
Abstract
Objectives
Currently, lncRNA plays an important role in the occurrence and development of acute myeloid leukemia (AML), including SNHG5. However, the role and mechanism of SNHG5 in AML remains unclear. In this study, we explored the regulatory mechanism of SNHG5 in the development of AML.
Methods and results
QRT-PCR was used to investigate the expression of SNHG5, miR-489-3p, and SOX. The proliferation and apoptosis of AML cells were analyzed by cell transfection, cell counting kit-8 (CCK8), and flow cytometric analysis. Moreover, the expression analysis of marker proteins was detected by western blot. Through luciferase activity assay, RNA pull-down, and RNA-binding protein immunoprecipitation (RIP), we proved that SNHG5 could bind miR-489-3p and SOX4 which might be the target gene of miR-489-3p.
Results
We first found that SNHG5 was up-regulated in both AML patient bone marrow samples and various AML cell lines. Second, we found that knockdown of SNHG5 inhibited proliferation of AML cells and promoted apoptosis. It was found that SNHG5 could bind miR-489-3p, and the relative expression of SNHG5 was negatively correlated with miR-489-3p. Further results suggested that SOX4 might be the target gene of miR-489-3p. Finally, our experimental data indicated that knockdown of SNHG5 could reduce the tumor volume and down-regulated SOX4 levels in vivo.
Conclusions
Our results demonstrated that SNHG5 affected the expression of SOX4 through binding miR-489-3p to regulate proliferation and apoptosis of AML, which might act as a prospective prognostic biological marker and a promising therapeutic target for AML.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Acute myeloid leukemia (AML) is a proliferation of abnormal myeloid blasts in the bone marrow which inhibits normal hematopoiesis, and has great heterogeneity in cytogenetics, morphology, and molecular biology [1, 2]. The pathogenesis of AML is very complex, and current research indicates that chromosomal abnormalities, reproducible genetic abnormalities, and epigenetic abnormalities are the main mechanisms of AML patients [1, 3]. Without treatment, AML is often fatal, and historically has been associated with poor prognosis [4]. Advances in diagnostic and therapeutic strategies have improved the clinical outcomes of AML patients and approximately 40% of AML patients achieve long-term survival [2, 5, 6]. However, the pathogenesis of AML is still not fully understood, thus hindering new treatments for AML. Therefore, there is an urgent need to identify new predictive markers and therapeutic targets for AML.
Long non-coding RNAs (lncRNAs) are defined as heterogeneous, evolutionarily conserved non-coding transcripts that are over 200 nucleotides in length and have extremely limited protein-coding capabilities [7]. A growing number of studies have shown that frequent abnormal expression of lncRNA was observed in all human malignancies, and acted as an oncogene or tumor suppressor to regulate tumor progression [8]. In AML, lncRNAs are an important biomarker factors detected by microarray analysis. Its expression distinguished acute subtypes of leukemia, which may provide a more accurate tool for classifying leukemia and stratified patients [9, 10]. LncRNA SNHG5 has been reported to be up-regulated and served as a potential prognostic biomarker for AML. In addition, SNHG5 can regulate imatinib resistance in chronic myeloid leukemia by modulating miR-205-5p [11]. However, the role and mechanism of SNHG5 in AML remains unclear.
Through Starbase website prediction and experimental verification, our results showed that SNHG5 could target to bind to miR-489-3p. A large amount of literature has proved that miR489-3p plays an important role in the occurrence and development of cancer, and has a significant inhibiting effect on the invasion, migration, and metastasis of tumor. Jiang et al. demonstrated that miR-489-3p directly regulated the target gene Smad3 in breast cancer, inhibited the expression of Smad3, and mediated the drug resistance for chemotherapy drug doxorubicin (ADM) via the miR-489-Smad3-EMT signaling pathway [12]. Xie et al. showed that miR-489-3p targeted SUZ12 expression through EMT signaling pathway and restrained proliferation, invasion, and migration of non-small cell lung cancer (NSCLC) [13].
It is predicted by Targetscan that miR-489-3p can target SOX4. SOX4 is a key tumorigenic target for C/EBPa mutant AML [14]. In addition, overexpression of SOX4 is associated with poor prognosis of AML and promotes the progression [15, 16]. However, whether miR-489-3p can regulate the role of SOX4 in AML has not been reported.
This study was designed to investigate the role of SNHG5 in the regulation of SOX4 expression in AML by competitive binding to miR-489-3p, thereby promoting AML progression.
Materials and methods
Clinical samples
Eighty AML patients were enrolled in this study from Zhongshan Hospital and Affiliated No. 2 Hospital of Xi’an Jiaotong University, which were diagnosed by bone marrow routine, immune phenotyping, fusion gene, and chromosome detection. The clinical samples were obtained with the patient’s informed consent. Forty healthy volunteers were from bone marrow transplantation donors with a normal health check-up, and all specimens were obtained with informed consent. The marrow fluid was obtained as follows: the most common site for bone marrow puncture is the ilium (or pelvic bone). And the patient can be supine from the anterior superior spine of the ilium, or the posterior superior spine of the ilium, and if necessary also from the sternum or the protrusion of the spine. The procedure is sterilized, a sterile surgical towel is laid out, and there is a hole in the surgical towel that exposes the puncture site. After local anesthesia with about 1–2 mL 2% procaine, 0.1–0.2 mL marrow fluid was extracted by suction of the syringe with a special bone marrow puncture needle. Then, pull out the needle, cover with a sterile gauze, and finish. At the moment of extraction, there may be a brief sensation of pain, which is a sign of successful puncture. In this study, we isolated and collected CD34+ stem/progenitors from the bone marrow samples of AML patients and healthy patients. This study was done in accordance with a protocol approved by Zhongshan Hospital and Affiliated No. 2 Hospital of Xi’an Jiaotong University.
