar angle that reaches a maximum of only 21° above the horizon at solar noon in mid-January. Yet, despite these conditions, watersheds with greater solar exposure were, as hypothesized, associated with greater diel temperature variation and increased thermal sensitivity. Thus the influence of solar exposure in winter was sufficient to produce these relatively small fluctuations in stream temperature, but insufficient to produce a detectable impact on overall winter stream temperature.
Watersheds with glacial material overlying bedrock were associated with colder winter stream temperatures; the direction of this influence was unexpected because previous studies reported that relatively permeable substrate (such as glacial drift) moderated seasonal extremes in stream and river temperatures through groundwater’s cooling influence in summer and warming influence in winter (Garner et al. 2014; Laizé et al. 2017). Permeable glacial outwash substrate was observed to be one of the strongest thermally moderating factors in a large-scale analysis of summer stream temperature variation in the Puget Sound region, Washington (Booth et al. 2014). Though it remains unclear why our findings differed from those of the aforementioned studies, we did observe evidence suggesting that glacial materials were associated with more stable stream temperatures, in their reduced diel stream temperature range and lower thermal sensitivity. A more complete understanding of the influence of geology on stream temperature in the study area would likely require dedicated sampling and a process-based modelling approach.
Climate influence
During the last three decades, there has been a warming trend in minimum daily stream temperature in the Pacific Northwest, particularly during winter (Isaak et al. 2012; Arismendi et al. 2013). During winter 2015, the air temperature anomaly of + 2.5 °C was comparable to projections for the mid-twenty-first century (Marlier et al. 2017). Most of our metrics of the winter thermal regime shifted in 2015 (warmer temperatures and greater diel variation), though thermal sensitivity did not. Despite these shifts in metrics, the influences of watershed characteristics on stream temperature in 2015 were not distinct compared to the other study years and even trended somewhat weaker. This may be because conditions in winter 2015 were less winter-like and so the thermal processes affected by watershed characteristics were not expressed as strongly that winter. Thus, if winter 2015 is a proxy for future winter climate conditions, future winter stream temperatures are expected to be warmer, with weakened influences of the watershed characteristics examined here. This would consequently have a homogenizing effect across the diversity of winter thermal regimes, with current differences among watersheds (e.g., Fig. 6a) declining, at least within the range in climate conditions observed in this study. A decline in thermal heterogeneity among streams due to warmer winters was documented in the Copper River Delta of Alaska, a result of reduced snowpack and ice in warm years (Adelfio et al. 2019). Our findings suggest that a climate-related decline in stream thermal heterogeneity across the landscape may extend to climates where snow and ice melt are not major drivers of stream temperature.
Implications
Winter stream temperature affects phenology of life-history transitions in aquatic organisms, competitive interactions, and food web dynamics (Cunjak and Power 1987; Bradley and Ormerod 2001; Kishi et al. 2005; Schindler and Rogers 2009; French et al. 2016), but the effects of a continually changing winter thermal regime remain challenging to predict. Salmonids, for example, will be influenced by species-specific thermal responses and tolerances at various life stages (Murray and McPhail 1988; Giannico and Hinch 2003; Crozier et al. 2008; Finstad et al. 2011; Shuter et al. 2012; Steel et al. 2012). Winter temperatures may affect the juvenile rearing stage for the study area’s steelhead, cutthroat trout, and coho salmon that are present during winter. For fall-spawning species, including chum salmon (O. keta), Chinook salmon (O. tshawytscha), and coho, winter temperatures are likely to influence timing of emergence and egg and fry development. A species’ habitat range will also determine how it is affected by a shifting winter thermal regime. In the study region, smaller, lower-elevation streams are often dominated by coho salmon and cutthroat trout, while higher-gradient headwater streams are occupied by cutthroat trout and, to a lesser degree, steelhead (Martens and Dunham 2021). A spatially explicit understanding of projected winter thermal regimes coupled with known thermal responses at the species level will facilitate targeted climate adaptation strategies (e.g., Halofsky et al. 2011) and help optimize allocation of management and monitoring activities.