Experimental ethics policy
All experimental procedures for clinical samples studies were approved by Zhongshan Hospital and Affiliated No. 2 Hospital of Xi’an Jiaotong University, and were performed on the basis of World Medical Association Declaration of Helsinki [17]. All experimental procedures for animal research were approved by Zhongshan Hospital and Affiliated No. 2 Hospital of Xi’an Jiaotong University, and were conducted according to Guide for the Care and Use of Laboratory Animals, and were approved by the local animal research council [18].
Cell culture
The human bone marrow stromal cell line HS-5 cells and adult acute myeloid leukemia cell lines HL-60, KG-1a, Kasumi-1, ME-1, and MOLM-14 cells were purchased from Nanjing Key Gen Biotech Co., Ltd. (Nanjing, China). All cells were fostered in Dulbecco's modified Eagle's medium (Thermo Fisher Scientific, Waltham, MA, USA) with 10% fetal bovine serum (FBS) (GE Healthcare, Logan, UT, USA) and anti-biotics (Sigma-Aldrich, St. Louis, MO, USA) in a humidified chamber at 37˚C with 5% CO2.
Animals
Sixteen 6–8-week-old male nude mice (Nanjing Medical University Experimental Animal Center, Nanjing) were selected. Animals are adapted to housing conditions for 10 days, light/dark cycle for 12 h, relative humidity of 45–65%, temperature of 18–23 ℃, diet, and water could be used at will. Animals were randomly divided into control group and experimental group, with eight rats in each group. All experiments in this project were ratified through the animal ethics committees of Affiliated No. 2 Hospital School of Medicine, Xi’an Jiaotong University. The experimental procedures of animal were carried out by conforming to the handbook of the Care and Use of Laboratory Animals from the National Institute of Health, China.
RNA extraction and quantitative real-time PCR (qRT-PCR)
Total RNA was extracted by using TRIzol-Reagent (Life Technologies) in accordance with the manufacturer's protocols. High Capacity cDNA Archive kit (Applied Biosystems, Foster City, CA) was used to reverse transcribed mRNA into complementary DNA (cDNA). qPCR was performed Using an The ABI PRISM 7700 System and Taq-Man reagents (Applied Biosystems). GAPDH was used as an internal reference gene. The expression of miR-489-3p was performed by the Taq-Man stem-loop reverse transcription–PCR described in the study [19], and the levels was detected using the PrimeScript1 miRNA RT-PCR kit (Life Technologies, Carlsbad, CA) in accordance with the manufacturer’s protocols. U6 was selected as the reference gene. The primer of miR-489-3p was purchased from RiboBio Co., Ltd (Guangzhou, China). The relative expression level was calculated using the 2−ΔΔCt method. Sequences of the applied biosystems primers are as in Table 1.
Total protein extraction and western blot
The cells were collected and washed twice with pre-cooled PBS in a 1.5 mL EP tube and the precipitate was retained. Lysis buffer is used to lyse cells and collect supernatant as total protein in a new EP tube. Proteins were separated through 12.5% SDS-PAGE and transferred onto NC membranes (Millipore, Bedford, MA). The membrane was blocked at 5% BSA for 1 h at 37 °C, and then incubated at 37 °C with primary antibodies against anti-cyclin D1, CDK4/CDK6, cyclin E1, p-Rb, Rb, Bcl-2, BAX, cleaved Caspase 3, cleaved Caspase 9, cleaved-PARP, PARP, E-cadherin, N-cadherin, and GAPDH (1:500, Cell Signaling, MA). Subsequently, after washed by PBS buffers, Goat anti-rabbit secondary antibody (1:3000, Bio-Rad, USA) labeled with horseradish peroxidase (HRP) was incubated with membranes at 37 °C for 1 h. The protein signals were exposed with a storage phosphor screen. GAPDH was used as an internal reference. The density of each band was analyzed by Image J software. All the experiments were performed in triplicate.
Cloning formation experiment
In brief, HL-60 and KG-1a cells (2 × 104) transfected with si-NC, si-SNHG5#1, and si-SNHG5#2 were respectively plated into three-well plates and cultured for 2 weeks. The colonies were fixed with 10% formaldehyde for 20 min and then stained with 0.1% crystal violet for 10 min. The number of colonies including ≥ 50 cells was counted through a microscope. All experiments were conducted three times.
Cell proliferation assay
Cell proliferation assay was conducted by Cell Counting Kit-8 (CCK8) (Dojindo Lab, Japan). Briefly, 3000 cells/well were cultured in 96-well plate at 37 °C after each well was added with 10 μL CCK-8 solution. Then, the spectrophotometric absorbance was measured at 450 nm every day by Thermo Scientific Varioskan Flash machine for each sample. All the experiments were performed in triplicate and repeated three times, and the mean value was calculated.
Cell transfection
SNHG5 small interfering (si-SNHG5) and control (si-NC) were purchased from Guangzhou RiboBio Co., Ltd. (Guangzhou, China). The miRNA-489-3p inhibitors/mimics (Thermo Fisher Scientific) were transfected into the HL-60 and KG-1a cells. The siRNA sequences were as follows: si-SNHG5#1, Sense: 5′-GAGGCCAGAUUGUCUUGGA-3′, Antisense: 5′-UCCAAGACAAUCUGGCCUCTT-3′, si-SNHG5#2: Sense: GCAACGAUUUCUGGCUAGUTT, Antisense: ACUAGCCAGAAAUCGUUGCTT.
NC siRNA: 5′-CAACAAGATGAAGAGCACCAA-3′. The HL-60 and KG-1a cells were transfected with si-SNHG5 or NC siRNA using Lipofectamine® 2000 (Invitrogen; Thermo Fisher Scientific, Inc.). The negative control empty vector (NC) and the pcDNA3.1-SNHG5 plasmid (SNHG5) were also transfected into KG-1a and HL-60 cells using Lipofectamine® 2000, in accordance with the manufacturer's protocol.
Flow cytometric analysis
KG-1a and HL-60 cells transfected with si-SNHG5 or si-NC were enriched after transfection. The propidium iodide (PI) and FITC-Annexin V double dyes were used to stain the KG-1a and HL-60 cells using the FITC-Annexin V Apoptosis Detection Kit (BD Biosciences) according to the manufacturer’s directions. After double staining, the cells were analyzed by flow cytometry (FACScan®; BD Biosciences). To investigate cell cycle, the HL-60 and KG-1a cells were stained with PI using the CycleTEST™ Plus DNA Reagent Kit (BD Biosciences) following the protocol, and analyzed by FACScan. The percentage of cells in G0/G1, S, and G2/M phase were calculated.
Luciferase activity assay
The KG-1a and HL-60 cells were cultured at 60–80% confluence for 12 h, and then transfected with different plasmids including (1) pmiRGLO with + NC mimic or miR-489-3p-mimic; (2) pmiRGLO-WT (SNHG5 or SOX4) with NC mimic or miR-489-3p-mimic; (3) pmiRGLO-MUT (SNHG5 or SOX4) with NC mimic or miR-489-3p-mimic.
After cultured for 48 h, the cells were washed using PBS for two times and added 100 μL PLB (Passive Lysis Buffer) in each well. After shaking for 15 min at RT, the cell lysate was collected. 20 μL cell lysate was added into luminous plate and was read the background value using GloMax bioluminescence detection instrument for 2 s. 100 μL LAR II working solution was added into each sample. After the results were detected for 2 s, and each well was treated by 100 μL Stop&Glo® Reagent and obtain the result using GloMAx for 2 s.
RNA pull-down
According to the Pierce Magnetic RNA–Protein Pull-Down Kit instructions (Life Technologies, Baltimore, MD, USA), the specific steps are as follows: (1) In vitro transcription and biotin random labeling using pcDNA3.1-SNHG5 plasmid as a template; biotin RNA was obtained. (2) Streptavidin beads’ pretreatment: washing two equal amounts of streptavidin beads with binding buffer. One of them was used to bind biotin RNA and incubated with 200 μL of binding buffer containing 0.5% BSA and 100 μg/mL tRNA for 15 min at room temperature. (3) Cell lysate pretreatment: the cell lysate was mixed with one of the above-mentioned non-BSA and tRNA-blocked beads, and incubated at room temperature for 15 min. Leave it clear and discard beads. (4) RNA pretreatment: take 51 μg of biotin RNA and fill it to 100 μL with binding buffer. Incubate at 56 °C for 5 min and incubate at 37° C for 10 min. (5) Incubation of RNA with cell lysate: the pretreated protein lysate was incubated with biotin RNA in 200 μL binding buffer for 30 min at room temperature. (6) Pre-blocked streptavidin beads were added to the above mixture, and incubated at room temperature for 15 min. Discard the supernatant, wash the beads five times with the binding buffer, and elute the protein of interest in the beads. Western blot analyzed the expression of the target protein.
RNA-binding protein immunoprecipitation (RIP)
According to the RIP RNA-Binding Protein Immunoprecipitation Kit (Millipore, Billerica, MA, USA) instructions, the specific steps are as follows: (1) cells in the culture dish were collected, and RIP lysate was added in an equal volume to obtain a cell lysate. (2) Resuspend the magnetic beads in RIP wash buffer, add protein antibody, and incubate for 30 min at room temperature to obtain magnetic beads of connexin antibody. (3) RIP immunoprecipitation buffer containing antibody-magnetic beads was added to the cell lysate, and incubated overnight at 4 °C; discarded the supernatant and obtained the RNA–protein complex. (4) The RNA–protein complex was resuspended in proteinase K buffer and incubated at 55 °C for 30 min; retained the supernatant and added RIP wash buffer, and then extracted the RNA by chloroform–isoamyl alcohol method. The RNA was reverse transcribed to detect the level of SNHG5.
Statistical analysis
All experiments were repeated for at least three times. All results were statistically expressed as mean soil standard deviation (xS). Data were processed using SPSS 10.0 statistical software (SPSS, Chicago). The overall survival (OS) was evaluated by the Kaplan–Meier analysis and log-rank test. Student data were analyzed by Student's t test. One-way ANOVA was used to analyze the data between the e groups above. p value < 0.05 was considered statistically significant.
Results
LncRNA SNHG5 is up-regulated in AML patients and cells
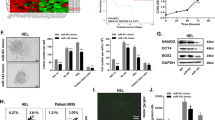
To explore whether the levels of SNHG5 change during the development of AML, we detected the expression levels of SNHG5 in 40 normal people and 80 AML patients. The results showed that the expression of SNHG5 was dramatically increased in the bone marrow tissues of AML patients compared with the normal people (Fig. 1a). To illuminate the relationship between SNHG5 accumulation levels and the survival of AML patients, the Kaplan–Meier analysis and log-rank test were used to evaluate the OS (overall survival) (Fig. 1b). The results of survival analysis indicated that overexpression of SNHG5 significantly reduced survival (p < 0.0486), which suggested that up-regulation of SNHG5 might contribute to the development of AML. Furthermore, we detected the expression levels of SNHG5 in HS-5, HL-60, KG-1a, Kasumi-1, ME-1, and MOLM-14 cells. Results showed that compared with HS-5 cells (bone marrow stromal cells of normal people), the expression of SNHG5 were also markedly increased in AML cell lines (Fig. 1c).