This study quantifies facets of the winter thermal regime of small fish-bearing streams and relates these facets to multiple watershed characteristics. Our direct measurements, and our predictions across the broader study region (Fig. 6a), show that even streams in adjacent small watersheds can have significant thermal differences if watershed characteristics differ. Given this spatial variation in winter thermal regime—and the fact that direct monitoring of all streams at this scale is rarely feasible—our findings point to the value of assessing winter temperatures through a modelling approach based on remotely sensed watershed characteristics. Because we were able to detect influences of watershed attributes within a study area characterized by a relatively homogeneous climate and a narrow elevational range, we expect that application of this approach at a broader geographic scale, with greater elevational, geologic, and climate variability, would provide additional insight regarding the influences of local watershed attributes on stream thermal regime. Whereas the influences of watershed characteristics are expected to vary by region owing to differences in climate, physiography, and vegetation, the capacity to develop large-scale predictive models using watershed characteristics is rapidly increasing with the growing accessibility of public stream temperature data sets (e.g., NorWeST, Isaak et al. 2017) and topographic data sets (i.e., DEMs), which have global coverage (U.S. Geological Service 2019).
The relatively small number of studies on winter stream temperature continues to hamper our understanding of how winter thermal regimes are distributed across landscapes and how aquatic biota may respond to altered or warmer winter regimes. Our models of the current distribution of winter thermal regimes and of watershed-scale drivers can help managers understand where on the landscape unique winter thermal regimes may exist now. Observations from an anomalous year suggest how these distributions may appear in the future and are a first step in designing management plans that account for winter thermal habitat needs.
Availability of data and material
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Code availability
The R code used for the analysis of data in this study are available from the corresponding author upon reasonable request.
References
Adelfio LA, Wondzell SM, Mantua NJ, Reeves GH (2019) Warm winters reduce landscape-scale variability in the duration of egg incubation for coho salmon (Oncorhynchus kisutch) on the Copper River Delta, Alaska. Can J Fish Aquat Sci 76:1362–1375. https://doi.org/10.1139/cjfas-2018-0152
Allan JD, Castillo MM (2007) Stream Ecology: structure and function of running waters, 2nd edn. Springer, Dordrecht
Arismendi I, Johnson SL, Dunham JB, Haggerty R (2013) Descriptors of natural thermal regimes in streams and their responsiveness to change in the Pacific Northwest of North America. Freshw Biol 58:880–894. https://doi.org/10.1111/fwb.12094
Bjornn TC, Reiser DW (1979) Habitat requirements of salmonids in streams. In: Meehan WR (ed) Influences of forest and rangeland management of salmonid fishes and their habitat. American Fisheries Society Special Publication, pp 83–138
Booth DB, Kraseski KA, Rhett Jackson C (2014) Local-scale and watershed-scale determinants of summertime urban stream temperatures. Hydrol Process 28:2427–2438. https://doi.org/10.1002/hyp.9810
Bradley DC, Ormerod SJ (2001) Community persistence among stream invertebrates tracks the North Atlantic Oscillation. J Anim Ecol 70:987–996. https://doi.org/10.1046/j.0021-8790.2001.00551.x