SNHG5 was up-regulation in AML tissues and cell lines. a qRT-PCR detected the expression level of SNHG5 in AML tissues and normal tissues. b Kaplan–Meier survival analysis of overall prognosis in AML cases with high or low SNHG5 expression. c Relative expression of SNHG5 in AML cell lines HL-60, KG-1a, Kasumi-1, ME-1, MOLM-14, and normal cells HS-5. *p < 0.05; **p < 0.01; ***p < 0.001
Knockdown of SNHG5 inhibits proliferation of AML cells
To investigate the biological function of SNHG5 in AML cells, negative controls (si-NC) and si-SNHG5 were, respectively, transfected into KG-1a and HL-60 cells. qRT-PCR results showed that the expression of SNHG5 was significantly down-regulated in HL-60 and KG-1a cells transfected with si-SNHG5 (Fig. 2a). Furthermore, the effect of SNHG5 on cell proliferation was confirmed by CCK-8 assay. The results revealed that down-regulation of SNHG5 with si-SNHG5 significantly suppressed the cell growth of KG-1a and HL-60 cells (Fig. 2b), suggesting that knockdown of SNHG5 inhibited AML cells proliferation.

Knockdown of SNHG5 inhibited the proliferation of AML cells. a qRT-PCR detected the expression level of SNHG5 in HL-60 and KG-1a cells transfected with si-NC or si-SNHG5. b Effect of SNHG5 knockdown on cell proliferation was assessed by CCK-8 assay in HL-60 and KG-1a cells. c Analysis of statistical results detected by flow cytometry for the effects of SNHG5 knockdown on cells cycle in KG-1a and HL-60 cells. d Western blot detected the expression of cell cycle-related protein in HL-60 and KG-1a cells transfected with si-NC or si-SNHG5. *p < 0.05; **p < 0.01; ***p < 0.001
Subsequently, we carried out cell cycle analysis and the result demonstrated that HL-60 and KG-1a cells transfected with si-SNHG5 were arrested in G1 phase (Figure S1). Next, we detected the expression of cell cycle-related genes, including Cyclin D1, Cyclin-dependent kinases-4 (CDK4), Cyclin-dependent kinases-6 (CDK6), Cyclin E1, p-Rb, and Rb. Results showed that the expression levels of Cyclin D1, CDK4, CDK6, Cyclin E1, and p-Rb were significantly decreased in HL-60 and KG-1a cells transfected with si-SNHG5 (Fig. 2d). These results indicated that SNHG5 could promote the cell proliferation and the knockdown of SNHG5 restrained the AML cell proliferation.
Knockdown of SNHG5 promotes apoptosis of AML cells
To investigate the function of SNHG5 on the apoptosis of AML cells, we performed cell apoptosis assay through the flow cytometry analysis. The experimental results showed that the apoptotic rate of HL-60 and KG-1a cells transfected with si-SNHG5 was obviously increased, compared with the control (Fig. 3a). In addition, we detected the accumulation level of apoptosis-related proteins, including B-cell lymphoma/leukernia-2(Bcl-2), bcl-2-associated x-protein (BAX), C-caspase 3, C-caspase 9, Cleaved-Poly (ADP-ribose) polymerases (C-PARP), and PARP. Western blot results showed that compared with the control group, the accumulation levels of Bcl-2 and PARP were decreased, while the accumulation levels of BAX, C-caspase 3, C-caspase 9, and C-PARP were increased in HL-60 and KG-1a cells transfected with si-SNHG5 (Fig. 3b). Taken together, these results suggested that knockdown of SNHG5 promoted the apoptosis of AML cells.

Knockdown of SNHG5 promoted AML cells apoptosis. a Flow cytometry detects the effects of SNHG5 knockdown on cells apoptosis in KG-1a and HL-60 cells. b Western blot detected the expression of cell apoptosis-related protein in HL-60 and KG-1a cells transfected with si-NC or si-SNHG5. *p < 0.05; **p < 0.01; ***p < 0.001
The expression of SNHG5 was negatively correlated with miR-489-3p
According to the prediction of starbase website, the sequence of SNHG5 was highly matched with miR-489-3p (Fig. 4a). Luciferase reporter assays were performed using the wild-type SNHG5 and mutant SNHG5 with or without miR-489-3p. The mutant SNHG5 sequence was unable to bind the miR-489-3p. Transfection of miR-489-3p obviously decreased the levels of the wild-type SNHG5 (Fig. 4b), which indicated that miR-489-3p could reduce the luciferase activity by binding to SNHG5, and miR-489-3p might be the target of SNHG5. Furthermore, RIP assay showed that SNHG5 enriched the miRNA ribonucleoprotein complex containing Ago2 compared to the control IgG (Fig. 4c).

Expression of SNHG5 was negatively correlated with miR-489-3p. a The matched sequence of wild-type or mutant-type SNHG5 with miR-489-3p. b The luciferase activity assay of HL-60 and KG-1a cells after transfected with empty vector (pmiRGLO), wild-type SNHG5, or mutant SNHG5 with miR-489-3p mimic or NC mimic, respectively. c RIP assay showed that SNHG5 enriched the miRNA ribonucleoprotein complex containing Ago2 compared to the control IgG from KG-1a and HL-60 cells lysates. d QRT-PCR examined the expression of SNHG5 in KG-1a and HL-60 cells transfected with NC or SNHG5 (left), and the expression of miR-489-3p in KG-1a and HL-60 cells transfected with si-NC, si-SNHG5, NC, or SNHG5 (right). e qRT-PCR detected the expression level of miR-489-3p in AML tissues and normal tissues (left). Correlation analysis indicated that relative SNHG5 level was negative correlated with miR-489-3p expression. ***p < 0.001
To further explore the relationship between SNHG5 and miR-489-3p expression. We transferred pcDNA3.1-SNHG5 into HL-60 and KG-1a cells for the overexpression of SNHG5, and the qRT-PCR results suggested that the expression level of SNHG5 was significantly increased (Fig. 4d). Then, we detected the expression levels of miR-489-3p in HL-60 and KG-1a cells transfected with si-NC, si-SNHG5, NC, or pcDNA3.1-SNHG5, respectively. Results showed that knockdown of SNHG5 promoted miR-489-3p expression, while overexpression of SNHG5 reduced the accumulation of miR-489-3p (Fig. 4d). Finally, we evaluated the level of miR-489-3p between AML patients and healthy controls. The results suggested that miR-489-3p was down-regulated in AML patients (Fig. 4e), and correlation analysis suggested that the relative expression of SNHG5 was negatively correlated with the expression of miR-489-3p (Fig. 4e).
MiR-489-3p targets the SOX4 gene
To determine the target gene of miR-489-3p, we used TargetScan to predict the potential target genes. It revealed that the 3′-UTR of SOX4 contained a potential binding site of miR-489-3p (Fig. 5a). Luciferase reporter assays of the wild-type and mutant 3′-UTRs of SOX4 were performed in the presence or absence of miR-489-3p overexpression. The mutant 3′-UTR sequence prevented miR-489-3p binding. Transfection of miR-489-3p apparently decreased the luciferase activity by binding to the 3′-UTR of SOX4 (Fig. 5b), which suggested that SOX4 was a target for miR-489-3p. The results of western blot and qRT-PCR suggested that the mRNA expression levels and protein accumulation of SOX4 were decreased when the miR-489-3p was up-regulated, whereas increased when down-regulation of miR-489-3p (Fig. 5c, d). Next, we detected the mRNA and protein accumulation of SOX4 in AML patients and healthy controls, and the results showed that the mRNA and protein expression of SOX4 were increased in AML patients (Fig. 5e, f), and correlation analysis suggested that the relative expression of SOX4 was negatively correlated with miR-489-3p (Fig. 5g). Taken together, these results verified that the SOX4 could be targeted by the miR-489-3p.

SOX4 is the target gene of miR-489-3p. a The wt or mutant type of SOX4 3′ UTR for miR-489-3p. b The luciferase activity of wild-type or mutant SOX4 3′ UTR in HL-60 and KG-1a cells transfected with miR-489-3p mimic or NC mimic. c qRT-PCR detected the expression of miR-489-3p (left) and SOX4 (right) in HL-60 and KG-1a cells transfected with miR-NC, miR-489-3p, NC inh, or miR-489-3p inh. d Western blot evaluated the expression of SOX4 protein in HL-60 and KG-1a cells transfected with miR-NC, miR-489-3p, NC inh or miR-489-3p inh. e, f, qRT-PCR (E) and western blot (F) detected the expression of SOX4 in AML tissues and normal tissues. g Correlation analysis showed that relative SOX4 expression was negative correlated with miR-489-3p expression. *p < 0.05; **p < 0.01; ***p < 0.001
SNHG5 regulates SOX4 by miR-489-3p to regulate proliferation and apoptosis
To further analyze the underlying mechanism of SNHG5, we knocked down of SNHG5 and inhibited miR-489-3p in HL-60 cells. CCK-8 assay showed that HL-60 cells proliferation was inhibited when knockdown of SNHG5, but rescued to normal level when inhibited miR-489-3p expression (Fig. 6a). Next, we conducted flow cytometry to analyze the cell cycle and apoptosis of HL-60 cells. Downregulation of SNHG5 arrested the cell in G1 phase and promoted apoptosis. However, the cell cycle and apoptosis recovered to the normal levels when inhibited the miR-489-3p expression (Figure S2 and Fig. 6c). Finally, we detected the expression of cell cycle-related, and apoptosis-related proteins, and the results were consistent with the above results (Fig. 6d). Taken together, these data suggested that SNHG5 adjusted the expression of SOX4 through binding the miR-489-3p to regulate cell proliferation and apoptosis.

SNHG5 regulates SOX4 by miR-489-3p to regulate proliferation and apoptosis. a CCK-8 assay assessed the cell proliferation in HL-60 and KG-1a cells transfected with inh NC and si-NC, inh NC and si-SNHG5, or si-SNHG5 and miR-489-3p inh. b, c, Analysis of statistical results of cells cycle (b) and apoptosis (c) in KG-1a and HL-60 cells transfected with inh NC and si-NC, inh NC and si-SNHG5, or si-SNHG5 and miR-489-3p inh. d Western blot detected the accumulation of proteins related to cell cycle, apoptosis, in HL-60 and KG-1a cells transfected with inh NC and si-NC, inh NC and si-SNHG5, or si-SNHG5 and miR-489-3p inh. *p < 0.05; **p < 0.01; ***p < 0.001
Knockdown of SNHG5 reduces tumor volume and down-regulates SOX4 levels in vivo
To further explore the effect of SNHG5 on the tumorigenicity in mice, we injected 107 HL-60 cells transfected with siSNHG5 or siNC into nude mice. First, we detected the expression level of SNHG5 in HL-60 cells transfected with siSNHG5 or siNC by qRT-PCR, and the results suggested that expression level of SNHG5 was significantly decreased in si-SNHG5 transfected HL-60 cell compared with si-NC (Fig. 7a). We measured the change in tumor volume and weight within 5 weeks, and we found that the knockdown of SNHG5 in HL-60 inhibited tumorigenesis of HL-60 cells in nude mice (Fig. 7b), including reducing the weight and volume of AML tumors (Fig. 7c). Moreover, our results indicated that knockdown of SNHG5 also down-regulated the expression of SOX4 in vivo (Fig. 7d). In summary, the above results showed that knockdown of SNHG5 reduced tumor growth and down-regulated SOX4 levels in vivo.