Briggs MA, Johnson ZC, Snyder CD et al (2018) Inferring watershed hydraulics and cold-water habitat persistence using multi-year air and stream temperature signals. Sci Total Environ 636:1117–1127. https://doi.org/10.1016/j.scitotenv.2018.04.344
Brown GW (1969) Predicting temperatures of small streams. Water Resour Res 5:68–75. https://doi.org/10.1029/WR005i001p00068
Caballero Y, Jomelli V, Chevallier P, Ribstein P (2002) Hydrological characteristics of slope deposits in high tropical mountains (Cordillera Real, Bolivia). CATENA 47:101–116. https://doi.org/10.1016/S0341-8162(01)00179-5
Caissie D (2006) The thermal regime of rivers: a review. Freshw Biol 51:1389–1406. https://doi.org/10.1111/j.1365-2427.2006.01597.x
Carlier C, Wirth SB, Cochand F et al (2018) Geology controls streamflow dynamics. J Hydrol 566:756–769. https://doi.org/10.1016/j.jhydrol.2018.08.069
Crandell DR (1964) Pleistocene glaciations of the southwestern Olympic Peninsula, Washington. U.S. Geological Survey Professional Paper 501. Washington D.C., pp B135–B139
Crisp DT, Howson G (1982) Effect of air temperature upon mean water temperature in streams in the north Pennines and English Lake District. Freshw Biol 12:359–367. https://doi.org/10.1111/j.1365-2427.1982.tb00629.x
Crozier LG, Hendry AP, Lawson PW et al (2008) Potential responses to climate change in organisms with complex life histories: evolution and plasticity in Pacific salmon. Evol Appl 1:252–270. https://doi.org/10.1111/j.1752-4571.2008.00033.x
Cunjak RA, Power G (1987) The feeding and energetics of stream-resident trout in winter. J Fish Biol 31:493–511. https://doi.org/10.1111/j.1095-8649.1987.tb05254.x
Danehy RJ, Colson CG, Parrett KB, Duke SD (2005) Patterns and sources of thermal heterogeneity in small mountain streams within a forested setting. For Ecol Manag 208:287–302. https://doi.org/10.1016/j.foreco.2004.12.006
Durance I, Ormerod SJ (2007) Climate change effects on upland stream macroinvertebrates over a 25-year period. Glob Chang Biol 13:942–957. https://doi.org/10.1111/j.1365-2486.2007.01340.x
Elliott JM, Elliott JA (2010) Temperature requirements of Atlantic salmon Salmo salar, brown trout Salmo trutta and Arctic charr Salvelinus alpinus: predicting the effects of climate change. J Fish Biol 77:1793–1817. https://doi.org/10.1111/j.1095-8649.2010.02762.x
Erickson TR, Stefan HG (2000) Linear air/water temperature correlations for streams during open water periods. J Hydrol Eng 5:317–321. https://doi.org/10.1061/(asce)1084-0699(2000)5:3(317)
ESRI (2016) ArcGIS desktop: release 10.5. Environmental Systems Research Institute, Redlands
Finstad AG, Forseth T, Jonsson B et al (2011) Competitive exclusion along climate gradients: Energy efficiency influences the distribution of two salmonid fishes. Glob Chang Biol 17:1703–1711. https://doi.org/10.1111/j.1365-2486.2010.02335.x
Foster AD, Claeson SM, Bisson PA, Heimburg J (2020) Aquatic and riparian ecosystem recovery from debris flows in two western Washington streams. Ecol Evol 10:2749–2777
Franklin JF, Dyrness CT (1973) Natural vegetation of Oregon and Washington. Oregon State University Press, Portland
French WE, Vondracek B, Ferrington LC et al (2016) Brown trout (Salmo trutta) growth and condition along a winter thermal gradient in temperate streams. Can J Fish Aquat Sci 74:56–64. https://doi.org/10.1139/cjfas-2016-0005
Fu P, Rich PM (2003) A geometric solar radiation model with applications in agriculture and forestry. Comput Electron Agric 37:25–35. https://doi.org/10.1016/S0168-1699(02)00115-1