Knockdown of SNHG5 reduces tumor volume and down-regulates SOX4 levels in vivo. a qRT-PCR detected the expression of SNHG5 in HL-60 transfected with siNC or siSNHG5. b, c The picture of tumor size (b), tumor weight, and tumor volume (c) in mice injected HL-60 cell with shNC or shSNHG5. d Western blot detected the expression of SOX4 in mice injected HL-60 cell with shNC or shSNHG5. *p < 0.05; ***p < 0.001
Discussion
In this study, we found that SNHG5 was up-regulated in the bone marrow tissues of AML patients and various AML cell lines, and its expression was negatively correlated with the overall survival of AML patients. Furthermore, knockdown of SNHG5 inhibited the proliferation of AML cells, and promoted the apoptosis of AML cells. In addition, our results first illuminated that SHNG5 could affect the expression of SOX4 through binding the miR-489-3p to regulate the proliferation and apoptosis of AML cells.
Studies have shown that the aberrant of miRNAs expression level played an important role in the occurrence, development, diagnosis, and prognosis of AML [20]. For example, high expressions of miR-155, miR-125b, and miR-100 play a role in the progression of disease in AML patients, whereas the expressions of miR-29a, miR-142-3p, and miR-223 are reduced in AML patients, act as the tumor suppressor gene [21, 22]. MiR-489-3p plays a vital role in inhibiting the development and progression of various cancers. For instance, studies have shown that miR-489-3p participates in the regulation of osteosarcoma (OS) metastasis, and targets the paired box gene 3 (PAX3) gene to reduce the expression of PAX3 to inhibit OS (osteosarcoma) metastasis [23]. In our study, our results first showed that miR-489-3p, which was positively related to the development of AML, regulated the proliferation and apoptosis, of AML cells through targeting SOX4 and reducing its expression. However, recent studies have confirmed that miR-489-3p also played a role in promoting cancer by promoting tumor cell proliferation, inhibiting apoptosis. MiR-489-3p is overexpressed in renal clear cell carcinoma and renal clear cell papillary carcinoma [24, 25]. These studies indicate that the expression profile of miR-489-3p is clearly cell-specific and tissue-specific.
Current research indicates that SNHG5 has an important function in different cancers. SNHG5 can regulate the expression of miR-155 to promote the proliferation of melanoma and induce apoptosis, and SNHG5 also promotes the cell proliferation of bladder cancer through targeting p27 [26,27,28]. In addition, SNHG5 is significantly down-regulated in gastric cancer (GC) and inhibited the proliferation of GC cells, which plays a role in the development of GC by interacting with metastasis-associated gene 2 (MTA2) [29]. The SNHG5/miR-32 axis can target the KLF4 to regulate the proliferation and migration of gastric cancer cell. In colorectal cancer (CRC), it has reported that SNHG5 is up-regulated and knockdown of lncRNA SNHG5 could inhibit the proliferation of colorectal cancer cells and promote the apoptosis [30]. In this study, we first found that the expression of SNHG5 was negatively correlated with the AML and it could increase the expression of SOX4 by competitively binding miR-489-3p. At the same time, our study proved for the first time that miR-489-3p could target SOX4 and reduced its expression.
Recent studies have shown that the SOX4 gene, as a member of the human sex-determining HMG domain family, is involved in tumorigenesis, including AML [31, 32]. Studies have shown that retroviruses carrying the SOX4 gene could synergize with myocyte enhancer factor 2 (Mef2c) and cause myeloid leukemia. The latest study showed that during the development of AML, miR-339-5p exerts its anti-proliferative effect in AML by directly targeting SOX4 [33]. The study found that AML patients with low SOX4 expression had higher remission rates and longer overall survival than AML patients with high SOX4 expression. In addition, other studies have shown that CASC15 regulated cell survival, proliferation, and expression of its chromosomal neighboring gene SOX4, and played an important role in RUNX1/AML translocation leukemia [16]. Here, our results indicated that miR-489-3p could target SOX4 and reduced its expression, which could directly regulate the proliferation and apoptosis of AML. In summary, our research revealed the clinical significance of SOX4 in AML and its role in the development of leukemia.
Cyclin and CDK are important proteins in the cell cycle regulatory system [34]. Overexpression of Cyclin D1 activated the activity of CDK4 and CDK6, shorten the G1 phase, dysregulated cell cycle regulation and abnormal cell proliferation, leading to tumorigenesis [35, 36]. In our study, the expressions of CylinD1, CDK4, and CDK6 were significantly reduced in SNHG5 knockdown cell lines, which restored normal cell cycle and inhibited the proliferation of the cells. Studies have demonstrated that the level of Bcl-2 and Bax protein formed a positive and negative regulation of apoptosis. When Bcl-2 was highly expressed and the Bcl-2/Bax ratio was up-regulated, the two proteins formed heterodimers and inhibited apoptosis [37]. When Bax was highly expressed and the Bcl-2/Bax ratio was down-regulated, the two proteins formed homodimers and promoted apoptosis [37, 38]. In this study, the expression of Bcl-2 was significantly reduced and the levels of Bax were increased in SNHG5 knockdown cell lines, which indicated that the Bcl-2/Bax might form the homodimer and promote apoptosis. We also detected the effect of SNHG5 in vivo. HL-60 cells transfected with shSNHG5 or shNC were injected into nude mice, and it was found that knockdown of SNHG5 reduced the tumor volume and down-regulated the SOX4 level. SCID-AML model is a common model used to study the proliferation, differentiation, and regulatory mechanism of leukemia [41]. The mouse acute leukemia model of immunodeficiency is one of the important methods to study the biological characteristics, treatment, and prognosis of AML [42]. In our further studies, the SCID-AML models will be employed to verify the regulatory effect of SNHG5 on the proliferation and metastasis of AML.
Conclusion
In summary, these results proved that SNHG5 could regulate the expression of SOX4 in AML by competitively binding mir-489-3p, which further affected the proliferation, and apoptosis of AML. Our research implied that SNHG5 might act as a prospective prognostic biological marker and a promising therapeutic target for AML.
Data availability
All data generated or analyzed during this study are included in this published article.
References
Houda H. Angiogenesis and acute myeloid leukemia. Hematology. 2014;19(6):311–23.
Grimwade D. The clinical significance of cytogenetic abnormalities in acute myeloid leukaemia. Best Pract Res Clin Haematol. 2001;14(3):497–529.
Omar AW, Levine RL. Mutations in epigenetic modifiers in the pathogenesis and therapy of acute myeloid leukemia. Blood. 2013;121(18):3563–72.
Alessandro I, Valentina S, Marilena C, Federica L, Giuseppe V, Sarah P, et al. The role of the immunosuppressive microenvironment in acute myeloid leukemia development and treatment. Expert Rev Hematol. 2014;7(6):807–18.
Ha PM, Cho SA, Kyung Hyun Y, Moon Hee Y, Young AJ, Hyo Soo L, et al. Gene expression profile related to prognosis of acute myeloid leukemia. Oncol Rep. 2007;18(6):1395–402.
Basturk A, Akinci S, Hacibekiroglu T, Guney T, Kutlucan A, Ceran F, et al. Prognostic significance of flow cytometry findings in Turkish adult acute leukemia patients. Eur Rev Med Pharmacol Sci. 2015;19(18):3360–6.
Yang G, Lu X, Yuan L. LncRNA: a link between RNA and cancer. Biochem Biophys Acta. 2014;1839(11):1097–109.
Gibb EA, Brown CJ, Wan LL. The functional role of long non-coding RNA in human carcinomas. Mol Cancer. 2011;10(1):38.
Fernando TR, Rodriguez-Malave NI, Waters EV, Yan W, Casero D, Basso G, et al. LncRNA expression discriminates karyotype and predicts survival in B-lymphoblastic leukemia. Mol Cancer Res. 2015;13(5):839–51.
Chen L, Wang W, Cao L, Li Z, Wang X. Long non-coding RNA CCAT1 acts as a competing endogenous RNA to regulate cell growth and differentiation in acute myeloid leukemia. Mol Cells. 2016;39(4):330–6.
He B, Bai Y, Kang W, Zhang X, Jiang X. LncRNA SNHG5 regulates imatinib resistance in chronic myeloid leukemia via acting as a CeRNA against MiR-205-5p. Am J Cancer Res. 2017;7(8):1704.
Jiang L, He D, Yang D, Chen Z, Pan Q, Mao A, et al. MiR-489 regulates chemoresistance in breast cancer via epithelial mesenchymal transition pathway. FEBS Lett. 2014;588(11):2009–155.
Xie Z, Cai L, Li R, Zheng J, Wu H, Yang X, et al. Down-regulation of miR-489 contributes into NSCLC cell invasion through targeting SUZ12. Tumour Biol. 2015;36(8):6497–505.
Zhang H, Alberich-Jorda M, Amabile G, Yang H, Staber PB, Di Ruscio A, et al. Sox4 is a key oncogenic target in C/EBPalpha mutant acute myeloid leukemia. Cancer Cell. 2013;24(5):575–88. https://doi.org/10.1016/j.ccr.2013.09.018.
Lu JW, Hsieh MS, Hou HA, Chen CY, Tien HF, Lin LI. Overexpression of SOX4 correlates with poor prognosis of acute myeloid leukemia and is leukemogenic in zebrafish. Blood Cancer J. 2017;7(8):e593. https://doi.org/10.1038/bcj.2017.74.
Fernando TR, Contreras JR, Zampini M, Rodriguez-Malave NI, Alberti MO, Anguiano J, et al. The lncRNA CASC15 regulates SOX4 expression in RUNX1-rearranged acute leukemia. Mol Cancer. 2017;16(1):126. https://doi.org/10.1186/s12943-017-0692-x.
Press OU. World Medical Association declaration of Helsinki. Gastroenterologia Japonica. 1991;26(2):269–70.
Clark JD, Gebhart GF, Gonder JC, Keeling ME, Kohn DF. The 1996 guide for thecare and use of laboratory animals. Ilar J. 1997;38(1):41–8.
Shi DB, Wang YW, Xing AY, Gao JW, Zhang H, Guo XY, et al. C/EBPα-induced miR-100 expression suppresses tumor metastasis and growth by targeting ZBTB7A in gastric cancer. Cancer Lett. 2015;369(2):376–85.
Wang X, Chen H, Bai J, He A. MicroRNA: an important regulator in acute myeloid leukemia. Cell Biol Int. 2017;41(9):936–45. https://doi.org/10.1002/cbin.10770.
Wallace JA, O'Connell RM. MicroRNAs and acute myeloid leukemia: therapeutic implications and emerging concepts. Blood. 2017;130(11):1290–301. https://doi.org/10.1182/blood-2016-10-697698.
Liao Q, Wang B, Li X, Jiang G. miRNAs in acute myeloid leukemia. Oncotarget. 2017;8(2):3666–822. https://doi.org/10.18632/oncotarget.12343.
Liu Q, Yang G, Qian Y. Loss of MicroRNA-489-3p promotes osteosarcoma metastasis by activating PAX3-MET pathway. Mol. Carcinog. 2016;56(4):1312–21.