Garner G, Hannah DM, Sadler JP, Orr HG (2014) River temperature regimes of England and Wales: Spatial patterns, inter-annual variability and climatic sensitivity. Hydrol Process 28:5583–5598. https://doi.org/10.1002/hyp.9992
Giannico GR, Hinch SG (2003) The effect of wood and temperature on juvenile coho salmon winter movement, growth, density and survival in side-channels. River Res Appl 19:219–231. https://doi.org/10.1002/rra.723
Halofsky JE, Peterson DL, O’Halloran KA, Hoffman CH (2011) Adapting to climate change at Olympic National Forest and Olympic National Park. USDA Forest Service - General Technical Report PNW-GTR-844
Hawkins BL, Fullerton AH, Sanderson BL, Steel EA (2020) Individual-based simulations suggest mixed impacts of warmer temperatures and a nonnative predator on Chinook salmon. Ecosphere 11:e03218. https://doi.org/10.1002/ecs2.3218
Holtby LB (1988) Effects of logging on stream temperatures in Carnation Creek British Columbia, and associated impacts on the coho salmon (Oncorhynchus kisutch). Can J Fish Aquat Sci 45:502–515. https://doi.org/10.1139/f88-060
Holtby LB, McMahon TE, Scrivener JC (1989) Stream Temperatures and Inter-Annual Variability in the Emigration Timing of Coho Salmon (Oncorhynchus kisutch) Smolts and fry and Chum Salmon (O. Keta) Fry from Carnation Creek, British Columbia. Can J Fish Aquat Sci 46:1396–1405
Isaak DJ, Wollrab S, Horan D, Chandler G (2012) Climate change effects on stream and river temperatures across the northwest U.S. from 1980–2009 and implications for salmonid fishes. Clim Change 113:499–524. https://doi.org/10.1007/s10584-011-0326-z
Isaak DJ, Wenger S, Peterson E et al (2017) The NorWeST summer stream temperature model and scenarios for the western U.S.: A crowd-sourced database and new geospatial tools foster a user community and predict broad climate warming of rivers and streams. Water Resour Res 53:9181–9205
Isaak DJ, Luce CH, Chandler GL et al (2018) Principle components of thermal regimes in mountain river networks. Hydrol Earth Syst Sci Discuss 22:6225–6240. https://doi.org/10.5194/hess-2018-266
Johnson SL (2004) Factors influencing stream temperatures in small streams: substrate effects and a shading experiment. Can J Fish Aquat Sci 61:913–923. https://doi.org/10.1002/mana.19790880109
Johnson ZC, Johnson BG, Briggs MA et al (2020) Paired air-water annual temperature patterns reveal hydrogeological controls on stream thermal regimes at watershed to continental scales. J Hydrol 587:124929. https://doi.org/10.1016/j.jhydrol.2020.124929
Käser D, Hunkeler D (2016) Contribution of alluvial groundwater to the outflow of mountainous catchments. Water Resour Res 52:680–697. https://doi.org/10.1002/2014WR016730
Kaushal SS, Likens GE, Jaworski NA et al (2010) Rising stream and river temperatures in the United States. Front Ecol Environ 8:461–466. https://doi.org/10.1890/090037
Kelleher C, Wagener T, Gooseff M et al (2012) Investigating controls on the thermal sensitivity of Pennsylvania streams. Hydrol Process 26:771–785. https://doi.org/10.1002/hyp.8186
Kelson KI, Wells SG (1989) Geologic influences on fluvial hydrology and bedload transport in small mountainous watersheds, Northern New Mexico, U.S.A. Earth Surf Process Landforms 14:671–690. https://doi.org/10.1002/esp.3290140803
Kishi D, Murakami M, Nakano S, Maekawa K (2005) Water temperature determines strength of top-down control in a stream food web. Freshw Biol 50:1315–1322. https://doi.org/10.1111/j.1365-2427.2005.01404.x