Apostolos Z, Lambrou GI, Nikos M, Patroklos K, Gregory P, Krinio G, et al. New miRNA profiles accurately distinguish renal cell carcinomas and upper tract urothelial carcinomas from the normal kidney. PLoS ONE. 2014;9(3):e91646.
Enrico M, Luigi M, Apurva C, Masamichi H, Guido M, Matteo B, et al. Clear cell papillary renal cell carcinoma: micro-RNA expression profiling and comparison with clear cell renal cell carcinoma and papillary renal cell carcinoma. J Urol. 2014;191(4):e244-e.
Ma Z, Xue S, Zeng B, Qiu D. lncRNA SNHG5 is associated with poor prognosis of bladder cancer and promotes bladder cancer cell proliferation through targeting p27. Oncol Lett. 2018;15(2):1924–30.
Wang Z, Pan L, Yu H, Wang Y. The long non-coding RNA SNHG5 regulates gefitinib resistance in lung adenocarcinoma cells by targeting miR-377/CASP1 axis. Biosci Rep. 2018;38(4):BSR20180400.
Lu Y, Wang S, Yue L, Tognetti L, Rui T, Kang Z, et al. SNHG5 promotes proliferation and induces apoptosis in melanoma by sponging miR-155. RSC Adv. 2018;8(11):6160–8.
Zhao L, Guo H, Zhou B, Feng J, Li Y, Han T, et al. Long non-coding RNA SNHG5 suppresses gastric cancer progression by trapping MTA2 in the cytosol. Oncogene. 2016;35(44):5770.
Damas ND, Marcatti M, Côme C, Christensen LL, Nielsen MM, Baumgartner R, et al. SNHG5 promotes colorectal cancer cell survival by counteracting STAU1-mediated mRNA destabilization. Nat Commun. 2016;7:13875.
Fung TK, Leung AY, So CW. Sox4you: a new player in C/EBPalpha leukemia. Cancer Cell. 2013;24(5):557–9. https://doi.org/10.1016/j.ccr.2013.10.016.
Chen J, Ju HL, Yuan XY, Wang TJ, Lai BQ. SOX4 is a potential prognostic factor in human cancers: a systematic review and meta-analysis. Clin Transl Oncol. 2016;18(1):65–72. https://doi.org/10.1007/s12094-015-1337-4.
Sun X, Liu H, Li T, Qin L. MicroRNA3395p inhibits cell proliferation of acute myeloid leukaemia by directly targeting SOX4. Mol Med Rep. 2018;18(6):5261–9. https://doi.org/10.3892/mmr.2018.9552.
Pines J. The cell cycle kinases. Semin Cancer Biol. 1994;5(4):305–13.
John RR, Malathi N, Ravindran C, Anandan S. Mini review: Multifaceted role played by cyclin D1 in tumor behavior. Indian J Dent Res. 2017;28(2):187–92. https://doi.org/10.4103/ijdr.IJDR_697_16.
Sherr CJ, Beach D, Shapiro GI. Targeting CDK4 and CDK6: from discovery to therapy. Cancer Discov. 2016;6(4):353–67. https://doi.org/10.1158/2159-8290.cd-15-0894.
Renault TT, Dejean LM, Manon S. A brewing understanding of the regulation of Bax function by Bcl-xL and Bcl-2. Mech Ageing Dev. 2017;161(Pt B):201–10. https://doi.org/10.1016/j.mad.2016.04.007.
Edlich F. BCL-2 proteins and apoptosis: recent insights and unknowns. Biochem Biophys Res Commun. 2018;500(1):26–34. https://doi.org/10.1016/j.bbrc.2017.06.190.
Bourboulia D, Stetler-Stevenson WG. Matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs): positive and negative regulators in tumor cell adhesion. Semin Cancer Biol. 2010;20(3):161–8.
Malemud CJ. Matrix metalloproteinases (MMPs) in health and disease: an overview. Front Biosci. 2006;11(2):1696–701.
Xu KH, Lu DP. Plumbagin induces ROS-mediated apoptosis in human promyelocytic leukemia cells in vivo. Leuk Res. 2010;34(5):658–65. https://doi.org/10.1016/j.leukres.2009.08.017.
Her Z, Yong KSM, Paramasivam K, Tan WWS, Chan XY, Tan SY, et al. An improved pre-clinical patient-derived liquid xenograft mouse model for acute myeloid leukemia. J Hematol Oncol. 2017;10(1):162. https://doi.org/10.1186/s13045-017-0532-x.
Author information
Authors and Affiliations
Contributions
XYY and WGZ conceived and designed the experiments, MYF and CCW analyzed and interpreted the results of the experiments, and LH and CMY performed the experiments.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interests, and all authors should confirm its accuracy.
Ethics approval and consent to participate
The animal use protocol listed below has been reviewed and approved by the Animal Ethical and Welfare Committee.
Additional information
Responsible Editor: John Di Battista.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.

11_2020_1345_MOESM1_ESM.jpg
Figure S1 The cell cycle examined by flow cytometry about the effects of SNHG5 knockdown on cells cycle in KG-1a and HL-60 cells. Figure S2 Cells cycle detected by flow cytometry in KG-1a and HL-60 cells transfected with inh NC and si-NC, inh NC and si-SNHG5, or si-SNHG5 and miR-489-3p inh (JPG 3063 kb)
Rights and permissions
About this article

{kind=link}
Cite this article
Ying, X., Zhang, W., Fang, M. et al. LncRNA SNHG5 regulates SOX4 expression through competitive binding to miR-489-3p in acute myeloid leukemia. Inflamm. Res. 69, 607–618 (2020). https://doi.org/10.1007/s00011-020-01345-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00011-020-01345-x