Krause S, Blume T, Cassidy NJ (2012) Investigating patterns and controls of groundwater up-welling in a lowland river by combining Fibre-optic Distributed Temperature Sensing with observations of vertical hydraulic gradients. Hydrol Earth Syst Sci 16:1775–1792. https://doi.org/10.5194/hess-16-1775-2012
Laizé CLR, Meredith CB, Dunbar MJ, Hannah DM (2017) Climate and basin drivers of seasonal river water temperature dynamics. Hydrol Earth Syst Sci 21:3231–3247. https://doi.org/10.5194/hess-21-3231-2017
Leach JA, Moore RD (2014) Winter stream temperature in the rain-on-snow zone of the Pacific Northwest: Influences of hillslope runoff and transient snow cover. Hydrol Earth Syst Sci 18:819–838. https://doi.org/10.5194/hess-18-819-2014
Liu Z, Jian Z, Yoshimura K et al (2015) Recent contrasting winter temperature changes over North America linked to enhanced positive Pacific-North American pattern. Geophys Res Lett 42:7750–7757. https://doi.org/10.1002/2015GL065656
Luce C, Staab B, Kramer M et al (2014) Sensitivity of summer stream temperatures to climate variability in the Pacific Northwest. Water Resour Res 50:3428–3443. https://doi.org/10.1002/2013WR014329
Macan TT (1958) The temperature of a small stony stream. Hydrobiologia 12:89–106
Maheu A, Poff NL, St-Hilaire A (2016) A classification of stream water temperature regimes in the conterminous USA. River Res Appl 32:896–906. https://doi.org/10.1002/rra.2906
Mantua NJ, Metzger R, Crain P et al (2011) Climate change, fish, and fish habitat management at Olympic National Forest and Olympic National Park. In: Halofsky JE, Peterson DL, O’Halloran KA, Hoffman CH (eds) Adapting to climate change at Olympic National Forest and Olympic National Park. U.S.D.A. Forest Service, Portland, pp 43–60
Marlier ME, Xiao M, Engel R et al (2017) The 2015 drought in Washington State: a harbinger of things to come? Environ Res Lett 12:114008. https://doi.org/10.1088/1748-9326/aa8fde
Martens KD, Dunham J (2021) Evaluating coexistence of fish species with coastal cutthroat trout in low order streams of western Oregon and Washington, USA. Fishes 6:4. https://doi.org/10.3390/fishes6010004
Martens KD, Devine WD, Minkova TV, Foster AD (2019) Stream conditions after 18 years of passive riparian restoration in small fish-bearing watersheds. Environ Manag 63:673–690. https://doi.org/10.1007/s00267-019-01146-x
Morin A, Lamoureux W, Busnarda J (1999) Empirical models predicting primary productivity from chlorophyll a and water temperature for stream periphyton and lake and ocean phytoplankton. J N Am Benthol Soc 18:299–307. https://doi.org/10.2307/1468446
Morrill JC, Bales RC, Conklin MH (2005) Estimating stream temperature from air temperature: implications for future water quality. J Environ Eng 131:139–146. https://doi.org/10.1061/(ASCE)0733-9372(2005)131:1(139)
Mote PW, Hamlet AF, Clark MP, Lettenmaier DP (2005) Declining mountain snowpack in western north America. Bull Am Meteorol Soc 86:39–49. https://doi.org/10.1175/BAMS-86-1-39
Murray CB, McPhail JD (1988) Effect of incubation temperature on the development of five species of Pacific salmon (Oncorhynchus) embryos and alevins. Can J Zool 66:266–273. https://doi.org/10.1139/z88-038
NASA (2018) POWER: Prediction of worldwide energy resources. https://power.larc.nasa.gov/. Accessed 19 Dec 2018
National Oceanic and Atmospheric Administration (2018) National Centers for Environmental Information. https://www.ncdc.noaa.gov/. Accessed 19 Dec 2018
Nolin AW, Daly C (2006) Mapping “at risk” snow in the Pacific Northwest. J Hydrometeorol 7:1164–1171. https://doi.org/10.1175/jhm543.1
O’Driscoll MA, DeWalle DR (2006) Stream-air temperature relations to classify stream-ground water interactions in a karst setting, central Pennsylvania, USA. J Hydrol 329:140–153. https://doi.org/10.1016/j.jhydrol.2006.02.010
Poff NL, Allan JD, Bain MB et al (1997) The Natural Flow Regime. Bioscience 47:769–784. https://doi.org/10.2307/1313099
Poole GC, Berman CH (2001) An ecological perspective on in-stream temperature: Natural heat dynamics and mechanisms of human-caused thermal degradation. Environ Manage 27:787–802. https://doi.org/10.1007/s002670010188
PRISM Climate Group (2018) 30-year normals. In: Oregon State Univ. http://prism.oregonstate.edu. Accessed 12 Dec 2018
R Core team (2016) R Core Team. R A Lang. Environ. Stat. Comput. R Found. Stat. Comput. Vienna, Austria. http//www.R-project.org/. Accessed 12 Dec 2020
Regonda SK, Rajagopalan B, Clark M, Pitlick J (2005) Seasonal cycle shifts in hydroclimatology over the western United States. J Clim 18:372–384. https://doi.org/10.1175/JCLI-3272.1
Rosenberry DO, Briggs MA, Delin G, Hare DK (2016) Combined use of thermal methods and seepage meters to efficiently locate, quantify, and monitor focused groundwater discharge to a sand-bed stream. Water Resour Res 52:4486–4503. https://doi.org/10.1002/2016WR018808
Schindler DE, Rogers LA (2009) Responses of pacific salmon populations to climate variation in freshwater ecosystems. In: Krueger CC, Zimmerman CE (eds) Pacific salmon: ecology and management of western Alaska’s populations. American Fisheries Society, Bethesda, Maryland, pp 1127–1142
Segura C, Caldwell P, Sun G et al (2015) A model to predict stream water temperature across the conterminous USA. Hydrol Process 29:2178–2195. https://doi.org/10.1002/hyp.10357
Shuter BJ, Finstad AG, Helland IP et al (2012) The role of winter phenology in shaping the ecology of freshwater fish and their sensitivities to climate change. Aquat Sci 74:637–657. https://doi.org/10.1007/s00027-012-0274-3
Soil Survey Staff (2018) Official soil series descriptions. In: Nat. Resour. Conserv. Serv. United States Dep. Agric. https://www.nrcs.usda.gov/wps/portal/nrcs/main/soils/survey/. Accessed 19 Dec 2018
Sowder C, Steel EA (2012) A note on the collection and cleaning of water temperature data. Water (Switzerland) 4:597–606. https://doi.org/10.3390/w4030597
Steel EA, Tillotson A, Larsen DA et al (2012) Beyond the mean: the role of variability in predicting ecological effects of stream temperature on salmon. Ecosphere 3:104. https://doi.org/10.1890/es12-00255.1
Steel EA, Beechie TJ, Torgersen CE, Fullerton AH (2017) Envisioning, quantifying, and managing thermal regimes on river networks. Bioscience 67:506–522. https://doi.org/10.1093/biosci/bix047
Steel EA, Marsha A, Fullerton AH et al (2019) Thermal landscapes in a changing climate: biological implications of water temperature patterns in an extreme year. Can J Fish Aquat Sci 76:1740–1756. https://doi.org/10.1139/cjfas-2018-0244
Strahler AN (1957) Quantitative analysis of watershed geomorphology. Eos Trans Am Geophys Union 38:913–920. https://doi.org/10.1029/TR038i006p00913
Tague CL, Farrell M, Grant G et al (2007) Hydrogeologic controls on summer stream temperatures in the McKenzie River basin, Oregon. Hydrol Process 21:3288–3300. https://doi.org/10.1002/hyp.6538
Taniguchi Y, Rahel FJ, Novinger DC, Gerow KG (1998) Temperature mediation of competitive interactions among three fish species that replace each other along longitudinal stream gradients. Can J Fish Aquat Sci 55:1894–1901
U.S. Environmental Protection Agency (1976) Quality criteria for water. Washington D.C.
U.S. Environmental Protection Agency (2003) EPA Region 10 Guidance For Pacific Northwest State and Tribal Temperature Water Quality Standards Acknowledgments. Seattle, WA
U.S. Geological Service (2019) Earth Explorer. https://earthexplorer.usgs.gov/. Accessed 30 May 2019
Van Vliet MTH, Franssen WHP, Yearsley JR et al (2013) Global river discharge and water temperature under climate change. Glob Environ Chang 23:450–464. https://doi.org/10.1016/j.gloenvcha.2012.11.002
Vannote RL, Sweeney BW (1980) Geographic analysis of thermal equilibria: a conceptual model for evaluating the effect of natural and modified thermal regimes on aquatic insect communities. Am Nat 115:667–695
Vasseur DA, DeLong JP, Gilbert B et al (2014) Increased temperature variation poses a greater risk to species than climate warming. Proc R Soc B Biol Sci 281:20132612. https://doi.org/10.1098/rspb.2013.2612
Ward JV (1985) Thermal characteristics of running waters. Hydrobiologia 125:31–46
Ward JV, Stanford JA (1982) Thermal responses in the evolutionary ecology of aquatic insects. Annu Rev Entomol 27:97–117. https://doi.org/10.1146/annurev.en.27.010182.000525
Ward JV, Malard F, Tockner K, Uehlinger U (1999) Influence of ground water on surface water conditions in a glacial flood plain of the Swiss Alps. Hydrol Process 13:277–293. https://doi.org/10.1002/(sici)1099-1085(19990228)13:3%3c277::aid-hyp738%3e3.3.co;2-e
Warner MD, Mass CF, Salathé EP (2015) Changes in winter atmospheric rivers along the North American west coast in CMIP5 climate models. J Hydrometeorol 16:118–128. https://doi.org/10.1175/JHM-D-14-0080.1
WDNR (1997) Final habitat conservation plan. Washington Department of Natural Resources, Olympia
Webb B, Zhang Y (1997) Spatial and seasonal variability in the components of the river heat budget. Hydrol Process 11:79–101. https://doi.org/10.1002/(SICI)1099-1085(199701)11:1%3c79::AID-HYP404%3e3.3.CO;2-E
Webb BW, Hannah DM, Moore RD et al (2008) Recent advances in stream and river temperature research. Hydrol Process 22:902–918. https://doi.org/10.1002/hyp.6994
Welsh HH, Hodgson GR (2008) Amphibians as metrics of critical biological thresholds in forested headwater streams of the Pacific Northwest, U.S.A. Freshw Biol 53:1470–1488. https://doi.org/10.1111/j.1365-2427.2008.01963.x
Wirth SB, Carlier C, Cochand F et al (2020) Lithological and tectonic control on groundwater contribution to stream discharge during low-flow conditions. Water (Switzerland) 12:821. https://doi.org/10.3390/w12030821
Acknowledgements
Funding was provided by Washington State Department of Natural Resources, with additional support from the U.S. Department of Agriculture Forest Service Pacific Northwest Research Station. The authors would like to thank scientific technicians Paul Dunnette, Ellis Cropper, Mitchell Vorwerk, David Grover, and Miles Mayer. We appreciate the comments and suggestions of two anonymous reviewers.
Funding
Funding was provided by Washington State Department of Natural Resources, with additional support from the U.S. Department of Agriculture Forest Service Pacific Northwest Research Station.
Author information
Authors and Affiliations
Contributions
All five authors contributed to the conception and design of the research that resulted in this manuscript. Data collection was accomplished by a team led by TM, with assistance from AF and WD and technicians named in Acknowledgements. Analytical design was led by EAS, with contributions from all other authors. Data analyses were performed by WD. The first draft of the manuscript was written by WD. All authors provided text contributing to the manuscript and commented on and edited each revision of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflicts of interest/Competing interests
The authors have no conflicts of interest to declare that are relevant to the content of this article.
Ethics approval
None; this study did not involve human participants or biological material. No experimental work was conducted on animals.
Consent to participate
None; this study did not involve human participants or biological material.
Consent for publication
None; this study did not involve human participants or biological material.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Devine, W.D., Steel, E.A., Foster, A.D. et al. Watershed characteristics influence winter stream temperature in a forested landscape. Aquat Sci 83, 45 (2021). https://doi.org/10.1007/s00027-021-00802-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00027-021-00802-x